
Influence of Melatonin Coating on Physiochemical Qualities and Enzymatic Activities in Banana Pericarp under Cold Storage



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
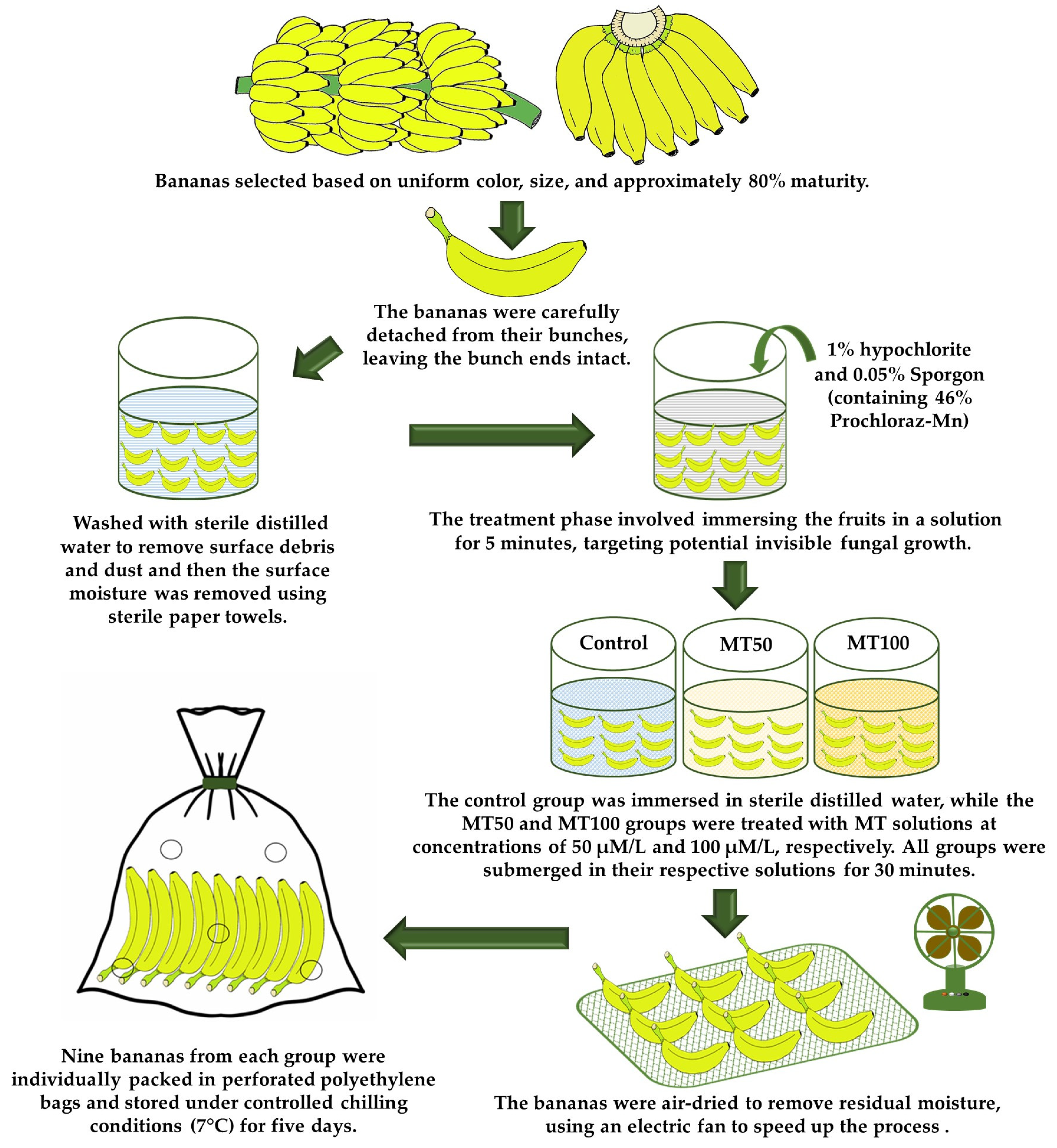
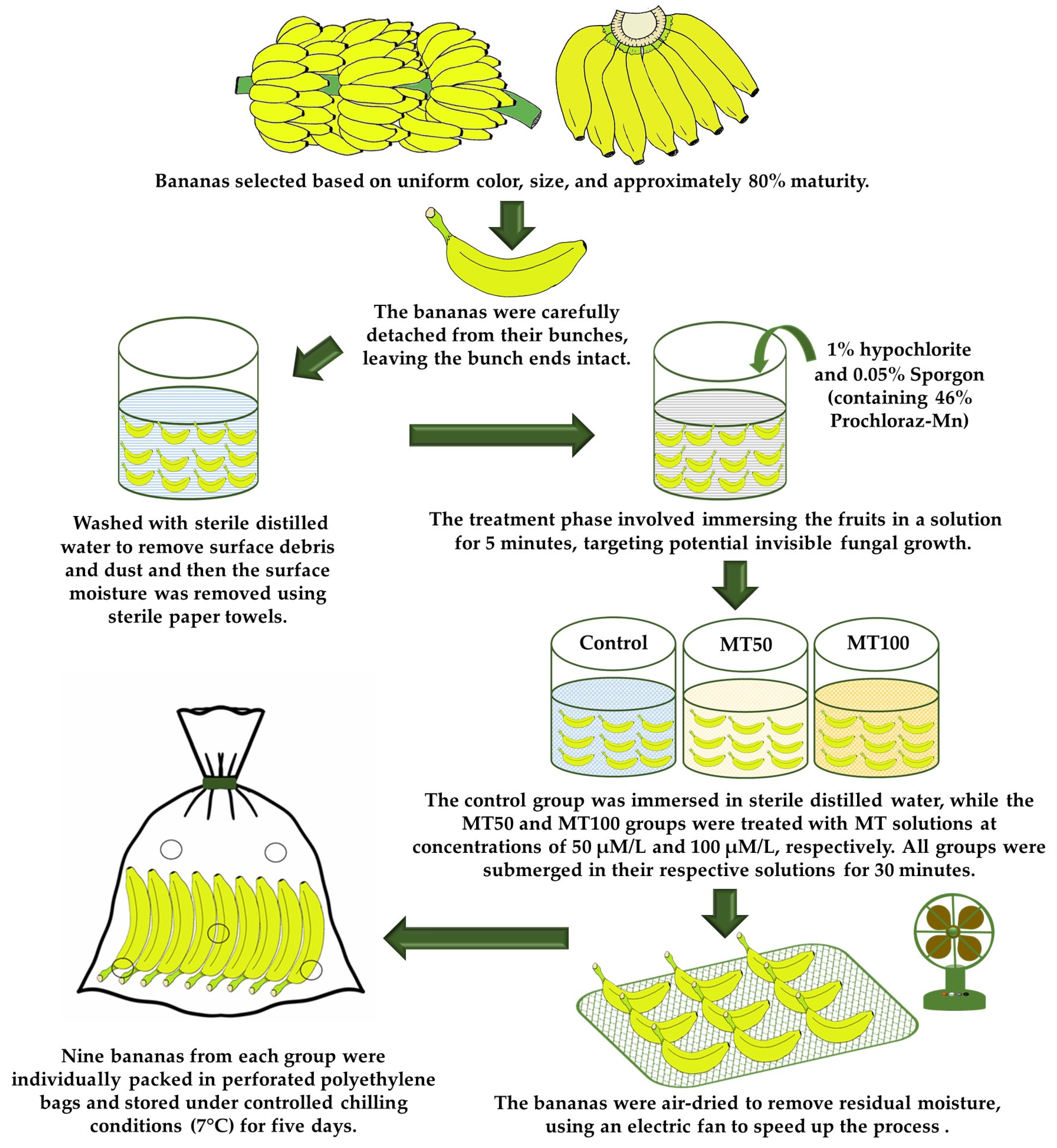
2.2. Plant Materials and Treatments
2.3. Quality Analysis
2.3.1. Determination of CI index
2.3.2. Determination of EL
2.3.3. Determination of MDA
2.3.4. ROS Determination
Determination of O2•− Radicals
Determination of H2O2 Radicals
Determination of •OH Radicals
2.3.5. Determination of GSH
2.3.6. Determination of AA
2.3.7. Determination of TPC
2.3.8. Determination of TCC
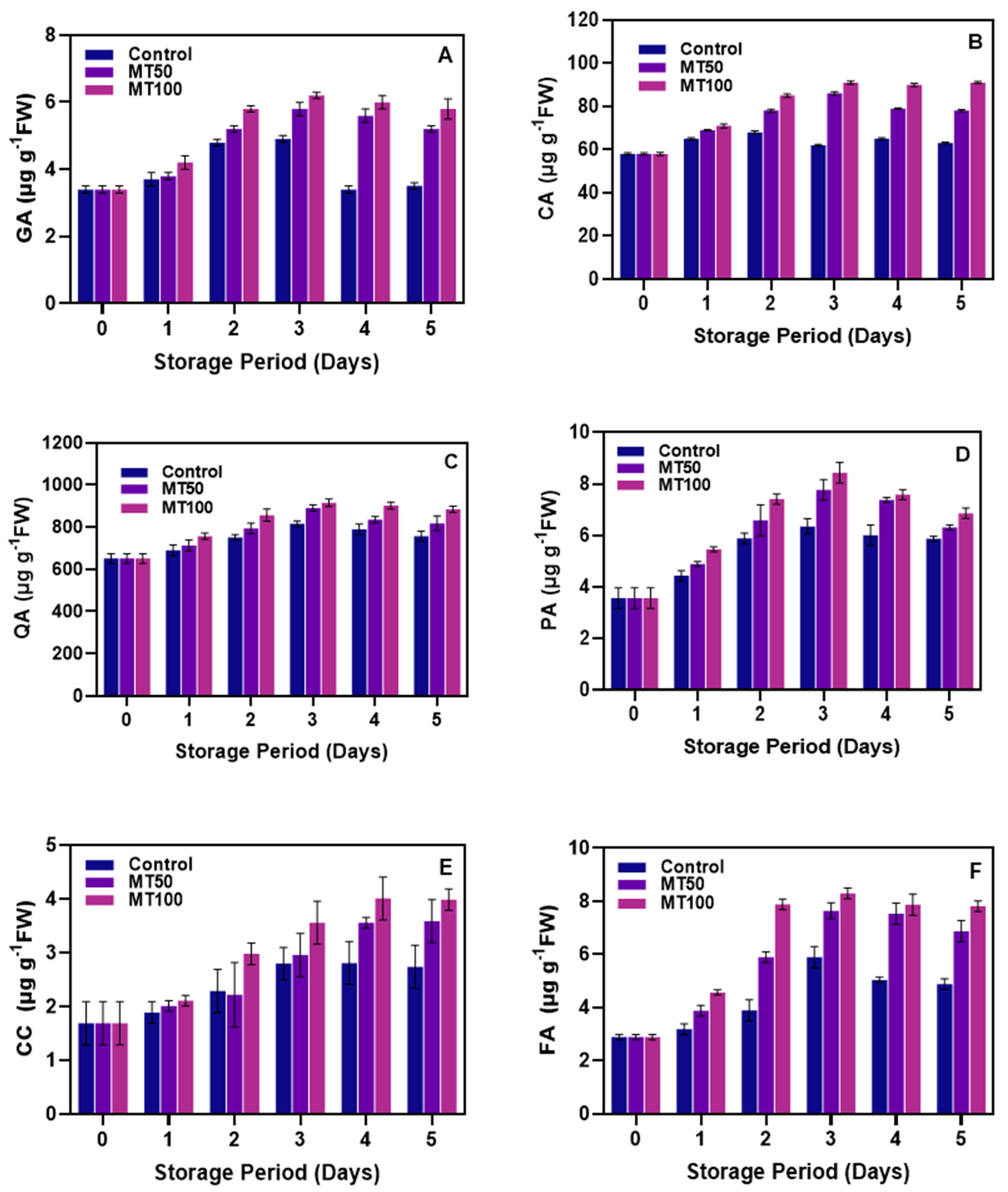
2.3.9. Determination of Phenolic Acids
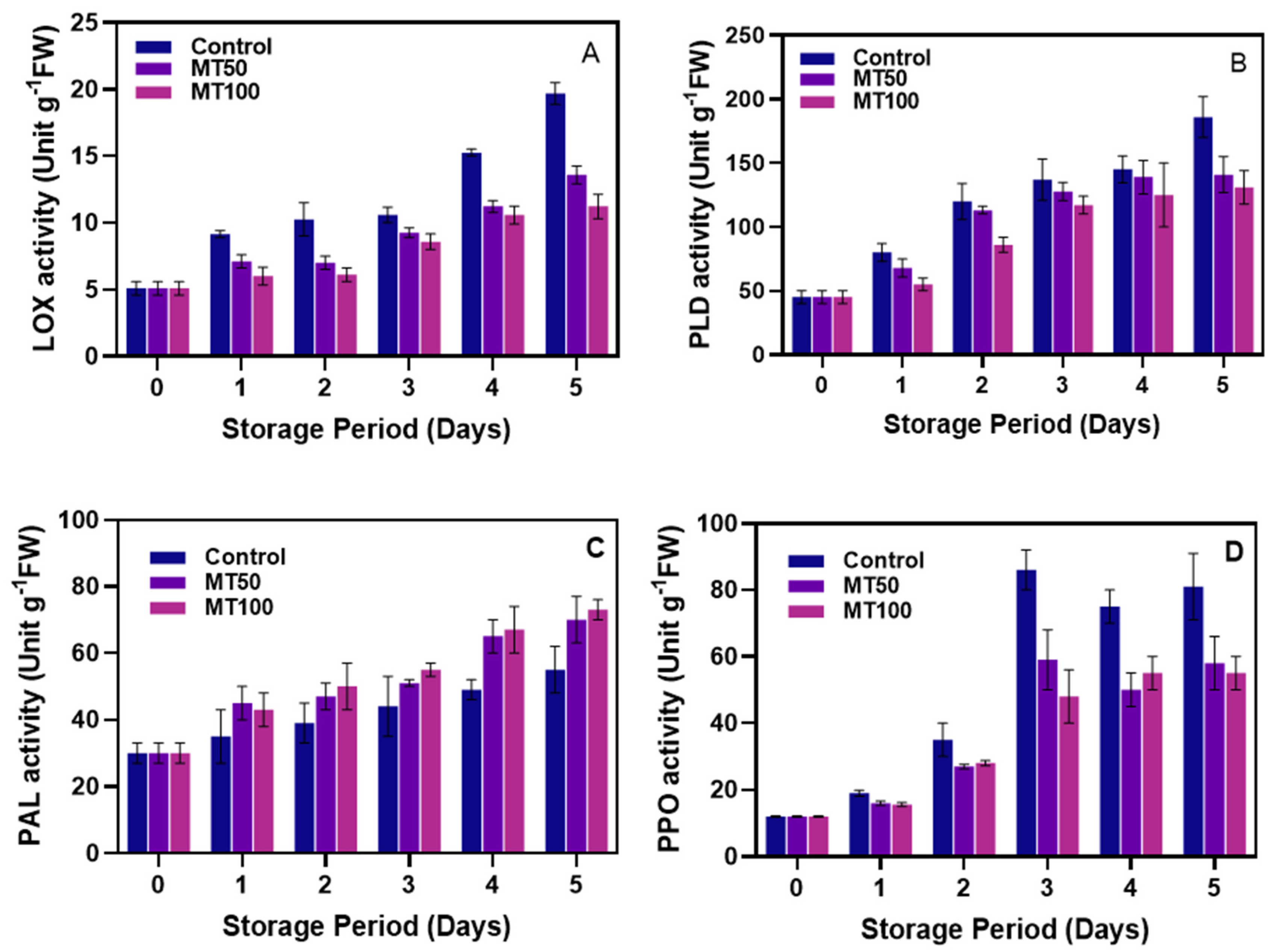
2.3.10. Determination of Enzyme Activities
LOX Activity
PLD Activity
PAL Activity
PPO Activity
2.4. Statistical Analysis
3. Results and Discussion
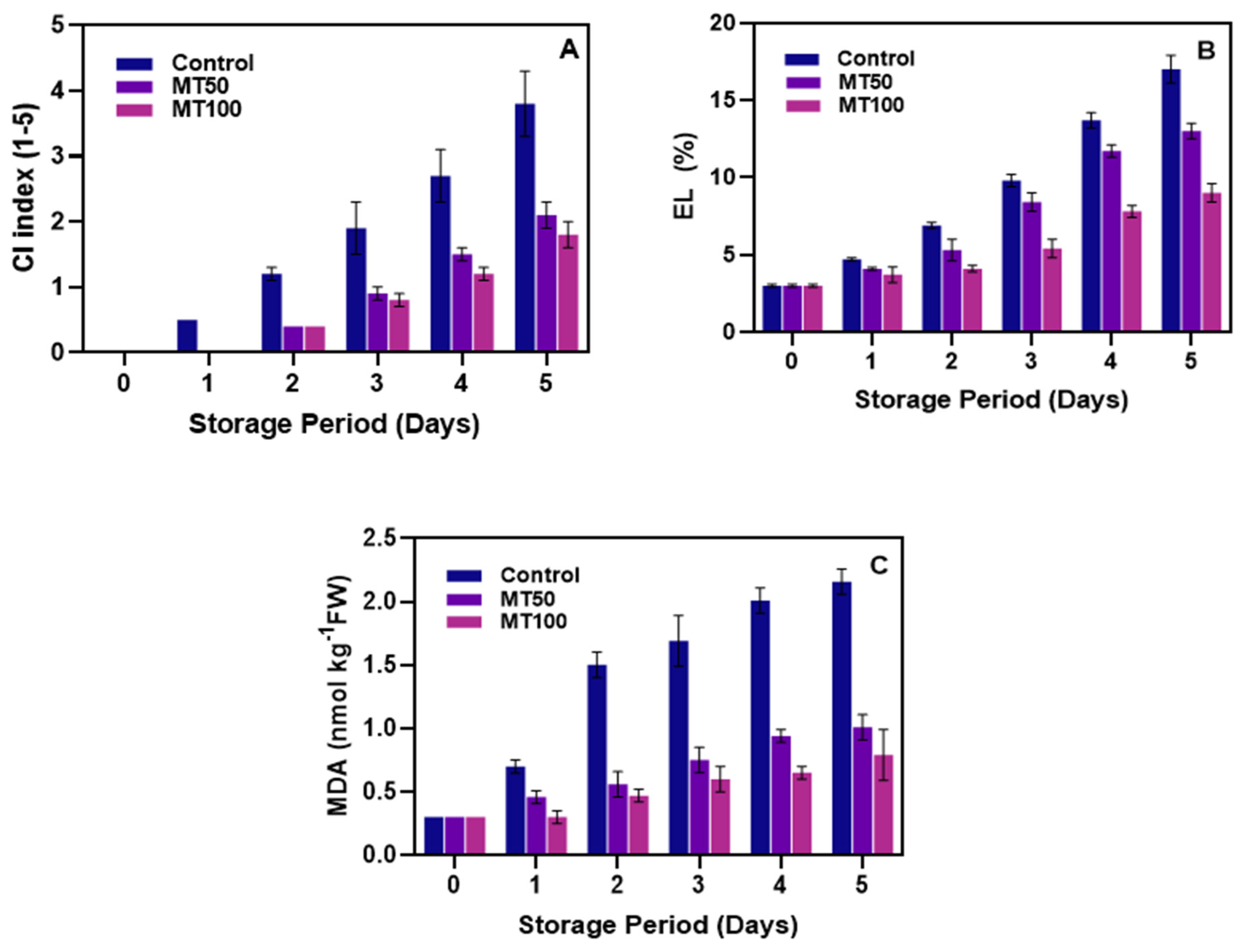
3.1. CI, Membrane Permeability, and MDA Levels
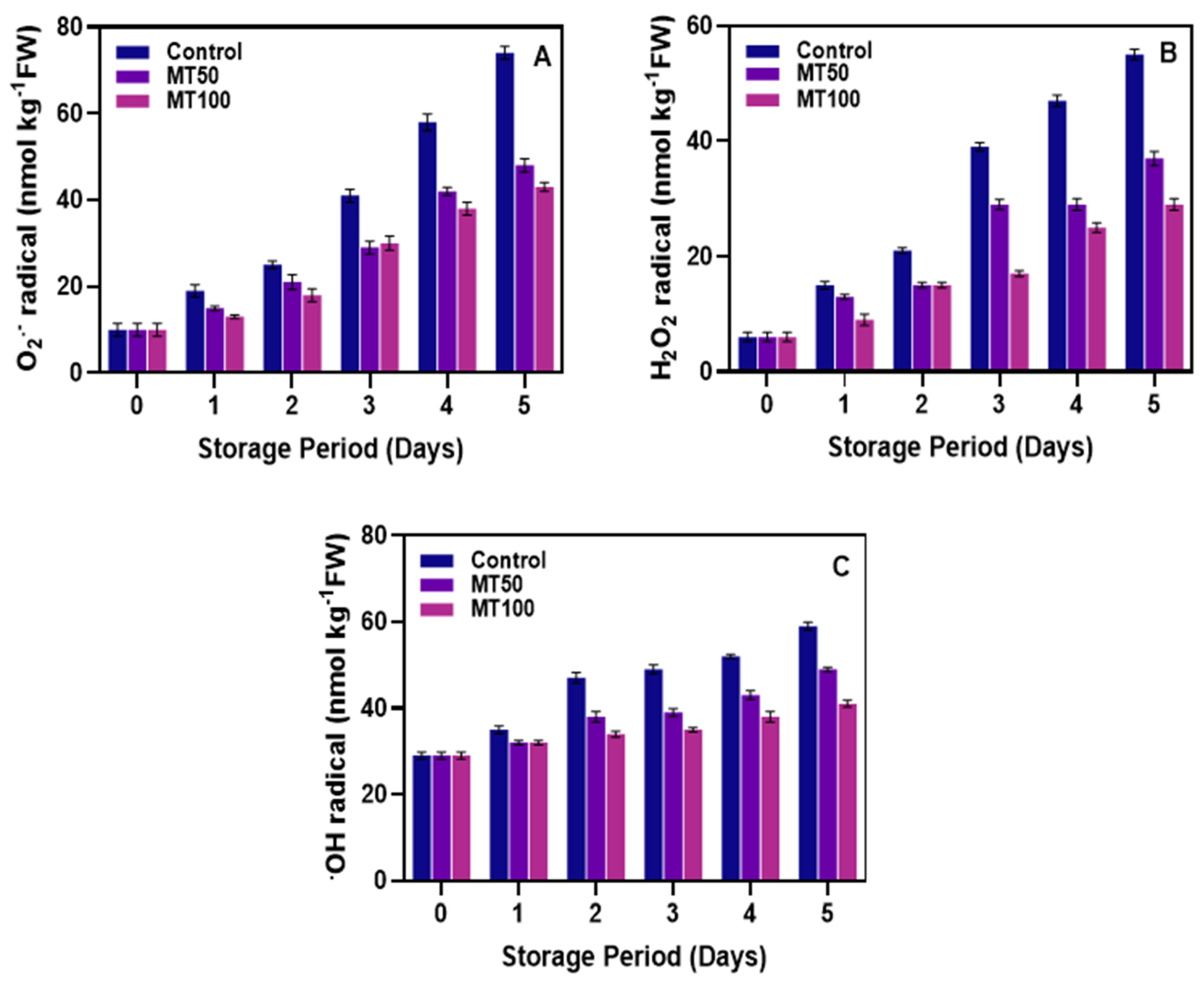
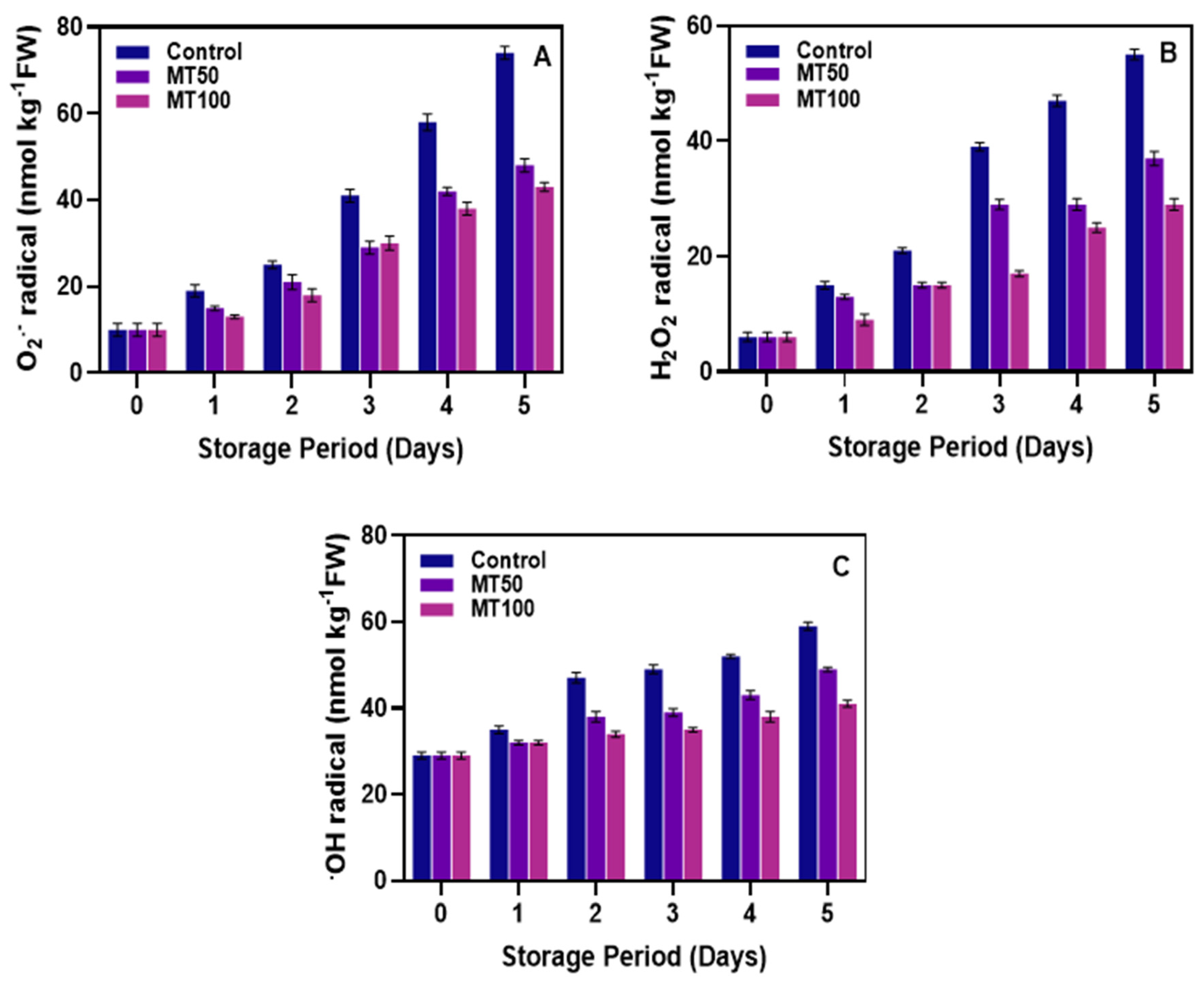
3.2. Production of ROS
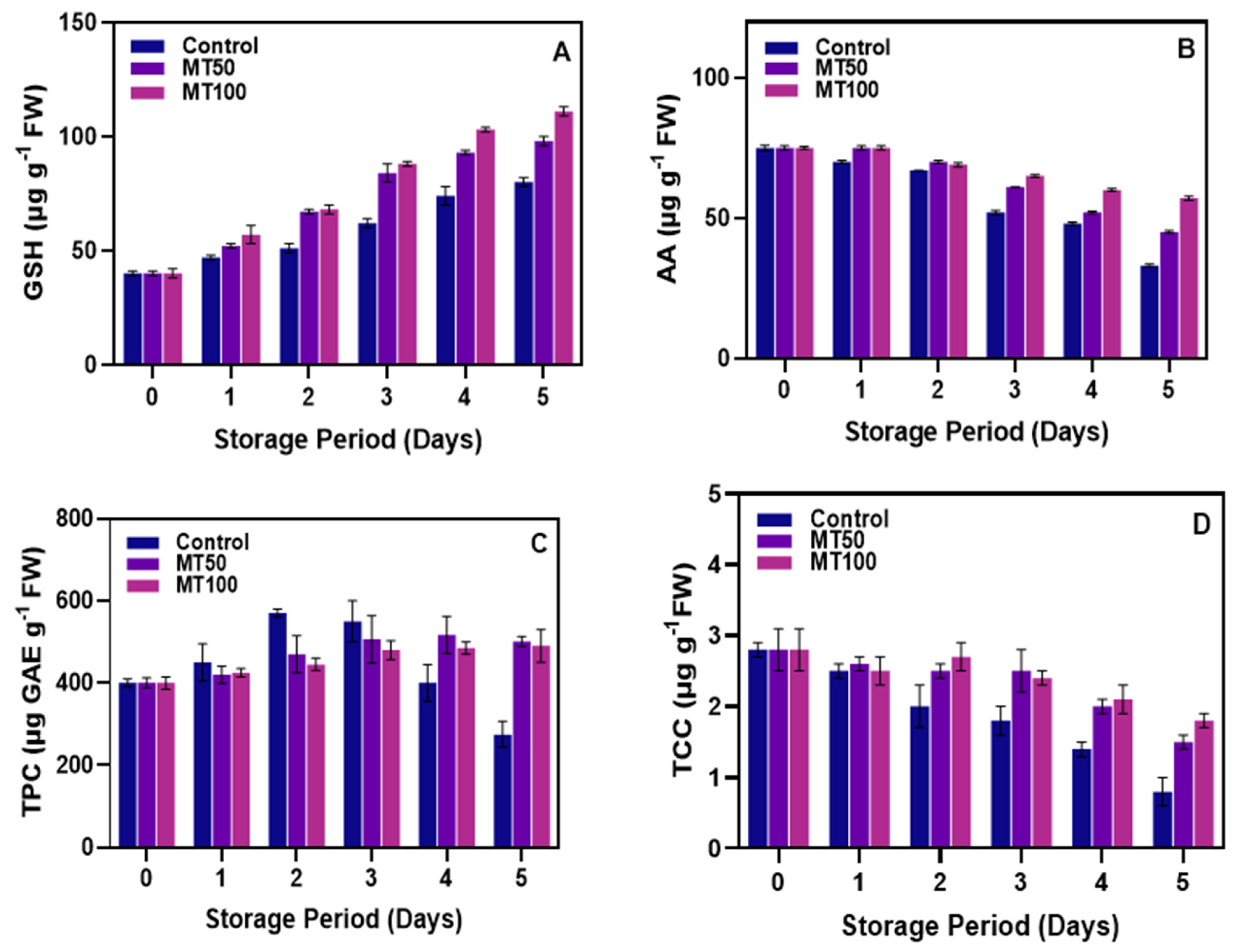
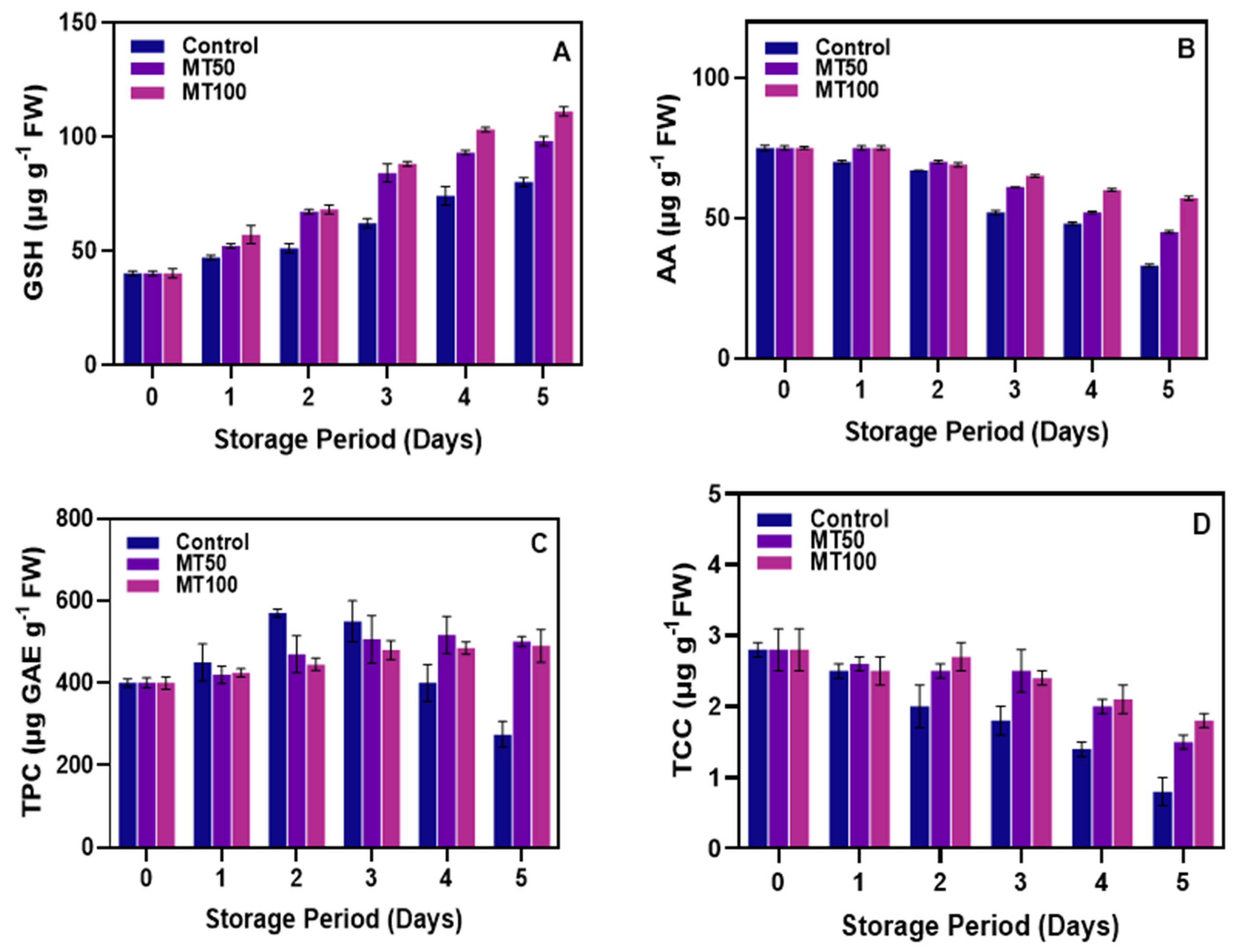
3.3. Changes in GSH, AA, TPC, and TCC Levels
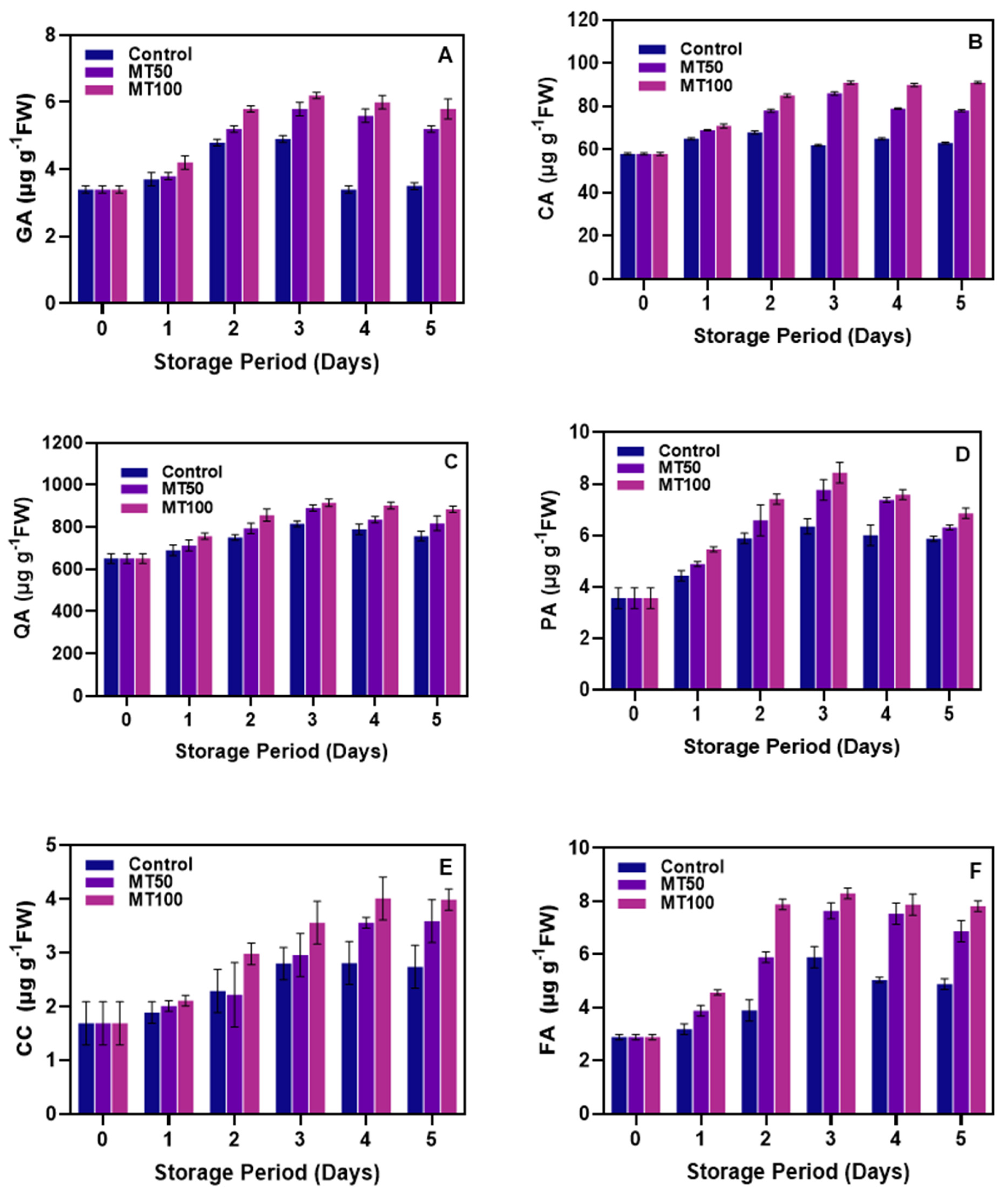
3.4. Changes in Phenolic Acids
3.5. Enzyme Activities
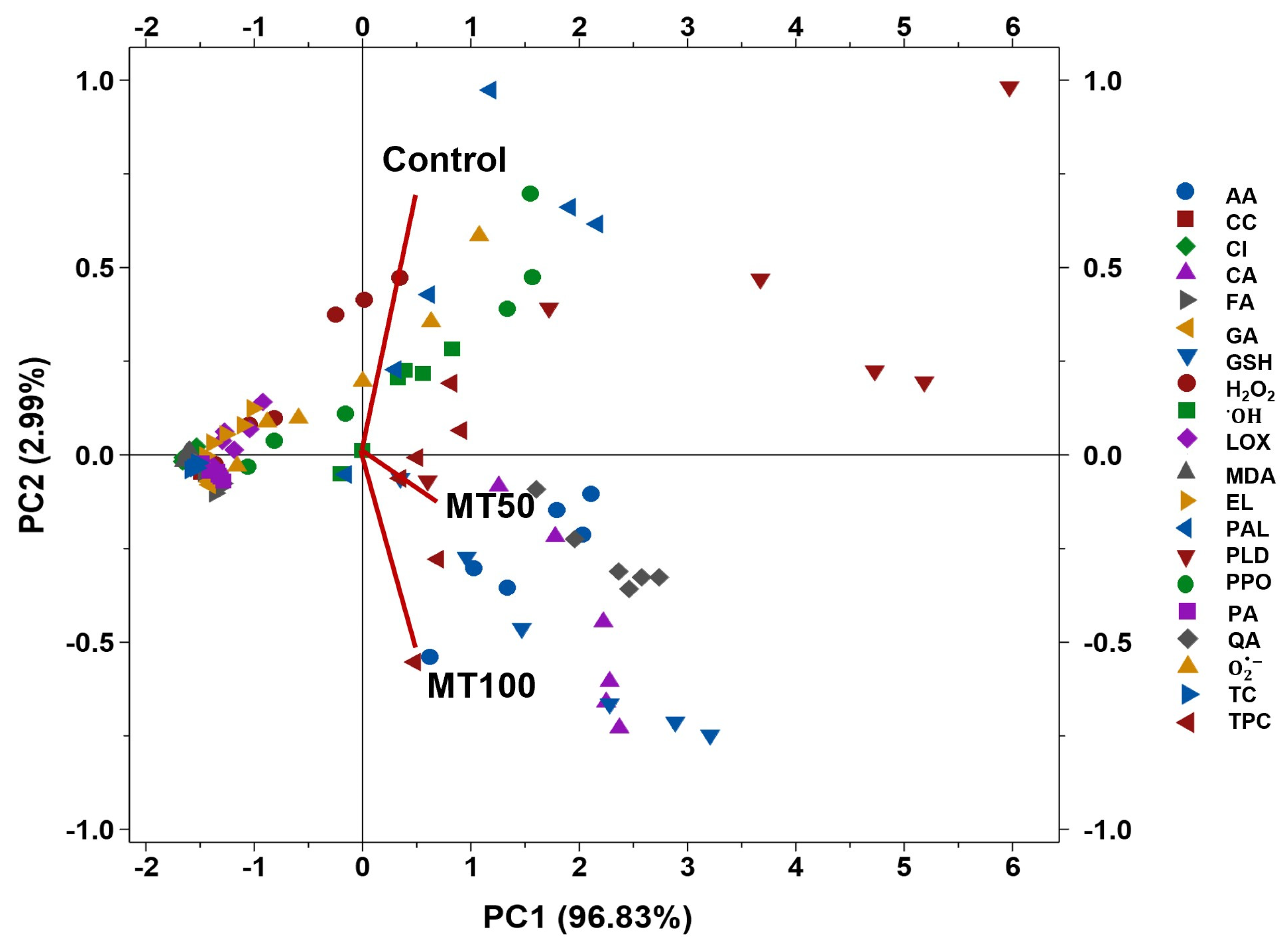
3.6. Correlation and Association of Coating Conditions and Physicochemical Parameters
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Maseko, K.H.; Regnier, T.; Meiring, B.; Wokadala, O.C.; Anyasi, T.A. Musa Species Variation, Production, and the Application of Its Processed Flour: A Review. Sci. Hortic. 2024, 325, 112688. [Google Scholar] [CrossRef]
- Aurore, G.; Parfait, B.; Fahrasmane, L. Bananas, Raw Materials for Making Processed Food Products. Trends Food Sci. Technol. 2009, 20, 78–91. [Google Scholar] [CrossRef]
- Hettiaratchi, U.P.K.; Ekanayake, S.; Welihinda, J. Chemical Compositions and Glycemic Responses to Banana Varieties. Int. J. Food Sci. Nutr. 2011, 62, 307–309. [Google Scholar] [CrossRef] [PubMed]
- Happi Emaga, T.; Andrianaivo, R.H.; Wathelet, B.; Tchango, J.T.; Paquot, M. Effects of the Stage of Maturation and Varieties on the Chemical Composition of Banana and Plantain Peels. Food Chem. 2007, 103, 590–600. [Google Scholar] [CrossRef]
- Anyasi, T.A.; Jideani, A.I.O.; Mchau, G.R.A. Functional Properties and Post-harvest Utilization of Commercial and Noncommercial Banana Cultivars. Compr. Rev. Food Sci. Food Saf. 2013, 12, 509–522. [Google Scholar] [CrossRef] [PubMed]
- Sarma, P.P.; Gurumayum, N.; Verma, A.K.; Devi, R. A Pharmacological Perspective of Banana: Implications Relating to Therapeutic Benefits and Molecular Docking. Food Funct. 2021, 12, 4749–4767. [Google Scholar] [CrossRef] [PubMed]
- Jin, Z.-Q.; Xu, B.-Y.; Liu, J.-H.; Su, W.; Zhang, J.-B.; Yang, X.-L.; Jia, C.-H.; Li, M.-Y. Identification of Genes Differentially Expressed at the Onset of the Ethylene Climacteric in Banana. Postharvest Biol. Technol. 2009, 52, 307–309. [Google Scholar] [CrossRef]
- Bapat, V.A.; Trivedi, P.K.; Ghosh, A.; Sane, V.A.; Ganapathi, T.R.; Nath, P. Ripening of Fleshy Fruit: Molecular Insight and the Role of Ethylene. Biotechnol. Adv. 2010, 28, 94–107. [Google Scholar] [CrossRef] [PubMed]
