Abstract
Aquatic plants, just like terrestrial plants, are exposed to oxidative stress. However, their responses are still under-researched. In this study, we examined the physiological and antioxidative responses of an aquatic plant, duckweed (Lemna minor L.), to four indole-3-acetic acid (IAA)-degrading and -producing Pseudomonas bacteria (Pseudomonas oryzihabitans D1-104/3, P. putida A3-104/5, P. gessardii C31-106/3 and P. yamanorum C44-104/1) and/or a supraphysiological level of IAA (10 mg L−1). Growth characteristics, total photosynthetic pigment content, histochemical localization of reactive oxygen species and antioxidant enzyme activity (SOD, CAT and POX) were evaluated at two time points, after 3 and 7 days of co-cultivation. Superoxide anion and hydrogen peroxide were produced and accumulated mainly in the roots, daughter fronds and veins of duckweeds. Duckweeds’ responses depended on the strain of Pseudomonas, time and exogenous IAA. Co-cultivation of duckweed with bacteria has positive or neutral effects. Exogenous application of IAA had a negative or neutral effect on enzyme activity and other parameters. Co-cultivation with P. gessardii C31-106/3 showed plant-growth-promoting effects on duckweed: increased biomass production, modulation of duckweeds’ antioxidant enzymatic activity and reduction in hydrogen peroxide content. This study widens our knowledge of aquatic plants and their response to oxidative stress, supports the hypothesis that plant growth-promoting bacteria (PGPB) induce tolerable levels of oxidative stress in plants and introduces a new PGPB strain, P. gessardii C31-106/3.
1. Introduction
Duckweed (Lemna minor L.) is an aquatic plant of simplified morphology, with high vegetative reproductive capacity, which results in a wide spectrum of aquatic niches occupied by this aquatic plant. Duckweed has applications in traditional medicine and in traditional foods of people and animals. It is even lauded as the food of the future, as these minute plants have low growth requirements and can be genetically modified to boost their nutritional value [1,2,3,4]. Furthermore, in recent decades, more attention has been directed to duckweed-based research, with the prospect of duckweeds even replacing A. thaliana, the golden standard of model organisms [5]. To survive (and thrive) even in hostile environments, duckweeds developed unique survival strategies, including a robust antioxidative response [1,6]. Many studies focused on the abiotic stress response of duckweed, as it has a prominent role in bioremediation and bioindication [7,8,9]. However, relatively little is known about its response to biotic stress.
In nature, the plants from the duckweed family (Lemnaceae), like all plants, do not exist in isolation, but create a functional entity, a holobiont, an ecological unit consisting of the plant and various microorganisms that inhabit it [10]. Due to their co-evolution with microorganisms, Lemnaceae adopted a survival strategy by developing a unique immune system that makes them resistant to various microorganisms, including bacteria [1]. It is unsurprising then that duckweeds have a complex rhizosphere microbiome, inhabited by various bacteria that can have an inhibitory, stimulating or neutral effect on the growth of duckweeds. The well-studied PGPB (plant-growth-promoting bacteria) employ aminocyclopropane-1-carboxylic acid (ACC) deaminase, indole-3-acetic acid (IAA) and gibberellic acid (GA) production, siderophores, phosphorus solubilization and transformation of nitrogen and consequently improve the growth of plants [11]. In the last decade, the modulation of oxidative response has been proposed as an additional mechanism for PGPB [12]. The consensus is that PGPB induce tolerable or very low levels of oxidative stress. Simultaneously, other mechanisms of PGPB ensure that potential benefits (e.g., increased availability of nitrogen, iron and potassium, phytohormones) outweigh the cost (increased oxidative stress) [12]. PGPB research is primarily driven by the transition to green agriculture, where synthetic fertilizers are substituted with biostimulants and biofertilizers (plant-growth-promoting microorganisms and their biochemical products) [13]. In horticulture, rhizosphere-associated PGPB are used to promote growth and improve the resistance of sensitive ornamental plants [14]. However, the oxidative aspect of PGPB effects on plants is still under-researched. What is well-established is that most PGPB produce IAA, which is mostly associated with root development and better absorption of nutrients. However, in duckweeds, roots are gradually losing their significance and becoming a vestigial organ; hence, their role is not as straightforward as in terrestrial plants [15]. Moreover, it is reported that the effects of exogenous IAA on duckweeds are not the same as in terrestrial plants [16]. But just like in other plants, the rhizosphere of duckweeds is still inhabited by many IAA-producing bacterial strains [17]. The presence of IAA-producing bacteria indicates that they serve a role in the life cycle of duckweeds, as a source of this phytohormone, while duckweeds provide nutrients and actively “recruit” rhizospheric bacteria by emitting signaling molecules [17,18]. Finally, one of the most recent developments in biotechnology is engineered microbiomes which rely on modifying the structure of naturally occurring microbiomes in vitro and enhancing their natural abilities [19]. To achieve this, our knowledge of PGPB needs to be continuously expanded, and new isolates from different sources need to be characterized.
In this research, our focus was on evaluating the physiological and antioxidative responses of duckweeds co-cultivated with rhizosphere-associated bacterial strains (Pseudomonas oryzihabitans D1-104/3, P. putida A3-104/5, P. gessardii C31-106/3 and P. yamanorum C44-104/1) in the absence and presence of exogenous IAA. According to previously reported data, these strains not only produce but also degrade IAA and may affect duckweed doubling times [17]. In addition to bacterial strains, in parallel, the effects of supraphysiological IAA (10 mg L−1) on the physiological and antioxidative response of duckweeds were analyzed. Finally, the main aim of this study was to expand the existing knowledge of structure and interactions in the rhizosphere microbiome for the future application of new biofertilizers in agriculture, including horticulture.
2. Materials and Methods
2.1. Duckweed (L. minor L.) Culture Conditions and Co-Cultivation with Pseudomonas Strains
Duckweeds purchased from a commercial nursery were surface-sterilized and maintained in Murashige–Skoog (MS) medium, at 24 ± 2 °C (under fluorescent light of 40 µmoL m−2 s−1 with 16 h light/8 h dark photoperiod) [17]. For the experiments, 150 ± 20 individual duckweeds were transferred into 50 mL MS medium, in a glass flask with flat bottom. For the co-cultivation with duckweeds, bacterial strains with a dual ability to degrade and produce IAA (P. oryzihabitans D1-104/3, P. putida A3-104/5, P. gessardii C31-106/3, P. yamanorum C44-104/1), characterized in our previous research [17], were selected for analyses in this study. Bacterial cultures were grown overnight in a thermal shaker (IKA KS 4000 i control, Staufen, Germany) at 220 rpm, 30 °C, in Luria–Bertani (LB) medium, as previously described [17]. Then, the bacterial suspensions were briefly centrifuged and washed in MS medium and transferred to flasks with surface-sterilized duckweeds (1:100; v/v; 108 CFU mL−1). Duckweeds and bacteria, as well as surface-sterilized duckweeds, were cultivated for 3 and 7 days. For the analysis of IAA effects on the antioxidative response, surface-sterilized duckweed cultures as well as bacteria–duckweed co-cultures were supplemented with IAA in a concentration of 10 mg L−1. Controls in all experiments were set as surface-sterilized duckweeds in IAA-free (IAA−) or IAA-containing (IAA+) MS medium. The plant material was then collected and frozen in liquid nitrogen and stored at −20 °C for further analyses, except for photosynthetic pigment content analysis and estimation of bacterial density, in which fresh duckweed mass was used. The surface density of bacteria was estimated by macerating 20 mg of fresh duckweeds and diluting this macerate in 1 ml of sterile double-distilled water. The ensuing suspension was then serially diluted and streaked onto LB nutrient agar in Petri dishes for colony-forming unit (CFU) counting. All samples were presented in triplicates. CFU values were calculated from the mean value of dilution plates, as described previously [17].
2.2. Photosynthetic Pigment Content
For the isolation of chlorophyll and carotenoids, plant material from culture (about 20 mg) was transferred into 2 mL Eppendorf tubes and soaked in 2 mL 96% ethanol (Zorka Pharma Hemija doo, Šabac, Serbia). Test tubes with ethanol and plant material were incubated in a water bath (Univeba JP Selecta, Barcelona, Spain) at 70 °C, for a period of 10 min. The content of chlorophyll and carotenoids was measured on a spectrophotometer (UV-VIS Agilent 8453, Agilent Technologies, Santa Clara, CA, USA). The absorbance was recorded at three wavelengths: 470 nm, 648 nm and 664 nm. Concentrations of chlorophyll and carotenoids were calculated according to Lichtenthaler as described in [20]. All extractions and measurements were repeated three times.
