

Biological Potential of Extremophilic Filamentous Fungi for the Production of New Compounds with Antimicrobial Effect

 ,
,  , ,
, ,
Abstract
1. Introduction
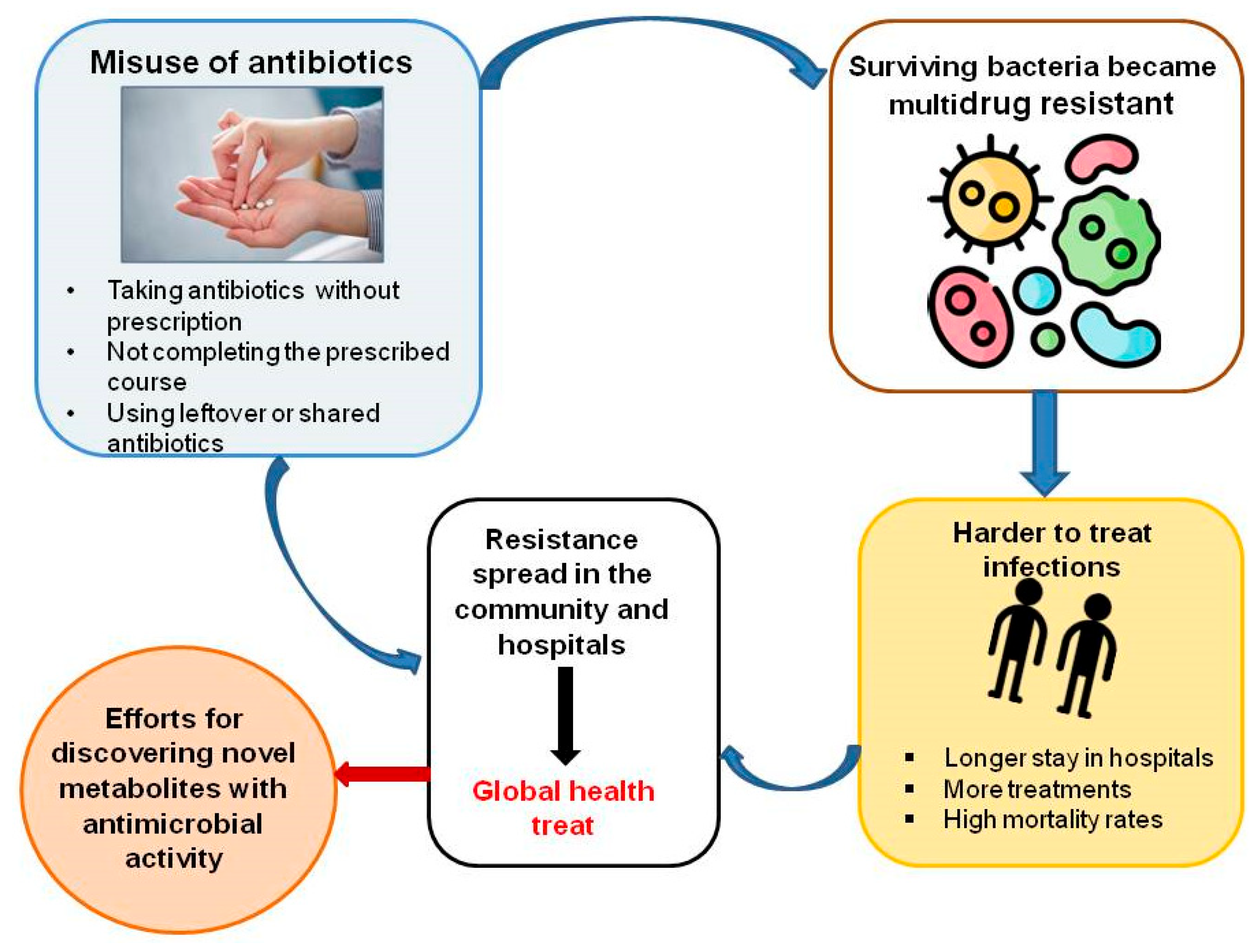
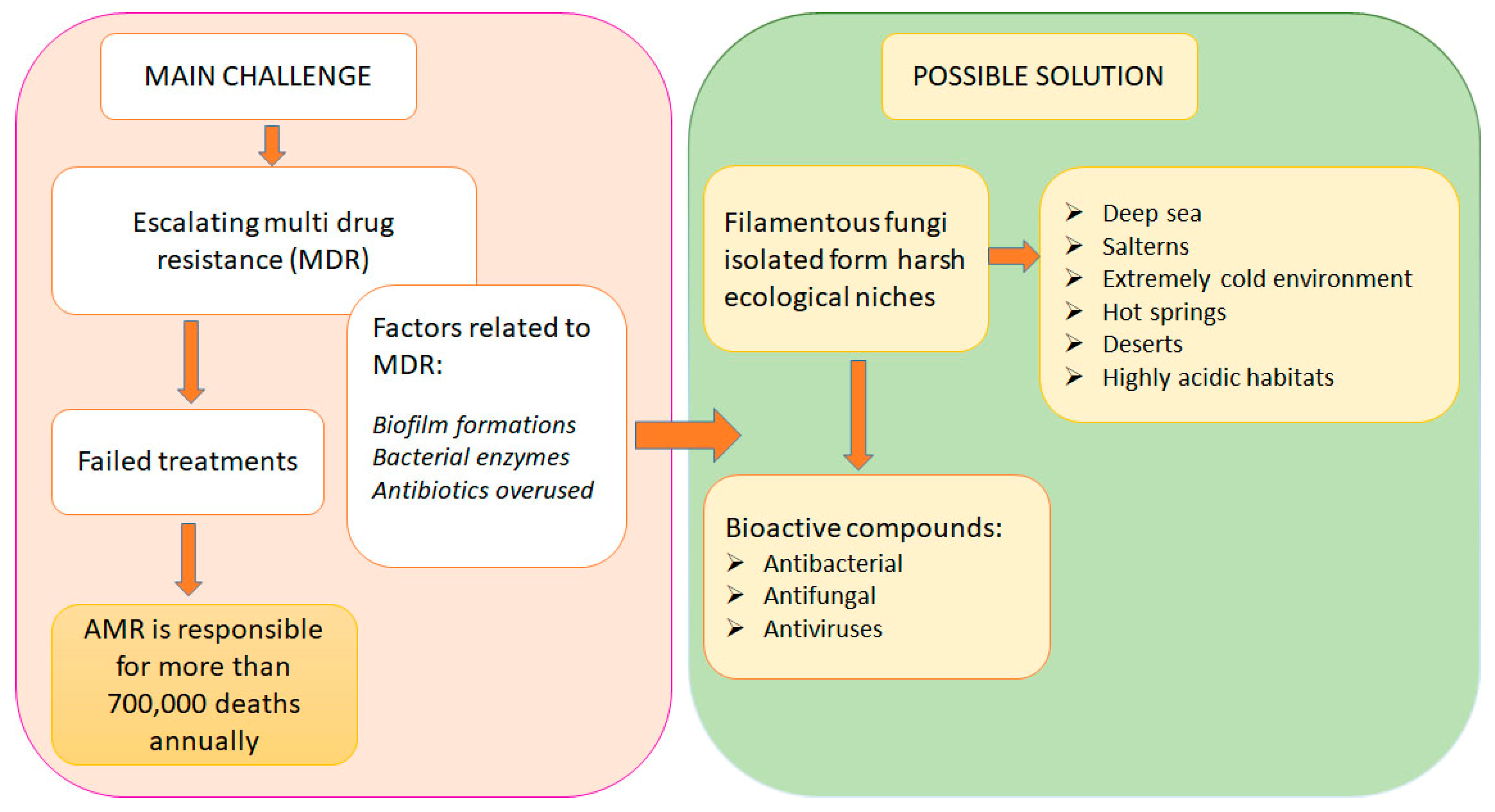
2. Antimicrobial Resistance
2.1. Contributing Factors to Antibiotic Failure and Underlying Mechanisms
2.1.1. Biofilm Formation
2.1.2. Adaptive Resilience and Non-Genetic Resistance
2.1.3. Modification of Antibiotic Target Sites
2.1.4. Enzymatic Degradation of Antibiotics
- β-lactamases, including extended-spectrum β-lactamases (ESBLs) and carbapenemases (e.g., KPC, NDM-1, OXA-48), which hydrolyze the β-lactam ring, rendering penicillins, cephalosporins, and carbapenems ineffective [12].
- Aminoglycoside-modifying enzymes (AMEs), which inactivate aminoglycosides by acetylation, phosphorylation, or nucleotidylation [20].
- Macrolide resistance enzymes modify the structure of macrolide antibiotics, preventing their inhibitory effects [21].
2.1.5. Reduced Membrane Permeability
- Porin mutations reduce the permeability of the bacterial cell wall, limiting the uptake of antibiotics. This has been observed in fluoroquinolone-resistant Escherichia coli and Klebsiella pneumoniae [22].
- Loss of outer membrane proteins (OMPs), particularly in Acinetobacter baumannii, has been associated with resistance to carbapenems [10].
