
New Observation in Biocontrol of Penicillium caperatum against Fusarium oxysporum on Saposhnikovia divaricata and as a Plant Growth Promoter

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Rhizospheric Fungi of S. divaricata
2.2. In Vitro Inhibition Activity and Inhibition Spectrum
2.3. Culture Characteristics and Phylogenetic Analysis of MR-16
2.4. Effects of MR-16 on Mycelial Growth of F. oxysporum
2.5. Effects of MR-16 on Conidial Germination of F. oxysporum
2.6. Soil Colonization Assays
2.7. Biocontrol Activity of MR-16 Isolate on S. divaricata Wilt
2.8. Plant Growth Promotion of MR-16 Isolate
2.9. Statistical Analysis
3. Results
3.1. Antagonistic Activities of Fungal Isolate against F. oxysporum
3.2. Antifungal Spectrum of MR-16 Isolate
3.3. Identification of Antagonistic Strain MR-16
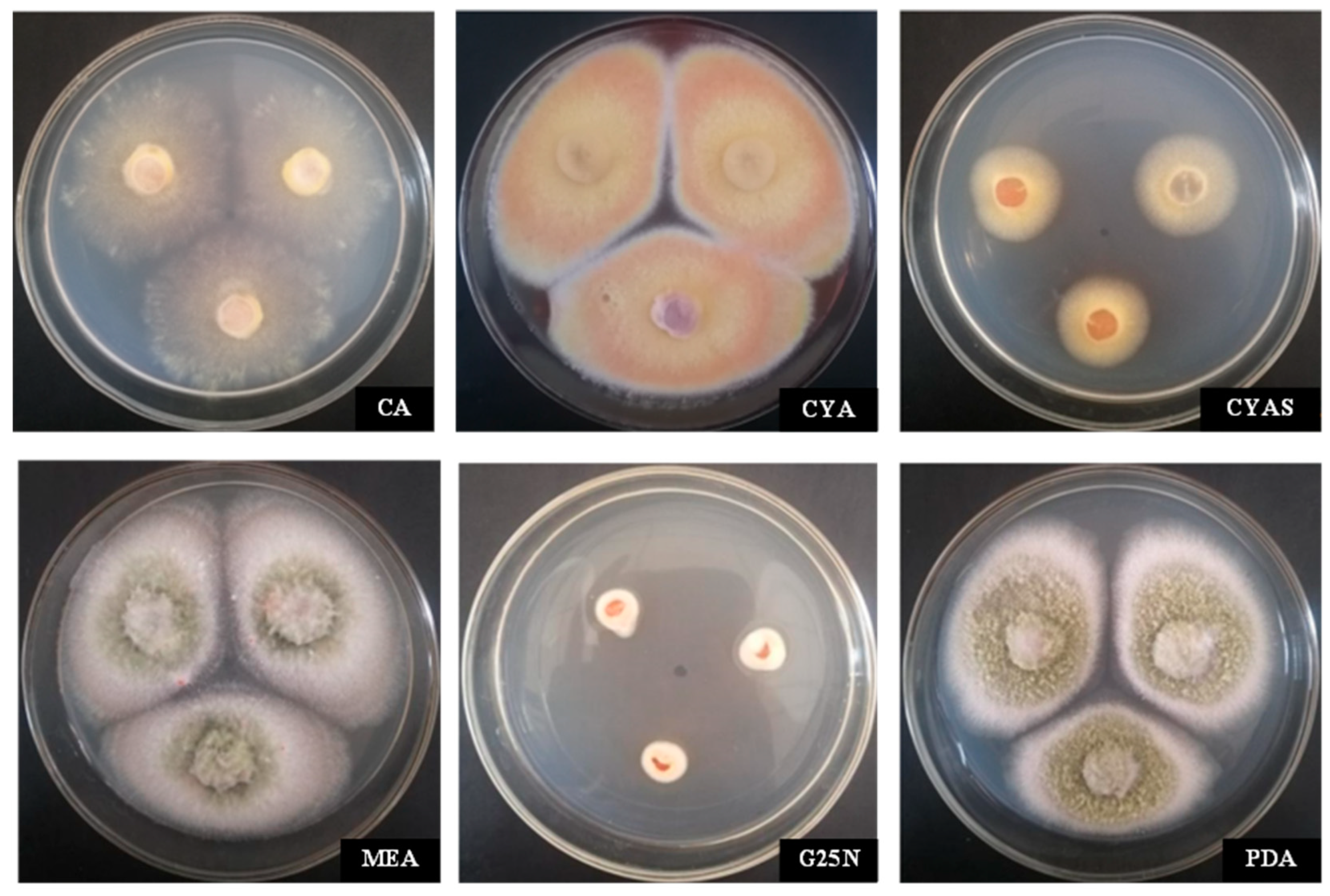
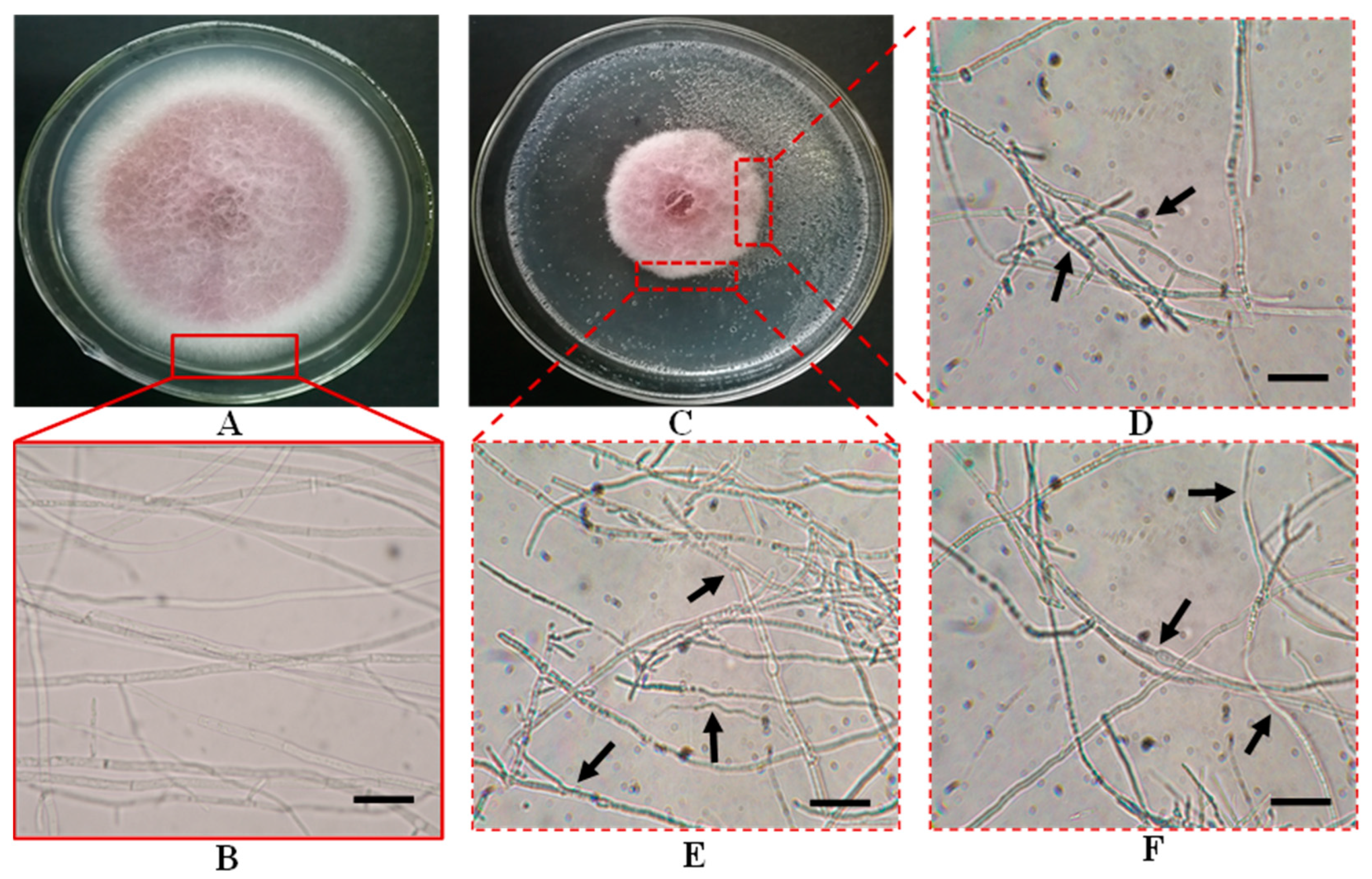
3.3.1. Identification of Culture Characteristics
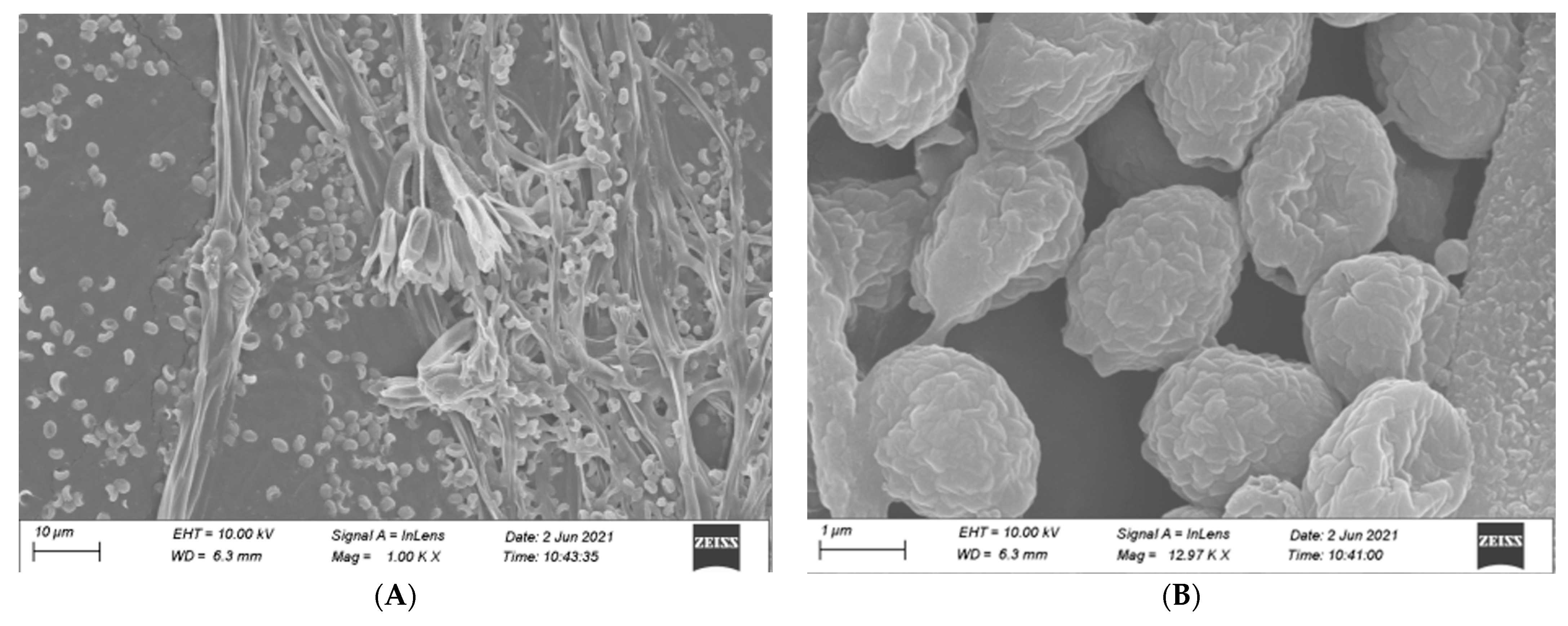
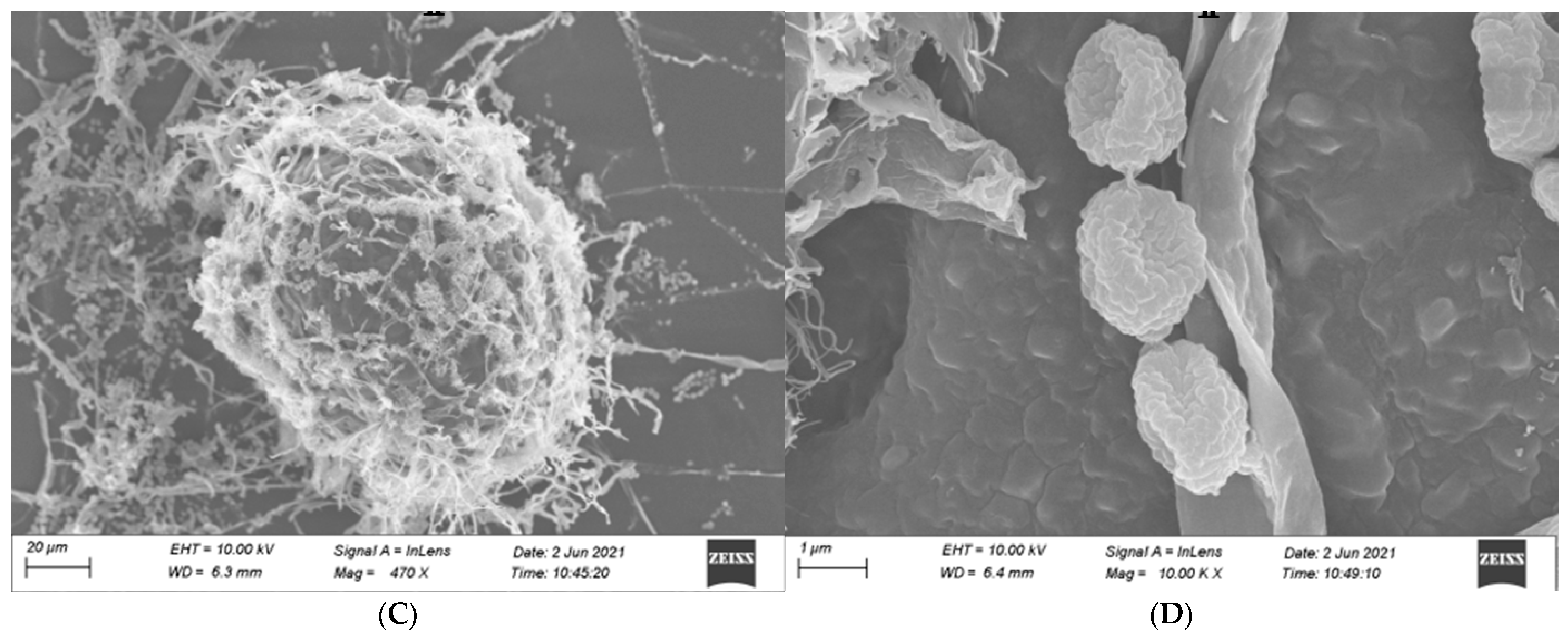
3.3.2. Identification of Microscopic Features
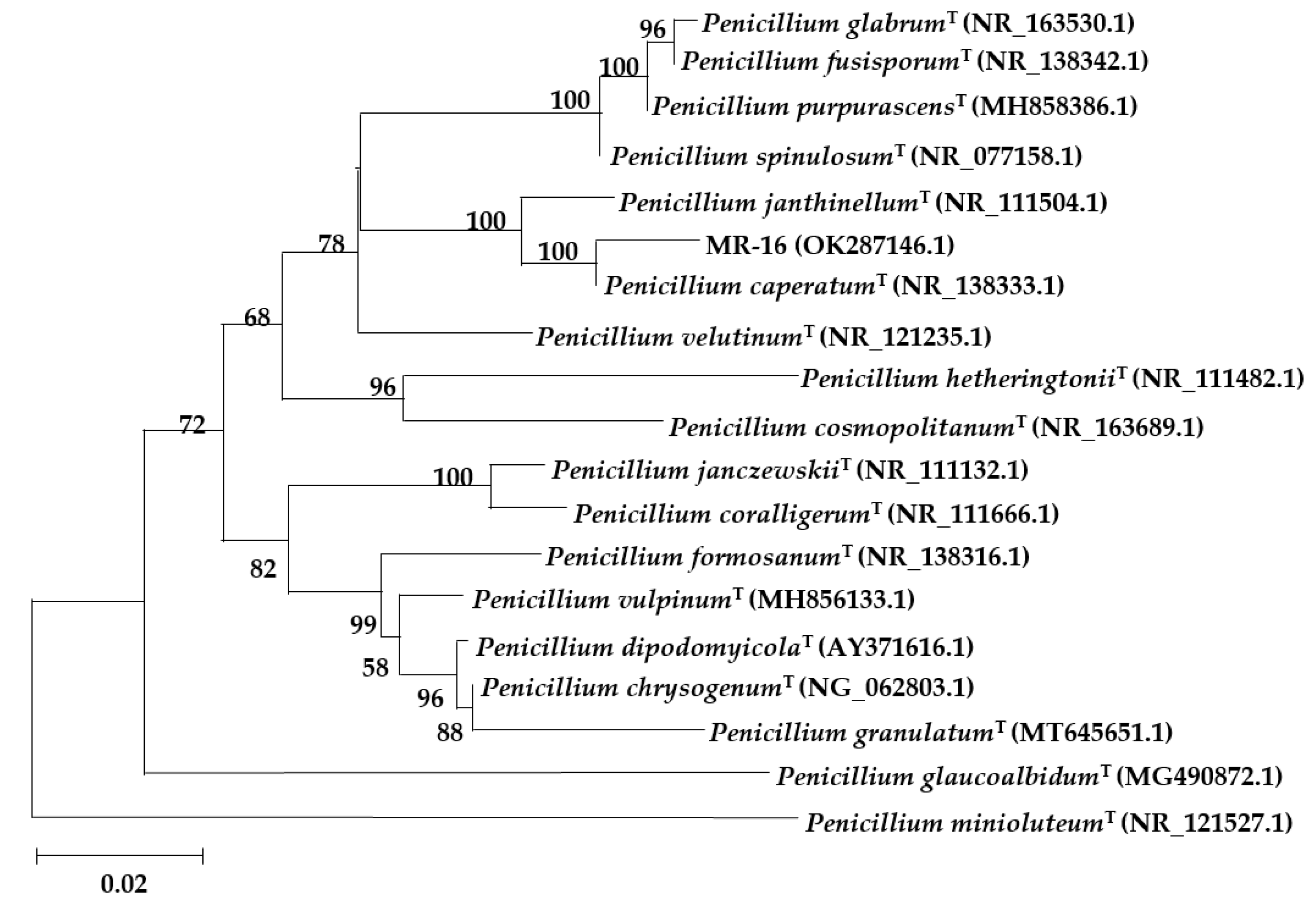
3.3.3. Molecular Identification
3.4. MR-16 Culture Filtrate Inhibition of F. oxysporum Mycelium
3.5. Culture Filtrate of MR-16 on Spore Germination of F. oxysporum in Co-Culture
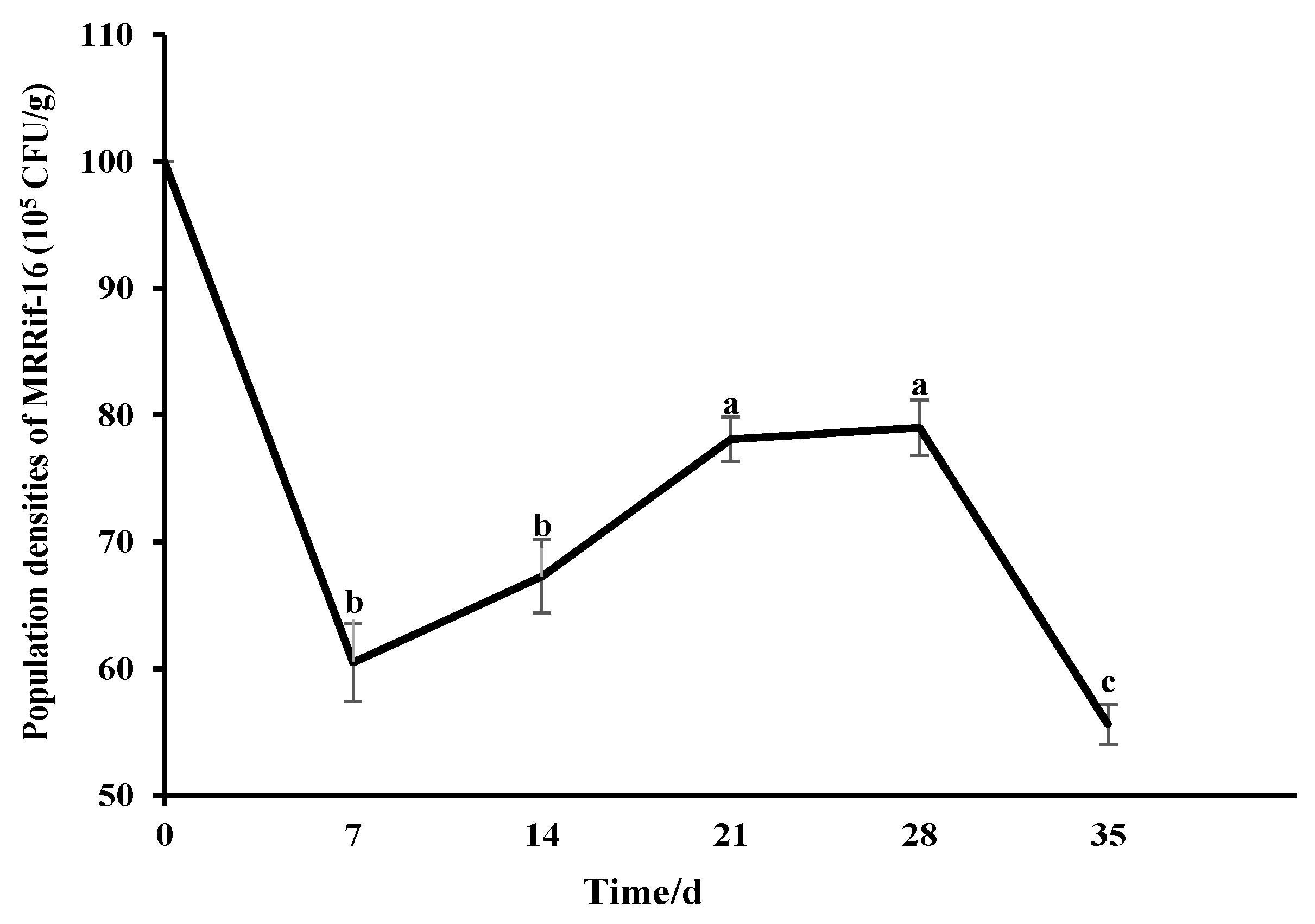
3.6. Soil Colonization Ability of MR-16 Isolate
3.7. Control Efficiency of MR-16 against Fusarium wilt of S. divaricata
3.8. The Growth-Promoting Effect of MR-16 on S. divaricata
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Asad, S.A. Mechanisms of action and biocontrol potential of Trichoderma against fungal plant diseases—A review. Ecol. Complex 2022, 49, 100978. [Google Scholar] [CrossRef]
- Michielse, C.B.; Rep, M. Pathogen profile update: Fusarium oxysporum. Mol. Plant Pathol. 2009, 10, 311–324. [Google Scholar] [CrossRef] [PubMed]
- National Pharmacopoeia Committee. Pharmacopoeia of the People’s Republic of China; China Medical Science Press: Beijing, China, 2020; Volume I, p. 156. ISBN 978-7-5067-7337-9. [Google Scholar]
