

Odour Fingerprints of Black (Tuber mesentericum) and Bianchetto (Tuber borchii) Truffles from Different Areas of the Campania Region

 , ,
, ,  , and
, and 
Abstract

1. Introduction
2. Materials and Methods
2.1. Samples
2.2. Electronic Nose Analysis
2.3. Analysis of Volatile Organic Compounds
2.3.1. Sample Preparation and SPME Extraction
2.3.2. Gas Chromatography/Mass Spectrometry Analysis
2.4. Statistical Analysis
3. Results and Discussion
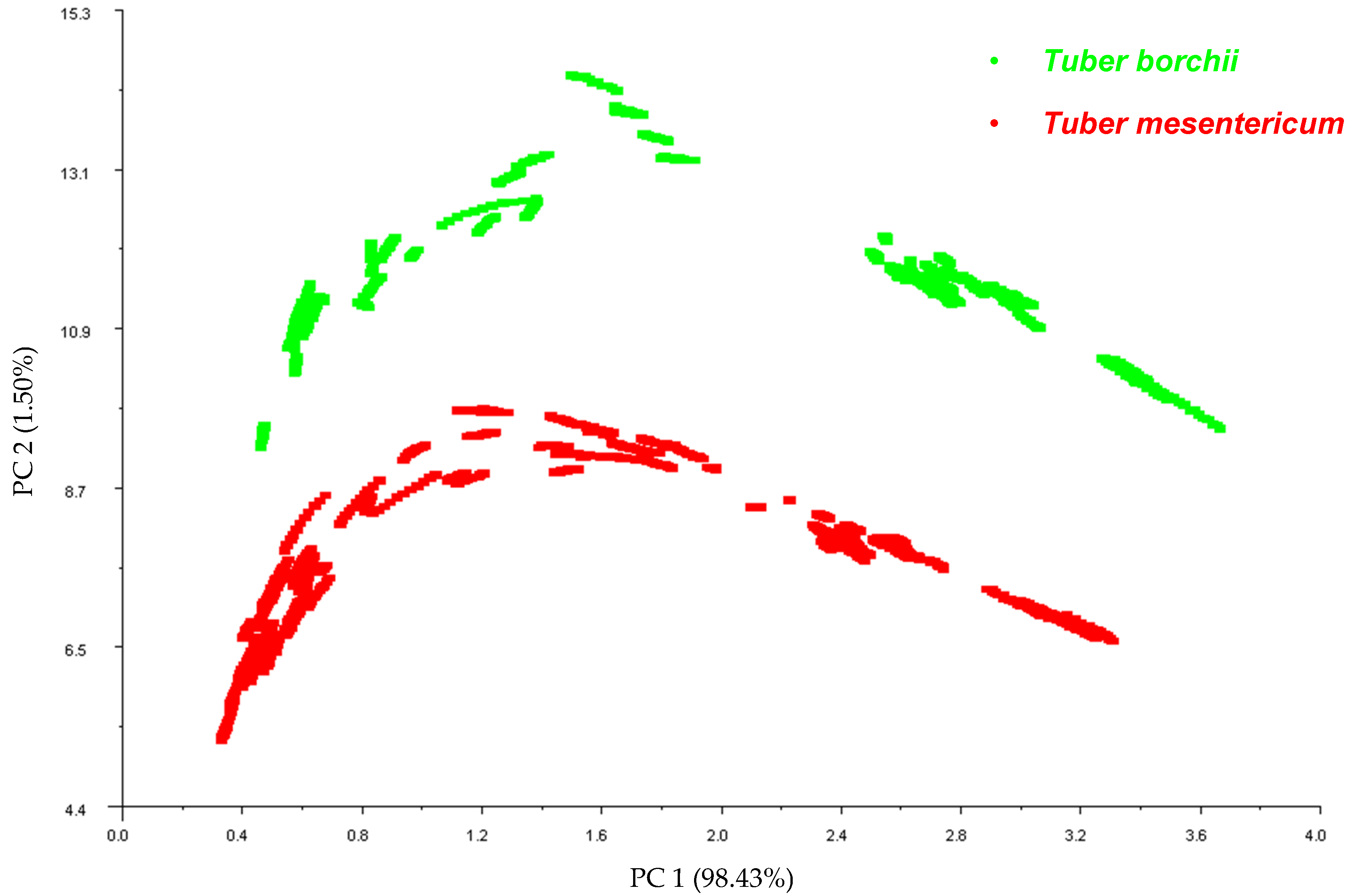
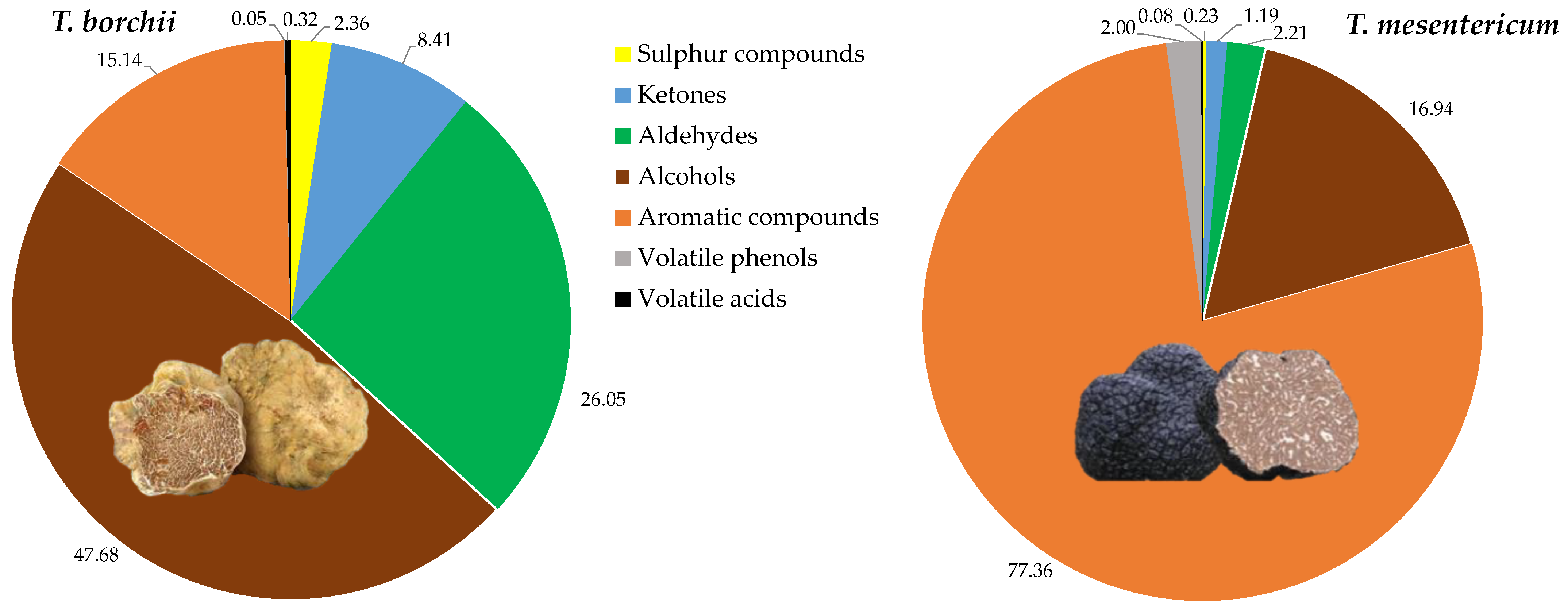
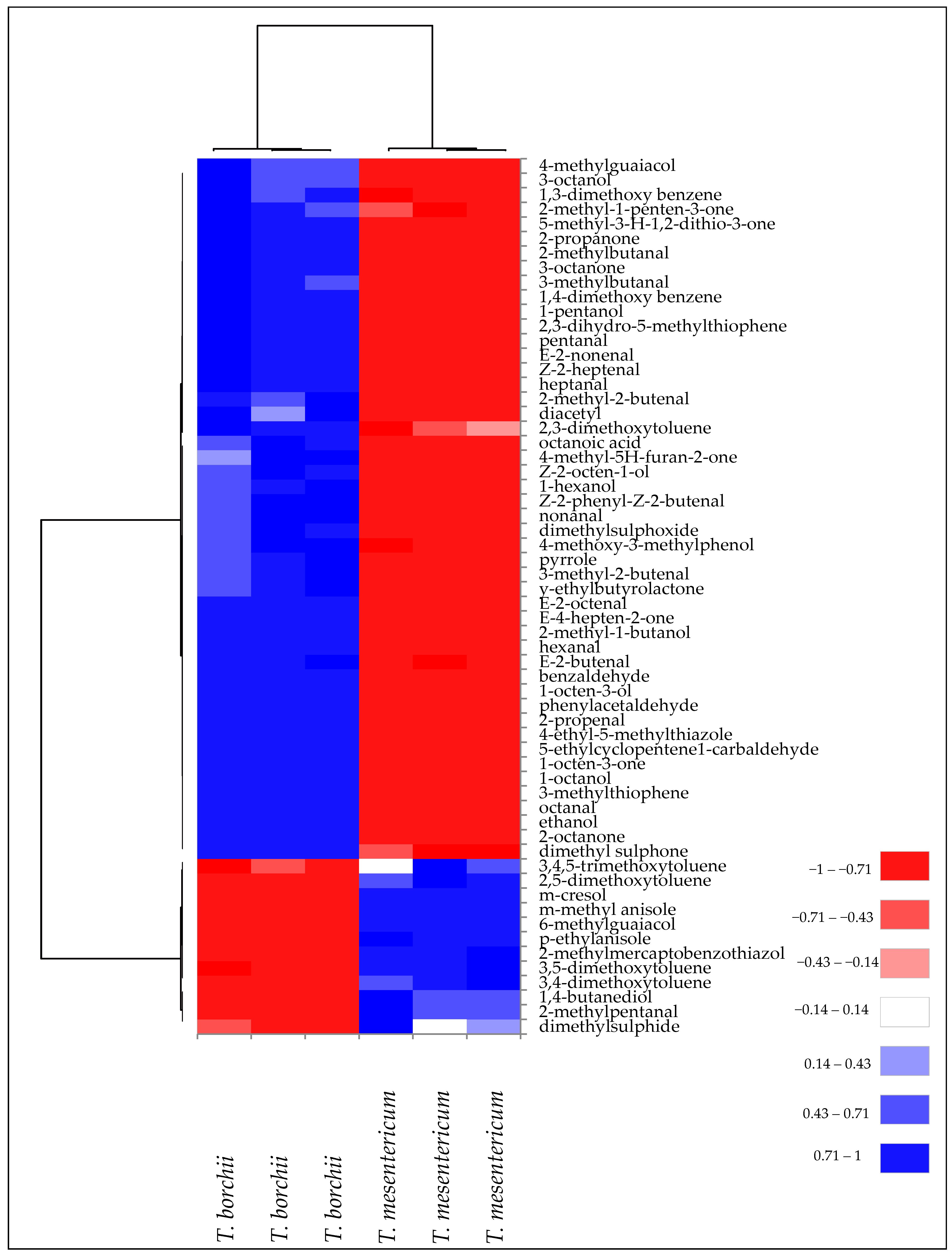
3.1. Specie Variability between Tuber borchii and Tuber mesentericum
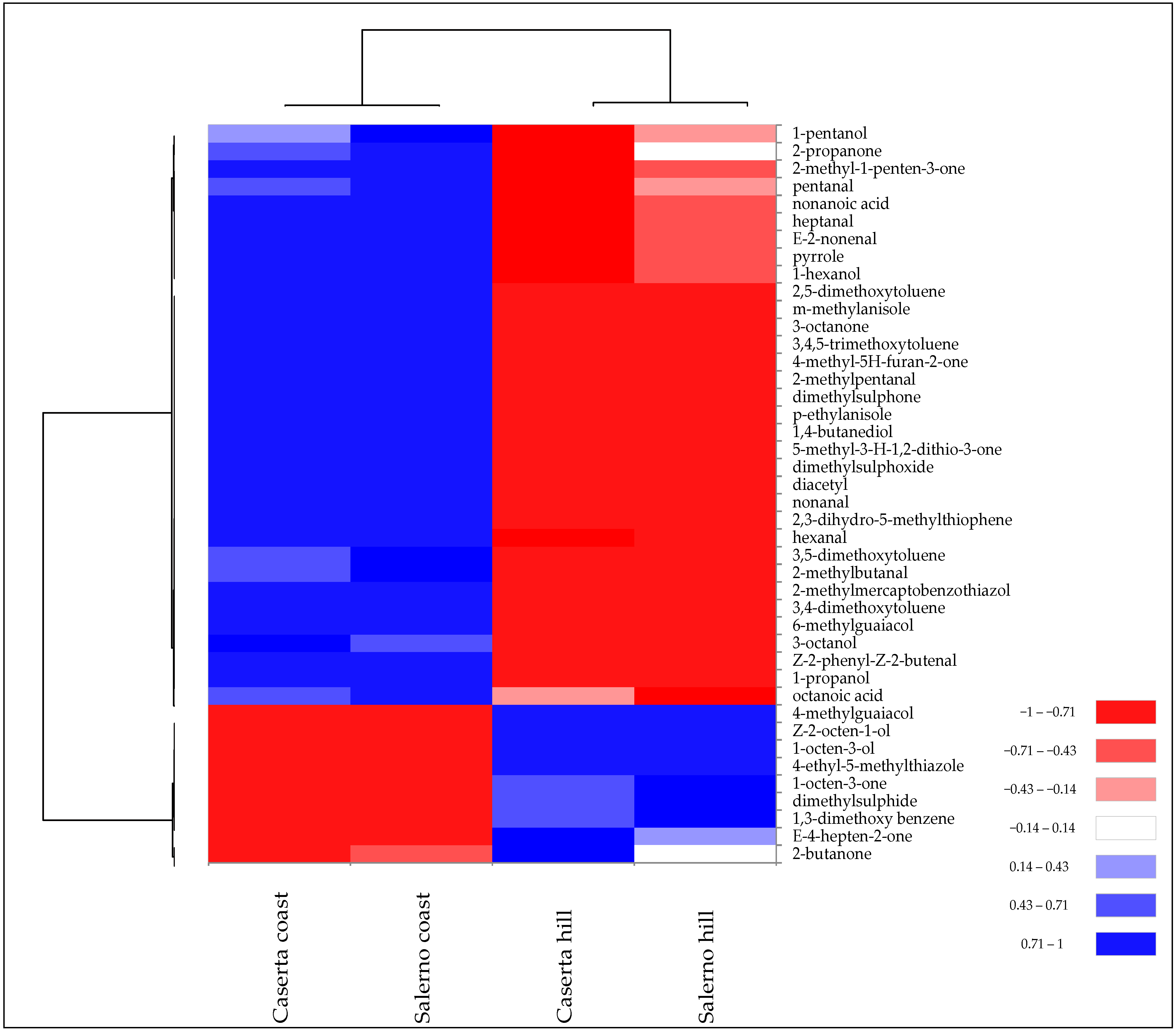
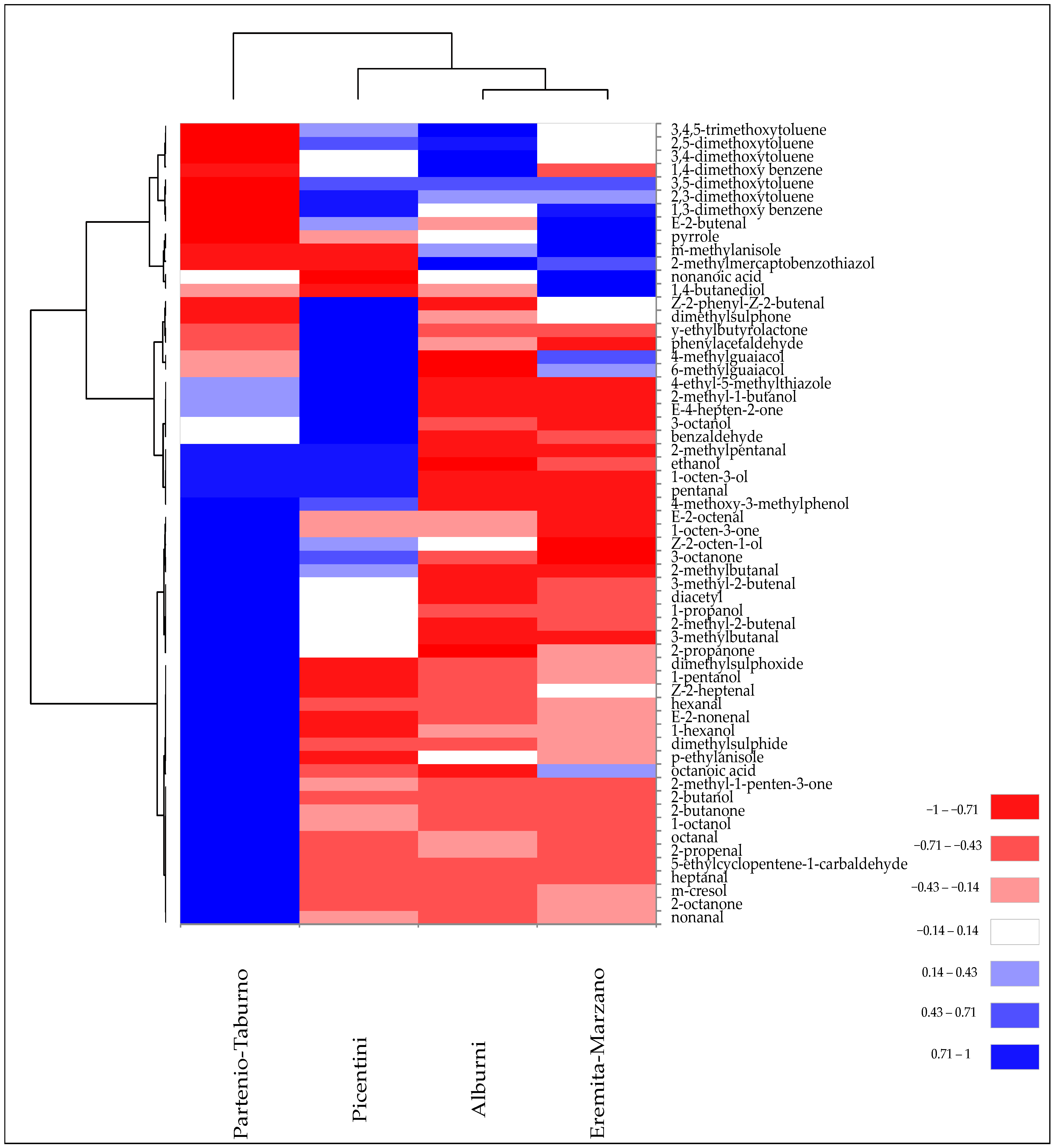
3.2. Geographical Variability of Tuber borchii and Tuber mesentericum
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mello, A.; Murat, C.; Bonfante, P. Truffles: Much more than a prized and local fungal delicacy. FEMS Microbiol. Lett. 2006, 260, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Mustafa, A.M.; Angeloni, S.; Nzekoue, F.K.; Abouelenein, D.; Sagratini, G.; Caprioli, G.; Torregiani, E. An Overview on Truffle Aroma and Main Volatile Compounds. Molecules 2020, 25, 5948. [Google Scholar] [CrossRef] [PubMed]
- Culleré, L.; Ferreira, V.; Chevret, B.; Venturini, M.E.; Sánchez-Gimeno, A.C.; Blanco, D. Characterisation of aroma active compounds in black truffles (Tuber melanosporum) and summer truffles (Tuber aestivum) by gas chromatography–olfactometry. Food Chem. 2010, 122, 300–306. [Google Scholar] [CrossRef]
- Splivallo, R.; Culleré, L. The smell of truffles: From aroma biosynthesis to product quality. In True Truffle (Tuber spp.) in the World: Soil Ecology, Systematics and Biochemistry; Zambonelli, A., Lotti, M., Murat, C., Eds.; Springer International Publishing: Cham, Switzerland, 2016; Volume 47, pp. 393–407. [Google Scholar] [CrossRef]
