
Pigment Production of Chlamydomonas Strains in Response to Norflurazon and ZnO Nanoparticles


Abstract
1. Introduction
2. Materials and Methods
2.1. Algal Strains and Culture Conditions
2.2. Growth and Cell Morphology
2.3. Cell Viability Assay
2.4. Photosynthetic Activity and Pigment Content
2.5. Statistical Analysis
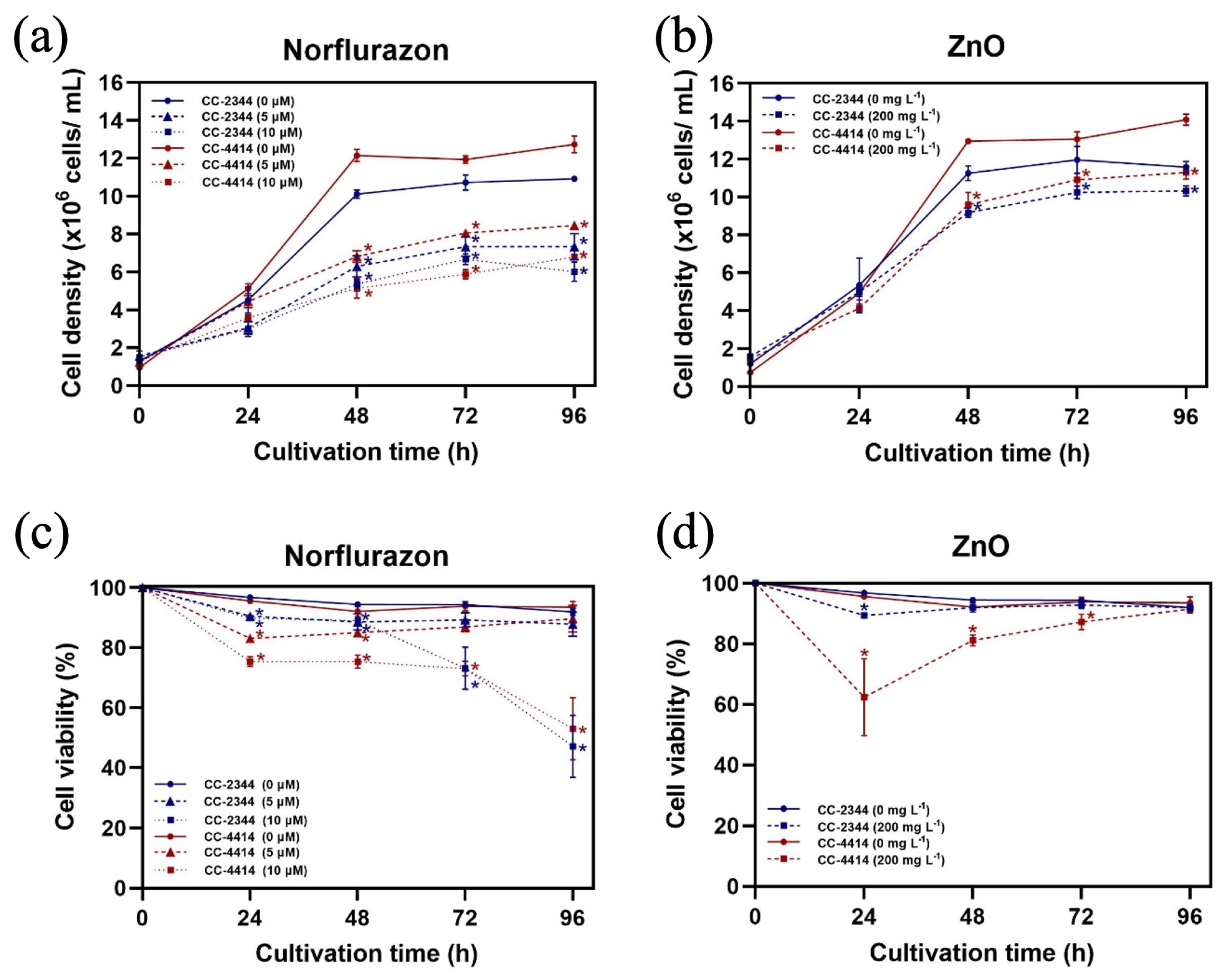
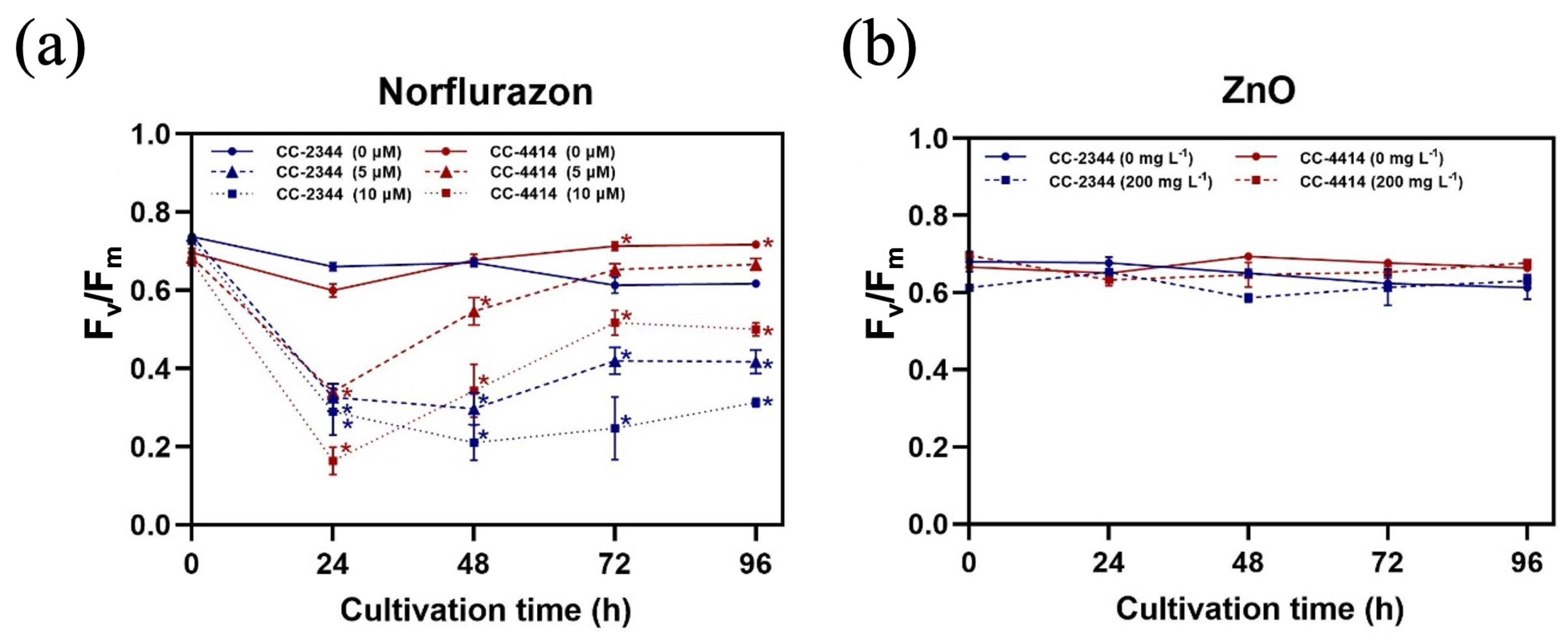
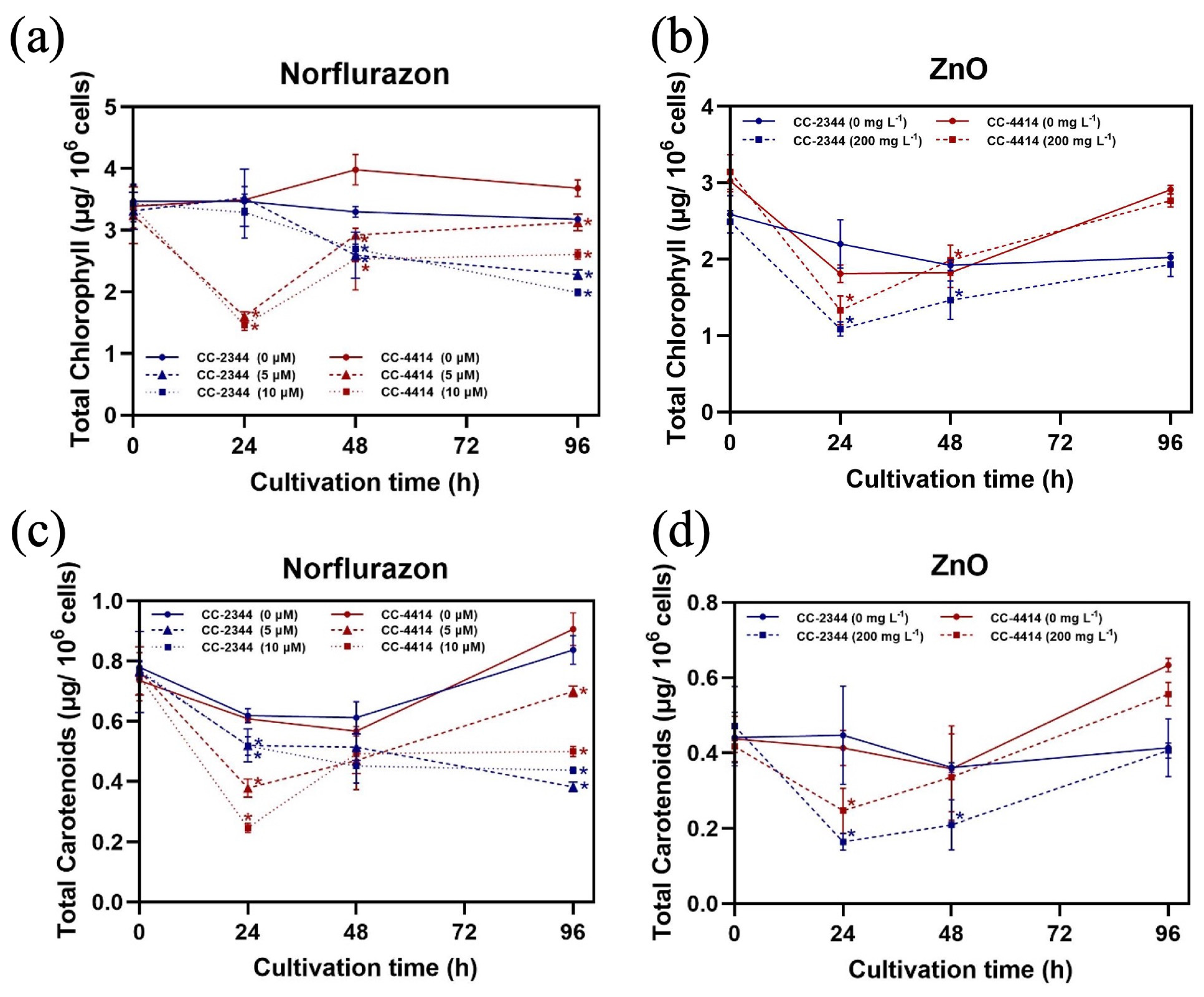
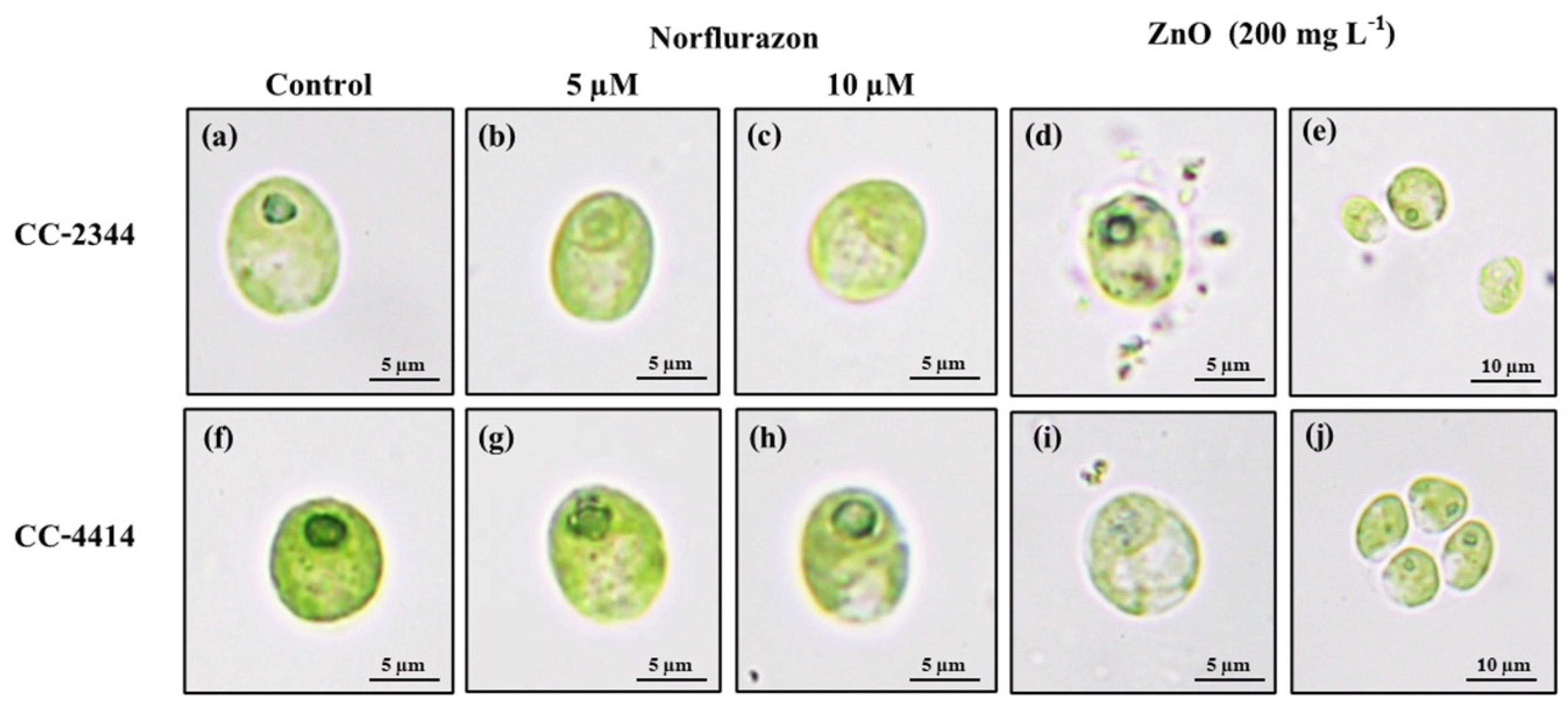
3. Results
3.1. Algal Growth and Responses to Chemicals
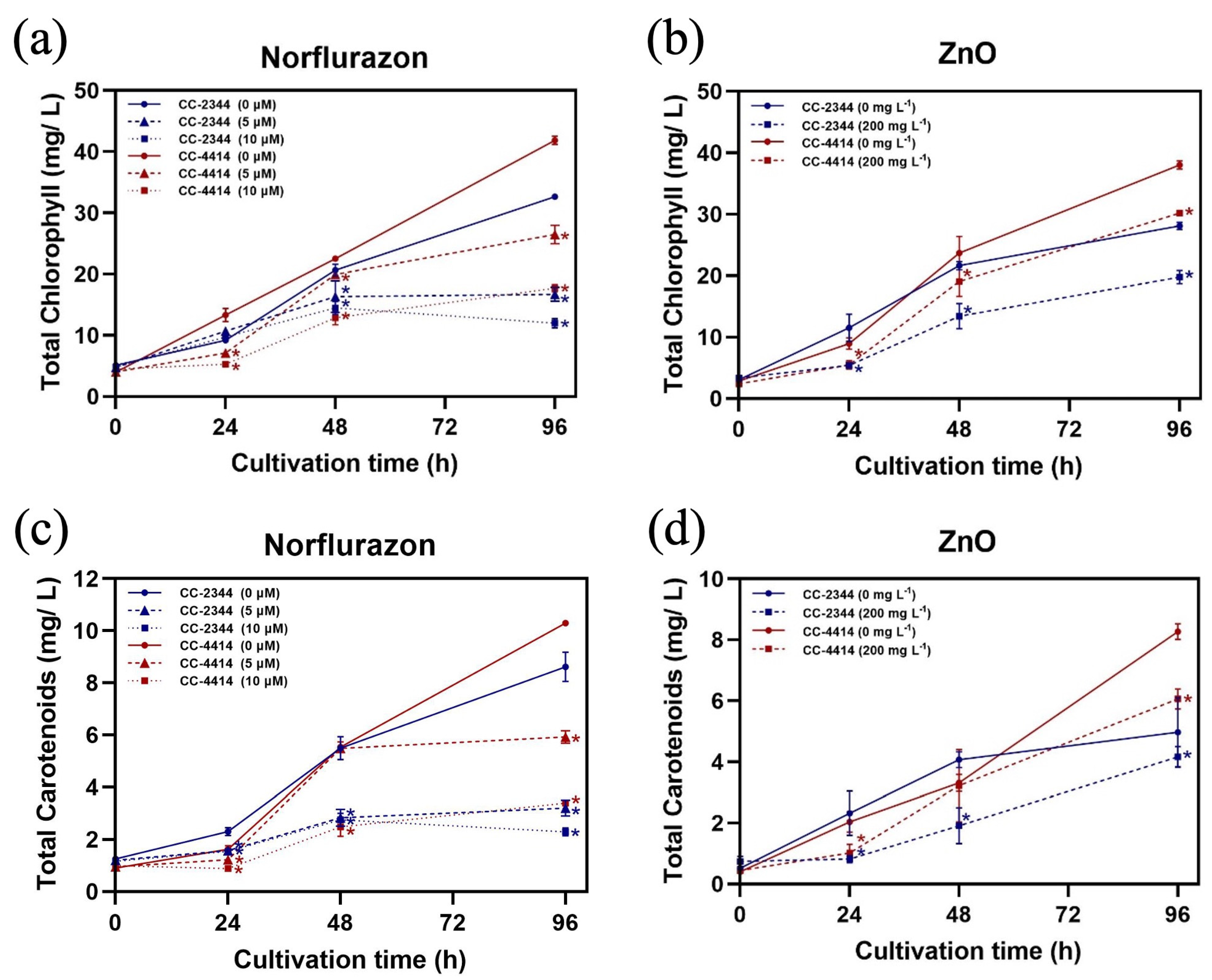
3.2. Photosynthesis and Pigment Content
3.3. Cell Morphology
3.4. Pigment Production
4. Discussion
4.1. Effects of Norflurazon
4.2. Effects of ZnO NPs
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Andersen, R. Diversity of eukaryotic algae. Biodivers. Conserv. 1992, 1, 267–292. [Google Scholar] [CrossRef]
- Hamed, I. The evolution and versatility of microalgal biotechnology: A review. Compr. Rev. Food Sci. Food Saf. 2016, 15, 1104–1123. [Google Scholar] [CrossRef] [PubMed]
- Sharma, N.K.; Rai, A.K. Biodiversity and biogeography of microalgae: Progress and pitfalls. Environ. Rev. 2011, 19, 1–15. [Google Scholar] [CrossRef]
- Jin, E.S.; Melis, A. Microalgal biotechnology: Carotenoid production by the green algaeDunaliella salina. Biotechnol. Bioprocess Eng. 2003, 8, 331–337. [Google Scholar] [CrossRef]
- Das, P.; Aziz, S.S.; Obbard, J.P. Two phase microalgae growth in the open system for enhanced lipid productivity. Renew. Energy 2011, 36, 2524–2528. [Google Scholar] [CrossRef]