- Kreiner, J.; Pang, E.; Lenon, G.B.; Yang, A.W.H. Saposhnikoviae divaricata: A phytochemical, pharmacological, and pharmacokinetic review. Chin. J. Nat. Med. 2017, 15, 255–264. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.M.; Jiang, H.; Dai, H.L.; Wang, Z.W.; Jia, G.Z.; Meng, X.C. Feeble antipyretic, analgesic, and anti-inflammatory activities were found with tegular dose 4′-O-beta-D-glucosyl-5-O-methylvisamminol, one of the conventional marker compounds for quality evaluation of Radix Saposhnikoviae. Pharmacogn. Mag. 2017, 13, 168–174. [Google Scholar]
- Ma, J.; Tian, Y.; Wang, Z.; Liu, G.; Wang, Z.; Zhang, M. Isolation and identification of endophytic antagonistic bacteria from Saposhnikovia divaricata. J. Jilin Agric. Univ. 2022, 33, 323–328. [Google Scholar]
- Guo, X.; Tian, X.; Hao, J.; Wang, Y.; Yang, L.; Han, M.; Han, Z. Relationships between Saposhnikovia divaricata chromone content and soil factors in different regions. J. S. China Agric. Univ. 2020, 41, 31–37. [Google Scholar]
- Zeng, L. Diseases and Insect Pests of Medicinal Plants; Guizhou Science and Technology Press: Guiyang, China, 2017; p. 222. [Google Scholar]
- Tleuova, A.B.; Wielogorska, E.; Talluri, V.; Stepanek, F.; Elliott, C.T.; Grigoriev, D.O. Recent advances and remaining barriers to producing novel formulations of fungicides for safe and sustainable agriculture. J. Control. Release 2020, 326, 468–481. [Google Scholar] [CrossRef]
- Gao, Y.Q.; Wang, Y.; Li, J.; Shang, S.B.; Song, Z.Q. Improved application of natural forest product terpene for discovery of potential botanical fungicide. Ind. Crop. Prod. 2018, 126, 103–112. [Google Scholar] [CrossRef]
- Sare, A.R.; Jijakli, M.H.; Massart, S. Microbial ecology to support integrative efficacy improvement of biocontrol agents for postharvest diseases management. Postharvest Biol. Technol. 2021, 179, 111572. [Google Scholar] [CrossRef]
- Bollmann-Giolai, A.; Malone, J.G.; Arora, S. Diversity, detection and exploitation: Linking soil fungi and plant disease. Curr. Opin. Microbiol. 2022, 70, 102199. [Google Scholar] [CrossRef]
