

Genome-Wide Identification, Characterization, and Expression of the HAK/KUP/KT Potassium Transporter Gene Family in Poncirus trifoliata and Functional Analysis of PtKUP10 under Salt Stress

 ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials and Treatments
2.2. Bioinformatics Analysis
2.3. RNA Isolation and Fluorescence Quantitative Real-Time PCR Analysis
2.4. Transformation of Arabidopsis and Phenotypic Analysis
2.5. Data Analysis
3. Results
3.1. Genome-Wide Identification of PtKUP Gene Family
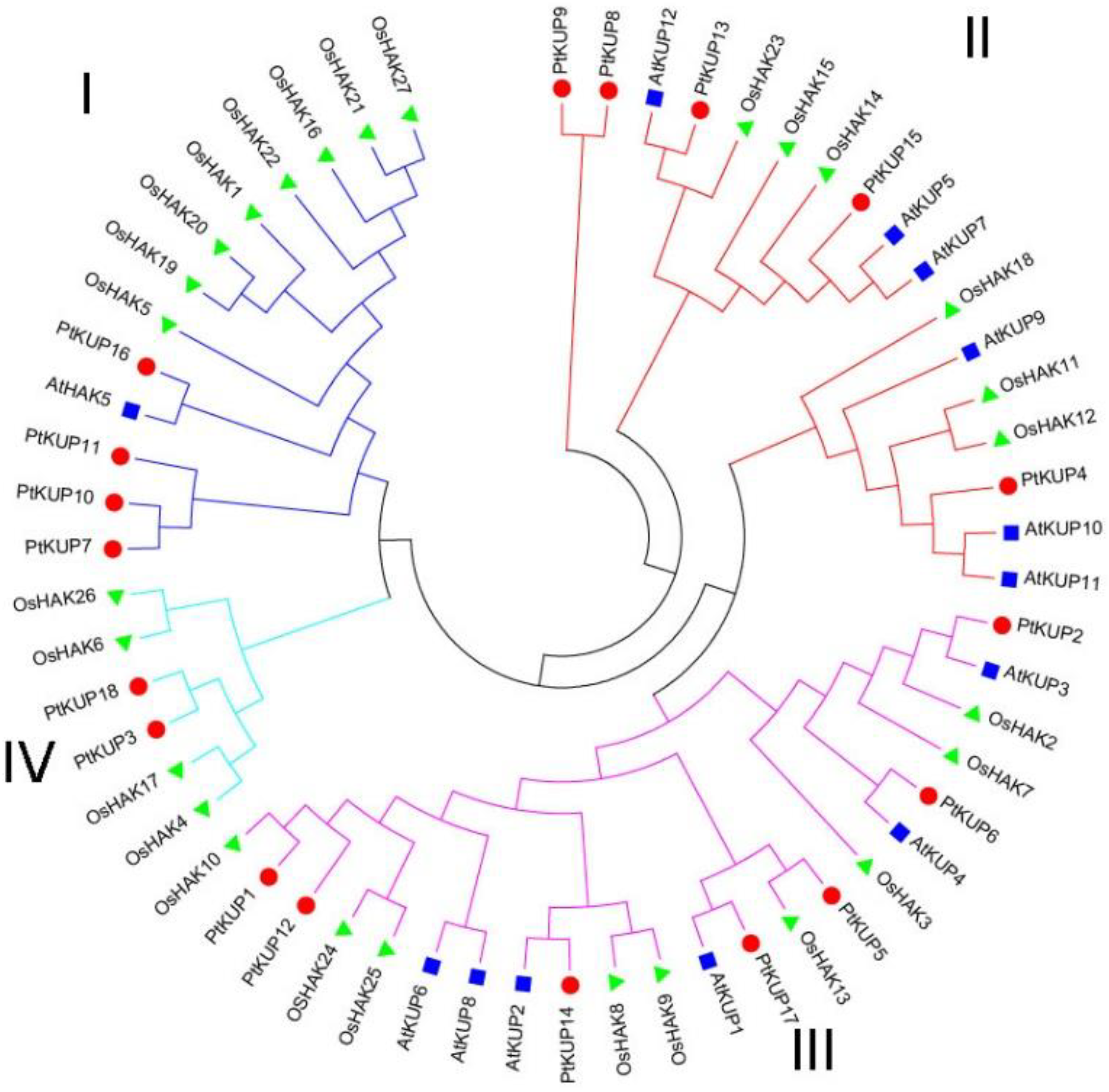
3.2. Phylogenetic Analyses of PtKUP Gene Family
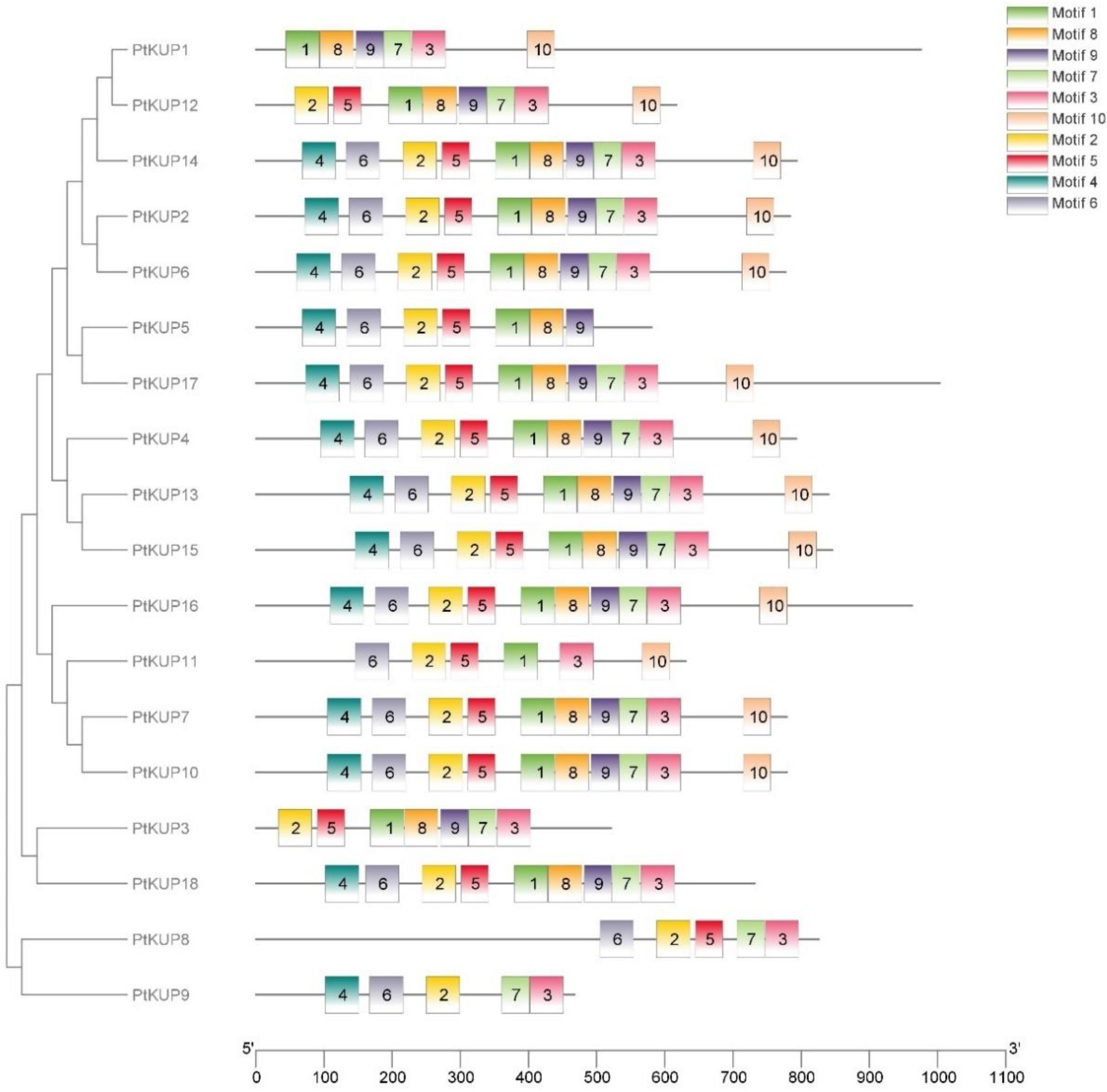
3.3. Conserved Motifs Analysis of PtKUP Gene Family
3.4. Expression Analysis of PtKUP Gene Family in Different Tissues
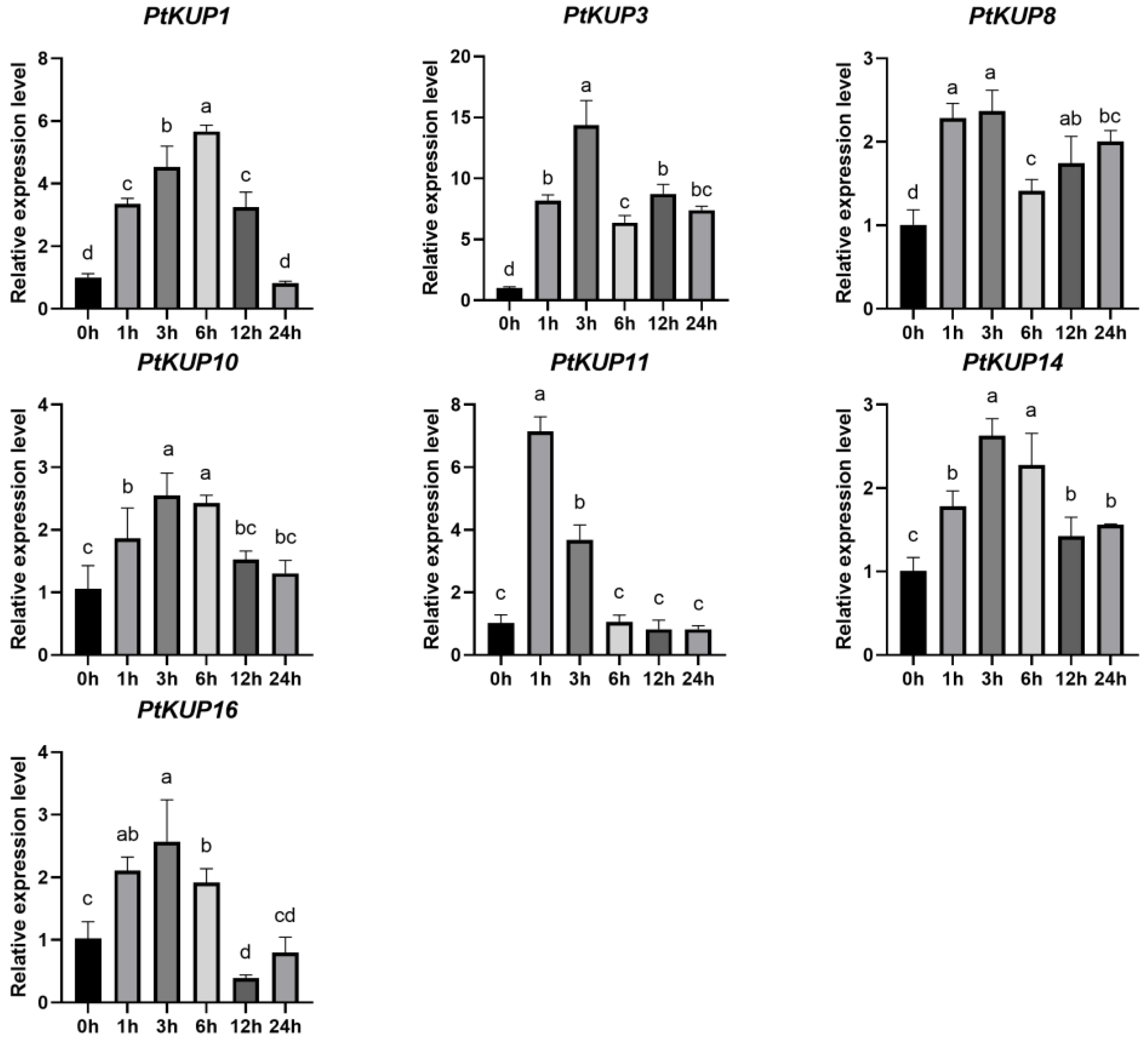
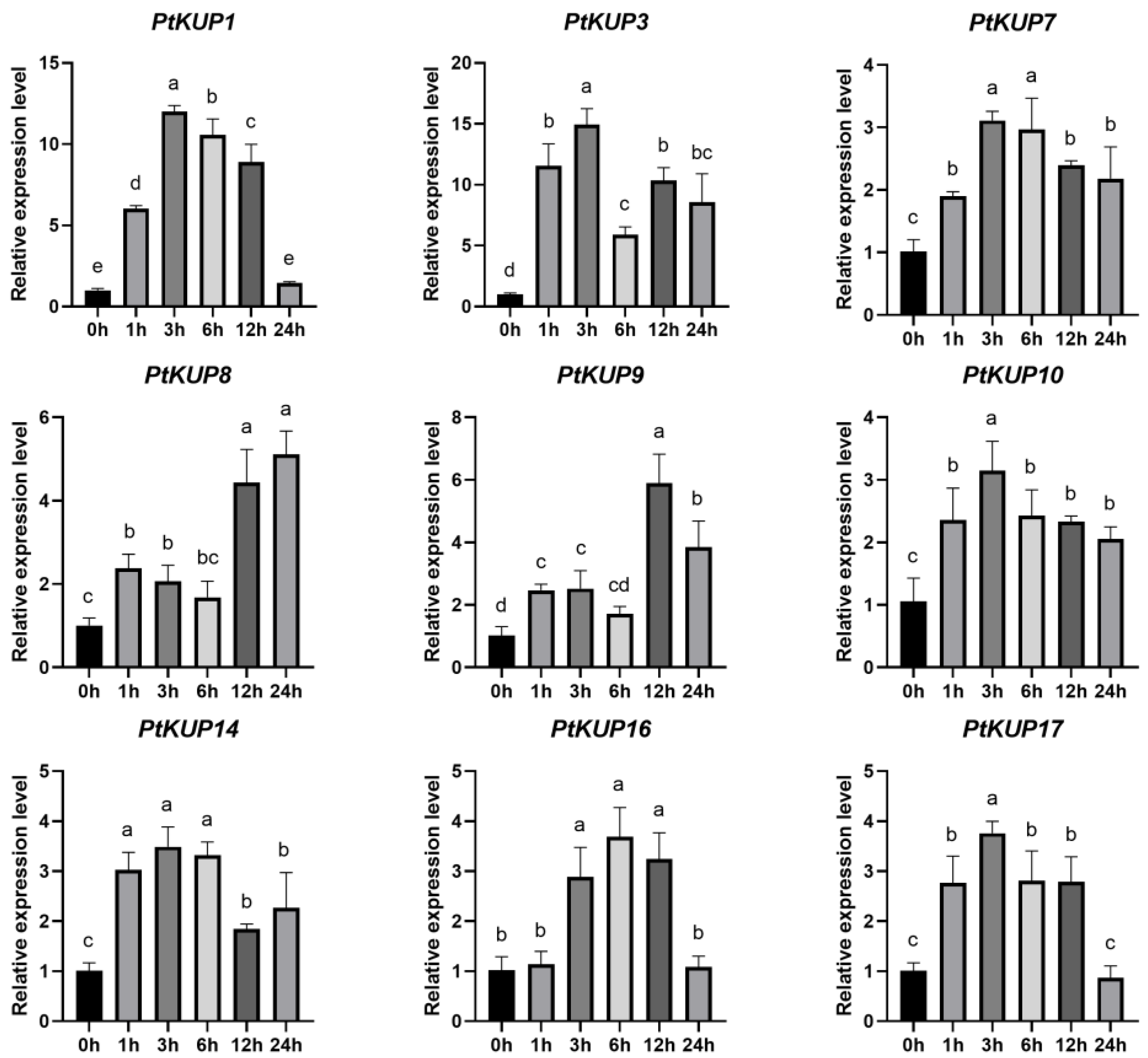
3.5. Expression Analysis of PtKUP Gene Family under Low K+ and Salt Stress
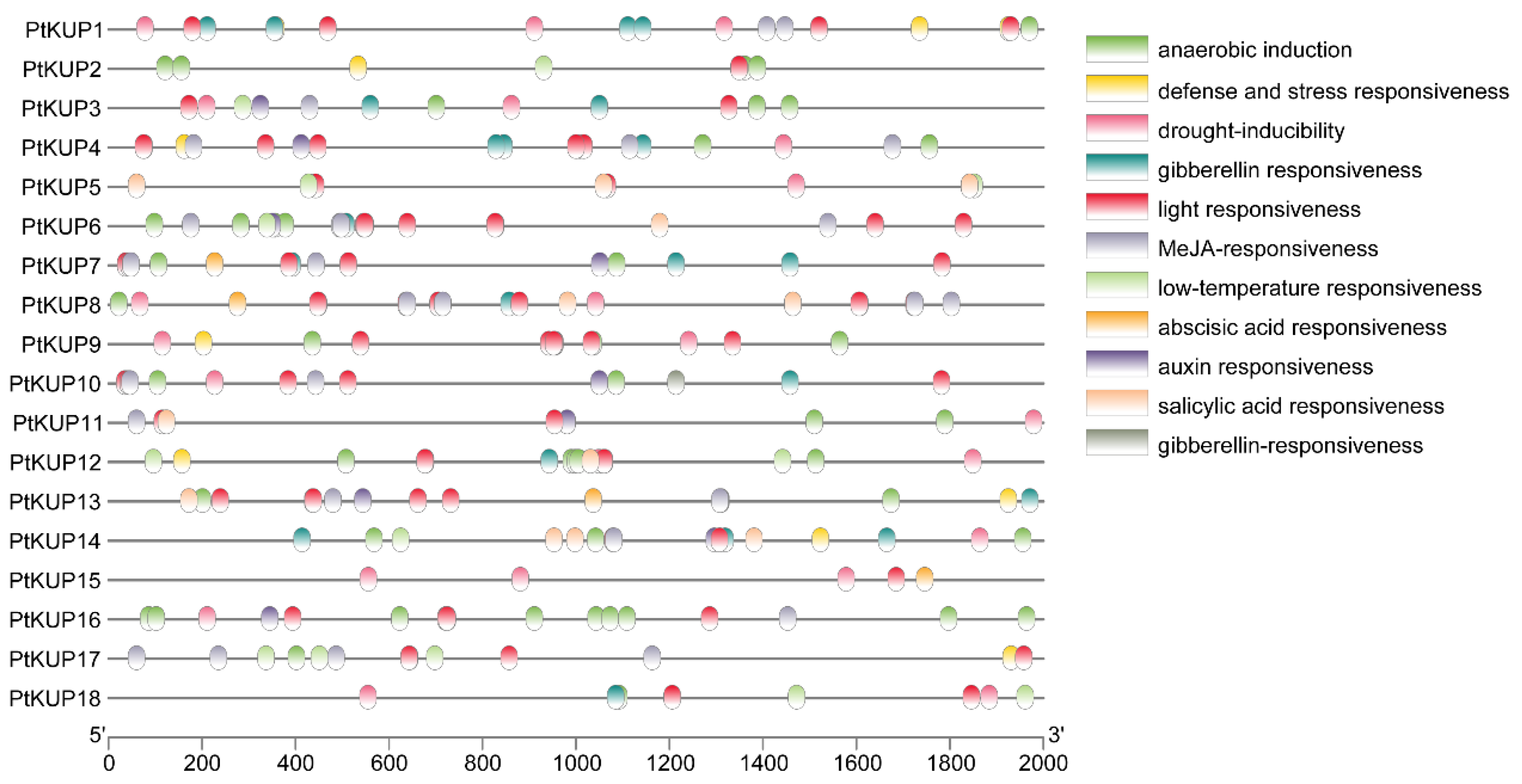
3.6. Promoter Component Analysis of PtKUP Gene Family
3.7. Functional Analysis of PtKUP10 under Salt Stress
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Pandey, G.K.; Mahiwal, S. Role of Potassium in Plants; Springer: Cham, Switzerland, 2020. [Google Scholar]
- Leigh, R.A.; Wyn Jones, R.G. A hypothesis relating critical potassium concentrations for growth to the distribution and functions of this ion in the plant cell. New Phytol. 1984, 97, 1–13. [Google Scholar] [CrossRef]
- Ou, W.; Mao, X.; Huang, C.; Tie, W.; Yan, Y.; Ding, Z.; Wu, C.; Xia, Z.; Wang, W.; Zhou, S.; et al. Genome-Wide Identification and Expression Analysis of the KUP Family under Abiotic Stress in Cassava (Manihot esculenta Crantz). Front. Physiol. 2018, 9, 17. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Bhuyan, M.; Nahar, K.; Hossain, M.; Mahmud, J.; Hossen, M.; Masud, A.; Moumita; Fujita, M. Potassium: A Vital Regulator of Plant Responses and Tolerance to Abiotic Stresses. Agronomy 2018, 8, 31. [Google Scholar] [CrossRef]
