

Unlocking the Potential of In Vitro Photoautotrophy for Eryngium foetidum: Biomass, Morphophysiology, and Acclimatization

 , , , ,
, , , ,  , ,
, ,  , and
, and 
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimental Design
2.2. Chlorophyll a Fluorescence
2.3. Determination of Photosynthetic Pigments
2.4. Quantification of Photosynthetic Performance
2.5. Growth Analyzes
2.6. Leaf Anatomical Analysis
2.7. Ex Vitro Acclimatization
2.8. Statistical Analyzes
3. Results
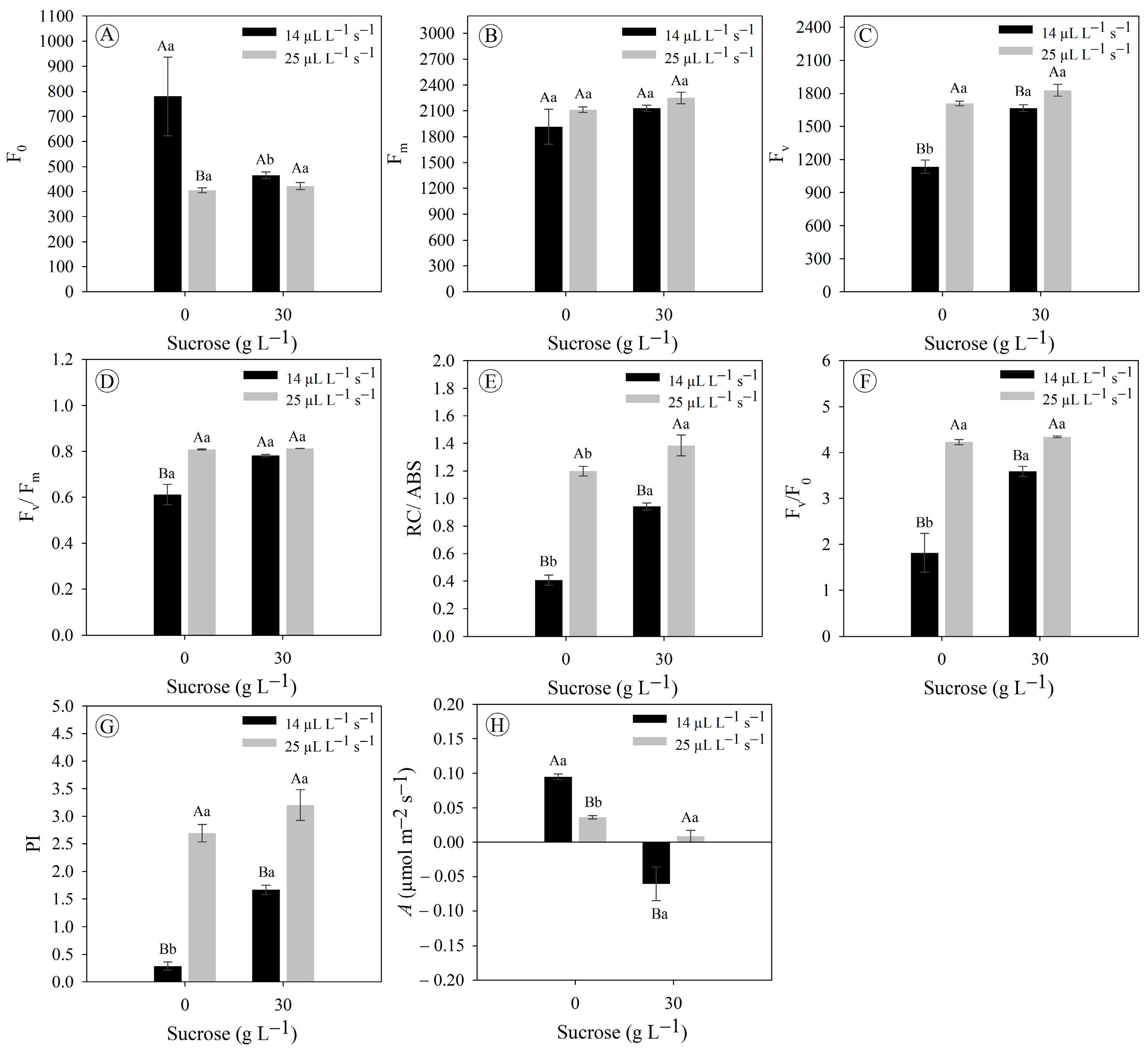
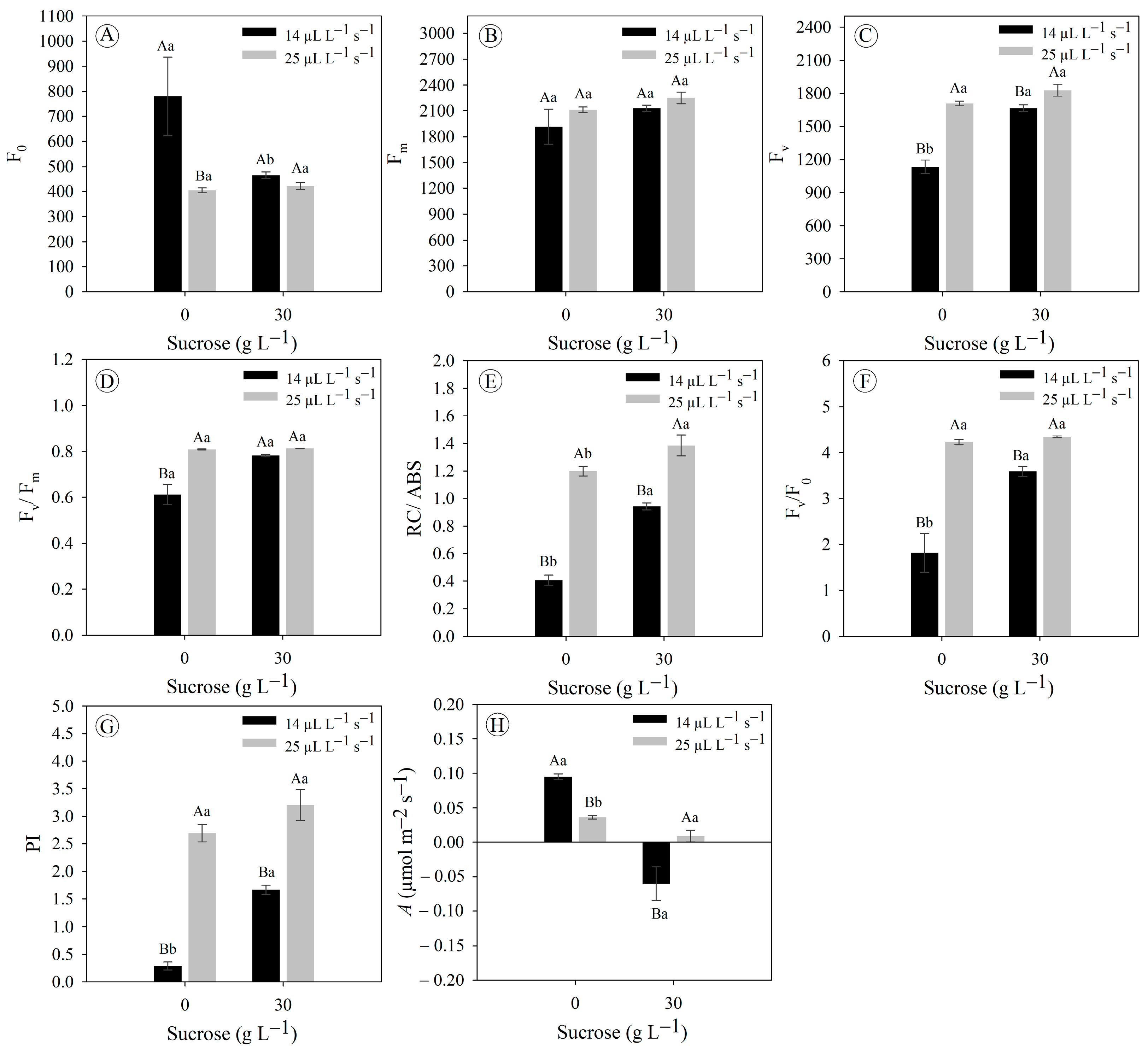
3.1. Gas Exchange Rates Improve Chlorophyll a Fluorescence and Photosynthetic Rate in Eryngium foetidum Grown In Vitro
3.2. Sucrose Concentrations and Gas Exchange Rates Do Not Modulate Pigment Concentration in Eryngium foetidum Grown In Vitro
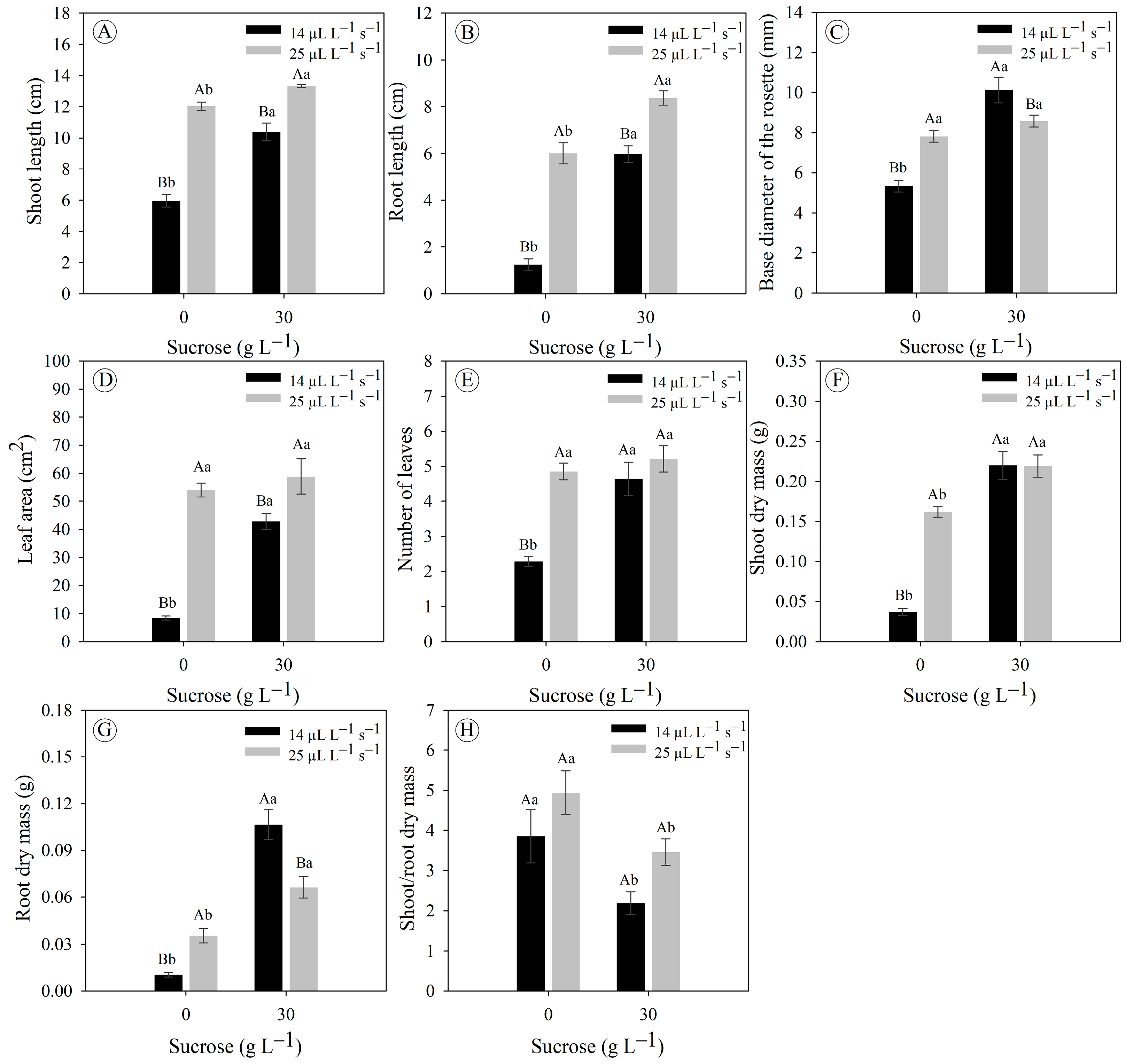
3.3. Sucrose Concentrations and Gas Exchange Rates Affect the In Vitro Growth and Development of Eryngium foetidum