- Der Agopian, R.G.; Peroni-Okita, F.H.G.; Soares, C.A.; Mainardi, J.A.; do Nascimento, J.R.O.; Cordenunsi, B.R.; Lajolo, F.M.; Purgatto, E. Low Temperature Induced Changes in Activity and Protein Levels of the Enzymes Associated to Conversion of Starch to Sucrose in Banana Fruit. Postharvest Biol. Technol. 2011, 62, 133–140. [Google Scholar] [CrossRef]
- Yang, X.; Zhang, Z.; Joyce, D.; Huang, X.; Xu, L.; Pang, X. Characterization of Chlorophyll Degradation in Banana and Plantain during Ripening at High Temperature. Food Chem. 2009, 114, 383–390. [Google Scholar] [CrossRef]
- Du, L.; Song, J.; Forney, C.; Palmer, L.C.; Fillmore, S.; Zhang, Z. Proteome Changes in Banana Fruit Peel Tissue in Response to Ethylene and High-Temperature Treatments. Hortic. Res. 2016, 3, 16012. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Joyce, D.C.; Jiang, W.; Lu, W. Effects of Chilling Temperatures on Ethylene Binding by Banana Fruit. Plant Growth Regul. 2004, 43, 109–115. [Google Scholar] [CrossRef]
- Li, W.; Shao, Y.; Chen, W.; Jia, W. The Effects of Harvest Maturity on Storage Quality and Sucrose-Metabolizing Enzymes During Banana Ripening. Food Bioprocess. Technol. 2011, 4, 1273–1280. [Google Scholar] [CrossRef]
- Jiang, Y.; Joyce, D.C.; Macnish, A.J. Extension of the Shelf Life of Banana Fruit by 1-Methylcyclopropene in Combination with Polyethylene Bags. Postharvest Biol. Technol. 1999, 16, 187–193. [Google Scholar] [CrossRef]
- Mulagund, J.; Porika, H.; Soorianathasundaram, K.; Deepika, C. Influence of Growth Regulators Combined with Chemicals to Improve Post-harvest Fruit Quality in Banana Cv. Nendran (Musa AAB). J. Food Process Technol. 2015, 6, 1000428. [Google Scholar] [CrossRef]
- Liu, J.; Sun, J.; Pan, Y.; Yun, Z.; Zhang, Z.; Jiang, G.; Jiang, Y. Endogenous Melatonin Generation Plays a Positive Role in Chilling Tolerance in Relation to Redox Homeostasis in Litchi Fruit during Refrigeration. Postharvest Biol. Technol. 2021, 178, 111554. [Google Scholar] [CrossRef]
- Jannatizadeh, A. Exogenous Melatonin Applying Confers Chilling Tolerance in Pomegranate Fruit during Cold Storage. Sci. Hortic. 2019, 246, 544–549. [Google Scholar] [CrossRef]
- Gao, H.; Lu, Z.; Yang, Y.; Wang, D.; Yang, T.; Cao, M.; Cao, W. Melatonin Treatment Reduces Chilling Injury in Peach Fruit through Its Regulation of Membrane Fatty Acid Contents and Phenolic Metabolism. Food Chem. 2018, 245, 659–666. [Google Scholar] [CrossRef]
- Dreux, C. Biochemical studies of methoxy indoles. II—Determination of methoxy-5 tryptamine by spectrofluorometry. Clin. Chim. Acta 1970, 30, 519–526. [Google Scholar] [CrossRef]
- Gonzalez-Gomez, D.; Lozano, M.; Fernandez-Leon, M.F.; Ayuso, M.C.; Bernalte, M.J.; Rodriguez, A.B. Detection and Quantification of Melatonin and Serotonin in Eight Sweet Cherry Cultivars (Prunus Avium L.). Eur. Food Res. Technol. 2009, 229, 223–229. [Google Scholar] [CrossRef]
- Sturtz, M.; Cerezo, A.B.; Cantos-Villar, E.; Garcia-Parrilla, M.C. Determination of the Melatonin Content of Different Varieties of Tomatoes (Lycopersicon Esculentum) and Strawberries (Fragariaananassa). Food Chem. 2011, 127, 1329–1334. [Google Scholar] [CrossRef] [PubMed]
- Kumari, A.; Singh, S.K.; Mathpal, B.; Verma, K.K.; Garg, V.K.; Bhattacharyya, M.; Bhatt, R. The Biosynthesis, Mechanism of Action, and Physiological Functions of Melatonin in Horticultural Plants: A Review. Horticulturae 2023, 9, 913. [Google Scholar] [CrossRef]