- Wang, M.; Zheng, Q.; Shen, Q.; Guo, S. The Critical Role of Potassium in Plant Stress Response. Int. J. Mol. Sci. 2013, 14, 7370–7390. [Google Scholar] [CrossRef] [PubMed]
- Almeida, D.M.; Oliveira, M.M.; Saibo, N.J.M. Regulation of Na+ and K+ homeostasis in plants: Towards improved salt stress tolerance in crop plants. Genet. Mol. Biol. 2017, 40, 326–345. [Google Scholar] [CrossRef] [PubMed]
- Amjad, M.; Akhtar, J.; Anwar-Ui-Haq, M.; Imran, S.; Jacobsen, S. Soil and foliar application of potassium enhances fruit yield and quality of tomato under salinity. Turk. J. Biol. 2014, 38, 208–218. [Google Scholar] [CrossRef]
- Gupta, M.; Qiu, X.; Wang, L.; Xie, W.; Zhang, C.; Xiong, L.; Lian, X.; Zhang, Q. KT/HAK/KUP potassium transporters gene family and their whole-life cycle expression profile in rice (Oryza sativa). Mol. Genet. Genom. 2008, 280, 437–452. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.; Chu, J.; Wang, Y.; Chang, J.; Zhou, Y.; Jiang, X. Positive Regulatory Roles of Manihot esculenta HAK5 under K+ Deficiency or High Salt Stress. Plants 2024, 13, 849. [Google Scholar] [CrossRef] [PubMed]
- Morton, M.J.L.; Awlia, M.; Al-Tamimi, N.; Saade, S.; Pailles, Y.; Negrão, S.; Tester, M. Salt stress under the scalpel—Dissecting the genetics of salt tolerance. Plant J. 2019, 97, 148–163. [Google Scholar] [CrossRef]
- Mäser, P.; Thomine, S.; Schroeder, J.I.; Ward, J.M.; Hirschi, K.; Sze, H.; Talke, I.N.; Amtmann, A.; Maathuis, F.J.; Sanders, D.; et al. Phylogenetic relationships within cation transporter families of Arabidopsis. Plant Physiol. 2001, 126, 1646–1667. [Google Scholar] [CrossRef]
- Rubio, F.; Nieves-Cordones, M.; Alemán, F.; Martínez, V. Relative contribution of AtHAK5 and AtAKT1 to K+ uptake in the high-affinity range of concentrations. Physiol. Plant. 2008, 134, 598–608. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.J.; Kwak, J.M.; Uozumi, N.; Schroeder, J.I. AtKUP1: An Arabidopsis gene encoding high-affinity potassium transport activity. Plant Cell 1998, 10, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Gierth, M.; Maser, P.; Schroeder, J.I. The potassium transporter AtHAK5 functions in K+ deprivation-induced high-affinity K+ uptake and AKT1 K+ channel contribution to K+ uptake kinetics in Arabidopsis roots. Plant Physiol. 2005, 137, 1105–1114. [Google Scholar] [CrossRef] [PubMed]
- Han, M.; Wu, W.; Wu, W.; Wang, Y. Potassium Transporter KUP7 Is Involved in K+ Acquisition and Translocation in Arabidopsis Root under K+-Limited Conditions. Mol. Plant 2016, 9, 437–446. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Hu, Q.; Luo, L.; Yang, T.; Zhang, S.; Hu, Y.; Yu, L.; Xu, G. Rice potassium transporter OsHAK1 is essential for maintaining potassium-mediated growth and functions in salt tolerance over low and high potassium concentration ranges. Plant Cell Environ. 2015, 38, 2747–2765. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Cao, H.; Cheng, J.; He, X.; Sohail, H.; Niu, M.; Huang, Y.; Bie, Z. Pumpkin CmHKT1;1 Controls Shoot Na⁺ Accumulation via Limiting Na⁺ Transport from Rootstock to Scion in Grafted Cucumber. Int. J. Mol. Sci. 2018, 19, 2648. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Chen, C.; Cai, H.; Wu, L. Overexpression of PeHKT1;1 Improves Salt Tolerance in Populus. Genes 2018, 9, 475. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Li, X.; Wang, X.; Feng, J.; Zhu, S. Potassium fulvic acid alleviates salt stress of citrus by regulating rhizosphere microbial community, osmotic substances, and enzyme activities. Front. Plant Sci. 2023, 14, 1161469. [Google Scholar] [CrossRef] [PubMed]
- Bonnin, M.; Favreau, B.; Soriano, A.; Leonhardt, N.; Oustric, J.; Lourkisti, R.; Ollitrault, P.; Morillon, R.; Berti, L.; Santini, J. Insight into Physiological and Biochemical Determinants of Salt Stress Tolerance in Tetraploid Citrus. Antioxidants 2023, 12, 1640. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Liu, X.; Mao, W.; Zhang, X.; Chen, S.; Zhan, K.; Bi, H.; Xu, H. Genome-Wide Identification and Analysis of HAK/KUP/KT Potassium Transporters Gene Family in Wheat (Triticum aestivum L.). Int. J. Mol. Sci. 2018, 19, 3969. [Google Scholar] [CrossRef]