3.4. Sucrose Supplementation and Gas Exchange Rates Affect Leaf Anatomy
3.5. Ex Vitro Acclimatization of Eryngium foetidum Grown under Sucrose Concentrations and Gas Exchange Rates
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Castro, T.F.D.; Carneiro, W.F.; Reichel, T.; Fabem, S.L.; Machado, M.R.F.; Souza, K.K.C.; Resende, L.V.; Murgas, L.D.S. The toxicological effects of Eryngium foetidum extracts on zebrafish embryos and larvae depend on the type of extract, dose, and exposure time. Toxicol. Res. 2022, 11, 891–899. [Google Scholar] [CrossRef] [PubMed]
- Lucas, D.B.; Cardozo, A.L. Eryngium in Flora e Funga do Brasil. Jardim Botânico do Rio de Janeiro. Available online: https://floradobrasil.jbrj.gov.br/FB15529 (accessed on 5 December 2023).
- POWO. Plants of the World Online. Available online: https://powo.science.kew.org/ (accessed on 30 November 2023).
- Singh, S.; Singh, D.R.; Banu, S.; Salim, K.M. Determination of bioactives and antioxidant activity in Eryngium foetidum L.: A traditional culinary and medicinal herb. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2013, 83, 453–460. [Google Scholar] [CrossRef]
- Anju, T.; Rai, N.K.S.; Uthirchamkavu, I.; Sreedharan, S.; Ndhlala, A.R.; Singh, P.; Kumar, A. Analysis of nutritional and antioxidant potential of three traditional leafy vegetables for food security and human wellbeing. S. Afr. J. Bot. 2022, 145, 99–110. [Google Scholar] [CrossRef]
- Do Socorro Teixeira Costa Leitão, D.; Pereira Barbosa-Carvalho, A.P.; Campos de Siqueira, F.; Pontes s Sousa, R.; Santos Lopes, A.; Campos Chisté, R. Extracts of Eryngium foetidum Leaves from the Amazonia Were Efficient Scavengers of ROS and RNS. Antioxidants 2023, 12, 1112. [Google Scholar] [CrossRef]
- Rodrigues, T.L.M.; Silva, M.E.P.; Gurgel, E.S.C.; Oliveira, M.S.; Lucas, F.C.A. Eryngium foetidum L. (Apiaceae): A literature review of traditional uses, chemical composition, and pharmacological activities. Evid. Based Complement. Alternat Med. 2022, 14, 2896895. [Google Scholar] [CrossRef] [PubMed]
- Paw, M.; Gogoi, R.; Sarma, N.; Saikia, S.; Chanda, S.K.; Lekhak, H.; Lal, M. Anti-microbial, anti-oxidant, anti-diabetic study of leaf essential oil of Eryngium foetidum L. along with the chemical profiling collected from North East India. J. Essent. Oil-Bear. 2022, 26, 814–829. [Google Scholar] [CrossRef]
- Zhang, X.; Chen, J.; Zhou, S.; Zhao, H. Ethanol extract of Eryngium foetidum leaves induces mitochondrial associated apoptosis via ROS generation in human gastric cancer cells. Nutr. Cancer 2022, 74, 2996–3006. [Google Scholar] [CrossRef]
- Rodrigues, T.L.M. Desempenho Fisiológico e Perfil Químico do Óleo Essencial. Orientadora: Eloisa Helena de Aguiar Andrade. Masters’s Thesis, Universidade Federal Rural da Amazônia, Museu Paraense Emílio Goeldi, Belém, Brazil, 2019; 60p. Available online: http://repositorio.ufra.edu.br/jspui/handle/123456789/812 (accessed on 10 December 2023).