- Barsanti, L.; Coltelli, P.; Evangelista, V.; Frassanito, A.M.; Passarelli, V.; Vesentini, N.; Gualtieri, P. Oddities and curiosities in the algal world. In Algal Toxins: Nature, Occurrence, Effect and Detection; Springer: Berlin/Heidelberg, Germany, 2008; pp. 353–391. [Google Scholar]
- Corcoran, E. Sick Water?: The Central Role of Wastewater Management in Sustainable Development: A Rapid Response Assessment; UNEP/Earthprint: Nairobi, Kenya, 2010. [Google Scholar]
- Boger, P. Carotenoid biosynthesis inhibitor herbicides-mode of action and resistance mechanism. Pestic. Outlook 1998, 9, 29–35. [Google Scholar]
- Park, J.-H.; Tran, L.H.; Jung, S. Perturbations in the photosynthetic pigment status result in photooxidation-induced crosstalk between carotenoid and porphyrin biosynthetic pathways. Front. Plant Sci. 2017, 8, 1992. [Google Scholar] [CrossRef]
- Guardini, Z.; Dall’Osto, L.; Barera, S.; Jaberi, M.; Cazzaniga, S.; Vitulo, N.; Bassi, R. High carotenoid mutants of Chlorella vulgaris show enhanced biomass yield under high irradiance. Plants 2021, 10, 911. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Gerken, H.; Huang, J.; Chen, F. Engineering of an endogenous phytoene desaturase gene as a dominant selectable marker for Chlamydomonas reinhardtii transformation and enhanced biosynthesis of carotenoids. Process Biochem. 2013, 48, 788–795. [Google Scholar] [CrossRef]