- Feng, X.; Wang, Y.; Zhang, N.; Wu, Z.; Zeng, Q.; Wu, J.; Wu, X.; Wang, L.; Zhang, J.; Qi, Y. Genome-wide systematic characterization of the HAK/KUP/KT gene family and its expression profile during plant growth and in response to low-K+ stress in Saccharum. BMC Plant Biol. 2020, 20, 20. [Google Scholar] [CrossRef]
- Chou, K.; Shen, H. Cell-PLoc: A package of Web servers for predicting subcellular localization of proteins in various organisms. Nat. Protoc. 2008, 3, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Li, X.; Ma, L.; Mei, X.; Liu, Y.; Huang, H.; Promponas, V.J. ggmotif: An R Package for the extraction and visualization of motifs from MEME software. PLoS ONE 2022, 17, e276979. [Google Scholar] [CrossRef] [PubMed]
- Rombauts, S.; Déhais, P.; Van Montagu, M.; Rouzé, P. PlantCARE, a plant cis-acting regulatory element database. Nucleic Acids Res. 1999, 27, 295–296. [Google Scholar] [CrossRef]
- Wang, M.; Zhang, X.; Liu, J.H. Deep sequencing-based characterization of transcriptome of trifoliate orange (Poncirus trifoliata (L.) Raf.) in response to cold stress. BMC Genom. 2015, 16, 555. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. Cell Mol. Biol. 1998, 16, 735–743. [Google Scholar] [CrossRef]
- Rus, A.; Lee, B.; Muñoz-Mayor, A.; Sharkhuu, A.; Miura, K.; Zhu, J.; Bressan, R.A.; Hasegawa, P.M. AtHKT1 facilitates Na+ homeostasis and K+ nutrition in planta. Plant Physiol. 2004, 136, 2500–2511. [Google Scholar] [CrossRef]
