
Freezing Points of Fruit from Different Kiwifruit Genotypes at Harvest and during Cold Storage

 ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Fruit Materials
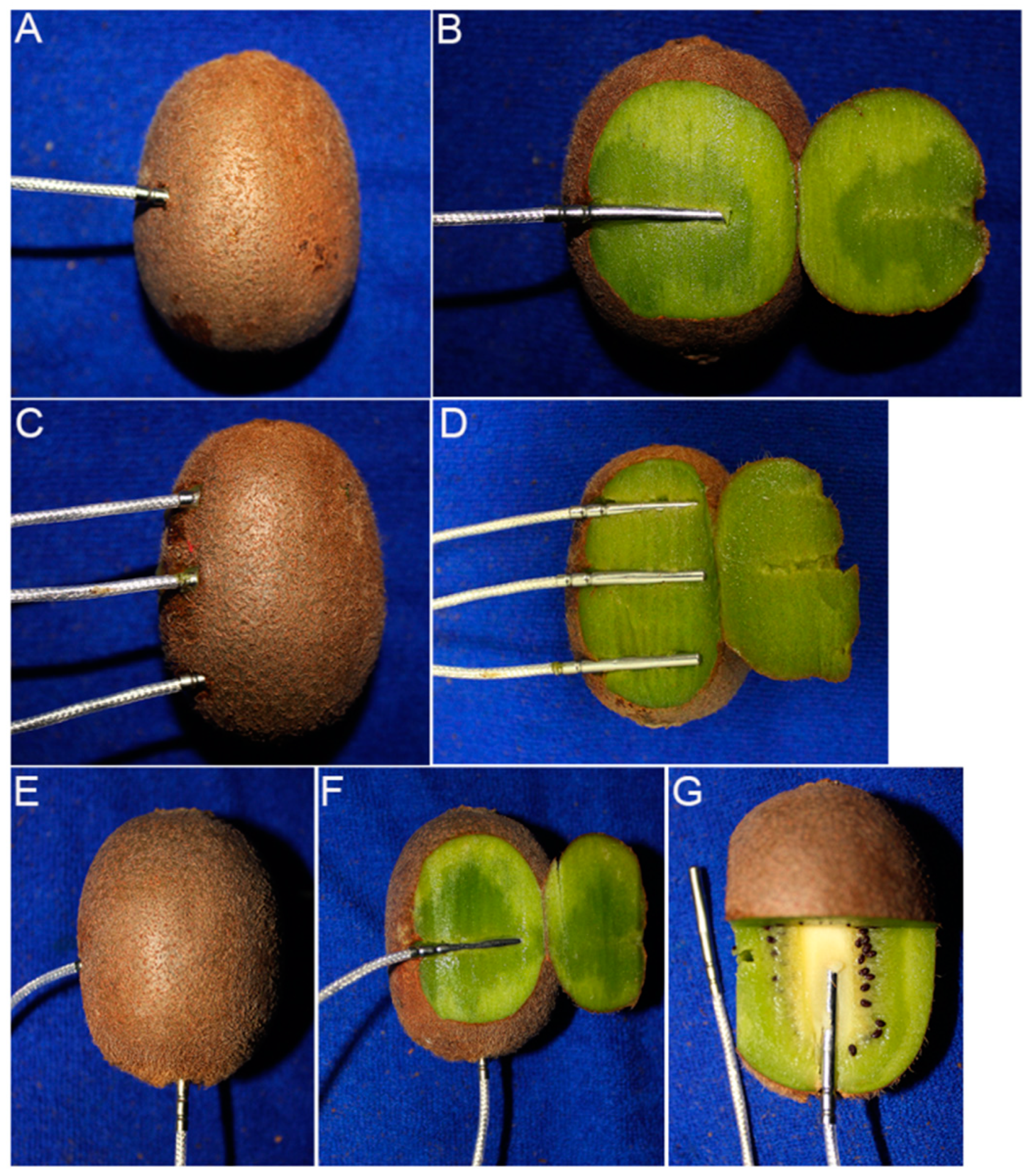
2.2. Trial Designs
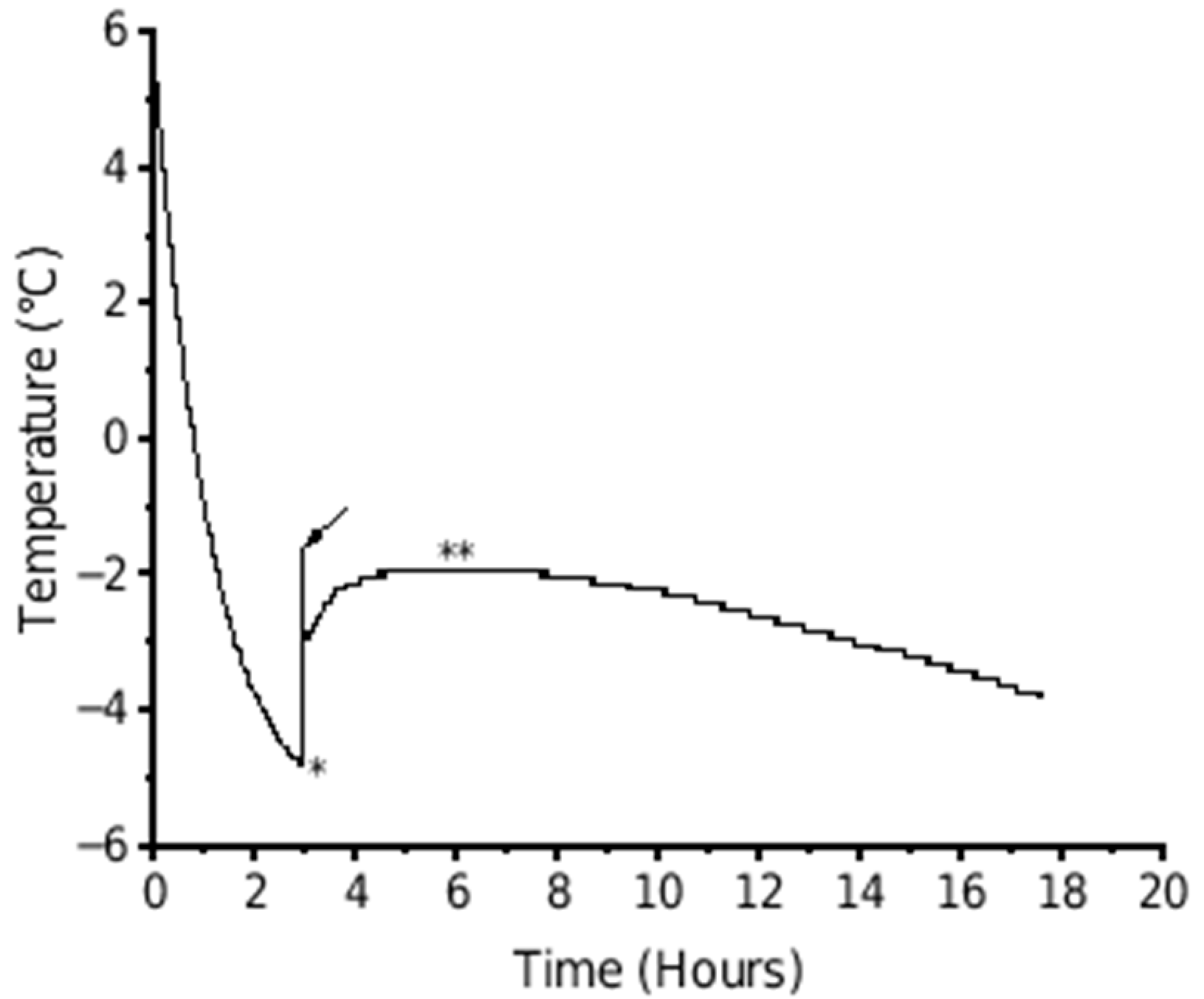
2.3. Freezing Point, Soluble Solids Content, and Water Content Measurements
2.4. Data Analysis
3. Results
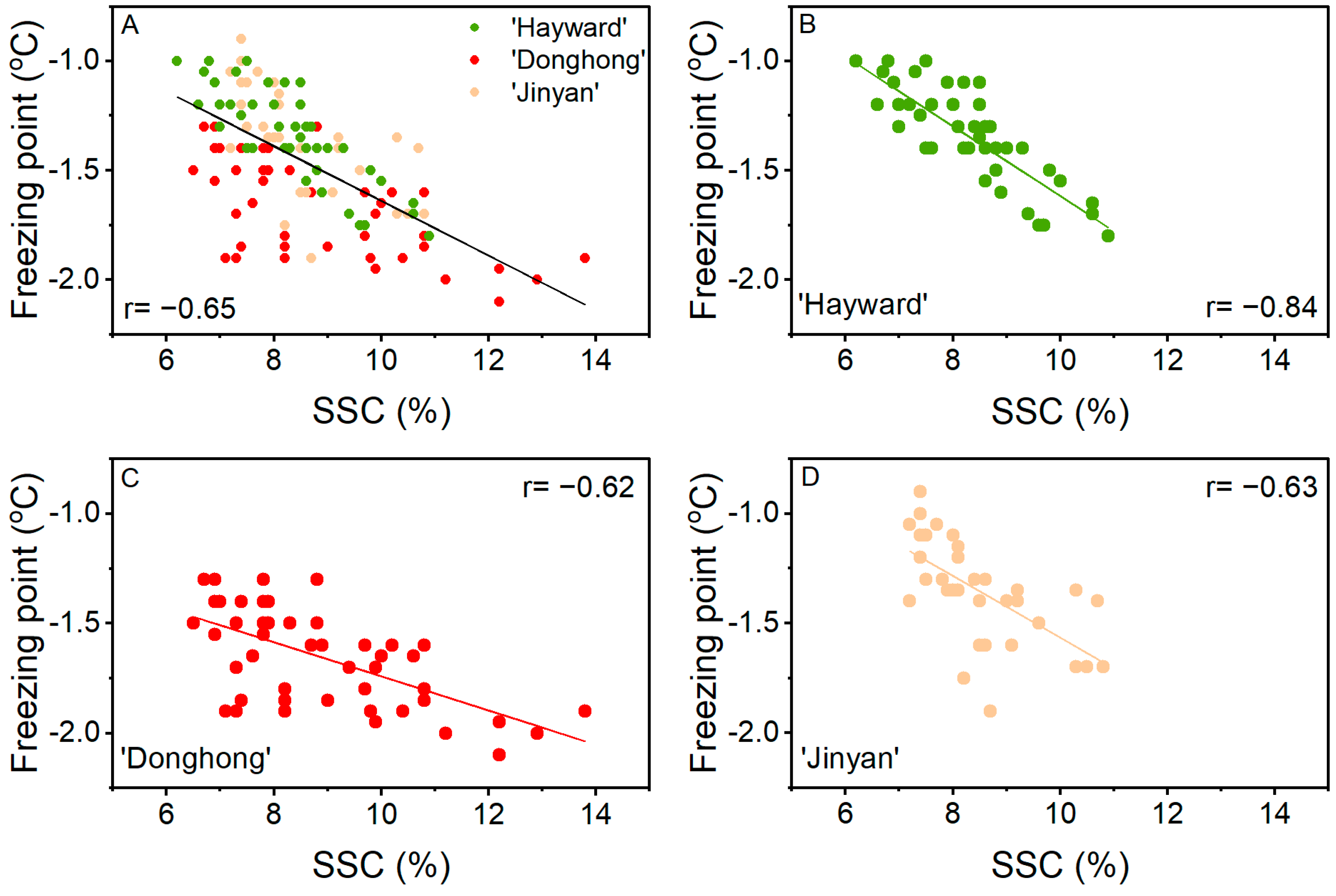
3.1. Trial 1: Freezing Point of Fruit of Different Kiwifruit Accessions at Harvest
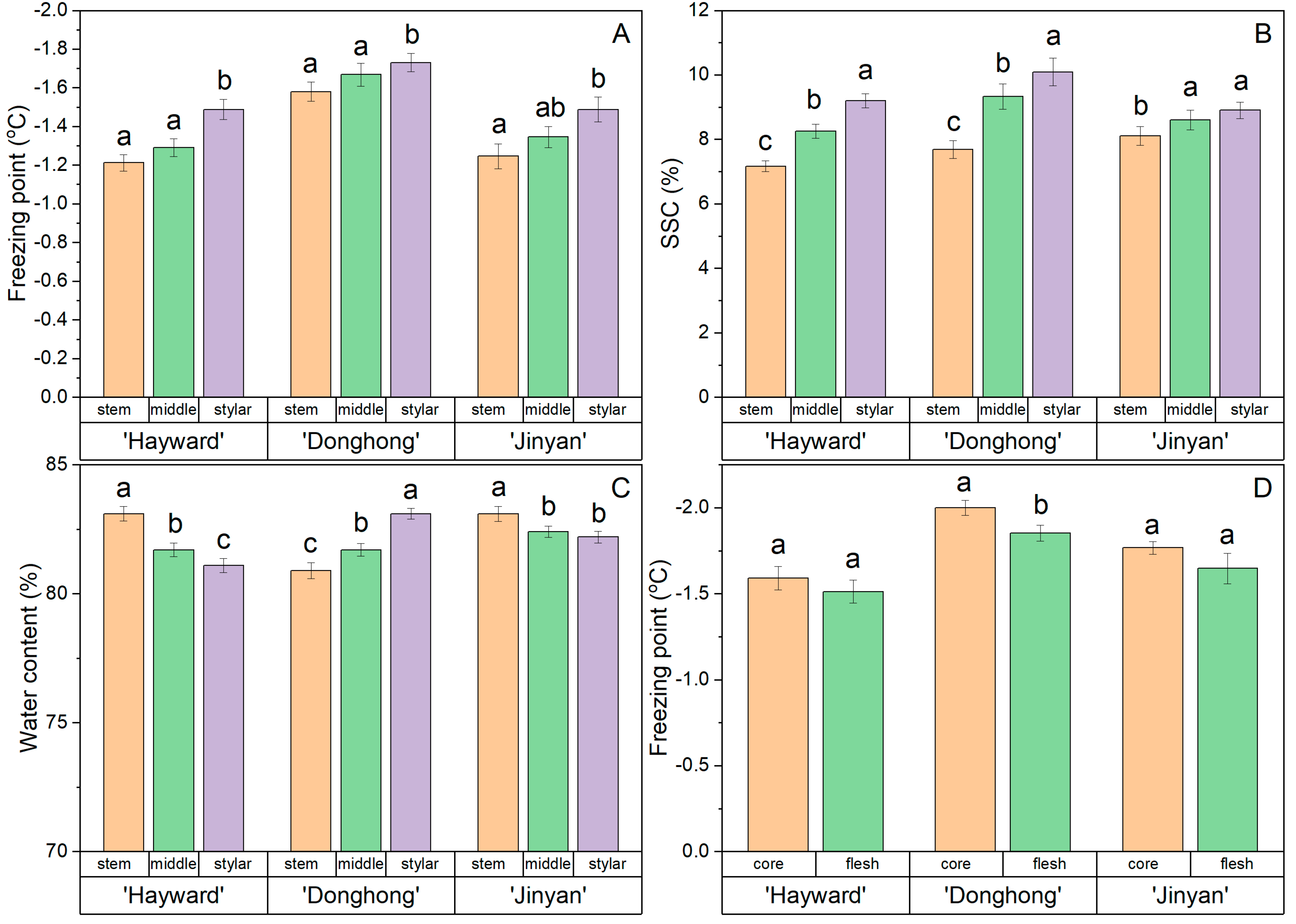
3.2. Trial 2: Freezing Point of Different Positions within Fruits
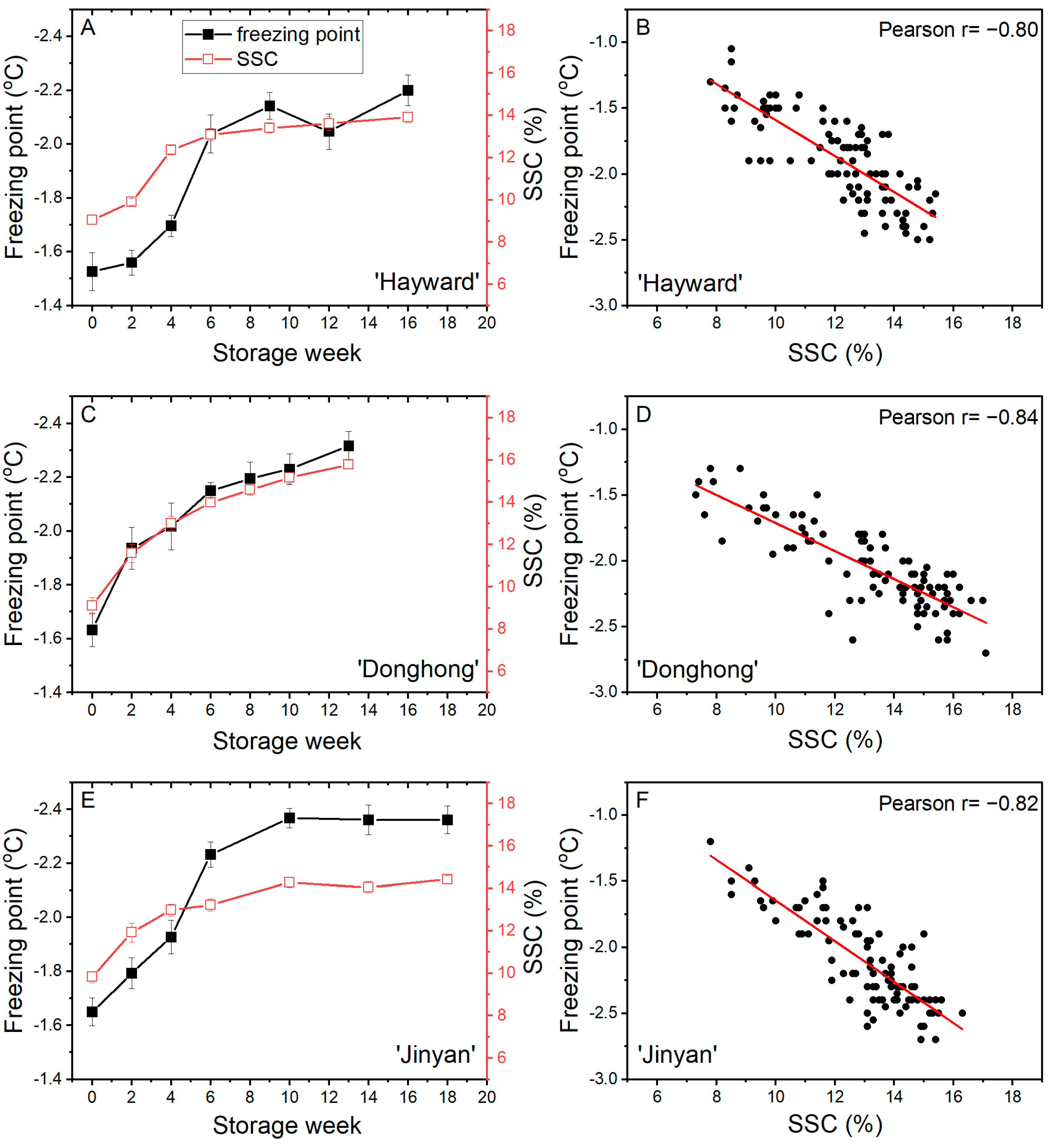
3.3. Trial 3: Freezing Point of Fruits during Cold Storage
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chamberlain, J.E. Global industry and markets. In Kiwifruit: Botany, Production and Uses; Richardson, A.C., Burdon, J.N., Ferguson, A.R., Eds.; CABI: Wallingford, UK, 2024; pp. 16–28. [Google Scholar]
- Garcia, C.V.; Quek, S.-Y.; Stevenson, R.J.; Winz, R.A. Kiwifruit flavour: A review. Trends Food Sci. Technol. 2012, 24, 82–91. [Google Scholar] [CrossRef]
- Richardson, D.P.; Ansell, J.; Drummond, L.N. The nutritional and health attributes of kiwifruit: A review. Eur. J. Nutr. 2018, 57, 2659–2676. [Google Scholar] [CrossRef] [PubMed]
- Burdon, J.N.; Wang, R. Postharvest: Fresh fruit harvest, storage and supply. In Kiwifruit: Botany, Production and Uses; Richardson, A.C., Burdon, J.N., Ferguson, A.R., Eds.; CABI: Wallingford, UK, 2024; pp. 353–374. [Google Scholar]