- Turk, H.; Erdal, S.; Genisel, M.; Atici, O.; Demir, Y.; Yanmis, D. The Regulatory Effect of Melatonin on Physiological, Biochemical and Molecular Parameters in Cold-Stressed Wheat Seedlings. Plant Growth Regul. 2014, 2, 139–152. [Google Scholar] [CrossRef]
- Liu, G.; Zhang, Y.; Yun, Z.; Hu, M.; Liu, J.; Jiang, Y.; Zhang, Z. Melatonin Enhances Cold Tolerance by Regulating Energy and Proline Metabolism in Litchi Fruit. Foods 2020, 9, 454. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Randhawa, M.S.; Azam, M.; Liu, H.; Ejaz, S.; Ilahy, R.; Qadri, R.; Khan, M.I.; Umer, M.A.; Khan, M.A.; et al. Exogenous Melatonin Treatment Reduces Post-harvest Senescence and Maintains the Quality of Papaya Fruit during Cold Storage. Front. Plant Sci. 2022, 13, 1039373. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Chang, J.; Zheng, J.; Dong, Y.; Liu, Q.; Yang, X.; Wei, C.; Zhang, Y.; Ma, J.; Zhang, X. Local Melatonin Application Induces Cold Tolerance in Distant Organs of Citrullus Lanatus L. via Long Distance Transport. Sci. Rep. 2017, 7, 40858. [Google Scholar] [CrossRef] [PubMed]
- Yang, N.; Han, M.-H.; Teng, R.-M.; Yang, Y.-Z.; Wang, Y.-H.; Xiong, A.-S.; Zhuang, J. Exogenous Melatonin Enhances Photosynthetic Capacity and Related Gene Expression in A Dose-Dependent Manner in the Tea Plant (Camellia Sinensis (L.) Kuntze). Int. J. Mol. Sci. 2022, 23, 6694. [Google Scholar] [CrossRef] [PubMed]
- Mirshekari, A.; Madani, B.; Yahia, E.M.; Golding, J.B.; Vand, S.H. Post-harvest Melatonin Treatment Reduces Chilling Injury in Sapota Fruit. J. Sci. Food Agric. 2020, 100, 1897–1903. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Chen, Y.; Kang, H. Melatonin Is a Potential Target for Improving Post-Harvest Preservation of Fruits and Vegetables. Front. Plant Sci. 2019, 10, 1388. [Google Scholar] [CrossRef]
- Gao, H.; Zhang, Z.K.; Chai, H.K.; Cheng, N.; Yang, Y.; Wang, D.N.; Yang, T.; Cao, W. Melatonin Treatment Delays Post-harvest Senescence and Regulates Reactive Oxygen Species Metabolism in Peach Fruit. Postharvest Biol. Technol. 2016, 118, 103–110. [Google Scholar] [CrossRef]
- Promyou, S.; Raruang, Y.; Chen, Z.-Y. Melatonin Treatment of Strawberry Fruit during Storage Extends Its Post-Harvest Quality and Reduces Infection Caused by Botrytis Cinerea. Foods 2023, 12, 1445. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.L.; Shan, W.; Cai, D.L.; Chen, J.Y.; Lu, W.J.; Su, X.G.; Kuang, J.F. Post-harvest Application of Glycine Betaine Ameliorates Chilling Injury in Cold-Stored Banana Fruit by Enhancing Antioxidant System. Sci. Hortic. 2021, 287, 110264. [Google Scholar] [CrossRef]
- Wang, Y.; Luo, Z.; Huang, X.; Yang, K.; Gao, S.; Du, R. Effect of Exogenous γ-Aminobutyric Acid (GABA) Treatment on Chilling Injury and Antioxidant Capacity in Banana Peel. Sci. Hortic. 2014, 168, 132–137. [Google Scholar] [CrossRef]
- Chen, J.; He, L.; Jiang, Y.; Wang, Y.; Joyce, D.C.; Ji, Z.; Lu, W. Role of Phenylalanine Ammonia-lyase in Heat Pretreatment-induced Chilling Tolerance in Banana Fruit. Physiol. Plant. 2008, 132, 318–328. [Google Scholar] [CrossRef]
- Yang, S.; Su, X.; Yang, B.; Cheng, G.; Chen, Y.; Yang, E.; Jiang, Y. Oxidation and Peroxidation of Post-harvest Banana Fruit during Softening. Pak. J. Bot. 2008, 40, 2023–2202. [Google Scholar]