- March, R.E.; Richards, D.S.; Ryan, R.W. Volatile compounds from six species of truffle–head-space analysis and vapor analysis at high mass resolution. Int. J. Mass Spectrom. 2006, 249, 60–67. [Google Scholar] [CrossRef]
- Kiss, M.; Csóka, M.; Győrfi, J.; Korány, K. Comparison of the fragrance constituents of Tuber aestivum and Tuber brumale gathered in Hungary. J. Appl. Bot. Food. Qual. 2011, 84, 102–110. [Google Scholar]
- Splivallo, R.; Deveau, A.; Valdez, N.; Kirchhoff, N.; Frey-Klett, P.; Karlovsky, P. Bacteria associated with truffle-fruiting bodies contribute to truffle aroma. Environ. Microbiol. 2015, 17, 2647–2660. [Google Scholar] [CrossRef] [PubMed]
- Gioacchini, A.M.; Menotta, M.; Guescini, M.; Saltarelli, R.; Ceccaroli, P.; Amicucci, A.; Barbieri, E.; Giomaro, G.; Stocchi, V. Geographical traceability of Italian white truffle (Tuber magnatum Pico) by the analysis of volatile organic compounds. Rapid Commun. Mass Spectrom. 2008, 22, 3147–3153. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, J.; Qiao, P.; Zhou, D.; Xing, Y.; Chen, J. Monitoring the volatile composition and change in different geographical regions and harvest time of Chinese truffle (Tuber indicum Cooke & Massee). Eur. Food Res. Technol. 2022, 248, 1663–1677. [Google Scholar] [CrossRef]
- Vita, F.; Taiti, C.; Pompeiano, A.; Bazihizina, N.; Lucarotti, V.; Mancuso, S.; Alpi, A. Volatile organic compounds in truffle (Tuber magnatum Pico): Comparison of samples from different regions of Italy and from different seasons. Sci. Rep. 2015, 5, 12629. [Google Scholar] [CrossRef]
- Strojnik, L.; Grebenc, T.; Ogrinc, N. Species and geographic variability in truffle aromas. Food Chem. Toxicol. 2020, 142, 111434. [Google Scholar] [CrossRef]
- Li, Y.; Xing, Y.M.; Murat, C.; Kohler, A.; Zhou, D.Y.; Yu, F.Q.; Chen, J. Transcriptome and metabolome analysis reveals stage-specific metabolite accumulation during maturity of Chinese black truffle Tuber indicum. Plant Physiol. Biochem. 2023, 205, 108158. [Google Scholar] [CrossRef]