2.3. Oxidative Stress Assessment
2.3.1. Localization of Superoxide Anion (O2•−) Production and Hydrogen Peroxide (H2O2) Accumulation
Localization of O2•− was determined using the nitro blue tetrazolium (NBT, Sigma-Aldrich, St. Louis, MO, USA) reduction staining method, while H2O2 accumulation was evaluated using the 3,3’-diaminobenzidine (DAB, Sigma-Aldrich, St. Louis, MO, USA) staining method [19]. Control plants and plants inoculated with appropriate bacterial strains and grown in IAA− or IAA+ medium were collected and immersed for 2 h in 0.2% solution of NBT prepared in 50 mM sodium phosphate buffer (pH 7.5) and 1.25 mg/mL DAB-HCl (pH 3.8). The samples were wrapped in aluminum foil and incubated at room temperature. The localization of endogenous O2•− in duckweeds was visualized as blue formazan precipitates due to the reduction of pale yellow NBT by O2•−. At the sites of endogenous H2O2 accumulation in duckweed tissue, DAB forms a deep brown polymerization product in the presence of peroxidases. As a staining protocol control, some samples were incubated in buffer or 10 mM ascorbic acid. Chlorophyll from frond tissue was extracted with acetic acid/glycerol/ethanol (1:1:3; v/v/v) solution for 5 min at 100 °C or until the frond color was removed. The bleached plants were submerged in glycerol/ethanol (1:4; v/v) storing solution before mounting on slides for light microscopy analysis. All samples were visualized by a Leica DMLB 2900 light microscope and the program LAS V4.11.
2.3.2. Lipid Peroxidation and Hydrogen Peroxide (H2O2) Content
To estimate lipid peroxidation, MDA (malondialdehyde) was detected and measured according to the procedure described by Health and Packer [21]. Briefly, 0.1 g of plant material was ground in liquid nitrogen, and the fine ice powder was dissolved in 1 mL of 0.1% TCA. The extract was centrifuged at 10,000× g for 15 min, at 4 °C. The supernatant was then mixed with 0.5% TBA and 20% TCA. The mixture was boiled at 95 °C for 30 min and then quickly cooled on ice. The absorbance was read spectrophotometrically at 532 nm and 600 nm wavelength. Hydrogen peroxide (H2O2) was quantified according to procedure described in [22]. Briefly, the same plant extract in 0.1% TCA used for MDA detection was mixed with potassium iodide reagent in 10 mM potassium phosphate buffer, pH 7.0, and 1 M potassium iodide. The absorbance of the resulting complex was measured spectrophotometrically at 390 nm. Spectrophotometric readings were performed on a Bio Tek Synergy H1 microplate reader, Agilent Technologies, Santa Clara, CA, USA. Concentrations of MDA and H2O2 were calculated as previously described [23].
2.3.3. Analysis of Antioxidant Enzyme Activity
To assess the activity of antioxidant enzymes CAT, POX and SOD, proteins were extracted as previously described [24]. Briefly, plant tissue was ground in liquid nitrogen with mortar and pestle, until fine powder was formed. Protein extraction buffer (50 mM Tris pH 8 and 1 mM EDTA with 30% glycerol and 1.5% PVPP) was added to the powdered tissue (3:1; v/w), with 10 mM DTT (dithiothreitol) and 1 mM PMSF (phenylmethylsulfonyl fluoride). The extracts were centrifuged at 10,000× g for 15 min, at 4 °C. The supernatant was kept in aliquots, at −20 °C, for further analyses. Peroxidase (POX, EC 1.11.1.7) activity was measured as described previously by [20]. Catalase (CAT, EC 1.11.1.6) activity was measured according to a modified protocol by Aebi (1984) with modifications described by Milošević et al. [24]. Superoxide dismutase (SOD, EC 1.15.1.1) activity was measured according to [25], with modifications described by [26]. Absorbances of POX and CAT were read spectrophotometrically. Absorbances for SOD were measured with a Bio Tek Synergy H1 microplate reader, Agilent Technologies, Santa Clara, CA, USA.
2.4. Statistical Analysis
Each experiment was performed in three biological samples (three flasks, each containing 150 ± 20 duckweeds at the onset of the experiment). In addition, the measurements for each sample were performed three times. The results were expressed as the mean ± standard error. Numerical data were analyzed by ANOVA, followed by Duncan’s test to separate the means at a confidence level of p < 0.05. Statistical analysis was performed using STATISTICA software, version 8. Graphs were generated in GraphPad Prism, version 8 (StatSoft, Hamburg, Germany).
3. Results and Discussion
3.1. Growth Parameters of Duckweeds and Surface Density of Pseudomonas Bacteria
The effect of bacteria (P. oryzihabitans D1-104/3, P. putida A3-104/5, P. gessardii C31-106/3 and P. yamanorum C44-104/1) in the absence (IAA−) or presence of IAA (IAA+) on duckweed growth was evaluated by light microscopy, and representative micrographs were selected (Figure 1). Budding daughter fronds (DFs) are clearly visible in all treatments, still attached to the mother frond (MF) (Figure 1a,b,d,e). Alive fronds are intensely green, without any visible macroscopic changes caused by bacteria (Figure 1b–e,g–j) or IAA (Figure 1b,g). Bleached representative duckweed specimens (Figure 1f–j) show transparent roots and fronds, except in duckweeds inoculated with P. oryzihabitans D1-104/3 and grown in presence of IAA, where the nodes appeared dark and non-transparent (Figure 1g), indicating possible morphological alterations. Structural changes in hosts induced by endophytes are not uncommon; the most investigated are the changes induced by Rhizobium. Since endophytic P. oryzihabitans strains are common [27,28], it is possible that the observed change is a response to an endophytic colonization. The addition of IAA apparently enhanced this effect on the nodes of duckweeds inoculated with P. oryzihabitans D1-104/3, possibly due to stimulatory effect on bacterial growth. In our previous study, we determined that P. oryzihabitans D1-104/3, as well as other strains from this study, had the ability to use IAA as the sole carbon source [27,28]. It is possible that bacteria preferred duckweed nodes as source of nutrients, whilst stimulation from exogenous IAA and the endophytic lifestyle of this strain caused the observed difference in morphology compared to other growth conditions (IAA− and IAA+ with other bacterial strains). However, additional analyses are needed to monitor colonization of duckweeds and potential structural changes caused by bacteria (e.g., scanning electron microscopy) or specific staining for detection of endophytes (e.g., propidium iodide staining). Finally, the complex lace-like organization of frond aerenchyma with air-filled gaps is clearly visible and well-defined, especially in bleached preparations (Figure 1f–j).
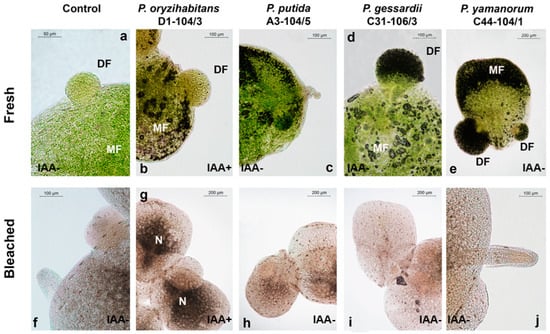
Figure 1.
Representative micrographs of fresh, unbleached (a–e) and bleached (f–j) duckweeds co-cultivated with P. oryzihabitans D1-104/3 (b,g), P. putida A3-104/5 (c,h), P. gessardii C31-106/3 (d,i) and P. yamanorum C44-104/1 (e,j). IAA− indicates an IAA− medium; IAA+ is an IAA+ medium. DF—daughter frond; MF—mother frond; N—node. (b,g)—duckweeds grown in medium with IAA (IAA+) and with P. oryzihabitans D1-104/3.
To assess the macroscopic effects of four Pseudomonas bacteria on duckweeds, the fresh weight of duckweeds was measured. The fresh weight (biomass) increased over time in all treatments and was not affected by IAA (Figure 2a). After 7 days, P. gessardii C31-106/3 significantly increased the fresh weight of duckweeds compared to other samples. There are multiple possible mechanisms by which P. gessardii C31-106/3 might have increased the growth of duckweeds [8]; in our previous study, this strain accelerated the doubling of duckweeds in an IAA+ medium and was able to produce as well as utilize IAA as the sole carbon source [17]. Cell division and growth of duckweeds are strongly associated with IAA: it is possible that P. gessardii C31-106/3 stimulates these processes by regulating IAA levels more efficiently than other strains. However, exact mechanisms should be determined in future characterization studies of its PGPB properties.