- Devi, P.B.; Deb, P.; Singh, H.B. Promotion, utilization, and commercial cultivation of local spices with special reference to eryngo (Eryngium foetidum L.) as a measure for livelihood improvement towards achieving the goal of sustainable development in the indo-burma biodiversity hotspot: A case study from manipur, North-East India. In Climate Change Adaptation, Risk Management and Sustainable Practices in the Himalay; Sharma, S., Kuniyal, J.C., Chand, P., Singh, P., Eds.; Springer International Publishing: Cham, Switzerland, 2023; pp. 253–266. [Google Scholar] [CrossRef]
- Rajamohan, S. Harbouring the potential of medicinal and aromatic plants of India: Novel biotechnological approach and extraction technologies. In Medicinal and Aromatic Plants of India Vol. 1. Medicinal and Aromatic Plants of the World, Vol. 8; Máthé, Á., Khan, I.A., Eds.; Springer International Publishing: Cham, Switzerland, 2022; pp. 323–339. [Google Scholar] [CrossRef]
- Karuppusamy, S. A review on trends in production of secondary metabolites from higher plants by in vitro tissue, organ and cell cultures. J. Med. Plants Res. 2009, 3, 1222–1239. [Google Scholar]
- Fazili, M.A.; Bashir, I.; Ahmad, M.; Yaqoob, U.; Geelani, S.N. In vitro strategies for the enhancement of secondary metabolite production in plants: A review. Bull. Natl. Res. Cent. 2022, 46, 35. [Google Scholar] [CrossRef]
- Nhut, D.T. General information: Some aspects of plant tissue culture. In Plant Tissue Culture: New Techniques and Application in Horticultural Species of Tropical Region; Nhut, D.T., Tung, H.T., Yeung, E.C.T., Eds.; Springer: Singapore, 2022; pp. 1–23. [Google Scholar] [CrossRef]
- Kozai, T.; Kubota, C. Concepts, definitions, ventilation methods, advantages and disadvantages. In Photoautotrophic (Sugar–Free Medium) Micropropagation as a New Micropropagation and Transplant Production System; Kozai, T., Afreen, F., Zobayed, S., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 19–30. [Google Scholar] [CrossRef]
- Ramos, J.C.M.; Ribeiro, L.M.; Nunes, G.P.; Soares, J.S.; Sorgato, J.C. In vitro and ex vitro production of Schomburgkia crispa: Effect of flask sealing systems and different light sources. Rodriguesia 2023, 74, e01062022. [Google Scholar] [CrossRef]
- Saldanha, C.W.; Otoni, C.G.; Azevedo, J.L.F.; Dias, L.L.C.; Rêgo, M.M.; Otoni, W.C. A low-cost alternative membrane system that promotes growth in nodal cultures of Brazilian ginseng [Pfaffia glomerata (Spreng.) Pedersen]. Plant Cell Tiss. Organ. Cult. 2012, 110, 413–422. [Google Scholar] [CrossRef]
- Fortini, E.A.; Batista, D.S.; Mamedes-Rodrigues, T.C.; Felipe, S.H.S.; Correia, L.N.F.; Chagas, K.; Silva, P.O.; Rocha, D.I.; Otoni, W.C. Gas exchange rates and sucrose concentrations affect plant growth and production of flavonoids in Vernonia condensata grown in vitro. Plant Cell Tiss. Organ. Cult. 2021, 144, 593–605. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Batista, D.S.; Dias, L.L.C.; Rêgo, M.M.; Saldanha, C.W.; Otoni, W.C. Flask sealing on in vitro seed germination and morphogenesis of two types of ornamental pepper explants. Ciênc. Rural. 2017, 47, e20150245. [Google Scholar] [CrossRef]