- Trovão, M.; Schüler, L.M.; Machado, A.; Bombo, G.; Navalho, S.; Barros, A.; Pereira, H.; Silva, J.; Freitas, F.; Varela, J. Random mutagenesis as a promising tool for microalgal strain improvement towards industrial production. Mar. Drugs 2022, 20, 440. [Google Scholar] [CrossRef]
- Dastjerdi, R.; Montazer, M. A review on the application of inorganic nano-structured materials in the modification of textiles: Focus on anti-microbial properties. Colloids Surf. B Biointerfaces 2010, 79, 5–18. [Google Scholar] [CrossRef]
- Osmond, M.J.; Mccall, M.J. Zinc oxide nanoparticles in modern sunscreens: An analysis of potential exposure and hazard. Nanotoxicology 2010, 4, 15–41. [Google Scholar] [CrossRef] [PubMed]
- Gunawan, C.; Sirimanoonphan, A.; Teoh, W.Y.; Marquis, C.P.; Amal, R. Submicron and nano formulations of titanium dioxide and zinc oxide stimulate unique cellular toxicological responses in the green microalga Chlamydomonas reinhardtii. J. Hazard. Mater. 2013, 260, 984–992. [Google Scholar] [CrossRef] [PubMed]
- Prach, M.; Stone, V.; Proudfoot, L. Zinc oxide nanoparticles and monocytes: Impact of size, charge and solubility on activation status. Toxicol. Appl. Pharmacol. 2013, 266, 19–26. [Google Scholar] [CrossRef]
- Song, W.; Zhang, J.; Guo, J.; Zhang, J.; Ding, F.; Li, L.; Sun, Z. Role of the dissolved zinc ion and reactive oxygen species in cytotoxicity of ZnO nanoparticles. Toxicol. Lett. 2010, 199, 389–397. [Google Scholar] [CrossRef]