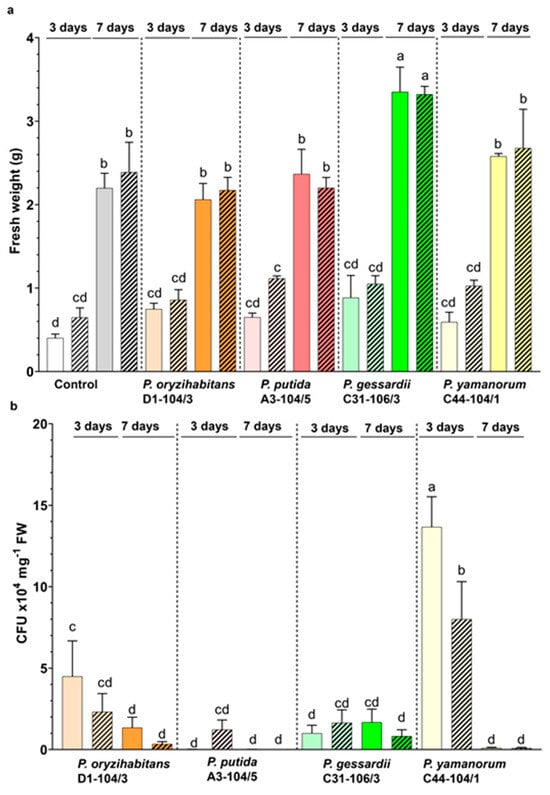
Figure 2.
Growth parameters of duckweeds and bacteria. (a) Fresh weight of duckweeds in IAA− (nonpatterned) and IAA+ (patterned) medium. (b) CFU numbers of P. oryzihabitans D1-104/3, P. putida A3-104/5, P. gessardii C31-106/3 and P. yamanorum C44-104/1 on the surface of duckweeds grown in IAA− (nonpatterned) and IAA+ (patterned) medium. White and grey: surface-sterilized (control) duckweeds. Orange: duckweeds inoculated with P. oryzihabitans D1-104/3; pink: P. putida A3-104/5; green: P. gessardii C31-106/3; yellow: P. yamanorum C44-104/1. Bars represent the mean of 3 replicates, with standard errors. At least one same letter signifies no statistical difference (ANOVA, p < 0.05).
In parallel, CFU numbers of bacteria on the surface of duckweeds were also estimated (Figure 2b). Colonies of P. yamanorum C44-104/1 had the highest density on day 3 of the experiment. Only P. gessardii C31-106/3 formed a relatively constant, high number of colonies throughout the experiment (Figure 2b). The surface density of bacteria also significantly fluctuated in the study of Ishizawa et al. [6]. The authors also noticed that even at very low surface density, PGPB exerted a positive effect. The fluctuations of surface density in this study could reflect each strain’s ecology, biofilm formation and chemotactic behavior: all strains, except for P. gessardii C31-106/3, exhibited chemotactic activity, and all formed biofilms in vitro [17]. Due to this, it is possible that P. gessardii C31-106/3 attaches to the surface of duckweeds via a biofilm more stable than that of other strains, which could explain its relatively constant and high surface density. This might also explain why P. gessardii C31-106/3 significantly increased duckweed biomass after 7 days compared to other strains: sessile lifestyle and biofilm formation are often correlated with a greater plant-growth-promoting effect [29]. Interestingly, P. putida A3-104/5 was the most chemotactically active [17] and in this study had the lowest surface density, pointing at the potential preference of this bacterial strain for the medium rather than the surface of the plants. In this study, the surface density of bacteria is mostly very high throughout the experiment; however, despite this, there was no negative effect on morphology or physiology of duckweeds. This suggests that duckweeds provide enough nutrients for bacteria even in MS medium without a carbon source and support the hypothesis that duckweeds can exist in an environment abundant with microorganisms [1]. In future studies, smaller inocula should be used to test the PGPB effects of these Pseudomonas strains, and fluorescence microscopy should be employed to simultaneously monitor the presence of bacteria on surface of plants and in a free medium.
3.2. Effects of Pseudomonas Strains and IAA on the Photosynthetic Pigment Content in Duckweeds
To assess the effects of bacteria (P. oryzihabitans D1-104/3, P. putida A3-104/5, P. gessardii C31-106/3 and P. yamanorum C44-104/1) co-cultivation in media with or without IAA on the physiological characteristics of duckweeds, the contents of photosynthetic pigments were estimated (Figure 3). The total chlorophyll (chlorophyll a and b) and carotenoid contents in duckweeds were evaluated at two time points: 3 and 7 days after onset of the experiment (Figure 3).
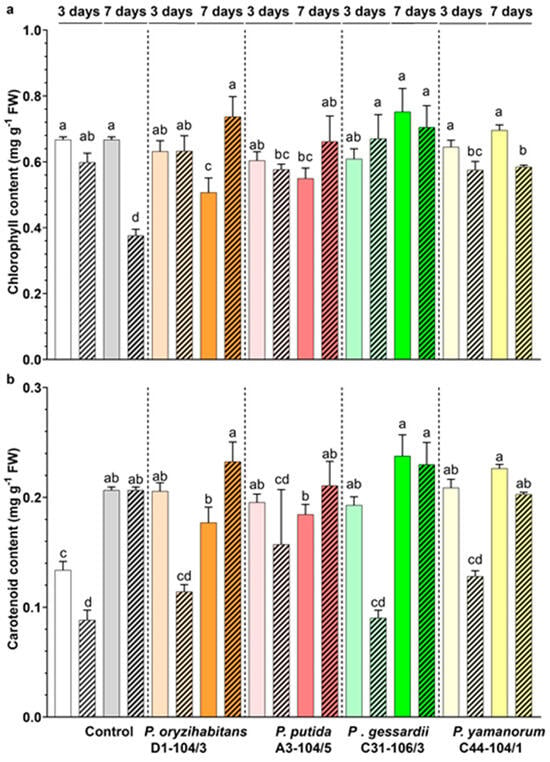
Figure 3.
Effects of Pseudomonas strains and IAA on the photosynthetic pigment content of duckweeds. (a) Chlorophyll content. (b) Carotenoid content in duckweeds co-cultivated with bacteria in IAA− (nonpatterned) and IAA+ MS medium (patterned). White and grey: surface-sterilized (control) duckweeds. Orange: duckweeds inoculated with P. oryzihabitans D1-104/3; pink: P. putida A3-104/5; green: P. gessardii C31-106/3; yellow: P. yamanorum C44-104/1. Bars represent the mean of 3 replicates, with standard errors. At least one same letter signifies no statistical difference (ANOVA, p < 0.05).
In the IAA− medium, total chlorophyll content mostly remained constant throughout the experiment, apart from duckweeds co-cultivated with P. oryzihabitans D1-104/3, in which it declined from day 3 until day 7 (Figure 3a). In the IAA+ medium, total chlorophyll declined only in surface-sterilized duckweeds (Figure 3a, patterned columns). Relative to control, P. putida A3-104/5 and P. yamanorum C44-104/1 had lower chlorophyll content in the IAA+ medium on day 3. In the IAA− medium on day 7, duckweeds with P. oryzihabitans D1-104/3 and P. putida A3-104/5 had higher chlorophyll content than the control. Interestingly, in the IAA+ medium on day 7, all inoculated duckweeds had more chlorophyll than control duckweeds (Figure 3a, patterned columns).
Similarly, the carotenoid content in duckweeds remained largely constant during the experiment, across all treatments (Figure 3b). However, in the IAA− medium, it rose over time in surface-sterilized duckweeds while it remained constant in inoculated samples. It also grew higher in the IAA+ medium in all samples (Figure 3b, patterned columns). Interestingly, on day 3, IAA induced a decrease in carotenoid content by 20–50%, while this effect apparently disappeared until day 7. Moreover, on day 3, bacteria apparently increased total carotenoid content in the IAA− medium (Figure 3, nonpatterned columns).
The content of photosynthetic pigments was mostly unaffected by bacteria, although some Pseudomonas strains had a slightly negative effect on the chlorophyll content of duckweeds (e.g., P. oryzihabitans D1-104/3, P. putida A3-104/5 and P. yamanorum C44-104/1). However, this effect changed over time and apparently depended on IAA as well: in the IAA+ medium, at the end of the experiment, all inoculated duckweeds had more chlorophyll in comparison to control ones. Chlorophyll and carotenoids are indicative of degree of oxidative stress as they also act as antioxidants [30]. However, elevated oxidative stress might become overwhelming to plants’ defense systems, which might induce a drop in photosynthetic pigments. At the beginning of the experiment, the colonization by some bacteria might induce a temporary stress response and a transient reduction in total chlorophyll. Moreover, bacteria showed a tendency to increase total carotenoid content in duckweeds in the first part of the experiment, indicating a protective response by the duckweeds colonized by bacteria. It is possible that IAA exerted a mildly toxic effect on duckweeds, reflected in a decline in pigment content (especially for carotenoids in the first part of the experiment), while bacteria ameliorated this effect over time. The accumulation of IAA leads to the overproduction of ethylene, which might lead to oxidative damage and, in this case, a temporary decline in photosynthetic pigment content [30]. Moreover, the carotenoid content might temporarily shrink because of stress-associated abscisic acid biosynthesis from carotenoids [31]. Although further analyses are needed, these preliminary results hint at the possibility of bacteria ameliorating the negative effects of supraphysiological levels of IAA over time, possibly by activating their own enzymes such as ACC deaminase to counteract ethylene overproduction and subsequently regulate oxidative stress in plants [32].