- Ling, Q.; Huang, W.; Jarvis, P. Use of a SPAD-502 meter to measure leaf chlorophyll concentration in Arabidopsis thaliana. Photosynth. Res. 2011, 107, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Jiao, Y.; Sha, C.; Shu, Q. Integrated Physiological and Metabolomic Analyses of the Effect of Potassium Fertilizer on Citrus Fruit Splitting. Plants 2022, 11, 499. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Guo, L.; Luo, L.; Liu, Y.; Peng, S. Identification of the magnesium transport (MGT) family in Poncirus trifoliata and functional characterization of PtrMGT5 in magnesium deficiency stress. Plant Mol. Biol. 2019, 101, 551–560. [Google Scholar] [CrossRef] [PubMed]
- Hussain, S.; Morillon, R.; Anjum, M.A.; Ollitrault, P.; Costantino, G.; Luro, F. Genetic diversity revealed by physiological behavior of citrus genotypes subjected to salt stress. Acta Physiol. Plant 2014, 37, 1740. [Google Scholar] [CrossRef]
- Rubio, F.; Santa-María, G.E.; Rodríguez-Navarro, A. Cloning of Arabidopsis and barley cDNAs encoding HAK potassium transporters in root and shoot cells. Physiol. Plant. 2000, 109, 34–43. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, J.; Chen, Y.; Li, R.; Wang, H.; Wei, J. Genome-wide analysis and identification of HAK potassium transporter gene family in maize (Zea mays L.). Mol. Biol. Rep. 2012, 39, 8465–8473. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Tan, J.; Jiao, Y.; Huang, B.; Ma, R.; Ramakrishnan, M.; Qi, G.; Zhang, Z. Genome-wide identification and expression analysis of the HAK/KUP/KT gene family in Moso bamboo. Front. Plant Sci. 2024, 15, 1331710. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Lu, X.; Wang, Y.; Xie, Y.; Ma, J.; Cheng, X.; Xia, E.; Wan, X.; Zhang, Z. HAK/KUP/KT family potassium transporter genes are involved in potassium deficiency and stress responses in tea plants (Camellia sinensis L.): Expression and functional analysis. BMC Genom. 2020, 21, 556. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Peng, L.; Xie, C.; Shi, X.; Dong, C.; Shen, Q.; Xu, Y. Genome-wide identification, characterization, and expression analyses of the HAK/KUP/KT potassium transporter gene family reveals their involvement in K+ deficient and abiotic stress responses in pear rootstock seedlings. Plant Growth Regul. 2018, 85, 187–198. [Google Scholar] [CrossRef]