- Daughton, C.G. Non-regulated water contaminants: Emerging research. Environ. Impact Assess. Rev. 2004, 24, 711–732. [Google Scholar] [CrossRef]
- Kim, H.; Hong, Y.; Ahn, J.H. A study on the management of micropollutants in water system considering climate change and other potential effects. Korean Chem. Eng. Res. 2013, 51, 645–654. [Google Scholar] [CrossRef]
- Harris, E.H. Chlamydomonas as a model organism. Annu. Rev. Plant Biol. 2001, 52, 363–406. [Google Scholar] [CrossRef] [PubMed]
- Sasso, S.; Stibor, H.; Mittag, M.; Grossman, A.R. From molecular manipulation of domesticated Chlamydomonas reinhardtii to survival in nature. eLife 2018, 7, e39233. [Google Scholar] [CrossRef] [PubMed]
- Brennan, L.; Owende, P. Biofuels from microalgae—A review of technologies for production, processing, and extractions of biofuels and co-products. Renew. Sustain. Energy Rev. 2010, 14, 557–577. [Google Scholar] [CrossRef]
- Gallaher, S.D.; Fitz-Gibbon, S.T.; Glaesener, A.G.; Pellegrini, M.; Merchant, S.S. Chlamydomonas genome resource for laboratory strains reveals a mosaic of sequence variation, identifies true strain histories, and enables strain-specific studies. Plant Cell 2015, 27, 2335–2352. [Google Scholar] [CrossRef]
- Flowers, J.M.; Hazzouri, K.M.; Pham, G.M.; Rosas, U.; Bahmani, T.; Khraiwesh, B.; Nelson, D.R.; Jijakli, K.; Abdrabu, R.; Harris, E.H. Whole-genome resequencing reveals extensive natural variation in the model green alga Chlamydomonas reinhardtii. Plant Cell 2015, 27, 2353–2369. [Google Scholar] [CrossRef] [PubMed]