3.3. Histochemical Assessment of O2•− Production and H2O2 Accumulation
To assess the effects of Pseudomonas bacteria and IAA on superoxide anion (O2•−) production and hydrogen peroxide localization in duckweeds, plants were stained with the NBT (Figure 4) and DAB (Figure 5) staining method.
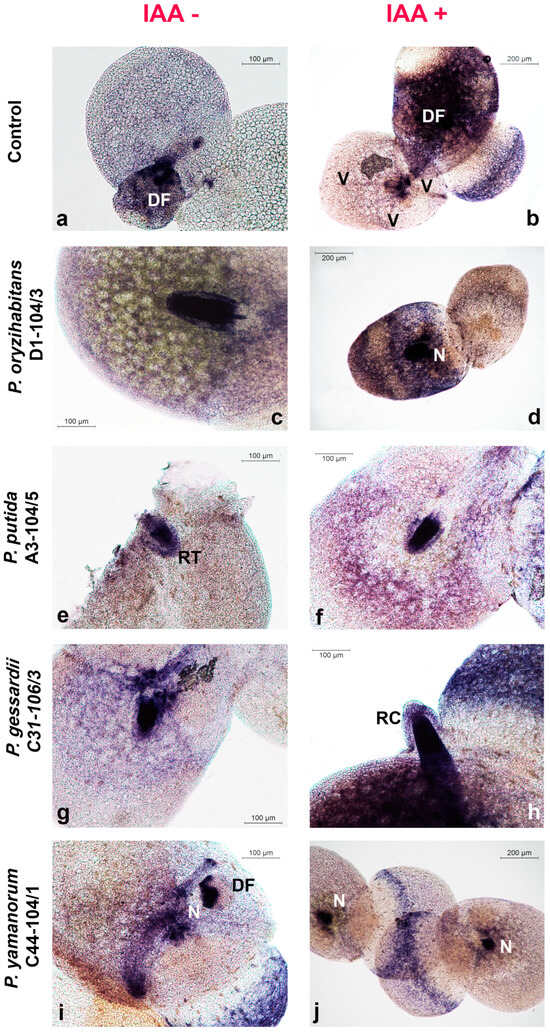
Figure 4.
Superoxide anion (O2•−) production in duckweeds cultivated in IAA− and IAA+ MS medium and inoculated with Pseudomonas strains. Micrographs: (a,b) control; (c,d) P. oryzihabitans D1-104/3; (e,f) P. putida A3-104/5; (g,h) P. gessardii C31-106/3; (i,j) P. yamanorum C44-104/1. DF—daughter frond; N—node; RC—root cap; V—vein. O2•− production localized as blue-colored blue formazan precipitates.
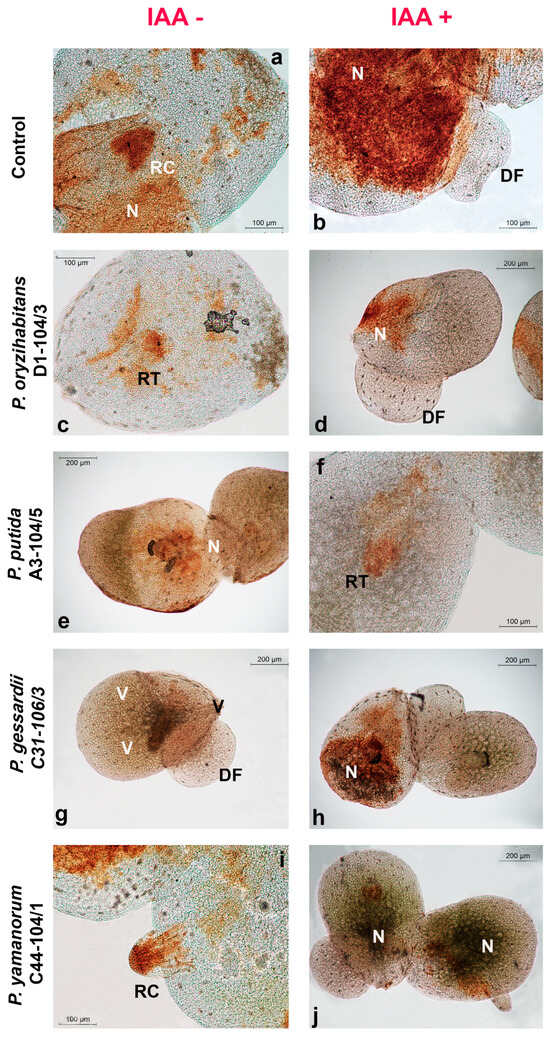
Figure 5.
Localization of hydrogen peroxide accumulation in duckweeds cultivated in IAA and IAA+ MS medium and inoculated with Pseudomonas strains. Micrographs: (a,b) control; (c,d) P. oryzihabitans D1-104/3; (e,f) P. putida A3-104/5; (g,h) P. gessardii C31-106/3; (i,j) P. yamanorum C44-104/1. DF—daughter frond; N—node; RC—root cap; RT—root tip; V—vein. H2O2 accumulation is visualized as deep brown polymerization product of DAB.
Generally, more O2•− was generated in budding daughter fronds (DFs), in root tips and root caps (RTs and RCs) and in the node (N)—the point where the root of the duckweed is attached to the frond (Figure 4). Similar results were observed for H2O2 accumulation: more H2O2 accumulated in root tips, root caps and nodes (Figure 4). Intensive O2•− production was observed in veins (Figure 4b), as well as higher H2O2 accumulation in comparison to the rest of the frond (Figure 5a). The aerenchyma of the fully formed fronds retained less NBT and DAB than roots or daughter fronds (Figure 4 and Figure 5).
Reactive oxygen species (ROS) are constantly produced during growth, photosynthesis and cell respiration under normal conditions. In addition, under abiotic or biotic stress conditions, including colonization by PGPB, ROS are generated intensively [33,34]. Although in this case, it is not possible to discriminate between normally produced ROS and ROS produced in response to bacteria, the pattern of ROS localization in roots, nodes, veins and daughter fronds of duckweeds can hypothetically be also attributed to bacterial preference for these anatomical parts, especially roots since they are highly metabolically active [35,36,37]. Superoxide anions were clearly localized in veins, which might be explained, at least in part, by duckweed–bacteria communication [6], the low reactivity of superoxide anions and the lack of superoxide dismutase and ascorbate in this region enabling O2•− to accumulate [38]. For instance, in veins of duckweeds co-cultivated with P. gessardii C31-106/3, a constantly high number of colonies was recorded, a significant increase in fresh weight was observed and this tissue retained more DAB (see Figure 5g), which indicated higher levels of H2O2 accumulation. Interestingly, P. gessardii C31-106/3 also apparently induced a transient increase in MDA content, indicative of lipid peroxidation with O2•− (see later). Although this increase in MDA content cannot be clearly inferred from micrographs, it is possible that P. gessardii C31-106/3 adhered more strongly to the duckweed surface, which was accompanied by more O2•− production, stimulation of metabolic activity in duckweeds and, consequently, growth promotion. However, further analyses of this strain’s PGP mechanisms are needed to explain its effects on biomass and ROS production. In our previous study, we detected bacteria from the genera Klebsiella, Lelliottia, Serratia and Hafnia concentrating in the midpoint of the duckweed root, which roughly corresponds to elongation zone [35,39]. Although different growth conditions and different staining methods were applied, making a comparison with Pseudomonas strains difficult, this points at tendency of bacteria to organize around metabolically active parts of duckweeds, such as their root elongation zone, which corroborates our hypothesis in this study, i.e., that locations of ROS production and bacterial localization may overlap. Furthermore, as duckweeds almost always reproduce vegetatively, they are usually found in nature in a chain of connected mother fronds and budding daughter fronds, young plants that are identical clones of the mother plant [40]. In this study, we observed many mother fronds with daughter fronds still attached (see Figure 1a,b,d,e). Daughter fronds of L. minor develop from the lateral meristems in the node of duckweeds, the anatomical point where the root is attached to the frond [40]. Superoxide anions (O2•−) are abundantly produced by meristemic tissues, while hydrogen peroxide is produced more by already differentiated tissues [41]. This might explain why daughter fronds stained more with NBT than with DAB. The veins of duckweeds are also very active metabolically, as they enable transport of nutrients through the plant. Moreover, ROS-mediated signaling in response to stress also travels via vascular bundles, which is another possible reason for ROS production localized in veins [6,38,42]. Regarding other visible microscopic features, aerenchyma, the specialized tissue of many water plants that regulates exposure to excess water, was stained faintly due to air-filled gaps between its cells [43]. Both ROS molecules were always concentrated in the roots, especially in root tips and root caps. This localization might also be attributed to the metabolic activity of the apical meristem [35,39]. In conclusion, the fact remains that duckweeds’ root is a habitat for microorganisms even though it is a morphologically simplified and even a redundant organ [5,15,44]. To compare the localization of ROS with bacterial localization, future studies should employ fluorescent staining for live bacterial cells or genetically modified bacteria that express fluorescent proteins to determine their distribution on the root surface of duckweeds and additional histochemical methods for ROS detection. It would be also of scientific interest to see if localized, bacteria-induced ROS production triggers defense mechanisms in duckweeds, since ROS act as signaling molecules that initiate the systemic resistance of plants, the coordinated and global defense response of plants in response to localized infection [45,46,47].