- Zhang, B.; Zhang, X.; Yan, L.; Kang, Z.; Tan, H.; Jia, D.; Yang, L.; Ye, L.; Li, X. Different maturities drive proteomic and metabolomic changes in Chinese black truffle. Food Chem. 2021, 342, 128233. [Google Scholar] [CrossRef] [PubMed]
- Dıaz, P.; Ibáñez, E.; Senorans, F.; Reglero, G. Truffle aroma characterization by headspace solid-phase microextraction. J. Chromatog. A 2003, 1017, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Niimi, J.; Deveau, A.; Splivallo, R. Geographical-based variations in white truffle Tuber magnatum aroma is explained by quantitative differences in key volatile compounds. New Phytol. 2021, 230, 1623–1638. [Google Scholar] [CrossRef] [PubMed]
- Bellesia, F.; Pinetti, A.; Tirillini, B.; Bianchi, A. Temperature-dependent evolution of volatile organic compounds in Tuber borchii from Italy. Flavour Fragr. J. 2001, 16, 1–6. [Google Scholar] [CrossRef]
- Culleré, L.; Ferreira, V.; Venturini, M.E.; Marco, P.; Blanco, D. Chemical and sensory effects of the freezing process on the aroma profile of black truffles (Tuber melanosporum). Food Chem. 2013, 136, 518–525. [Google Scholar] [CrossRef] [PubMed]
- Wei, G.; Dan, M.; Zhao, G.; Wang, D. Recent advances in chromatography-mass spectrometry and electronic nose technology in food flavor analysis and detection. Food Chem. 2023, 405, 134814. [Google Scholar] [CrossRef] [PubMed]
- Yakubu, H.G.; Kovacs, Z.; Toth, T.; Bazar, G. Trends in artificial aroma sensing by means of electronic nose technologies to advance dairy production–a review. Crit. Rev. Food Sci. Nutr. 2022, 63, 234–248. [Google Scholar] [CrossRef] [PubMed]
- Gliszczyńska-Świgło, A.; Chmielewski, J. Electronic nose as a tool for monitoring the authenticity of food. A review. Food Anal. Methods 2017, 10, 1800–1816. [Google Scholar] [CrossRef]
- Deshmukh, S.; Bandyopadhyay, R.; Bhattacharyya, N.; Pandey, R.; Jana, A. Application of electronic nose for industrial odors and gaseous emissions measurement and monitoring–an overview. Talanta 2015, 144, 329–340. [Google Scholar] [CrossRef]