- Samaras, A.; Karaoglanidis, G.S.; Tzelepis, G. Insights into the multitrophic interactions between the biocontrol agent Bacillus subtilis MBI 600, the pathogen Botrytis cinerea and their plant host. Microbiol. Res. 2021, 248, 126752. [Google Scholar] [CrossRef] [PubMed]
- Syed-Ab-Rahman, S.F.; Carvalhais, L.C.; Chua, E.; Xiao, Y.; Wass, T.J.; Schenk, P.M. Identification of soil bacterial isolates suppressing different phytophthora spp. and promoting plant growth. Front. Plant Sci. 2018, 9, 1502. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Yan, Y.; Wang, Y.; Wu, M.; Mao, Q.; Chen, Y.; Ren, J.; Liu, A.; Lin, X.; Ahammed, G.J. Trichoderma asperellum reduces phoxim residue in roots by promoting plant detoxification potential in Solanum lycopersicum L. Environ. Pollut. 2020, 259, 113893. [Google Scholar] [CrossRef] [PubMed]
- Wonglom, P.; Daengsuwan, W.; Ito, S.I.; Sunpapao, A. Biological control of Sclerotium fruit rot of snake fruit and stem rot of lettuce by Trichoderma sp. T76-12/2 and the mechanisms involved. Physiol. Mol. Plant Pathol. 2019, 107, 1–7. [Google Scholar] [CrossRef]
- Choi, H.W.; Ahsan, S.M. Biocontrol activity of Aspergillus terreus anu-301 against two distinct plant diseases, tomato fusarium wilt and potato soft rot. Plant Pathol. J. 2022, 38, 33–45. [Google Scholar] [CrossRef]
- Bunbury-Blanchette, A.L.; Walker, A.K. Trichoderma species show biocontrol potential in dual culture and greenhouse bioassays against Fusarium basal rot of onion. Biol. Control 2019, 130, 127–135. [Google Scholar] [CrossRef]
- Costa, D.; Tavares, R.M.; Baptista, P.; Lino-Neto, T. Cork oak endophytic fungi as potential biocontrol agents against Biscogniauxia mediterranea and Diplodia corticola. J. Fungi 2020, 6, 287. [Google Scholar] [CrossRef]
- Frisvad, J.C.; Samson, R.A. Polyphasic taxonomy of Penicillium subgenus Penicillium. A guide to identification of food and air-borne terverticillate Penicillia and their mycotoxins. Stud. Mycol. 2004, 49, 1–174. [Google Scholar]
- Visagie, C.M.; Renaud, J.B.; Burgess, K.M.; Malloch, D.W.; Clark, D.; Ketch, L.; Urb, M.; Louis-Seize, G.; Assabgui, R.; Sumarah, M.W.; et al. Fifteen new species of Penicillium. Persoonia 2016, 36, 247–280. [Google Scholar] [CrossRef]