- Schopfer, P.; Plachy, C.; Frahry, G. Release of Reactive Oxygen Intermediates (Superoxide Radicals, Hydrogen Peroxide, and Hydroxyl Radicals) and Peroxidase in Germinating Radish Seeds Controlled by Light, Gibberellin, and Abscisic Acid. Plant Physiol. 2001, 125, 1591–1602. [Google Scholar] [CrossRef]
- Pongprasert, N.; Sekozawa, Y.; Sugaya, S.; Gemma, H. The role and mode of action of UV-C hormesis in reducing cellular oxidative stress and the consequential chilling injury of banana fruit peel. Int. Food Res. J. 2011, 18, 741–749. [Google Scholar]
- Chotikakham, S.; Faiyue, B.; Uthaibutra, J.; Saengnil, K. Exogenous Methyl Salicylate Alleviates Senescent Spotting by Enhancing the Activity of Antioxidative Ascorbate-Glutathione Cycle in Harvested’ Sucrier’ Bananas. Sci. Hortic. 2020, 267, 109324. [Google Scholar] [CrossRef]
- Nielsen, S.S. Vitamin C Determination by Indophenol Method. In Food Analysis Laboratory Manual; Food Science Text Series; Springer: Cham, Switzerland, 2017. [Google Scholar] [CrossRef]
- Alberti, A.; Zielinski, A.A.F.; Zardo, D.M.; Demiate, I.M.; Nogueira, A.; Mafra, L.I. Optimisation of the Extraction of Phenolic Compounds from Apples Using Response Surface Methodology. Food Chem. 2014, 149, 151–158. [Google Scholar] [CrossRef]
- Liu, J.; Li, F.; Liang, L.; Jiang, Y.; Chen, J. Fibroin Delays Chilling Injury of Post-harvest Banana Fruit via Enhanced Antioxidant Capability during Cold Storage. Metabolites 2019, 9, 152. [Google Scholar] [CrossRef]
- Liu, H.; Cao, J.; Jiang, W. Changes in Phenolics and Antioxidant Property of Peach Fruit during Ripening and Responses to 1-Methylcyclopropene. Postharvest Biol. Technol. 2015, 108, 111–118. [Google Scholar] [CrossRef]
- Lin, Y.; Lin, Y.; Lin, H.; Zhang, S.; Chen, Y.; Shi, J. Inhibitory Effects of Propyl Gallate on Browning and Its Relationship to Active Oxygen Metabolism in Pericarp of Harvested Longan Fruit. LWT Food Sci. Technol. 2015, 60, 1122–1128. [Google Scholar] [CrossRef]
- Yi, C.; Qu, H.X.; Jiang, Y.M.; Shi, J.; Duan, X.W.; Joyce, D.C.; Li, Y.B. ATP-Induced Changes in Energy Status and Membrane Integrity of Harvested Litchi Fruit and Its Relation to Pathogen Resistance. J. Phytopathol. 2008, 156, 365–371. [Google Scholar] [CrossRef]
- Chen, X.; Ren, L.; Li, M.; Qian, J.; Fan, J.; Du, B. Effects of Clove Essential Oil and Eugenol on Quality and Browning Control of Fresh-Cut Lettuce. Food Chem. 2017, 214, 432–439. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y. Role of Anthocyanins, Polyphenol Oxidase and Phenols in Lychee Pericarp Browning. J. Sci. Food Agric. 2000, 80, 305–310. [Google Scholar] [CrossRef]
- Cainelli, N.; Ruperti, B. Biochemistry and Molecular Biology in Fruits during Cold Storage. In Annual Plant Reviews Online; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2019; pp. 659–688. [Google Scholar] [CrossRef]
- Lukatkin, A.; Brazaityte, A.; Bobinas, C.; Duchovskis, P. Chilling Injury in Chilling-Sensitive Plants: A Review. Zemdirb. Agric. 2012, 99, 111–124. [Google Scholar]
- Chang, L.-Y.; Sargent, S.A.; Kim, J.; Brecht, J.K. Delaying Ripening Using 1-MCP Reveals Chilling Injury Symptom Development at the Putative Chilling Threshold Temperature for Mature Green Banana. Front. Plant Sci. 2022, 13, 966789. [Google Scholar] [CrossRef] [PubMed]