- Pacioni, G.; Cerretani, L.; Procida, G.; Cichelli, A. Composition of commercial truffle flavored oils with GC–MS analysis and discrimination with an electronic nose. Food Chem. 2014, 146, 30–35. [Google Scholar] [CrossRef]
- Costa, R.; Fanali, C.; Pennazza, G.; Tedone, L.; Dugo, L.; Santonico, M.; Sciarrone, D.; Cacciola, F.; Cucchiarini, L.; Dachà, M. Screening of volatile compounds composition of white truffle during storage by GCxGC-(FID/MS) and gas sensor array analyses. LWT-Food Sci. Technol. 2015, 60, 905–913. [Google Scholar] [CrossRef]
- Balivo, A.; Cipolletta, S.; Tudisco, R.; Iommelli, P.; Sacchi, R.; Genovese, A. Electronic Nose Analysis to Detect Milk Obtained from Pasture-Raised Goats. Appl. Sci. 2023, 13, 861. [Google Scholar] [CrossRef]
- Schmidberger, P.C.; Schieberle, P. Characterization of the key aroma compounds in white Alba truffle (Tuber magnatum pico) and Burgundy truffle (Tuber uncinatum) by means of the sensomics approach. J. Agric. Food Chem. 2017, 65, 9287–9296. [Google Scholar] [CrossRef]
- Splivallo, R.; Bossi, S.; Maffei, M.; Bonfante, P. Discrimination of truffle fruiting body versus mycelial aromas by stir bar sorptive extraction. Phytochemistry 2007, 68, 2584–2598. [Google Scholar] [CrossRef] [PubMed]
- Czerny, M.; Brueckner, R.; Kirchhoff, E.; Schmitt, R.; Buettner, A. The Influence of Molecular Structure on Odor Qualities and Odor Detection Thresholds of Volatile Alkylated Phenols. Chem. Senses 2011, 36, 539–553. [Google Scholar] [CrossRef]
- Splivallo, R.; Ebeler, S.E. Sulfur volatiles of microbial origin are key contributors to human-sensed truffle aroma. Appl. Microbiol. Biotechnol. 2015, 99, 2583–2592. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Marcone, M.F. The biochemistry and biological properties of the world’s most expensive underground edible mushroom: Truffles. Food Res. Int. 2011, 44, 2567–2581. [Google Scholar] [CrossRef]