- Bolhar-Nordenkampf, H.R.; Long, S.P.; Baker, N.R.; Oquist, G.; Schreiber, U.L.E.G.; Lechner, E.G. Chlorophyll fluorescence as a probe of the photosynthetic competence of leaves in the field: A review of current instrumentation. Funct. Ecol. 1989, 3, 497–514. [Google Scholar] [CrossRef]
- Santos, R.P.; Ferreira Da Cruz, A.C.; Iarema, L.; Kuki, K.N.; Campos Otoni, W. Protocolo para extração de pigmentos foliares em porta-enxertos de videira micropropagados. Rev. Ceres 2008, 55, 356–364. [Google Scholar]
- Wellburn, A.R. The spectral determination of chlorophylls a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Castro, K.M.; Batista, D.S.; Fortini, E.A.; Silva, T.D.; Felipe, S.H.S.; Fernandes, A.M.; Sousa, R.M.J.; Nascimento, L.S.Q.; Campos, V.R.; Grazul, R.M.; et al. Photoperiod modulates growth, morphoanatomy, and linalool content in Lippia alba L. (Verbenaceae) cultured in vitro. Plant Cell Tiss. Organ. Cult. 2019, 139, 139–153. [Google Scholar] [CrossRef]
- Silva, T.D.; Batista, D.S.; Fortini, E.A.; Castro, K.M.; Felipe, S.H.S.; Fernandes, A.M.; Sousa, R.M.J.; Chagas, K.; Silva, J.V.S.; Correia, L.N.F.; et al. Blue and red light affects morphogenesis and 20-hydroxyecdisone content of in vitro Pfaffia glomerata accessions. J. Photochem. Photobiol. B Biol. 2020, 203, 111761. [Google Scholar] [CrossRef]
- Johansen, D.A. Plant Microtechnique; Mac Graw-Hill Book Co., Inc.: New York, NY, USA, 1940; 523p. [Google Scholar]
- O’Brien, T.P.; McCully, M.E. The Study of Plant Structure: Principles and Selected Methods; Termarcarphi Pty., Ltd.: Melbourne, Australia, 1981. [Google Scholar]
- Cruz, C.D. Genes: A software package for analysis in experimental statistics and quantitative genetics. Acta Sci. Agron. 2013, 35, 271–276. [Google Scholar] [CrossRef]
- Ferreira, P.R.B.; Cruz, A.C.F.; Batista, D.S.; Nery, L.A.; Andrade, I.G.; Rocha, D.I.; Felipe, S.H.S.; Koehler, A.D.; Nunes-Nesi, A. CO2 enrichment and supporting material impact the primary metabolism and 20-hydroxyecdysone levels in Brazilian ginseng grown under photoautotrophy. Plant Cell Tiss. Organ. Cult. 2019, 139, 77–89. [Google Scholar] [CrossRef]
- Louback, E.; Batista, D.S.; Pereira, T.A.R.; Mamedes-Rodrigues, T.C.; Silva, T.D.; Felipe, S.H.S.; Rocha, D.I.; Steinmacher, D.A.; Otoni, W.C. CO2 enrichment leads to altered cell wall composition in plants of Pfaffia glomerata (Spreng.) Pedersen (Amaranthaceae). Plant Cell Tiss. Organ. Cult. 2021, 145, 603–613. [Google Scholar] [CrossRef]
- Luis, S.J.; Jabín, B.J. CO2–enriched air in a temporary immersion system induces photomixotrophism during in vitro multiplication in vanilla. Plant Cell Tiss. Organ. Cult. 2023, 155, 29–39. [Google Scholar] [CrossRef]
- Wolf, S.; Kalman-Rotem, N.; Yakir, D.; Zrv, M. Autotrophic and heterotrophic carbon assimilation of in vitro grown potato (Solanum tuberosum L) plants. J. Plant Physiol. 1998, 153, 574–580. [Google Scholar] [CrossRef]
- Shin, K.S.; Park, S.Y.; Paek, K.Y. Sugar metabolism, photosynthesis, and growth of in vitro plantlets of Doritaenopsis under controlled microenvironmental conditions. In Vitro Cell Dev. Biol. Plant 2013, 49, 445–454. [Google Scholar] [CrossRef]