- Paull, R.E. Chilling injury of crops of tropical and subtropical origin. In Chilling Injury of Horticultural Crops; Wang, C., Ed.; CRC Press: Boca Raton, FL, USA, 1990. [Google Scholar]
- Lallu, N. Low temperature breakdown in kiwifruit. Acta Hortic. 1997, 444, 579–585. [Google Scholar] [CrossRef]
- Jie, W.; Lite, L.; Yang, D. The correlation between freezing point and soluble solids of fruits. J. Food Eng. 2003, 60, 481–484. [Google Scholar] [CrossRef]
- Mitchell, F.; Mayer, G.; Maxie, E.; Coates, W. Cold storage effects on fresh market peaches, nectarines & plums estimating freezing points using low temperatures to delay internal breakdown. Calif. Agric. 1974, 28, 12–14. [Google Scholar]
- Sirijariyawat, A.; Charoenrein, S. Freezing characteristics and texture variation after freezing and thawing of four fruit types. Songklanakarin J. Sci. Technol. 2012, 34, 517–523. [Google Scholar]
- Dan, W.; Lei, S.; Jiazheng, S.; Qian, Z.; Mosen, L. A study of freezing properties of blueberry fruits and influencing factors. Acta Agric. Univ. Jiangxiensis 2013, 35, 940–944. [Google Scholar] [CrossRef]
- Weirong, H.; Zhaoqi, Z.; Yueming, J.; Zuoliang, J. Study on the parameter of ice-temperature storage in litchi (Litchi chinensis Sonn.). Sci. Agric. Sincia 2005, 38, 797–802. [Google Scholar]
- Nanos, G.D.; Fourtouni, A.; Kaltsoula, E.; Sfakiotakis, E.M. Freezing point estimation of kiwifruit. Acta Hortic. 1997, 444, 229–232. [Google Scholar] [CrossRef]