- Bertault, G.; Raymond, M.; Berthomieu, A.; Callot, G.; Fernandez, D. Trifling variation in truffles. Nature 1998, 394, 734. [Google Scholar] [CrossRef]
- Lu, B.; Perez-Moreno, J.; Zhang, F.; Rinaldi, A.C.; Yu, F. Aroma profile of two commercial truffle species from Yunnan and Sichuan, China: Inter-and intraspecific variability and shared key compounds. Food Sci. Hum. Wellness 2021, 10, 163–173. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample ID | Truffle Species | No. Fruiting Bodies | Harvesting Site | Harvesting Date | Altitude (m) | Host Tree |
|---|---|---|---|---|---|---|
| Caserta hill | Tuber borchii | 6 | Caserta | February 2023 | 370 | Oak |
| Caserta coast | Tuber borchii | 23 | Caserta | March 2023 | 0 | Aleppo and Stone pine |
| Salerno hill | Tuber borchii | 12 | Salerno | February, March, and April 2023 | 800 | Oak |
| Salerno coast | Tuber borchii | 19 | Salerno | February and March 2023 | 0 | Aleppo and Stone pine |
| Alburni | Tuber mesentericum | 25 | Alburni-Vallo di Diano | February, March, and April 2023 | 1200–1300 | Beech |
| Eremita-Marzano | Tuber mesentericum | 18 | Eremita-Marzano | January, February, and March 2023 | 1000 | Beech |
| Partenio-Taburno | Tuber mesentericum | 39 | Partenio-Taburno | January, February, March, and April 2023 | 900–1100 | Beech |
| Picentini | Tuber mesentericum | 25 | Picentini | February, March, and April 2023 | 800–1500 | Beech |
| Compound | Odour Descriptor | Peak Area (%) | |
|---|---|---|---|
| T. borchii | T. mesentericum | ||
| Sulphur compounds | |||
| dimethylsulphide | Sulphur, cabbage-like, onion | 0.11 ± 0.00 b | 0.21 ± 0.05 a |
| 3-methylthiophene | Onion, savoury, roast, truffle, garlic, butter | 0.51 ± 0.02 | n.d. |
| 2-methyl-4,5-dihydrothiophene | Aged cheese, rubber | 1.57 ± 0.14 | n.d. |
| dimethylsulphoxide | Cheesy, garlic, mushroom | 0.08 ± 0.01 a | 0.01 ± 0.00 b |