- Pyo, Y.J.; Gierth, M.; Schroeder, J.I.; Cho, M.H. High-affinity K+ transport in Arabidopsis: AtHAK5 and AKT1 are vital for seedling establishment and post germination growth under low-potassium conditions. Plant Physiol. 2010, 153, 863–875. [Google Scholar] [CrossRef]
- Ahn, S.J.; Shin, R.; Schachtman, D.P. Expression of KT/KUP genes in Arabidopsis and the role of root hairs in K+ uptake. Plant Physiol. 2004, 134, 1135–1145. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Zhang, S.; Hu, Y.; Wu, F.; Hu, Q.; Chen, G.; Cai, J.; Wu, T.; Moran, N.; Yu, L.; et al. The Role of a Potassium Transporter OsHAK5 in Potassium Acquisition and Transport from Roots to Shoots in Rice at Low Potassium Supply Levels. Plant Physiol. 2014, 166, 945–959. [Google Scholar] [CrossRef] [PubMed]
- Feng, H.; Tang, Q.; Cai, J.; Xu, B.; Xu, G.; Yu, L. Rice OsHAK16 functions in potassium uptake and translocation in shoot, maintaining potassium homeostasis and salt tolerance. Planta 2019, 250, 549–561. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; Ben-Gal, A.; Shtein, I.; Bustan, A.; Dag, A.; Erel, R. Root structural plasticity enhances salt tolerance in mature olives. Environ. Exp. Bot. 2020, 179, 104224. [Google Scholar] [CrossRef]
- Skodra, C.; Michailidis, M.; Dasenaki, M.; Ganopoulos, I.; Thomaidis, N.S.; Tanou, G.; Molassiotis, A. Unraveling salt-responsive tissue-specific metabolic pathways in olive tree. Physiol. Plant. 2021, 173, 1643–1656. [Google Scholar] [CrossRef]
- Zhu, J. Abiotic Stress Signaling and Responses in Plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Zhou, T.; Wu, P.; Chen, J.; Du, X.; Feng, Y.; Hua, Y. Pectin demethylation-mediated cell wall Na+ retention positively regulates salt stress tolerance in oilseed rape. Theor. Appl. Genet. 2024, 137, 54. [Google Scholar] [CrossRef] [PubMed]