- Ribeiro, I.S.; Ribeiro, L.M.; Soares, J.S.; Ramos, J.C.M.; Sorgato, J.C. Light condition, flask sealing, and cultivation time on the germination and initial in vitro development of Dendrobium nobile Lindl. Ornam. Hortic. 2022, 28, 407–413. [Google Scholar] [CrossRef]
- Vinh, B.V.T.; Tung, H.T.; Mai, N.T.N.; Khai, H.D.; Luan, V.Q.; Phong, T.H.; Phong, T.N.P.; Nhut, D.T. Enhanced efficient micropropagation and reduced abnormal phenomena in Phyllanthus amarus plantlets cultured on medium containing silver nanoparticles. S. Afr. J. Bot. 2023, 163, 217–225. [Google Scholar] [CrossRef]
- Askari, N.; Aliniaeifard, S.; Visser, R.G.F. Low CO2 Levels are detrimental for in vitro plantlets through disturbance of photosynthetic functionality and accumulation of reactive oxygen species. Horticulturae 2022, 8, 44. [Google Scholar] [CrossRef]
- Silva, L.M.; Carvalho, V.S.; Generoso, A.L.; Miranda, D.P.; Costa Júnior, O.D.; Simioni, P.F.; Santana, D.B.; Cunha, M.; Oliveira, J.G.; Viana, A.P. Micropropagation of interspecific hybrids of Vitis spp. in microenvironments with different gas exchanges. Sci. Hortic. 2022, 305, 111413. [Google Scholar] [CrossRef]
- Carrari-Santos, R.; Vettorazzi, R.G.; Pinto, V.B.; Pinto, V.B.; Sena, E.O.A.; Oliveira, J.G.; Campostrini, E.; Silveira, V.; Santa-Catarina, C. Microporous membrane and culture medium affect in vitro seedling development of Dalbergia nigra (Vell.) Ex Benth. (Fabaceae) by modulation of the protein profile and accumulation of ethylene and CO2. Plant Cell Tiss. Organ. Cult. 2023, 153, 559–576. [Google Scholar] [CrossRef]
- Iarema, L.; Cruz, A.C.F.; Saldanha, C.W.; Dias, L.L.C.; Vieira, R.F.; Oliveira, E.J.; Otoni, W.C. Photoautotrophic propagation of Brazilian ginseng [Pfaffia glomerata (Spreng.) Pedersen]. Plant Cell Tiss. Organ. Cult. 2012, 110, 227–238. [Google Scholar] [CrossRef]
- Wang, K.L.C.; Li, H.; Ecker, J.R. Ethylene biosynthesis and signaling networks. Plant Cell 2002, 14 (Suppl. S1), S131–S151. [Google Scholar] [CrossRef] [PubMed]
- Kubota, C. Concepts and background of photoautotrophic micropropagation. In Molecular Breeding of Woody Plants; Morohoshi, N., Komamine, A., Eds.; Elsevier: Amsterdam, The Netherlands, 2001; pp. 325–334. [Google Scholar]
- Cournac, L.; Dimon, B.; Carrier, P.; Lohou, A.; Chagvardieff, P. Growth and photosynthetic characteristics of Solanum tuberosum plantlets cultivated in vitro in different conditions of aeration, sucrose supply, and CO2 enrichment. Plant Physiol. 1991, 97, 112–117. [Google Scholar] [CrossRef]
- Desjardins, Y.; Hdider, C.; Riek, J. Carbon nutrition in vitro—Regulation and manipulation of carbon assimilation in micropropagated systems. In Automation and Environmental Control in Plant Tissue Culture; Aitken-Christie, J., Kozai, T., Smith, M.A.L., Eds.; Springer: Dordrecht, The Netherlands, 1995; pp. 441–471. [Google Scholar] [CrossRef]
- Mayak, S.; Tirosh, T.; Ilan, A.; Duvdevani, A.; Khayat, E. Growth and development of pineapple (Ananas comosus L.) plantlets cultured in vitro at enriched and ambient CO2 environments. In International Symposium on Biotechnology of Tropical and Subtropical Species Part 2; ISHS: Brisbane, Australia, 1998; Volume 461, pp. 225–230. [Google Scholar] [CrossRef]