- Tian, S.; Qin, G.; Li, B. Reactive Oxygen Species Involved in Regulating Fruit Senescence and Fungal Pathogenicity. Plant Mol. Biol. 2013, 82, 593–602. [Google Scholar] [CrossRef] [PubMed]
- Patel, B.; Tandel, Y.; Patel, A.H.; Patel, B. Chilling Injury in Tropical and Subtropical Fruit: A Cola Storage Problem and Its Remedies: A Review. Int. J. Sci. Env. Technol. 2016, 5, 1882–1887. [Google Scholar]
- Liu, J.; Wu, H.; Wang, B.; Zhang, Y.; Wang, J.; Cheng, C.; Huang, Y. Exogenous Melatonin Enhances Cold Resistance by Improving Antioxidant Defense and Cold-Responsive Genes Expression in Banana. Horticulturae 2022, 8, 260. [Google Scholar] [CrossRef]
- Wang, Z.; Pu, H.; Shan, S.; Zhang, P.; Li, J.; Song, H.; Xu, X. Melatonin Enhanced Chilling Tolerance and Alleviated Peel Browning of Banana Fruit under Low Temperature Storage. Postharvest Biol. Technol. 2021, 179, 111571. [Google Scholar] [CrossRef]
- Qari, S.H.; Hassan, M.U.; Chattha, M.U.; Mahmood, A.; Naqve, M.; Nawaz, M.; Barbanti, L.; Alahdal, M.A.; Aljabri, M. Melatonin Induced Cold Tolerance in Plants: Physiological and Molecular Responses. Front. Plant Sci. 2022, 13, 843071. [Google Scholar] [CrossRef] [PubMed]
- Kong, X.; Ge, W.; Wei, B.; Zhou, Q.; Zhou, X.; Zhao, Y.; Ji, S. Melatonin Ameliorates Chilling Injury in Green Bell Peppers during Storage by Regulating Membrane Lipid Metabolism and Antioxidant Capacity. Postharvest Biol. Technol. 2020, 170, 111315. [Google Scholar] [CrossRef]
- Raza, A.; Charagh, S.; Garcia-Caparros, P.; Rahman, M.A.; Ogwugwa, V.H.; Saeed, F.; Jin, W. Melatonin-Mediated Temperature Stress Tolerance in Plants. GM Crops Food 2022, 13, 196–217. [Google Scholar] [CrossRef]
- Liang, D.; Shen, Y.; Ni, Z.; Wang, Q.; Lei, Z.; Xu, N.; Deng, Q.; Lin, L.; Wang, J.; Lv, X.; et al. Exogenous Melatonin Application Delays Senescence of Kiwifruit Leaves by Regulating the Antioxidant Capacity and Biosynthesis of Flavonoids. Front. Plant Sci. 2018, 9, 426. [Google Scholar] [CrossRef]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive Oxygen Species, Oxidative Damage, and Antioxidative Defense Mechanism in Plants under Stressful Conditions. J. Bot. 2012, 2012, e217037. [Google Scholar] [CrossRef]
- Samiasih, A.; Subagio, W.H.; Dharmana; Susanto, H.; Sadhana, U.; Sunoko, H.R.; Santosa, B. Banana Peels Extract (Musa Paradisiaca Var Kepok) Decreased MDA in New Zealand White Rabbit with DM Hyperlipidemia. IOP Conf. Ser. Earth Environ. Sci. 2019, 292, 012008. [Google Scholar] [CrossRef]
- Liu, X.; Zhou, Y.; Xiao, J.; Bao, F. Effects of Chilling on the Structure, Function and Development of Chloroplasts. Front. Plant Sci. 2018, 9, 1715. [Google Scholar] [CrossRef] [PubMed]
- Miura, K.; Furumoto, T. Cold Signaling and Cold Response in Plants. Int. J. Mol. Sci. 2013, 14, 5312–5337. [Google Scholar] [CrossRef]