- Harris, S.; McDonald, B. Physical data for kiwifruit (Actinidia chinensis). N. Z. J. Sci. 1975, 18, 307–312. [Google Scholar]
- Gerasopoulos, D.; Chlioumis, G.; Sfakiotakis, E. Non-freezing points below zero induce low-temperature breakdown of kiwifruit at harvest. J. Sci. Food Agric. 2006, 86, 886–890. [Google Scholar] [CrossRef]
- Zhong, Z.; Zhang, M.; Yang, L.; Chen, J.; Che, Z. Study on relation between freezing and physiological and biochemical indexes of fruits and vegetables. Sci. Technol. Food Ind. 2011, 32, 76–78. [Google Scholar]
- Yan, R.; Jia, N.; Song, M.; Zhao, Y.; Zhang, P. Study on correlationship among freezing point and contents of both soluble solids and water. Food Sci. 2007, 28, 554–557. [Google Scholar]
- Ma, Y.; Zhang, X.; Li, L.; Yang, W.; Zhang, W.; Cheng, S.; Guo, M.; Chen, G. Delaying fruit softening of ‘France’ prune (Prunus domestica L.) using near-freezing temperature storage. LWT 2022, 172, 114165. [Google Scholar] [CrossRef]
- Fan, X.; Xi, Y.; Zhao, H.; Liu, B.; Cao, J.; Jiang, W. Improving fresh apricot (Prunus armeniaca L.) quality and antioxidant capacity by storage at near freezing temperature. Sci. Hortic. 2018, 231, 1–10. [Google Scholar] [CrossRef]
- Liu, B.; Jiao, W.; Wang, B.; Shen, J.; Zhao, H.; Jiang, W. Near freezing point storage compared with conventional low temperature storage on apricot fruit flavor quality (volatile, sugar, organic acid) promotion during storage and related shelf life. Sci. Hortic. 2019, 249, 100–109. [Google Scholar] [CrossRef]