- Han, Z.; Cui, Y.; Wang, Y.; Wang, Y.; Sun, Z.; Han, M.; Yang, L. Effect of rhizospheric fungus on biological control of root rot (Fusarium equiseti) disease of Saposhnikovia divaricata. Agronomy 2022, 12, 2906. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Millan, A.F.S.; Larraya, L.; Farran, I.; Ancin, M.; Veramendi, J. Successful biocontrol of major postharvest and soil-borne plant pathogenic fungi by antagonistic yeasts. Biol. Control 2021, 160, 104683. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhao, L.; Li, Z.; Li, C.; Li, B.; Gu, X.; Zhang, X.; Zhang, H. Screening and identification of an antagonistic yeast controlling postharvest blue mold decay of pears and the possible mechanisms involved. Biol. Control 2019, 133, 26–33. [Google Scholar] [CrossRef]
- Darma, S.; Ambara, A.; Aman, A.T.; Annisa, L.; Nurrokhman; Nuryastuti, T.; Wibawa, T. High frequency of azole resistant Candida spp. colonization among presumptive multidrug resistant tuberculosis (MDR-TB) patients. PLoS ONE 2020, 15, e242542. [Google Scholar] [CrossRef] [PubMed]
- Gholami, M.; Amini, J.; Abdollahzadeh, J.; Ashengroph, M. Basidiomycetes fungi as biocontrol agents against take-all disease of wheat. Biol. Control 2019, 130, 34–43. [Google Scholar] [CrossRef]
- Luo, M.; Chen, Y.; He, J.; Tang, X.; Wu, X.; Xu, C. Identification of a new Talaromyces strain DYM25 isolated from the Yap Trench as a biocontrol agent against Fusarium wilt of cucumber. Microbiol. Res. 2021, 251, 126841. [Google Scholar] [CrossRef]
- Mulero-Aparicio, A.; Agustí-Brisach, C.; Varo, Á.; López-Escudero, F.J.; Trapero, A. A non-pathogenic strain of Fusarium oxysporum as a potential biocontrol agent against Verticillium wilt of olive. Biol. Control 2019, 139, 104045. [Google Scholar] [CrossRef]
- Visagie, C.M.; Houbraken, J.; Frisvad, J.C.; Hong, S.B.; Klaassen, C.H.; Perrone, G.; Seifert, K.A.; Varga, J.; Yaguchi, T.; Samson, R.A. Identification and nomenclature of the genus Penicillium. Stud. Mycol. 2014, 78, 343–371. [Google Scholar] [CrossRef]