- Harris, E.H. Chlamydomonas Sourcebook; Academic Press: San Diego, CA, USA, 1989; Volume 2. [Google Scholar]
- Pérez-Martín, M.; Pérez-Pérez, M.E.; Lemaire, S.D.; Crespo, J.L. Oxidative stress contributes to autophagy induction in response to endoplasmic reticulum stress in Chlamydomonas reinhardtii. Plant Physiol. 2014, 166, 997–1008. [Google Scholar] [CrossRef] [PubMed]
- Delledonne, M.; Zeier, J.; Marocco, A.; Lamb, C. Signal interactions between nitric oxide and reactive oxygen intermediates in the plant hypersensitive disease resistance response. Proc. Natl. Acad. Sci. USA 2001, 98, 13454–13459. [Google Scholar] [CrossRef]
- Sirisha, V.; Sinha, M.; D’Souza, J.S. Menadione-induced caspase-dependent programmed cell death in the green chlorophyte C hlamydomonas reinhardtii. J. Phycol. 2014, 50, 587–601. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. [34] Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1987; Volume 148, pp. 350–382. [Google Scholar]
- MELIS, A.; Spangfort, M.; Andersson, B. Light-absorption and electron-transport balance between photosystem II and photosystem I in spinach chloroplasts. Photochem. Photobiol. 1987, 45, 129–136. [Google Scholar] [CrossRef]
- Yogesh Taparia, A.Z.; Leu, S.; Zarivach, R.; Boussiba, S.; Khozin-Goldberg, I. A novel endogenous selection marker for the diatom Phaeodactylum tricornutumbased on a unique mutation in phytoene desaturase. Sci. Rep. 2019, 9, 8271. [Google Scholar]