- Marques, I.; Fernandes, I.; Paulo, O.S.; Lidon, F.C.; DaMatta, F.M.; Ramalho, J.C.; Ribeiro-Barros, A.I. A transcriptomic approach to understanding the combined impacts of supra-optimal temperatures and CO2 revealed different responses in the polyploid Coffea arabica and its diploid progenitor C. canephora. Int. J. Mol. Sci. 2021, 22, 3125. [Google Scholar] [CrossRef] [PubMed]
- Martins, J.P.R.; Almeida-Rodrigues, L.C.; Santos, E.R.; Gontijo, A.B.P.L.; Falqueto, A.R. Impacts of photoautotrophic, photomixotrophic, and heterotrophic conditions on the anatomy and photosystem II of in vitro-propagated Aechmea blanchetiana (Baker) L.B. Sm. (Bromeliaceae). In Vitro Cell. Dev. Biol. Plant 2020, 56, 350–361. [Google Scholar] [CrossRef]
- Rocha, T.T.; Araújo, D.X.; Silva, A.M.; Oliveira, J.P.V.; Carvalho, A.A.; Gavilanes, M.L.; Bertolucci, S.K.V.; Alves, E.A.; Pinto, J.E.B.P. Morphoanatomy and changes in antioxidant defense associated with the natural ventilation system of micropropagated Lippia dulcis plantlets. Plant Cell Tiss. Organ. Cult. 2022, 151, 467–481. [Google Scholar] [CrossRef]
- Soares, J.S.; Ramos, J.C.M.; Sorgato, J.C.; Ribeiro, L.M.; Reis, L.C. Brassavola tuberculata Hook.: In vitro growth and ex vitro establishment as a function of the micropropagation system and sucrose. Braz. J. Biol. 2023, 83, e270892. [Google Scholar] [CrossRef]
- Gonçalves, V.P.; Andrade, F.L.N.; Gomes, R.F.; Santos, L.S. Genetic diversity in creole genotypes of Amazon chicory. Acta Amazon. 2022, 52, 89–95. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, D.R.; Soares, M.N.B.; Silva, M.C.R.; Lima, M.C.; Silva-Moraes, V.K.d.O.; Alves, G.L.; Ríos-Ríos, A.M.; Lima, A.d.S.; Pinheiro, M.V.M.; Corrêa, T.R.; et al. Unlocking the Potential of In Vitro Photoautotrophy for Eryngium foetidum: Biomass, Morphophysiology, and Acclimatization. Horticulturae 2024, 10, 107. https://doi.org/10.3390/horticulturae10010107
Silva DR, Soares MNB, Silva MCR, Lima MC, Silva-Moraes VKdO, Alves GL, Ríos-Ríos AM, Lima AdS, Pinheiro MVM, Corrêa TR, et al. Unlocking the Potential of In Vitro Photoautotrophy for Eryngium foetidum: Biomass, Morphophysiology, and Acclimatization. Horticulturae. 2024; 10(1):107. https://doi.org/10.3390/horticulturae10010107
Chicago/Turabian StyleSilva, Darlyara Reis, Marion Nayon Braga Soares, Maria Cristina Rocha Silva, Mariana Costa Lima, Vitória Karla de Oliveira Silva-Moraes, Givago Lopes Alves, Anyela Marcela Ríos-Ríos, Aldilene da Silva Lima, Marcos Vinícius Marques Pinheiro, Thais Roseli Corrêa, and et al. 2024. "Unlocking the Potential of In Vitro Photoautotrophy for Eryngium foetidum: Biomass, Morphophysiology, and Acclimatization" Horticulturae 10, no. 1: 107. https://doi.org/10.3390/horticulturae10010107
APA StyleSilva, D. R., Soares, M. N. B., Silva, M. C. R., Lima, M. C., Silva-Moraes, V. K. d. O., Alves, G. L., Ríos-Ríos, A. M., Lima, A. d. S., Pinheiro, M. V. M., Corrêa, T. R., Batista, D. S., Figueiredo, F. A. M. M. d. A., Reis, F. d. O., Ferraz, T. M., & Felipe, S. H. S. (2024). Unlocking the Potential of In Vitro Photoautotrophy for Eryngium foetidum: Biomass, Morphophysiology, and Acclimatization. Horticulturae, 10(1), 107. https://doi.org/10.3390/horticulturae10010107