- Houbraken, J.; Samson, R.A. Phylogeny of Penicillium and the segregation of Trichocomaceae into three families. Stud. Mycol. 2011, 70, 1–51. [Google Scholar] [CrossRef]
- Kong, H. Flora fungorum sinicorum. In Penicillium et Teleomorphi Cognati; Science Press: Beijing, China, 2007; Volume 35, pp. 40–199. ISBN 978-7-0301-9262-2. [Google Scholar]
- Houbraken, J.; de Vries, R.P.; Samson, R.A. Modern taxonomy of biotechnologically important Aspergillus and Penicillium species. Adv. Appl. Microbiol. 2014, 86, 199–249. [Google Scholar] [CrossRef]
- Yu, L.; Qiao, N.; Zhao, J.; Zhang, H.; Tian, F.; Zhai, Q.; Chen, W. Postharvest control of Penicillium expansum in fruits: A review. Food Biosci. 2020, 36, 100633. [Google Scholar] [CrossRef]
- Radulesco, T.; Varoquaux, A.; Ranque, S.; Dessi, P.; Michel, J.; Cassagne, C. A Case of fungus ball-type maxillary sinusitis due to Penicillium roqueforti. Mycopathologia 2018, 183, 439–443. [Google Scholar] [CrossRef] [PubMed]
- Zakariya, N.A.; Majeed, S.; Jusof, W.H.W. Investigation of antioxidant and antibacterial activity of iron oxide nanoparticles (IONPS) synthesized from the aqueous extract of Penicillium spp. Sens. Int. 2022, 3, 100164. [Google Scholar] [CrossRef]
- Mareze, J.; Ramos-Pereira, J.; Santos, J.A.; Beloti, V.; López-Díaz, T.M. Identification and characterisation of lactobacilli isolated from an artisanal cheese with antifungal and antibacterial activity against cheese spoilage and mycotoxigenic Penicillium spp. Int. Dairy J. 2022, 130, 105367. [Google Scholar] [CrossRef]
- Patil, N.S.; Jadhav, J.P. Significance of Penicillium ochrochloron chitinase as a biocontrol agent against pest Helicoverpa armigera. Chemosphere 2015, 128, 231–235. [Google Scholar] [CrossRef]
- Murali, M.; Amruthesh, K.N. Plant growth-promoting fungus Penicillium oxalicum enhances plant growth and induces resistance in pearl millet against downy mildew disease. J. Phytopathol. 2015, 163, 743–754. [Google Scholar] [CrossRef]
- Din, G.; Hassan, A.; Dunlap, J.; Ripp, S.; Shah, A.A. Cadmium tolerance and bioremediation potential of filamentous fungus Penicillium chrysogenum FMS2 isolated from soil. Int. J. Environ. Sci. Technol. 2022, 19, 2761–2770. [Google Scholar] [CrossRef]