- Zhekisheva, M.; Zarka, A.; Khozin-Goldberg, I.; Cohen, Z.; Boussiba, S. Inhibition of astaxanthin synthesis under high irradiance does not abolish triacylglycerol accumulation in the green alga haematococcus pluvialis (Chlorophyceae) 1. J. Phycol. 2005, 41, 819–826. [Google Scholar] [CrossRef]
- Kitajima, K.; Hogan, K.P. Increases of chlorophyll a/b ratios during acclimation of tropical woody seedlings to nitrogen limitation and high light. Plant Cell Environ. 2003, 26, 857–865. [Google Scholar] [CrossRef] [PubMed]
- Cardona, T.; Shao, S.; Nixon, P.J. Enhancing photosynthesis in plants: The light reactions. Essays Biochem. 2018, 62, 85–94. [Google Scholar]
- Genevière, A.-M.; Derelle, E.; Escande, M.-L.; Grimsley, N.; Klopp, C.; Ménager, C.; Michel, A.; Moreau, H. Responses to iron oxide and zinc oxide nanoparticles in echinoderm embryos and microalgae: Uptake, growth, morphology, and transcriptomic analysis. Nanotoxicology 2020, 14, 1342–1361. [Google Scholar] [CrossRef]
- Suman, T.; Rajasree, S.R.; Kirubagaran, R. Evaluation of zinc oxide nanoparticles toxicity on marine algae Chlorella vulgaris through flow cytometric, cytotoxicity and oxidative stress analysis. Ecotoxicol. Environ. Saf. 2015, 113, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Bhuvaneshwari, M.; Iswarya, V.; Archanaa, S.; Madhu, G.; Kumar, G.S.; Nagarajan, R.; Chandrasekaran, N.; Mukherjee, A. Cytotoxicity of ZnO NPs towards fresh water algae Scenedesmus obliquus at low exposure concentrations in UV-C, visible and dark conditions. Aquat. Toxicol. 