- Zhao, H.; Liu, B.; Zhang, W.; Cao, J.; Jiang, W. Enhancement of quality and antioxidant metabolism of sweet cherry fruit by near-freezing temperature storage. Postharvest Biol. Technol. 2019, 147, 113–122. [Google Scholar] [CrossRef]
- Zhao, H.; Wang, B.; Cui, K.; Cao, J.; Jiang, W. Improving postharvest quality and antioxidant capacity of sweet cherry fruit by storage at near-freezing temperature. Sci. Hortic. 2019, 246, 68–78. [Google Scholar] [CrossRef]
- Zhao, H.; Jiao, W.; Cui, K.; Fan, X.; Shu, C.; Zhang, W.; Cao, J.; Jiang, W. Near-freezing temperature storage enhances chilling tolerance in nectarine fruit through its regulation of soluble sugars and energy metabolism. Food Chem. 2019, 289, 426–435. [Google Scholar] [CrossRef]
- Zhong, C.; Huang, W.; Wang, Z.; Li, L.; Li, D.; Zhang, Q.; Zhao, T.; Zhang, P. The breeding progress and development status of the kiwifruit industry in China. Acta Hortic. 2022, 1332, 445–454. [Google Scholar] [CrossRef]
- Liao, G.; Li, X.; Chen, L.; Huang, C.; Zhong, M.; Xu, X. Difference analysis on fruit quality among different parts of kiwifruit. North. Hortic. 2019, 06, 34–42. [Google Scholar] [CrossRef]
- MacRae, E.A.; Bowen, J.H.; Stec, M.G. Maturation of kiwifruit (Actinidia deliciosa cv Hayward) from two orchards: Differences in composition of the tissue zones. J. Sci. Food Agric. 1989, 47, 401–416. [Google Scholar] [CrossRef]