- Sharma, B.; Tiwari, S.; Bisht, N.; Tewari, L. Eco-friendly bioprocess using agar plug immobilized Penicillium crustosum PWWS-6 biomass for treatment of wastewater contaminated with toxic Congo red dye for use in agriculture. Ind. Crop. Prod. 2021, 170, 113755. [Google Scholar] [CrossRef]
- Urooj, F.; Farhat, H.; Tariq, A.; Moin, S.; Sohail, N.; Sultana, V.; Hameedi, S.F.; Shams, Z.I.; Ehteshamul-Haque, S. Role of endophytic Penicillium species and Pseudomonas monteilii in inducing the systemic resistance in okra against root rotting fungi and their effect on some physiochemical properties of okra fruit. J. Appl. Microbiol. 2021, 130, 604–616. [Google Scholar] [CrossRef]
- Udagawa, S.; Horie, Y. Some Eupenicillium from soils of New Guinea. Trans. Mycol. Soc. Jpn. 1973, 14, 370–387. [Google Scholar]
- Kepler, R.M.; Maul, J.E.; Rehner, S.A. Managing the plant microbiome for biocontrol fungi: Examples from Hypocreales. Curr. Opin. Microbiol. 2017, 37, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Wei, X.L.; Xue, L.; Zhang, Z.F.; Zhang, P. Antimicrobial meroterpenoids and erythritol derivatives isolated from the marine-algal-derived endophytic fungus Penicillium chrysogenum XNM-12. Mar. Drugs 2020, 18, 578. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Hou, M.; Chang, Z.; Zhang, J.; Xu, Y. Purification, identification and bioactivity of antagonistic chemicals produced by biocontrol agents of Penicillium striatisporum Pst10. Jiangsu J. Agric. Sci. 2013, 5, 1011–1018. [Google Scholar]
- Ting, A.S.Y.; Mah, S.W.; Tee, C.S. Evaluating the feasibility of induced host resistance by endophytic isolate Penicillium citrinum BTF08 as a control mechanism for Fusarium wilt in banana plantlets. Biol. Control 2012, 61, 155–159. [Google Scholar] [CrossRef]
- Xian, H.Q.; Liu, L.; Li, Y.H.; Yang, Y.N.; Yang, S. Molecular tagging of biocontrol fungus Trichoderma asperellum and its colonization in soil. J. Appl. Microbiol. 2020, 128, 255–264. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Chang, Z.; Zhao, J.; Zhou, M. Antifungal activity of Penicillium striatisporum Pst10 and its biocontrol effect on Phytophthora root rot of chilli pepper. Biol. Control 2008, 44, 24–31. [Google Scholar] [CrossRef]