2015, 162, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Djearamane, S.; Lim, Y.M.; Wong, L.S.; Lee, P.F. Cytotoxic effects of zinc oxide nanoparticles on cyanobacterium Spirulina (Arthrospira) platensis. PeerJ 2018, 6, e4682. [Google Scholar] [CrossRef] [PubMed]
- De Almeida, A.C.O.; dos Santos, L.F.; Vicentini, D.S.; Matias, W.G.; Melegari, S.P. Evaluation of toxicity of zinc oxide nanorods on green microalgae of freshwater and marine ecosystems. Environ. Chem. Ecotoxicol. 2021, 3, 85–90. [Google Scholar] [CrossRef]
- Lee, W.-M.; An, Y.-J. Effects of zinc oxide and titanium dioxide nanoparticles on green algae under visible, UVA, and UVB irradiations: No evidence of enhanced algal toxicity under UV pre-irradiation. Chemosphere 2013, 91, 536–544. [Google Scholar] [CrossRef] [PubMed]
- Djearamane, S.; Lim, Y.M.; Wong, L.S.; Lee, P.F. Cellular accumulation and cytotoxic effects of zinc oxide nanoparticles in microalga Haematococcus pluvialis. PeerJ 2019, 7, e7582. [Google Scholar] [CrossRef]
- Chen, P.; Powell, B.A.; Mortimer, M.; Ke, P.C. Adaptive interactions between zinc oxide nanoparticles and Chlorella sp. Environ. Sci. Technol. 2012, 46, 12178–12185. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | Norflurazon (µM) | ZnO (mg L−1) | ||
|---|---|---|---|---|
| 0 | 5 | 10 | 200 | |
| Total Chlorophyll (mg L−1) | 30.37 ± 0.73 | 16.69 ± 1.13 * | 11.98 ± 0.77 * | 19.76 ± 1.11 * |
| Chlorophyll a (mg L−1) | 18.44 ± 0.54 | 11.73 ± 0.78 * | 8.63 ± 0.57 * | 12.47 ± 0.55 * |