- Zhong, C.; Han, F.; Li, D.; Liu, X.; Zhang, Q.; Jiang, Z.; Huang, H. Breeding of red-fleshed kiwifruit cultivar ‘Donghong’. J. Fruit Sci. 2016, 33, 1596–1599. [Google Scholar]
- Zhong, C.; Wang, S.; Jiang, Z.; Huang, H. ‘Jinyan’, an interspecific hybrid kiwifruit with brilliant yellow flesh and good storage quality. HortScience 2012, 47, 1187–1190. [Google Scholar] [CrossRef]
- Rahman, M.S.; Guizani, N.; Al-Khaseibi, M.; Ali Al-Hinai, S.; Al-Maskri, S.S.; Al-Hamhami, K. Analysis of cooling curve to determine the end point of freezing. Food Hydrocoll. 2002, 16, 653–659. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession | Genetic Background | Freezing Point (°C) | SSC (%) | WC (%) | Correlation (r) |
|---|---|---|---|---|---|
| SZ18 | A. rufa × A. chinensis | −2.61 ± 0.05 | 14.2 ± 0.12 | 78.2 ± 0.26 | −0.37 |
| ‘Dongmei’ | A. chinensis var. deliciosa | −2.34 ± 0.05 | 15.6 ± 0.31 | 76.4 ± 0.55 | −0.17 |
| ‘Jinměi’ | A. chinensis var. deliciosa | −2.32 ± 0.05 | 16.6 ± 0.18 | 76.0 ± 0.51 | 0.19 |
| ‘Jinzao’ | A. chinensis var. chinensis | −2.28 ± 0.06 | 14.8 ± 0.33 | 79.7 ± 0.47 | −0.74 |
| ‘Huayou’ | A. chinensis var. chinensis | −2.25 ± 0.04 | 13.5 ± 0.27 | 76.8 ± 0.30 | −0.81 |
| ZK8 | A. rufa × A. chinensis | −2.21 ± 0.06 | 13.1 ± 0.15 | 76.9 ± 0.16 | −0.79 |
| ZK4 | A. chinensis var. deliciosa | −2.18 ± 0.05 | 13.4 ± 0.31 | 79.7 ± 0.19 | −0.39 |
| SZ17 | A. rufa × A. chinensis | −2.10 ± 0.02 | 13.1 ± 0.36 | 78.7 ± 0.19 | −0.21 |