3.4. Effects of Pseudomonas Strains and IAA on the MDA and H2O2 Content in Duckweeds
To assess the effects of different Pseudomonas strains and IAA on the H2O2 and MDA content of duckweeds co-cultivated with four bacteria in media with or without IAA, two selected time points (3 and 7 days after inoculation) were evaluated (Figure 6).
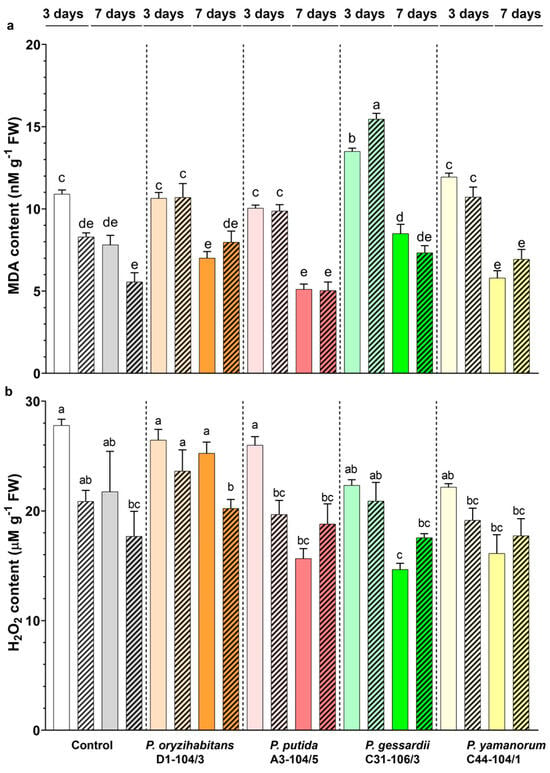
Figure 6.
MDA (a) and H2O2 (b) content of duckweeds cultivated in IAA− (nonpatterned) and IAA+ medium (patterned), with or without bacteria. Control samples are surface-sterilized duckweeds. White and grey: surface-sterilized (control) duckweeds. Orange: duckweeds inoculated with P. oryzihabitans D1-104/3; pink: P. putida A3-104/5; green: P. gessardii C31-106/3; yellow: P. yamanorum C44-104/1. Bars represent the mean of 3 replicates, with standard errors. At least one same letter signifies no statistical difference (ANOVA, p < 0.05).
In all samples grown in the IAA− medium, the MDA content was significantly reduced on day 7 of the experiment, compared to day 3 (Figure 6a). In control duckweeds, grown in the IAA+ medium, the MDA content remained the same from day 3 until day 7, whereas it was reduced in inoculated samples. On day 3, in duckweeds co-cultured with P. gessardii C31-106/3, the MDA content was significantly increased in both types of media, compared to appropriate controls. On day 3, in samples inoculated with other bacterial strains, MDA content was the same as that in control duckweeds in the IAA− medium. In the IAA+ medium, all bacteria increased the MDA content. On day 7, the MDA content was the same in all samples.
In the IAA− medium, the H2O2 content was significantly reduced on day 7 of the experiment in duckweeds inoculated with P. putida A3-104/5 and P. gessardii C31-106/3, compared to day 3 (Figure 4b). In control and in samples inoculated with other bacteria, it remained the same. In the IAA+ medium, in duckweeds inoculated with P. oryzihabitans D1-104/3 the H2O2 content decreased from day 3 to day 7. In other samples, it remained the same. On day 7, compared with the control in the IAA− medium, P. gessardii C31-106/3 had a significantly lower H2O2 content, whereas it was the same in other samples. The addition of IAA had no effect except in duckweeds inoculated with P. oryzihabitans D1-104/3 (day 7) and P. putida A3-104/5 (day 3).
It is evident from our results that duckweeds tolerate all bacteria used in this study since they cause a low level of oxidative stress during cultivation. This is consistent with some of our previous results and studies of other authors [6]. It is obvious that the antioxidant defense system of duckweeds efficiently removed O2•− over time. In this study, we found that the presence of bacteria only had a slightly negative effect if IAA was added to culture media, by increasing O2•− production at the beginning of the co-cultivation. The reason for this observed increase might be due to O2•− being overproduced during the colonization of plants by bacteria and serving as a signaling molecule that is upregulated by IAA [6,38,48]. In general, co-cultivation with bacteria or the exogenous application of IAA did not increase H2O2 levels in duckweeds. Interestingly, exogenous IAA application also did not produce more ROS, although a concentration of 10 mg L−1 of IAA is above physiological concentrations that apparently induced a transient decrease in carotenoid content, as presented in this study, and prolonged doubling times of duckweeds [16]. Moreover, in the presence of P. gessardii C31-106/3, an increase in lipid peroxidation (indicative of O2•− production) was observed on day 3 in both types of media; compared to control (surface-sterilized) duckweeds, the H2O2 content was reduced by the end of the experimental period. As mentioned above, duckweeds inoculated with this strain exhibited stimulated biomass production. In our previous study [17], this strain showed a positive effect on growth and increased the relative growth rate of duckweeds. According to the results of the current study, we can hypothesize that the positive effect of this bacterium on biomass production and relative growth rates of duckweed can be partially attributed to its antioxidative modulation capability. In general, levels of ROS produced in this experiment did not hinder the growth of duckweeds, either in response to bacteria or IAA. Any detrimental effect of interactions with bacteria is outweighed by potential benefits, e.g., more efficient nutrient uptake [49]. This might be an additional explanation for the biomass increase in duckweeds inoculated with P. gessardii C31-106. Finally, some Pseudomonas strains can activate plant defense pathways, leading to increased ROS production [45,47]. This activation could vary among strains and also depends on other factors (in this case, IAA) affecting MDA and H2O2 levels produced by the plants. This effect might account for the observed differences in ROS production between duckweeds inoculated with different bacterial strains used in this study.
3.5. Effects of Pseudomonas Strains and IAA on the Antioxidant Enzyme Activity of Duckweeds
To assess the effects of P. oryzihabitans D1-104/3, P. putida A3-104/5, P. gessardii C31-106/3 and P. yamanorum C44-104/1 and IAA on the activity of antioxidant enzymes (SOD, CAT and POX) of duckweeds co-cultivated with bacteria in media with or without IAA, we evaluated their activity at two time points (3 and 7 days after onset of the experiment, Figure 7). In duckweeds grown in the IAA− medium, 3 days after cultivation, the SOD activity was at the same level in all samples except in duckweeds inoculated with P. putida A3-104/5, in which significantly higher SOD activity was recorded in comparison to other treatments (Figure 7a). We recorded significantly lower levels of SOD activity in duckweeds grown in the IAA+ medium after 3 days of cultivation except for P. gessardii C31-106/3 and P. yamanorum C44-104/1 in comparison to IAA− medium. Seven days after inoculation, a significant decrease in SOD activity was observed in duckweeds grown in the IAA-free medium and co-cultivated with P. putida A3-104/5 and P. yamanorum C44-104/1. In duckweeds grown in the IAA+ medium, after seven days, there was an increase in SOD activity in control plants and a decrease in SOD activity in plants co-cultivated with P. gessardii C31-106/3 and P. yamanorum C44-104/1; in plants co-cultivated with P. oryzihabitans D1-104/3 and P. putida A3-104/5, the SOD activity remained at same level as that at 3 days of cultivation (Figure 7a).
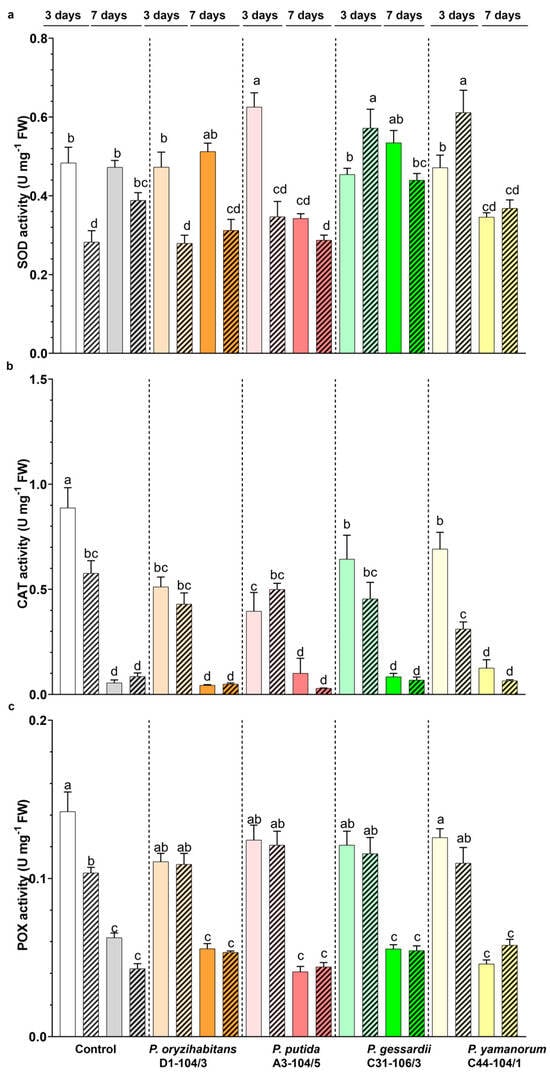
Figure 7.