| dimethylsulphone | Truffle, onion, garlic | 0.01 ± 0.00 a | 0.01 ± 0.00 a |
| Ketones and lactones | |||
| 2-propanone | Ethereal, fruity | 0.46 ± 0.04 a | 0.08 ± 0.01 b |
| 2-butanone | Ethereal, butterscotch | 0.90 ± 0.17 a | 0.89 ± 0.09 a |
| 2,3-butanedione | Buttery | 0.09 ± 0.02 a | 0.01 ± 0.00 b |
| 2-methyl-1-penten-3-one | 0.11 ± 0.01 a | 0.03 ± 0.01 b | |
| 3-octanone | Mushroom, herbal | 2.90 ± 0.40 a | 0.11 ± 0.02 b |
| 2-octanone | Fruity, mushroom | 0.03 ± 0.00 a | 0.01 ± 0.00 b |
| 1-octen-3-one | Mushroom, musty-mouldy | 1.73 ± 0.05 a | 0.06 ± 0.01 b |
| E-4-hepten-2-one | 0.11 ± 0.00 a | 0.01 ± 0.00 b | |
| 5-methyl-3-H-1,2-dithiol-3-one | 1.86 ± 0.20 | n.d. | |
| γ-ethyl-butyrolactone | Stale, burnt plastic, melon | 0.04 ± 0.01 a | 0.01 ± 0.00 b |
| 4-methyl-5H-furan-2-one | 0.08 ± 0.02 | n.d. | |
| Aldehydes | |||
| 2-propenal | Fruity | 0.02 ± 0.00 a | 0.01 ± 0.00 b |
| 2-methylbutanal | Musty, cocoa, nutty, coffee | 1.78 ± 0.16 a | 0.07 ± 0.01 b |
| 3-methylbutanal | Chocolate, peach, fruity | 3.42 ± 0.38 a | 0.48 ± 0.09 b |
| pentanal | Coffee, nutty, chocolate | 0.32 ± 0.02 a | 0.02 ± 0.00 b |
| E-2-butenal | 0.70 ± 0.03 a | 0.25 ± 0.03 b | |
| hexanal | Grassy, leafy | 8.21 ± 0.14 a | 0.58 ± 0.08 b |
| 2-methyl-2-butenal | Green | 0.43 ± 0.07 a | 0.04 ± 0.01 b |
| 3-methyl-2-butenal | Fruity | 0.06 ± 0.01 a | 0.01 ± 0.00 b |
| 2-methyl pentanal | Ethereal | 0.01 ± 0.00 a | 0.01 ± 0.00 a |
| heptanal | Fruity | 1.64 ± 0.06 a | 0.09 ± 0.02 b |
| octanal | Fruity | 0.18 ± 0.01 a | 0.01 ± 0.00 b |
| Z-2-heptenal | Green, slightly citrus | 0.36 ± 0.02 a | 0.07 ± 0.01 b |
| nonanal | Fruity, floral, rose-orange | 0.34 ± 0.05 a | 0.02 ± 0.00 b |
| 5-ethylcyclopentene-1-carbaldehyde | Grilled lamb | 2.72 ± 0.08 a | 0.24 ± 0.02 b |
| E-2-octenal | Fresh cucumber, leafy | 4.62 ± 0.04 a | 0.44 ± 0.04 b |
| E-2-nonenal | Green | 0.14 ± 0.01 a | 0.03 ± 0.00 b |
| Alcohols | |||
| ethanol | Alcoholic | 0.27 ± 0.01 a | 0.06 ± 0.01 b |