- Ziska, L.H.; DeJong, T.M.; Hoffman, G.F.; Mead, R.M. Sodium and chloride distribution in salt-stressed Prunus salicina, a deciduous tree species. Tree Physiol. 1991, 8, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Parida, A.K.; Das, A.B. Salt tolerance and salinity effects on plants: A review. Ecotox Environ. Safe 2005, 60, 324–349. [Google Scholar] [CrossRef]
- Li, W.; Xu, G.; Alli, A.; Yu, L. Plant HAK/KUP/KT K+ transporters: Function and regulation. Semin. Cell Dev. Biol. 2018, 74, 133–141. [Google Scholar] [CrossRef]
- Qiu, J.; Henderson, S.W.; Tester, M.; Roy, S.J.; Gilliham, M. SLAH1, a homologue of the slow type anion channel SLAC1, modulates shoot Cl⁻ accumulation and salt tolerance in Arabidopsis thaliana. J. Exp. Bot. 2016, 67, 4495–4505. [Google Scholar] [CrossRef] [PubMed]
- Tester, M.; Davenport, R. Na+ tolerance and Na+ transport in higher plants. Ann. Bot. 2003, 91, 503–527. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Accession | AA No. | MW (kD) | pI | Gravy | Chromosome Location | Exons No. | Subcellular Localization |
|---|---|---|---|---|---|---|---|---|
| PtKUP1 | Pt2g007790.1 | 975 | 109.66 | 7.81 | −0.147 | 2 | 11 | plasmalemma |
| PtKUP2 | Pt2g009280.1 | 783 | 87.36 | 8.41 | 0.432 | 2 | 9 | plasmalemma |
| PtKUP3 | Pt2g017510.1 | 520 | 59.26 | 8.26 | 0.369 | 2 | 6 | plasmalemma |
| PtKUP4 | Pt2g021520.1 | 792 | 88.48 | 8.38 | 0.369 | 2 | 8 | plasmalemma |
| PtKUP5 | Pt2g022230.1 | 580 | 64.19 | 9.00 | 0.645 | 2 | 10 | plasmalemma |
| PtKUP6 | Pt3g002490.1 | 776 | 87.28 | 9.35 | 0.318 | 3 | 8 | plasmalemma |
| PtKUP7 | Pt3g033530.1 | 778 | 87.26 | 6.88 | 0.249 | 3 | 8 | plasmalemma |
| PtKUP8 | Pt3g033540.1 | 825 | 93.32 | 8.90 | −0.166 | 3 | 16 | plasmalemma |
| PtKUP9 | Pt3g037370.1 | 467 | 52.66 | 9.41 | 0.383 | 3 | 7 | plasmalemma |
| PtKUP10 | Pt3g037380.1 | 778 | 87.20 | 6.67 | 0.254 | 3 | 8 | plasmalemma |
| PtKUP11 | Pt4g001420.1 | 630 | 69.65 | 6.68 | 0.254 | 4 | 9 | plasmalemma |