Effects of Pseudomonas strains and IAA on the activity of antioxidant enzymes SOD (a), CAT (b) and POX (c) of duckweeds co-cultivated with bacteria in IAA− (nonpatterned) or IAA+ MS medium (patterned). White and grey: surface-sterilized (control) duckweeds. Orange: duckweeds inoculated with P. oryzihabitans D1-104/3; pink: P. putida A3-104/5; green: P. gessardii C31-106/3; yellow: P. yamanorum C44-104/1. Bars represent the mean of 3 replicates, with standard errors. At least one same letter signifies no statistical difference (ANOVA, p < 0.05).
The highest level of CAT activity was observed in control plants 3 days after inoculation in the IAA− medium (Figure 7b). In all inoculated duckweeds, a lower level of CAT activity was recorded in comparison to control plants 3 days after cultivation in the IAA− medium. In duckweeds grown in the IAA+ medium, in the same period, a decrease in CAT activity was observed in control and plants co-cultivated with P. yamanorum C44-104/1 compared to IAA− conditions. Apparently, the exogenous application of IAA decreased this enzyme’s activity by 30% in control plants and by 50% in plants inoculated with P. yamanorum C44-104/1. Seven days after cultivation, a significant drop in CAT activity was recorded in all samples (Figure 7b).
Like CAT activity, POX activity was higher in control plants compared to duckweeds co-cultivated with bacteria and grown in the IAA medium (Figure 5c). Similarly, exogenous application of IAA decreased the POX activity level in control plants, while in inoculated plants, POX activity remained at the same level. (Figure 7c). After seven days of cultivation, there were no differences in POX activity between the IAA− and IAA+ samples (Figure 7c).
Antioxidant enzymes act as protectors of plants against oxidative stress, and most of the time, their increased activity indicates an efficient antioxidative response. Although IAA is a plant phytohormone that promotes the growth and development of plants, especially their roots, its effects are dose-dependent: at higher concentrations, IAA induces ethylene and abscisic acid biosynthesis, which is considered as the underlying mechanism of auxin toxicity [30]. In Arabidopsis exposed to oxidative stress, ethylene signaling will induce auxin accumulation in the root and inhibit its growth [31]. Superoxide anions have an important role in the colonization of plants by bacteria [48,50,51,52]. Plants produce superoxide anions in response to recognition of PAMPs (pathogen-associated molecular patterns), molecules characteristic of microorganisms (e.g., bacterial lipopolysaccharide, flagellin, glycans, bacterial nucleic acid). The ability of bacteria to remove superoxide anions and to modulate the host’s SOD is therefore very important during the colonization of the plant. The dynamics of SOD activity can be explained by bacteria regulating the host’s response and by duckweeds recognizing the presence of bacteria. According to our results, SOD activity as a first line of defense was differentially induced depending on bacterial strains as well as exogenous IAA application. For instance, there was a significant increase in SOD activity after 3 days only in plants co-cultivated with P. gessardii C31-106/3 and P. yamanorum C44-104/1 with IAA application. The prolonged application of IAA increased the level of SOD activity in control plants, while SOD activity was significantly decreased (P. gessardii C31-106/3 and P. yamanorum C44-104/1) or remained the same (P. oryzihabitans D1-104/3, P. putida A3-104/5) in co-cultivated plants. In contrast to SOD, CAT and POX activities were significantly decreased after prolonged growth regardless of the presence IAA in the medium and comparable to activity in control plants. Recently, an IAA-producing bacterium Leclercia adecarboxylata was reported to induce an increase in the CAT activity of cucumber exposed to zinc [53]. Similarly, a PGPB Pseudomonas strain ameliorated the toxicity of chromium in tomato and sunflower by increasing CAT activity and decreasing MDA content [54]. Catalases may also act as the first line of defense in some plants; hence, pathogens will tend to decrease CAT activity [24]. According to our study, the activity of CAT was reduced in inoculated duckweeds in the IAA− medium but remained comparable to control when exogenous IAA was applied. However, this decrease apparently was insufficient to stress duckweeds. It is also worth noting that duckweeds are routinely used in toxicity testing: a recent study of bioremediation of cadmium by L. minor and its bacteria showed a similar pattern of decrease for peroxidases and catalases [1,55]. Plants’ peroxidases act not only to remove excess hydrogen peroxide but also as the first line of defense against the pathogens and toxic compounds in many plants [51]. Moreover, various PGPB are reported to increase the activity of antioxidant enzymes and thereby increase the resistance of plants to stress [32]. It is evident from our results that POX enzymes represent the early response of duckweeds to rhizosphere-associated Pseudomonas strains and exogenous IAA. In later stages, peroxidases are less active, probably because they secured tolerable levels of hydrogen peroxide. In addition, the dynamics of enzymatic activation in these types of studies may depend significantly on the clone of duckweed used and experimental conditions [6]. In this study, there were two sources of carbon: exogenous IAA and photosynthetic activity of duckweeds. However, after 3 days, the IAA was undetectable by Salkowski reagent, suggesting that the bulk of IAA was removed by duckweeds and bacteria. This is in accordance with our previous findings, where these Pseudomonas strains grew in minimal medium with 20 mg L−1 of IAA as the sole carbon source and removed IAA at an initial concentration as high as 875 mg L−1 [17]. Ultimately, plants also possess defense mechanisms similar to animal innate immunity: they activate their enzymes and produce ROS differently, depending on the type of microorganism or foreign threat they are exposed to [6,45,46,47]. This might explain the differences in activity rates of enzymes reported in this study. Of three tested antioxidant enzymes, only SOD markedly responded to bacterial presence, making it a good biomarker for duckweed–bacteria interactions. The neutral effects on POX and CAT reflect the non-pathogenic nature of these strains, while a slight reduction in POX and CAT activities in the first part of the experiment in the IAA+ medium confirms dose-dependent toxicity of IAA. Interestingly, P. gessardii C31-106/3 and P. yamanorum C44-104/1 partially ameliorated this effect by increasing SOD activity. In future studies, analyses of differential expression of genes for antioxidant enzymes would elucidate the molecular responses of duckweeds to bacteria and to IAA and clarify their role in plant growth promotion.
4. Conclusions
This study shed more light on oxidative stress and antioxidative responses of duckweeds to four rhizosphere-associated Pseudomonas bacteria and exogenous application of IAA. ROS production and accumulation were localized in the most metabolically active regions of duckweeds: roots, daughter fronds and veins. The presented results also reinforce the hypothesis that plant-growth-promoting bacteria (PGPB) cause a tolerable level of oxidative stress in plants; however, this oxidative stress does not affect biomass production negatively. The PGPB effect of P. gessardii C31-106/3, reflected in a significant increase in biomass production, can be partially explained by the modulation of antioxidant enzymes of duckweeds. Based on the results presented in this study, carotenoids and SOD activity can be the most informative biomarkers of duckweed–bacteria interactions during abiotic stress, at least when the stressor is exogenous IAA at supraphysiological levels. Overall, this study expands the knowledge of aquatic plants’ physiology and responses to stress, which are still insufficiently researched compared to terrestrial plants. These observations may find their future practical application in agriculture and horticulture, by increasing yields and resistance of plants to oxidative stress.
Author Contributions
Conceptualization, O.R.; methodology, T.P., O.R., S.J. and S.M.; validation, T.P., M.Đ., O.R., S.S. and B.U.; formal analysis, O.R.; investigation, T.P., O.R. and S.J.; resources, T.P.; data curation, O.R. and S.S.; writing—original draft preparation, T.P. and O.R.; writing—review and editing, T.P., O.R., S.J., S.M. and B.U.; visualization, M.Đ. and S.J.; supervision, O.R.; project administration, O.R. All authors have read and agreed to the published version of the manuscript.
Funding
This research was financially supported by the Ministry of Science, Technological Development and Innovation of the Republic of Serbia through Grants number 451-03-66/2024-03/200007 and 451-03-65/2024-03/200178.