- Song, L.; Tan, Z.; Zhang, W.; Li, Q.; Jiang, Z.; Shen, S.; Luo, S.; Chen, X. Exogenous Melatonin Improves the Chilling Tolerance and Preharvest Fruit Shelf Life in Eggplant by Affecting ROS- and Senescence-Related Processes. Hortic. Plant J. 2023, 9, 523–540. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, L.; Duan, W.; Li, W.; Wang, Q.; Li, J.; Song, H.; Xu, X. Melatonin Maintained Higher Contents of Unsaturated Fatty Acid and Cell Membrane Structure Integrity in Banana Peel and Alleviated Post-harvest Chilling Injury. Food Chem. 2022, 397, 133836. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.; Nawaz, A.; Naz, S.; Ali, M.; Ejaz, S.; Azam, M.; Razzaq, K. Exogenous Melatonin Mitigates Chilling Injury in Zucchini Fruit by Enhancing Antioxidant System Activity, Promoting Endogenous Proline and GABA Accumulation, and Preserving Cell Wall Stability. Postharvest Biol. Technol. 2023, 204, 112445. [Google Scholar] [CrossRef]
- Wang, L.; Shen, X.; Chen, X.; Ouyang, Q.; Tan, X.; Tao, N. Exogenous Application of Melatonin to Green Horn Pepper Fruit Reduces Chilling Injury during Postharvest Cold Storage by Regulating Enzymatic Activities in the Antioxidant System. Plants 2022, 11, 2367. [Google Scholar] [CrossRef] [PubMed]
- Madebo, M.P.; Hu, S.; Zheng, Y.; Jin, P. Mechanisms of Chilling Tolerance in Melatonin Treated Post-harvest Fruits and Vegetables: A Review. J. Future Foods 2021, 1, 156–167. [Google Scholar] [CrossRef]
- Das, K.; Roychoudhury, A. Reactive Oxygen Species (ROS) and Response of Antioxidants as ROS-Scavengers during Environmental Stress in Plants. Front. Environ. Sci. 2014, 2, 53. [Google Scholar] [CrossRef]
- Kan, C.; Gao, Y.; Wan, C.; Chen, M.; Zhao, X.; Liu, S.; Chen, J. Influence of Different Cold Storage Times on Quality of “Cuiguan” Pear Fruits during Shelf Life. J. Food Process. Preserv. 2019, 43, e14245. [Google Scholar] [CrossRef]
- Buccheri, M.; Picchi, V.; Grassi, M.; Gandin, D.; Bianchi, G.; Scalzo, R.L. Dynamic Changes of Antioxidants and Fermentative Metabolites in Apple Peel in Relation to Storage, Controlled Atmosphere, and Initial Low Oxygen Stress. Sci. Hortic. 2021, 288, 110312. [Google Scholar] [CrossRef]
- Gapper, N.E.; Hertog, M.L.A.T.M.; Lee, J.; Buchanan, D.A.; Leisso, R.S.; Fei, Z.; Qu, G.; Giovannoni, J.J.; Johnston, J.W.; Schaffer, R.J.; et al. Delayed Response to Cold Stress Is Characterized by Successive Metabolic Shifts Culminating in Apple Fruit Peel Necrosis. BMC Plant Biol. 2017, 17, 77. [Google Scholar] [CrossRef]
- Jiao, J.; Jin, M.; Liu, H.; Suo, J.; Yin, X.; Zhu, Q.; Rao, J. Application of Melatonin in Kiwifruit (Actinidia Chinensis) Alleviated Chilling Injury during Cold Storage. Sci. Hortic. 2022, 296, 110876. [Google Scholar] [CrossRef]
- Galano, A.; Castaneda-Arriaga, R.; Perez-Gonzalez, A.; Tan, D.-X.; Reiter, R.J. Phenolic Melatonin-Related Compounds: Their Role as Chemical Protectors against Oxidative Stress. Molecules 2016, 21, 1442. [Google Scholar] [CrossRef]
- Haminiuk, C.W.I.; Maciel, G.M.; Plata-Oviedo, M.S.V.; Peralta, R.M. Phenolic Compounds in Fruits—An Overview. Int. J. Food Sci. Technol. 2012, 47, 2023–2044. [Google Scholar] [CrossRef]
- Khademi, O.; Ashtari, M.; Razavi, F. Effects of Salicylic Acid and Ultrasound Treatments on Chilling Injury Control and Quality Preservation in Banana Fruit during Cold Storage. Sci. Hortic. 2019, 249, 334–339. [Google Scholar] [CrossRef]
- Navina, B.; Keshav Huthaash, K.; Velmurugan, N.K.; Korumilli, T. Insights into Recent Innovations in Anti Browning Strategies for Fruit and Vegetable Preservation. Trends Food Sci. Technol. 2023, 139, 104128. [Google Scholar] [CrossRef]
- Kan, E.; Sargent, S.; Simonne, A.; Shaw, N.; Cantliffe, D. Changes in the Post-harvest Quality of Datil Hot Peppers as Affected by Storage Temperature. Proc. Fla. State Hort. Soc. 2007, 120, 246–250. [Google Scholar]