| ZH-CD611 | A. chinensis var. chinensis | −2.10 ± 0.09 | 11.7 ± 0.64 | 78.2 ± 0.22 | −0.86 |
| ZK3 | A. chinensis var. deliciosa | −2.08 ± 0.11 | 12.5 ± 0.50 | 80.0 ± 0.33 | −0.94 |
| ‘Wuzhi 3’ | A. chinensis var. chinensis | −2.05 ± 0.07 | 12.3 ± 0.43 | 80.6 ± 0.40 | −0.62 |
| ‘Jinfeng’ | A. chinensis var. chinensis | −2.03 ± 0.07 | 11.4 ± 0.26 | 81.9 ± 0.23 | −0.73 |
| ‘Qinxiang’ | A. chinensis var. deliciosa | −2.03 ± 0.06 | 12.2 ± 0.25 | 80.2 ± 0.25 | −0.67 |
| SZ15 | A. rufa × A. chinensis | −2.02 ± 0.06 | 12.3 ± 0.28 | 79.2 ± 0.26 | −0.79 |
| ZK9 | A. rufa × A. chinensis | −2.01 ± 0.04 | 12.6 ± 0.21 | 79.4 ± 0.24 | −0.75 |
| ‘Jinyuan’ | A. chinensis × A. eriantha | −2.00 ± 0.07 | 11.4 ± 0.39 | 80.7 ± 0.37 | −0.82 |
| ZK12 | A. chinensis × A. eriantha | −1.99 ± 0.03 | 13.4 ± 0.21 | 79.5 ± 0.24 | −0.22 |
| SZ14 | A. rufa × A. chinensis | −1.98 ± 0.04 | 12.8 ± 0.28 | 79.4 ± 0.23 | −0.59 |
| ZK5 | A. rufa × A. chinensis | −1.97 ± 0.05 | 12.2 ± 0.30 | 80.0 ± 0.21 | −0.59 |
| ‘Jinyu’ | A. chinensis var. chinensis | −1.97 ± 0.04 | 13.8 ± 0.42 | 77.4 ± 0.29 | −0.43 |
| ‘Jinnong’ | A. chinensis var. chinensis | −1.87 ± 0.06 | 12.7 ± 0.27 | 80.5 ± 0.18 | −0.61 |
| ZK6 | A. rufa × A. chinensis | −1.86 ± 0.06 | 11.4 ± 0.25 | 81.3 ± 0.23 | −0.41 |
| Ace1 | A. chinensis × A. eriantha | −1.86 ± 0.05 | 10.4 ± 0.15 | 82.7 ± 0.28 | 0.14 |
| ZH-CD11129 | A. chinensis var. chinensis | −1.80 ± 0.09 | 8.6 ± 0.22 | 80.1 ± 0.27 | −0.08 |
| Xuxiang-L | A. chinensis var. deliciosa | −1.76 ± 0.05 | 11.0 ± 0.23 | 81.2 ± 0.27 | −0.08 |