| 2-butanol | Fruity | 0.07 ± 0.00 a | 0.09 ± 0.02 a |
| 1-propanol | Alcoholic | 0.03 ± 0.01 a | 0.02 ± 0.00 a |
| 2-methyl-1-butanol | Fusel, alcoholic, whiskey | 2.49 ± 0.03 a | 0.10 ± 0.02 b |
| 1-pentanol | 0.20 ± 0.02 a | 0.01 ± 0.00 b | |
| 1-hexanol | Ethereal, fusel, fruity | 0.24 ± 0.02 a | 0.05 ± 0.01 b |
| 3-octanol | Earthy, mushroom, woody, citrus | 1.56 ± 0.31 a | 0.06 ± 0.01 b |
| 1-octen-3-ol | Earthy, mushroom | 40.87 ± 1.37 a | 16.27 ± 0.50 b |
| 1-octanol | Waxy | 0.11 ± 0.00 a | 0.01 ± 0.00 b |
| Z-2-octen-1-ol | Floral | 1.51 ± 0.13 a | 0.33 ± 0.07 b |
| Aromatic compounds | |||
| 4-ethyl-5-methylthiazole | Nutty | 0.44 ± 0.03 a | 0.01 ± 0.00 b |
| pyrrole | Nutty | 0.03 ± 0.01 a | 0.01 ± 0.00 b |
| 2-methylmercapto-benzothiazol | 0.02 ± 0.00 b | 0.14 ± 0.03 a | |
| 3-methylanisole | Floral, spicy | 11.96 ± 1.47 b | 66.63 ± 2.38 a |
| p-ethylanisole | Naphthyl | 0.02 ± 0.00 b | 0.54 ± 0.04 a |
| benzaldehyde | Almond | 0.89 ± 0.04 a | 0.16 ± 0.03 b |
| phenylacetaldehyde | Green, floral | 0.60 ± 0.03 a | 0.04 ± 0.01 b |
| 2,3-dimethoxytoluene | 0.18 ± 0.00 a | 0.14 ± 0.04 a | |
| 1,4-dimethoxy benzene | Nut-like | 1.32 ± 0.10 a | 0.04 ± 0.00 b |
| 1,3-dimethoxy benzene | Coconut, hazelnut, earthy | 0.09 ± 0.01 a | 0.04 ± 0.01 b |
| 2,5-dimethoxytoluene | Floral | 0.05 ± 0.01 b | 8.49 ± 2.04 a |
| 3,4-dimethoxytoluene | 0.01 ± 0.00 b | 0.95 ± 0.23 a | |
| 3,5-dimethoxytoluene | 0.15 ± 0.03 b | 0.88 ± 0.14 a | |
| Z-2-phenyl-Z-2-butenal | Musty, cocoa | 0.08 ± 0.01 a | 0.02 ± 0.00 b |
| 3,4,5-trimethoxytoluene | 0.03 ± 0.00 b | 0.05 ± 0.01 a | |
| Volatile phenols | |||
| 4-methylguaiacol | Spicy, smoky | 0.03 ± 0.01 a | 0.03 ± 0.00 a |
| m-cresol | Phenolic, medicinal-leathery, smoky | n.d. | 2.13 ± 0.21 |
| 4-methoxy-3-methylphenol | Phenolic | 0.02 ± 0.00 a | 0.02 ± 0.00 a |
| Volatile acids | |||
| octanoic acid | Musty, pungent | 0.13 ± 0.02 a | 0.03 ± 0.00 b |