| Chlorophyll b (mg L−1) | 11.71 ± 0.31 | 4.73 ± 0.33 * | 3.20 ± 0.19 * | 7.28 ± 0.59 * |
| Carotenoids (mg L−1) | 6.34 ± 0.41 | 3.21 ± 0.30 * | 2.30 ± 0.16 * | 4.16 ± 0.34 * |
| Control | Norflurazon (µM) | ZnO (mg L−1) | ||
|---|---|---|---|---|
| 0 | 5 | 10 | 200 | |
| Total Chlorophyll (mg L−1) | 39.98 ± 0.69 | 26.49 ± 1.51 * | 17.70 ± 0.61 * | 30.19 ± 0.43 * |
| Chlorophyll a (mg L−1) | 26.94 ± 0.39 | 19.13 ± 1.26 * | 12.70 ± 0.39 * | 19.06 ± 0.44* |
| Chlorophyll b (mg L−1) | 12.72 ± 0.31 | 7.02 ± 0.39 * | 4.77 ± 0.21 * | 11.13 ± 0.05 * |
| Carotenoid (mg L−1) | 9.28 ± 0.18 | 5.92 ± 0.24 * | 3.39 ± 0.11 * | 6.06 ± 0.32 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Intha, T.; Sirikhachornkit, A. Pigment Production of Chlamydomonas Strains in Response to Norflurazon and ZnO Nanoparticles. Fermentation 2023, 9, 193. https://doi.org/10.3390/fermentation9020193
Intha T, Sirikhachornkit A. Pigment Production of Chlamydomonas Strains in Response to Norflurazon and ZnO Nanoparticles. Fermentation. 2023; 9(2):193. https://doi.org/10.3390/fermentation9020193
Chicago/Turabian StyleIntha, Thanaporn, and Anchalee Sirikhachornkit. 2023. "Pigment Production of Chlamydomonas Strains in Response to Norflurazon and ZnO Nanoparticles" Fermentation 9, no. 2: 193. https://doi.org/10.3390/fermentation9020193
APA StyleIntha, T., & Sirikhachornkit, A. (2023). Pigment Production of Chlamydomonas Strains in Response to Norflurazon and ZnO Nanoparticles. Fermentation, 9(2), 193. https://doi.org/10.3390/fermentation9020193