Data Availability Statement
The data presented in this study are not readily available due to technical/time limitations. Requests to access the datasets should be directed to the corresponding author.
Acknowledgments
The authors would like to thank Martin Raspor for his collegial support during the preparation of this manuscript.
Conflicts of Interest
The author Tatjana Popržen is an employee of MDPI; however, she did not work for this journal (Horticulturae) at the time of submission and publication. The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Abbreviations
| PGPB | plant-growth-promoting bacteria |
| ACC | aminocyclopropane-1-carboxylic acid |
| IAA | indole-3-acetic acid |
| GA | gibberellic acid |
| CAT | catalase |
| POX | peroxidase |
| SOD | superoxide dismutase |
| MDA | malondialdehyde |
| H2O2 | hydrogen peroxide |
| O2•− | superoxide anion |
| ROS | reactive oxygen species |
| TCA | trichloroacetic acid |
| TBA | tiobarbiturate |
| MS medium | Murashige–Skoog medium |
| PVPP | polyvynilpyrrolidone phosphate |
| DTT | dithiothreitol |
| PMSF | phenylmethylsulfonyl fluoride |
| NBT | nitroblue tetrazolium |
| DAB | diaminobenzidine-3,3’-tetrachloride |
References
- Ziegler, P.; Appenroth, K.J.; Sree, K.S. Survival Strategies of Duckweeds, the World’s Smallest Angiosperms. Plants 2023, 12, 2215. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, P.; Sree, K.S.; Appenroth, K.J. Duckweeds for water remediation and toxicity testing. Toxicol. Environ. Chem. 2016, 98, 1127–1154. [Google Scholar] [CrossRef]
- Escobar, C.M.; Escobar, A.C. Duckweed: A tiny aquatic plant with enormous potential for bioregenerative life support systems. In Proceedings of the International Conference on Environmental Systems, Charleston, South Carolina, 16–20 July 2017; pp. 1–9. Available online: http://www.fao.org/ag/againfo/resources/documents/DW/Dw2.htm (accessed on 16 April 2024).
- Xu, J.; Shen, Y.; Zheng, Y.; Smith, G.; Sun, X.S.; Wang, D.; Zhao, Y.; Zhang, W.; Li, Y. Duckweed (Lemnaceae) for potentially nutritious human food: A review. Food Rev. Int. 2023, 39, 3620–3634. [Google Scholar] [CrossRef]
- Acosta, K.; Appenroth, K.J.; Borisjuk, L.; Edelman, M.; Heinig, U.; Jansen, M.A.; Oyama, T.; Pasaribu, B.; Schubert, I.; Sorrels, S.; et al. Return of the Lemnaceae: Duckweed as a model plant system in the genomics and postgenomics era. Plant Cell 2021, 33, 3207–3234. [Google Scholar] [CrossRef] [PubMed]
- Ishizawa, H.; Kuroda, M.; Morikawa, M.; Ike, M. Differential oxidative and antioxidative response of duckweed Lemna minor toward plant growth promoting/inhibiting bacteria. Plant Physiol. Biochem. 2017, 118, 667–673. [Google Scholar] [CrossRef] [PubMed]
- Varga, M.; Horvatić, J.; Čelić, A. Short term exposure of Lemna minor and Lemna gibba to mercury, cadmium and chromium. Cent. Eur. J. Biol. 2013, 8, 1083–1093. [Google Scholar] [CrossRef]
- Ishizawa, H.; Tada, M.; Kuroda, M.; Inoue, D.; Ike, M. Performance of plant growth-promoting bacterium of duckweed under different kinds of abiotic stress factors. Biocatal. Agric. Biotechnol. 2019, 19, 101146. [Google Scholar] [CrossRef]
- Basiglini, E.; Pintore, M.; Forni, C. Effects of treated industrial wastewaters and temperatures on growth and enzymatic activities of duckweed (Lemna minor L.). Ecotoxicol. Environ. Saf. 2018, 153, 54–59. [Google Scholar] [CrossRef]
- Vandenkoornhuyse, P.; Quaiser, A.; Duhamel, M.; Le Van, A.; Dufresne, A. The importance of the microbiome of the plant holobiont. New Phytol. 2015, 206, 1196–1206. [Google Scholar] [CrossRef]
- Ishizawa, H.; Kuroda, M.; Morikawa, M.; Ike, M. Evaluation of environmental bacterial communities as a factor affecting the growth of duckweed Lemna minor. Biotechnol. Biofuels 2017, 10, 62. [Google Scholar] [CrossRef]
- Gómez-Godínez, L.J.; Aguirre-Noyola, J.L.; Martínez-Romero, E.; Arteaga-Garibay, R.I.; Ireta-Moreno, J.; Ruvalcaba-Gómez, J.M. A Look at Plant-Growth-Promoting Bacteria. Plants 2023, 12, 1668. [Google Scholar] [CrossRef] [PubMed]
- Rouphael, Y.; Colla, G. Editorial: Biostimulants in Agriculture. Front. Plant Sci. 2020, 11, 40. [Google Scholar] [CrossRef] [PubMed]
- Nordstedt, N.P.; Jones, M.L. Isolation of Rhizosphere Bacteria That Improve Quality and Water Stress Tolerance in Greenhouse Ornamentals. Front. Plant Sci. 2020, 11, 826. [Google Scholar] [CrossRef] [PubMed]
- Ware, A.; Jones, D.H.; Flis, P.; Chrysanthou, E.; Smith, K.E.; Kümpers, B.M.; Yant, L.; Atkinson, J.A.; Wells, D.M.; Bhosale, R.; et al. Loss of ancestral function in duckweed roots is accompanied by progressive anatomical reduction and a re-distribution of nutrient transporters. Curr. Biol. 2023, 33, 1795–1802.e4. [Google Scholar] [CrossRef] [PubMed]
- Utami, D.; Kawahata, A.; Sugawara, M.; Jog, R.N.; Miwa, K.; Morikawa, M. Effect of exogenous general plant growth regulators on the growth of the Duckweed Lemna minor. Front. Chem. 2018, 6, 251. [Google Scholar] [CrossRef] [PubMed]
- Popržen, T.; Nikoli, I. Characterization of the IAA-Producing and -Degrading Pseudomonas Strains Regulating Growth of the Common Duckweed (Lemna minor L.). Int. J. Mol. Sci. 2023, 24, 17207. [Google Scholar] [CrossRef]
- Bakker, P.A.H.M.; Berendsen, R.L.; Van Pelt, J.A.; Vismans, G.; Yu, K.; Li, E.; Van Bentum, S.; Poppeliers, S.W.M.; Gil, J.J.S.; Zhang, H.; et al. The Soil-Borne Identity and Microbiome-Assisted Agriculture: Looking Back to the Future. Mol. Plant 2020, 13, 1394–1401. [Google Scholar] [CrossRef] [PubMed]
- Albright, M.B.N.; Louca, S.; Winkler, D.E.; Feeser, K.L.; Haig, S.-J.; Whiteson, K.L.; Emerson, J.B.; Dunbar, J. Solutions in microbiome engineering: Prioritizing barriers to organism establishment. ISME J. 2022, 16, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Đurić, M.; Subotić, A.; Prokić, L.; Trifunović-Momčilov, M.; Milošević, S. Alterations in Physiological, Biochemical, and Molecular Responses of Impatiens walleriana to Drought by Methyl Jasmonate Foliar Application. Genes 2023, 14, 1072. [Google Scholar] [CrossRef]
- Lipid Peroxidation Protocol (Health & Packer, 1968). Available online: https://plant-stress.weebly.com/uploads/7/6/3/3/7633398/lipid_peroxidation_protocol.pdf (accessed on 16 April 2024).