| PtKUP12 | Pt4g004240.1 | 616 | 68.81 | 8.78 | 0.458 | 4 | 7 | plasmalemma |
| PtKUP13 | Pt4g004250.1 | 839 | 93.01 | 5.99 | 0.385 | 4 | 9 | plasmalemma |
| PtKUP14 | Pt4g020610.1 | 793 | 88.49 | 7.59 | 0.366 | 4 | 9 | plasmalemma |
| PtKUP15 | Pt7g001340.1 | 845 | 93.85 | 5.37 | 0.305 | 7 | 10 | plasmalemma |
| PtKUP16 | Pt7g012630.1 | 962 | 107.35 | 8.85 | 0.195 | 7 | 10 | plasmalemma |
| PtKUP17 | Pt8g012100.1 | 1002 | 110.51 | 6.46 | 0.230 | 8 | 16 | plasmalemma |
| PtKUP18 | PtUn023130.1 | 731 | 81.77 | 8.69 | 0.363 | Un | 8 | plasmalemma |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, L.; Yin, X.; Wen, M.; Huang, B.; Liu, F.; Li, X.; Wang, P. Genome-Wide Identification, Characterization, and Expression of the HAK/KUP/KT Potassium Transporter Gene Family in Poncirus trifoliata and Functional Analysis of PtKUP10 under Salt Stress. Horticulturae 2024, 10, 628. https://doi.org/10.3390/horticulturae10060628
Jin L, Yin X, Wen M, Huang B, Liu F, Li X, Wang P. Genome-Wide Identification, Characterization, and Expression of the HAK/KUP/KT Potassium Transporter Gene Family in Poncirus trifoliata and Functional Analysis of PtKUP10 under Salt Stress. Horticulturae. 2024; 10(6):628. https://doi.org/10.3390/horticulturae10060628
Chicago/Turabian StyleJin, Longfei, Xinxing Yin, Mingxia Wen, Bei Huang, Feng Liu, Xinguo Li, and Peng Wang. 2024. "Genome-Wide Identification, Characterization, and Expression of the HAK/KUP/KT Potassium Transporter Gene Family in Poncirus trifoliata and Functional Analysis of PtKUP10 under Salt Stress" Horticulturae 10, no. 6: 628. https://doi.org/10.3390/horticulturae10060628
APA StyleJin, L., Yin, X., Wen, M., Huang, B., Liu, F., Li, X., & Wang, P. (2024). Genome-Wide Identification, Characterization, and Expression of the HAK/KUP/KT Potassium Transporter Gene Family in Poncirus trifoliata and Functional Analysis of PtKUP10 under Salt Stress. Horticulturae, 10(6), 628. https://doi.org/10.3390/horticulturae10060628