- Calvo-Brenes, P.; O’Hare, T. Effect of Freezing and Cool Storage on Carotenoid Content and Quality of Zeaxanthin-Biofortified and Standard Yellow Sweet-Corn (Zea Mays L.). J. Food Compos. Anal. 2020, 86, 103353. [Google Scholar] [CrossRef]
- Manafi, H.; Baninasab, B.; Gholami, M.; Talebi, M.; Khanizadeh, S. Exogenous Melatonin Alleviates Heat-Induced Oxidative Damage in Strawberry (Fragaria × Ananassa Duch. Cv. Ventana) Plant. J. Plant Growth Regul. 2022, 41, 52–64. [Google Scholar] [CrossRef]
- Aludatt, M.H.; Rababah, T.; Alhamad, M.N.; Al-Mahasneh, M.A.; Almajwal, A.; Gammoh, S.; Ereifej, K.; Johargy, A.; Alli, I. A Review of Phenolic Compounds in Oil-Bearing Plants: Distribution, Identification and Occurrence of Phenolic Compounds. Food Chem. 2017, 218, 99–106. [Google Scholar] [CrossRef]
- Ramirez-Bolanos, S.; Perez-Jimenez, J.; Diaz, S.; Robaina, L. A Potential of Banana Flower and Pseudo-Stem as Novel Ingredients Rich in Phenolic Compounds. Int. J. Food Sci. Technol. 2021, 56, 5601–5608. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, Y.; Yang, B.; Li, Y.; Liu, L.; Zhou, W.; Zheng, S.-J. Profiling of Phenolic Compounds of Fruit Peels of Different Ecotype Bananas Derived from Domestic and Imported Cultivars with Different Maturity. Horticulturae 2022, 8, 70. [Google Scholar] [CrossRef]
- Clifford, M.N.; Jaganath, I.B.; Ludwig, I.A.; Crozier, A. Chlorogenic Acids and the Acyl-Quinic Acids: Discovery, Biosynthesis, Bioavailability and Bioactivity. Nat. Prod. Rep. 2017, 34, 1391–1421. [Google Scholar] [CrossRef]
- Valanciene, E.; Malys, N. Advances in Production of Hydroxycinnamoyl-Quinic Acids: From Natural Sources to Biotechnology. Antioxidants 2022, 11, 2427. [Google Scholar] [CrossRef] [PubMed]
- Saravanan, K.; Aradhya, S.M. Polyphenols of Pseudostem of Different Banana Cultivars and Their Antioxidant Activities. J. Agric. Food Chem. 2011, 59, 3613–3623. [Google Scholar] [CrossRef]
- Siqueira, M.D.S.B.; Barbosa, N.C.; Cordenunsi, B.R.; Hassimotto, N.M.A.; de Resende, E.D. Chemical Changes in Prata-Ana Banana Stored under Reduced O2 and Increased CO2 Levels. Acta Sci. Technol. 2018, 40, 20219951313. [Google Scholar] [CrossRef]
- Ali, S.; Nawaz, A.; Naz, S.; Ejaz, S.; Maqbool, M.; Siddiqui, M.H.; Kalaji, H.M.; Wróbel, J.; Telesiński, A.; Auriga, A. Hydrogen Sulfide Mitigates Chilling Injury of Post-harvest Banana Fruits by Regulating γ-Aminobutyric Acid Shunt Pathway and Ascorbate–Glutathione Cycle. Front. Plant Sci. 2022, 13, 941246. [Google Scholar] [CrossRef]
- Ahammed, G.; Wu, M.; Wang, Y.; Yan, Y.; Mao, Q.; Ren, J.; Ma, R.; Liu, A.; Chen, S. Melatonin Alleviates Iron Stress by Improving Iron Homeostasis, Antioxidant Defense and Secondary Metabolism in Cucumber. Sci. Hortic. 2020, 265, 109205. [Google Scholar] [CrossRef]
- Li, S.; Xu, Y.; Bi, Y.; Zhang, B.; Shen, S.; Jiang, T.; Zheng, X. Melatonin Treatment Inhibits Gray Mold and Induces Disease Resistance in Cherry Tomato Fruit during Post-harvest. Postharvest Biol. Technol. 2019, 157, 110962. [Google Scholar] [CrossRef]