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants protective role of exogenous polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Radulović, O.; Stanković, S.; Stanojević, O.; Vujčić, Z.; Dojnov, B.; Trifunović-Momčilov, M.; Marković, M. Antioxidative responses of duckweed (Lemna minor L.) to phenol and rhizosphere-associated bacterial strain hafnia paralvei c32-106/3. Antioxidants 2021, 10, 1719. [Google Scholar] [CrossRef] [PubMed]
- Milošević, S.; Simonović, A.; Cingel, A.; Jevremović, S.; Todorović, S.; Filipović, B.; Subotić, A. Response of antioxidative enzymes to long-term Tomato spotted wilt virus infection and virus elimination by meristem-tip culture in two Impatiens species. Physiol. Mol. Plant Pathol. 2012, 79, 79–88. [Google Scholar] [CrossRef]
- Beyer, W.F.; Fridovich, I. Assaying for superoxide dismutase activity: Some large consequences of minor changes in conditions. Anal. Biochem. 1987, 161, 559–566. [Google Scholar] [CrossRef] [PubMed]
- Antonić, D.; Milošević, S.; Cingel, A.; Lojić, M.; Trifunović-Momčilov, M.; Petrić, M.; Subotić, A.; Simonović, A. Effects of exogenous salicylic acid on Impatiens walleriana L. grown in vitro under polyethylene glycol-imposed drought. S. Afr. J. Bot. 2016, 105, 226–233. [Google Scholar] [CrossRef]
- Rariz, G.; Ferrando, L.; Echegoyen, N.; Scavino, A.F. Antagonism between Azospirillum brasilense Az39 and Pseudomonas oryzihabitans, a seed-borne endophyte, in growing rice plants. Rev. Agronómica Noroeste Argent. 2017, 37, 45–56. [Google Scholar]
- Hardoim, P.R.; Hardoim, C.C.P.; van Overbeek, L.S.; van Elsas, J.D. Dynamics of seed-borne rice endophytes on early plant growth stages. PLoS ONE 2012, 7, e30438. [Google Scholar] [CrossRef] [PubMed]
- Pandit, A.; Adholeya, A.; Cahill, D.; Brau, L.; Kochar, M. Microbial biofilms in nature: Unlocking their potential for agricultural applications. J. Appl. Microbiol. 2020, 129, 199–211. [Google Scholar] [CrossRef] [PubMed]
- Kraft, M.; Kuglitsch, R.; Kwiatkowski, J.; Frank, M.; Grossmann, K. Indole-3-acetic acid and auxin herbicides up-regulate 9-cis-epoxycarotenoid dioxygenase gene expression and abscisic acid accumulation in cleavers (Galium aparine): Interaction with ethylene. J. Exp. Bot. 2007, 58, 1497–1503. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Alonso, M.-M.; Ortiz-García, P.; Moya-Cuevas, J.; Lehmann, T.; Sánchez-Parra, B.; Björk, R.G.; Karim, S.; Amirjani, M.R.; Aronsson, H.; Wilkinson, M.D.; et al. Endogenous indole-3-acetamide levels contribute to the crosstalk between auxin and abscisic acid, and trigger plant stress responses in Arabidopsis. J. Exp. Bot. 2021, 72, 459–475. [Google Scholar] [CrossRef]
- Gamalero, E.; Glick, B.R. Recent Advances in Bacterial Amelioration of Plant Drought and Salt Stress. Biology 2022, 11, 437. [Google Scholar] [CrossRef]
- Berrios, L.; Rentsch, J.D. Linking Reactive Oxygen Species (ROS) to Abiotic and Biotic Feedbacks in Plant Microbiomes: The Dose Makes the Poison. Int. J. Mol. Sci. 2022, 23, 4402. [Google Scholar] [CrossRef]
- Tzipilevich, E.; Russ, D.; Dangl, J.L.; Benfey, P.N. Plant immune system activation is necessary for efficient root colonization by auxin-secreting beneficial bacteria. Cell Host Microbe 2021, 29, 1507–1520.e4. [Google Scholar] [CrossRef] [PubMed]
- López-Farfán, D.; Reyes-Darias, J.A.; Matilla, M.A.; Krell, T. Concentration dependent effect of plant root exudates on the chemosensory systems of Pseudomonas putida KT2440. Front. Microbiol. 2019, 10, 432640. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Xu, Z.; Chen, L.; Xun, W.; Shu, X.; Chen, Y.; Sun, X.; Wang, Z.; Ren, Y.; Shen, Q.; et al. Root colonization by beneficial rhizobacteria. FEMS Microbiol. Rev. 2024, 48, fuad066. [Google Scholar] [CrossRef] [PubMed]
- Schlechter, R.O.; Miebach, M.; Remus-Emsermann, M.N.P. Driving factors of epiphytic bacterial communities: A review. J. Adv. Res. 2019, 19, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Karpinska, B.; Foyer, C.H. Superoxide signalling and antioxidant processing in the plant nucleus. J. Exp. Bot. 2024, erae090. [Google Scholar] [CrossRef] [PubMed]
- Radulović, O.; Stanković, S.; Uzelac, B.; Tadić, V.; Trifunović-Momčilov, M.; Lozo, J.; Marković, M. Phenol removal capacity of the common duckweed (Lemna minor L.) and six phenol-resistant bacterial strains from its rhizosphere: In vitro evaluation at high phenol concentrations. Plants 2020, 9, 599. [Google Scholar] [CrossRef]
- Lemon, G.D.; Posluszny, U. Comparative shoot development and evolution in the Lemnaceae. Int. J. Plant Sci. 2000, 161, 733–748. [Google Scholar] [CrossRef]
- Huang, H.; Ullah, F.; Zhou, D.X.; Yi, M.; Zhao, Y. Mechanisms of ROS regulation of plant development and stress responses. Front. Plant Sci. 2019, 10, 800. [Google Scholar] [CrossRef]
- Zandalinas, S.I.; Fichman, Y.; Mittler, R. Vascular bundles mediate systemic reactive oxygen signaling during light stress. Plant Cell 2020, 32, 3425–3435. [Google Scholar] [CrossRef]
- Takahashi, H.; Yamauchi, T.; Colmer, T.D.; Nakazono, M. Aerenchyma formation in plants. Plant Cell Monogr. 2014, 21, 247–265. [Google Scholar] [CrossRef] [PubMed]
- Ishizawa, H.; Kuroda, M.; Inoue, D.; Morikawa, M.; Ike, M. Community dynamics of duckweed-associated bacteria upon inoculation of plant growth-promoting bacteria. FEMS Microbiol. Ecol. 2020, 96, fiaa101. [Google Scholar] [CrossRef] [PubMed]
- De Vleesschauwer, D.; Djavaheri, M.; Bakker, P.A.H.M.; Höfte, M. Pseudomonas fluorescens WCS374r-induced systemic resistance in rice against Magnaporthe oryzae is based on pseudobactin-mediated priming for a salicylic acid-repressible multifaceted defense response. Plant Physiol. 2008, 148, 1996–2012. [Google Scholar] [CrossRef] [PubMed]
- Xing, Z.; Wu, X.; Zhao, J.; Zhao, X.; Zhu, X.; Wang, Y.; Fan, H.; Chen, L.; Liu, X.; Duan, Y. Isolation and identification of induced systemic resistance determinants from Bacillus simplex Sneb545 against Heterodera glycines. Sci. Rep. 2020, 10, 11586. [Google Scholar] [CrossRef] [PubMed]
- Bakker, P.A.H.M.; Pieterse, C.M.J.; Van Loon, L.C. Induced systemic resistance by fluorescent Pseudomonas spp. Phytopathology 2007, 97, 239–243. [Google Scholar] [CrossRef] [PubMed]
- Alquéres, S.; Meneses, C.; Rouws, L.; Rothballer, M.; Baldani, I.; Schmid, M.; Hartmann, A. The bacterial superoxide dismutase and glutathione reductase are crucial for endophytic colonization of rice roots by Gluconacetobacter diazotrophicus PAL5. Mol. Plant-Microbe Interact. 2013, 26, 937–945. [Google Scholar] [CrossRef] [PubMed]
- Tarkowski, Ł.P.; Signorelli, S.; Considine, M.J.; Montrichard, F. Integration of reactive oxygen species and nutrient signalling to shape root system architecture. Plant Cell Environ. 2023, 46, 379–390. [Google Scholar] [CrossRef]
- Santos, A.D.; Silveira, J.A.; Bonifacio, A.; Rodrigues, A.C.; Figueiredo, M.D. Antioxidant response of cowpea co-inoculated with plant growth-promoting bacteria under salt stress. Braz. J. Microbiol. 2018, 49, 513–521. [Google Scholar] [CrossRef] [PubMed]
- Santos, C.D.; Franco, O.L. Pathogenesis-Related Proteins (PRs) with Enzyme Activity Activating Plant Defense Responses. Plants 2023, 12, 2226. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Peng, Y.; Guo, L.; Li, C. Root colonization of encapsulated klebsiella oxytoca rs-5 on cotton plants and its promoting growth performance under salinity stress. Eur. J. Soil Biol. 2014, 60, 81–87. [Google Scholar] [CrossRef]
- Kang, S.M.; Shahzad, R.; Khan, M.A.; Hasnain, Z.; Lee, K.E.; Park, H.S.; Kim, L.R.; Lee, I.J. Ameliorative effect of indole-3-acetic acid- and siderophore-producing Leclercia adecarboxylata MO1 on cucumber plants under zinc stress. J. Plant Interact. 2021, 16, 30–41. [Google Scholar] [CrossRef]
- Fatima, H.; Ahmed, A. Indole-3-acetic acid synthesizing chromium-resistant bacteria can mitigate chromium toxicity in Helianthus annuus L. Plant Soil Environ. 2020, 66, 216–221. [Google Scholar] [CrossRef]
- Song, R.; Xia, Y.; Zhao, Z.; Yang, X.; Zhang, N. Effects of plant growth regulators on the contents of rutin, hyperoside and quercetin in Hypericum attenuatum Choisy. PLoS ONE 2023, 18, e0285134. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).