| SZ16 | A. rufa × A. chinensis | −1.74 ± 0.07 | 9.2 ± 0.15 | 80.2 ± 0.38 | −0.62 |
| ‘Jinmēi’ | A. chinensis × A. eriantha | −1.70 ± 0.07 | 8.9 ± 0.26 | 84.8 ± 0.18 | −0.27 |
| ‘Donghong’ | A. chinensis var. chinensis | −1.68 ± 0.06 | 9.2 ± 0.35 | 81.7 ± 0.21 | −0.75 |
| ZK2 | A. chinensis var. deliciosa | −1.67 ± 0.05 | 9.0 ± 0.28 | 81.4 ± 0.22 | −0.56 |
| ‘Mantianhong’ | A. chinensis × A. eriantha | −1.66 ± 0.06 | 10.5 ± 0.44 | 76.8 ± 0.45 | −0.93 |
| ZK7 | A. rufa × A. chinensis | −1.64 ± 0.03 | 10.2 ± 0.17 | 79.8 ± 0.17 | −0.73 |
| ‘Jinyan’ | A. eriantha × A. chinensis | −1.63 ± 0.05 | 10.1 ± 0.26 | 81.9 ± 0.16 | −0.80 |
| ‘Jinxia’ | A. chinensis var. chinensis | −1.62 ± 0.05 | 9.1 ± 0.17 | 80.6 ± 0.26 | −0.69 |
| ‘Jintao’ | A. chinensis var. chinensis | −1.62 ± 0.05 | 9.8 ± 0.21 | 82.3 ± 0.22 | −0.78 |
| ‘Hongyang’ | A. chinensis var. chinensis | −1.62 ± 0.07 | 8.9 ± 0.35 | 79.5 ± 0.23 | −0.87 |
| ‘Jinkui’ | A. chinensis var. deliciosa | −1.57 ± 0.09 | 11.2 ± 0.54 | 78.3 ± 0.26 | −0.69 |
| ‘Miliang 1’ | A. chinensis var. deliciosa | −1.55 ± 0.07 | 9.6 ± 0.28 | 80.5 ± 0.35 | −0.93 |
| ZK1 | A. chinensis var. deliciosa | −1.54 ± 0.05 | 8.8 ± 0.17 | 82.5 ± 0.18 | −0.70 |
| ‘Mantianhong 2’ | A. chinensis × A. eriantha | −1.53 ± 0.05 | 9.3 ± 0.30 | 80.3 ± 0.22 | −0.84 |
| ‘Hort16A’ | A. chinensis var. chinensis | −1.50 ± 0.07 | 9.2 ± 0.26 | 79.8 ± 0.28 | −0.59 |
| ‘Cuiyu’ | A. chinensis var. chinensis | −1.46 ± 0.05 | 8.1 ± 0.32 | 82.9 ± 0.44 | −0.75 |