- Jahan, M.S.; Guo, S.; Baloch, A.R.; Sun, J.; Shu, S.; Wang, Y.; Ahammed, G.J.; Kabir, K.; Roy, R. Melatonin Alleviates Nickel Phytotoxicity by Improving Photosynthesis, Secondary Metabolism and Oxidative Stress Tolerance in Tomato Seedlings. Ecotoxicol. Env. Saf. 2020, 197, 110593. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Huber, D.J.; Hu, M.; Jiang, G.; Gao, Z.; Xu, X.; Jiang, Y.; Zhang, Z. Delay of Post-harvest Browning in Litchi Fruit by Melatonin via the Enhancing of Antioxidative Processes and Oxidation Repair. J. Agric. Food Chem. 2018, 66, 7475–7484. [Google Scholar] [CrossRef] [PubMed]
- Jayanna, S.K.; Umesha, S. Enhancement of the Expression of Defense Genes in Tomato against Ralstonia Solanacearum by N-Octanoyl-L-Homoserine Lactone. Afr. J. Microbiol. Res. 2017, 11, 194–203. [Google Scholar] [CrossRef]
- Kaur, S.; Samota, M.K.; Choudhary, M.; Choudhary, M.; Pandey, A.K.; Sharma, A.; Thakur, J. How Do Plants Defend Themselves against Pathogens-Biochemical Mechanisms and Genetic Interventions. Physiol. Mol. Biol. Plants 2022, 28, 485–504. [Google Scholar] [CrossRef]
- Hu, W.; Yang, H.; Tie, W.; Yan, Y.; Ding, Z.; Liu, Y.; Wu, C.; Wang, J.; Reiter, R.J.; Tan, D.-X.; et al. Natural Variation in Banana Varieties Highlights the Role of Melatonin in Post-harvest Ripening and Quality. J. Agric. Food Chem. 2017, 65, 9987–9994. [Google Scholar] [CrossRef] [PubMed]
- Aghdam, M.S.; Jannatizadeh, A.; Luo, Z.; Paliyath, G. Ensuring Sufficient Intracellular ATP Supplying and Friendly Extracellular ATP Signaling Attenuates Stresses, Delays Senescence and Maintains Quality in Horticultural Crops during Post-harvest Life. Trends Food Sci. Technol. 2018, 76, 67–81. [Google Scholar] [CrossRef]
- Wang, Y.; Reiter, R.J.; Chan, Z. Phytomelatonin: A Universal Abiotic Stress Regulator. J. Exp. Bot. 2018, 69, 963–974. [Google Scholar] [CrossRef] [PubMed]
- Bartwal, A.; Mall, R.; Lohani, P.; Guru, S.K.; Arora, S. Role of Secondary Metabolites and Brassinosteroids in Plant Defense Against Environmental Stresses. J. Plant Growth Regul. 2013, 32, 216–232. [Google Scholar] [CrossRef]
- Venkatachalam, K.; Charoenphun, N.; Lekjing, S.; Noonim, P. Investigation of Melatonin Incorporated CMC-Gelatin Based Edible Coating on the Alleviation of Chilling Injury Induced Pericarp Browning in Longkong. Foods 2024, 13, 72. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Charoenphun, N.; Chucherd, P.; Paulraj, B.; Venkatachalam, K. Influence of Melatonin Coating on Physiochemical Qualities and Enzymatic Activities in Banana Pericarp under Cold Storage. Horticulturae 2024, 10, 364. https://doi.org/10.3390/horticulturae10040364
Charoenphun N, Chucherd P, Paulraj B, Venkatachalam K. Influence of Melatonin Coating on Physiochemical Qualities and Enzymatic Activities in Banana Pericarp under Cold Storage. Horticulturae. 2024; 10(4):364. https://doi.org/10.3390/horticulturae10040364
Chicago/Turabian StyleCharoenphun, Narin, Pakawan Chucherd, Balaji Paulraj, and Karthikeyan Venkatachalam. 2024. "Influence of Melatonin Coating on Physiochemical Qualities and Enzymatic Activities in Banana Pericarp under Cold Storage" Horticulturae 10, no. 4: 364. https://doi.org/10.3390/horticulturae10040364
APA StyleCharoenphun, N., Chucherd, P., Paulraj, B., & Venkatachalam, K. (2024). Influence of Melatonin Coating on Physiochemical Qualities and Enzymatic Activities in Banana Pericarp under Cold Storage. Horticulturae, 10(4), 364. https://doi.org/10.3390/horticulturae10040364