| nonanoic acid | Waxy | 0.17 ± 0.09 a | 0.06 ± 0.01 a |
| From\To | Caserta Hill | Salerno Hill | Caserta Coast | Salerno Coast | Total | % Correct | % Correct (Coast vs. Hill) |
|---|---|---|---|---|---|---|---|
| Caserta hill | 4 | 0 | 2 | 0 | 6 | 67% | 67% |
| Salerno hill | 3 | 5 | 0 | 4 | 12 | 42% | 67% |
| Caserta coast | 0 | 2 | 17 | 4 | 23 | 74% | 91% |
| Salerno coast | 2 | 0 | 4 | 13 | 19 | 68% | 89% |
| From\To | Alburni | Eremita-Marzano | Partenio-Taburno | Picentini | Total | % Correct |
|---|---|---|---|---|---|---|
| Alburni | 14 | 1 | 7 | 3 | 25 | 56% |
| Eremita-Marzano | 3 | 10 | 3 | 2 | 18 | 56% |
| Partenio-Taburno | 6 | 4 | 20 | 9 | 39 | 51% |
| Picentini | 5 | 0 | 4 | 16 | 25 | 64% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balivo, A.; De Falco, E.; Branca, L.; Caputo, M.; Sacchi, R.; Genovese, A. Odour Fingerprints of Black (Tuber mesentericum) and Bianchetto (Tuber borchii) Truffles from Different Areas of the Campania Region. Horticulturae 2024, 10, 557. https://doi.org/10.3390/horticulturae10060557
Balivo A, De Falco E, Branca L, Caputo M, Sacchi R, Genovese A. Odour Fingerprints of Black (Tuber mesentericum) and Bianchetto (Tuber borchii) Truffles from Different Areas of the Campania Region. Horticulturae. 2024; 10(6):557. https://doi.org/10.3390/horticulturae10060557
Chicago/Turabian StyleBalivo, Andrea, Enrica De Falco, Luca Branca, Michele Caputo, Raffaele Sacchi, and Alessandro Genovese. 2024. "Odour Fingerprints of Black (Tuber mesentericum) and Bianchetto (Tuber borchii) Truffles from Different Areas of the Campania Region" Horticulturae 10, no. 6: 557. https://doi.org/10.3390/horticulturae10060557
APA StyleBalivo, A., De Falco, E., Branca, L., Caputo, M., Sacchi, R., & Genovese, A. (2024). Odour Fingerprints of Black (Tuber mesentericum) and Bianchetto (Tuber borchii) Truffles from Different Areas of the Campania Region. Horticulturae, 10(6), 557. https://doi.org/10.3390/horticulturae10060557