| ZK13 | A. chinensis var. deliciosa | −1.46 ± 0.06 | 9.3 ± 0.39 | 82.3 ± 0.21 | −0.58 |
| ZK10 | A.- chinensis × A. eriantha | −1.41 ± 0.04 | 8.5 ± 0.20 | 79.9 ± 0.37 | −0.69 |
| ‘Xuxiang’ | A. chinensis var. deliciosa | −1.38 ± 0.06 | 8.4 ± 0.24 | 81.0 ± 0.37 | −0.70 |
| ‘Hayward’ | A. chinensis var. deliciosa | −1.23 ± 0.02 | 7.9 ± 0.15 | 81.5 ± 0.18 | −0.36 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, W.; Shen, S.; Wang, Z.; Yang, J.; Lv, H.; Tian, H.; Burdon, J.; Zhong, C. Freezing Points of Fruit from Different Kiwifruit Genotypes at Harvest and during Cold Storage. Horticulturae 2024, 10, 624. https://doi.org/10.3390/horticulturae10060624
Huang W, Shen S, Wang Z, Yang J, Lv H, Tian H, Burdon J, Zhong C. Freezing Points of Fruit from Different Kiwifruit Genotypes at Harvest and during Cold Storage. Horticulturae. 2024; 10(6):624. https://doi.org/10.3390/horticulturae10060624
Chicago/Turabian StyleHuang, Wenjun, Suyun Shen, Zhouqian Wang, Jie Yang, Haiyan Lv, Hua Tian, Jeremy Burdon, and Caihong Zhong. 2024. "Freezing Points of Fruit from Different Kiwifruit Genotypes at Harvest and during Cold Storage" Horticulturae 10, no. 6: 624. https://doi.org/10.3390/horticulturae10060624
APA StyleHuang, W., Shen, S., Wang, Z., Yang, J., Lv, H., Tian, H., Burdon, J., & Zhong, C. (2024). Freezing Points of Fruit from Different Kiwifruit Genotypes at Harvest and during Cold Storage. Horticulturae, 10(6), 624. https://doi.org/10.3390/horticulturae10060624