- Jiang, Z.; Guo, J. Hazard analysis of the impact of biocontrolling microbes on soil micro-ecosystem. J. Microbiol. 2006, 26, 85–88. [Google Scholar] [CrossRef]
- Ma, C.; Gong, X.; Gao, H.; Wu, J.; Li, D.; Chen, X.; Li, H.; Liu, M. Legacy impacts on the relationships between soil microbial community and the invasion potential of non-indigenous bacteria. Acta Ecol. Sin. 2019, 39, 1–9. [Google Scholar] [CrossRef]
- Hartmann, A.; Schmid, M.; Tuinen, D.V.; Berg, G. Plant-driven selection of microbes. Plant Soil 2009, 321, 235–257. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Inhibition Rate (%) | Inhibition Zone (mm) |
|---|---|---|
| MR-16 | 66.67 ± 2.22 a | 10.05 ± 0.19 a |
| MR-82 | 63.70 ± 1.28 b | 9.00 ± 0.19 bc |
| MR-87 | 62.59 ± 1.70 bc | 4.71 ± 0.32 h |
| MR-32 | 61.35 ± 0.92 bcd | 8.53 ± 0.31 c |
| MR-52 | 61.10 ± 0.78 bcde | 6.10 ± 0.40 f |
| MR-73 | 60.12 ± 0.82 cdef | 6.46 ± 0.26 ef |
| MR-43 | 59.63 ± 1.73 cdef | 7.42 ± 0.18 d |
| MR-39 | 58.89 ± 2.94 defg | 6.69 ± 0.23 e |
| MR-59 | 58.15 ± 0.64 efg | 8.95 ± 0.28 bc |
| MR-84 | 57.83 ± 0.30 fg | 9.20 ± 0.13 b |
| MR-96 | 57.41 ± 1.28 fg | 4.29 ± 0.32 h |
| MR-38 | 56.30 ± 2.31 g | 5.36 ± 0.35 g |
| Plant Pathogen | Inhibition Rate (%) 1 |
|---|---|
| Phytophthora cactorum | 80.00 ± 1.28 a |
| Cylindrocarpon destructans | 78.88 ± 0.64 ab |
| Mycocentrospora acerina | 77.11 ± 0.56 b |
| Alternaria liriodendron | 73.77 ± 0.93 c |
| Alternaria tenuissima | 72.22 ± 0.64 c |
| Fusarium solani | 71.88 ± 0.21 c |
| Fusarium equiseti | 68.22 ± 0.42 d |
| Botrytis cinerea | 68.14 ± 1.28 d |
| Rhizoctonia solani | 54.44 ± 1.11 e |
| Treatment | Spore Germination Rate (%) 1 | Inhibition Rate at 48 h (%) | |||
|---|---|---|---|---|---|
| 6 h | 12 h | 24 h | 48 h | ||
| CK | 2.70 ± 0.35 a | 9.56 ± 3.34 a | 22.83 ± 3.01 a | 59.7 ± 5.97 a | — |
| MR-16 | 1.1 ± 0.4 b | 1.69 ± 0.23 b | 7.4 ± 0.37 b | 9.04 ± 2.68 b | 84.88 ± 3.94 |
| Treatment 2 | Disease Severity 1 | Disease Control (%) 1 |
|---|---|---|
| A | 19.85 ± 2.06 b | 60.76 ± 4.08 a |
| B | 16.00 ± 4.23 b | 68.38 ± 8.36 a |
| C | 22.18 ± 3.72 b | 56.16 ± 7.35 a |
| D | 19.59 ± 2.28 b | 61.28 ± 4.51 a |
| CK | 50.59 ± 4.29 a | — |
| Treatment 1 | Plant Height/cm | Root Length/cm | Plant Fresh Biomass/g | Root Fresh Biomass/g | Plan Dry Biomass/g | Root Dry Biomass/g |
|---|---|---|---|---|---|---|
| A | 56.30 ± 4.47 a | 29.87 ± 3.34 a | 9.85 ± 0.49 a | 3.98 ± 1.03 a | 2.70 ± 0.67 a | 1.42 ± 0.28 a |
| B | 51.63 ± 3.58 b | 29.34 ± 2.15 a | 7.66 ± 1.62 b | 3.04 ± 0.47 b | 2.49 ± 0.97 a | 1.04 ± 0.30 b |
| C | 51.35 ± 4.14 b | 28.63 ± 2.84 a | 7.89 ± 1.98 b | 3.10 ± 0.39 b | 2.54 ± 1.01 a | 1.06 ± 0.25 b |
| CK | 50.99 ± 4.57 b | 28.25 ± 2.43 a | 6.62 ± 1.73 c | 2.33 ± 0.79 c | 1.88 ± 0.57 b | 0.82 ± 0.19 c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, Z.; Wang, J.; Ding, Y.; Sun, Z.; Wang, Y.; Wang, Y.; Yang, L.; Wang, Y. New Observation in Biocontrol of Penicillium caperatum against Fusarium oxysporum on Saposhnikovia divaricata and as a Plant Growth Promoter. Fermentation 2023, 9, 361. https://doi.org/10.3390/fermentation9040361
Han Z, Wang J, Ding Y, Sun Z, Wang Y, Wang Y, Yang L, Wang Y. New Observation in Biocontrol of Penicillium caperatum against Fusarium oxysporum on Saposhnikovia divaricata and as a Plant Growth Promoter. Fermentation. 2023; 9(4):361. https://doi.org/10.3390/fermentation9040361
Chicago/Turabian StyleHan, Zhongming, Jiao Wang, Yanzhe Ding, Zhuo Sun, Yan Wang, Yuyi Wang, Limin Yang, and Yunhe Wang. 2023. "New Observation in Biocontrol of Penicillium caperatum against Fusarium oxysporum on Saposhnikovia divaricata and as a Plant Growth Promoter" Fermentation 9, no. 4: 361. https://doi.org/10.3390/fermentation9040361
APA StyleHan, Z., Wang, J., Ding, Y., Sun, Z., Wang, Y., Wang, Y., Yang, L., & Wang, Y. (2023). New Observation in Biocontrol of Penicillium caperatum against Fusarium oxysporum on Saposhnikovia divaricata and as a Plant Growth Promoter. Fermentation, 9(4), 361. https://doi.org/10.3390/fermentation9040361