2.1.6. Efflux Pumps
- The Resistance-Nodulation-Division (RND) family in Gram-negative bacteria efficiently removes β-lactams, fluoroquinolones, tetracyclines, and macrolides [24].
- The major facilitator superfamily (MFS) and ATP-binding cassette (ABC) transporters contribute to multidrug resistance by effluxing a broad spectrum of antibiotics [25].
2.1.7. Factors Accelerating the Rate of AMR
- ✓
- Misuse and Overuse of Antibiotics
- ✓
- Agricultural Use of Antibiotics
- ✓
- Increase in Income Levels
- ✓
- Easy Travel Routes
- ✓
- Biological Factors
2.2. Clinical and Economic Impact of Antimicrobial Resistance
2.3. Strategies to Combat Antimicrobial Resistance
3. Extremophilic Fungi: Diversity, Adaptations, and Ecological Roles
3.1. Thermophilic Fungi: Classification, Ecology, and Thermotolerance Mechanisms
3.2. Psychrophilic Fungi: Adaptations to Cold Environments
3.3. Halophilic Fungi
3.4. Acidophiles and Alkaliphiles: Adaptations to Extreme pH Environments
3.5. Other Extremes
4. Extremophilic Fungi as Producers of Antimicrobial Substances
5. Characterization of the Active Substances Produced by Extremophilic Fungi
6. Mechanisms of Action of Antimicrobial Compounds from Filamentous Fungi
6.1. Inhibition of Cell Wall Synthesis
6.2. Disruption of Cell Membrane Integrity
6.3. Inhibition of Protein Synthesis
6.4. Inhibition of Nucleic Acid Synthesis
6.5. Inhibition of Enzymatic Functions
7. The Relationship Between Extreme Environmental Conditions and the Production of Bioactive Compounds by Extremophilic Fungi
7.1. Low-Temperature Environments—Psychrophilic and Psychrotrophic Fungi
7.2. Permafrost Soils and Cold-Adapted Fungi
7.3. High-Temperature Environments: Thermophilic and Hyperthermophilic Fungi
7.4. Desert Ecosystems
7.5. Highly Acidic Habitats: Acidophilic Fungi
7.6. Saline and Hypersaline Habitats: Halotolerant and Halophilic Fungi
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| MDR | Multidrug Resistance |
| AMR | Antimicrobial resistance |
References
- Libertucci, J.; Young, V.B. The role of the microbiota in infectious diseases. Nat. Microbiol. 2018, 4, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Hutchings, M.I.; Truman, A.W.; Wilkinson, B. Antibiotics: Past, present and future. Curr. Opin. Microbiol. 2019, 51, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Muteeb, G.; Rehman, M.T.; Shahwan, M.; Aatif, M. Origin of Antibiotics and Antibiotic Resistance, and Their Impacts on Drug Development: A Narrative Review. Pharmaceuticals 2023, 16, 1615. [Google Scholar] [CrossRef] [PubMed]
- Salam, M.A.; Al-Amin, M.Y.; Salam, M.T.; Pawar, J.S.; Akhter, N.; Rabaan, A.A.; Alqumber, M.A. Antimicrobial resistance: A growing serious threat for global public health. Healthcare 2023, 11, 1946. [Google Scholar] [CrossRef]
- de la Fuente-Nunez, C.; Cesaro, A.; Hancock, R.E. Antibiotic failure: Beyond antimicrobial resistance. Drug Resist. Updates 2023, 71, 101012. [Google Scholar] [CrossRef]
- Adebisi, Y.A.; Ogunkola, I.O. The global antimicrobial resistance response effort must not exclude marginalised populations. Trop. Med. Health 2023, 51, 33. [Google Scholar] [CrossRef] [PubMed]
- Yam, E.L.Y.; Hsu, L.Y.; Yap, E.P.-H.; Yeo, T.W.; Lee, V.; Schlundt, J.; Lwin, M.O.; Limmathurotsakul, D.; Jit, M.; Dedon, P.; et al. Antimicrobial resistance in the Asia Pacific region: A meeting report. Antimicrob. Resist. Infect. Control 2019, 8, 202. [Google Scholar] [CrossRef]
- Figueroa, L.; Jiménez, C.; Rodríguez, J.; Areche, C.; Chávez, R.; Henríquez, M.; de la Cruz, M.; Díaz, C.; Segade, Y.; Vaca, I. 3-Nitroasterric acid derivatives from an Antarctic sponge-derived Pseudogymnoascus sp. fungus. J. Nat. Prod. 2015, 78, 919–923. [Google Scholar] [CrossRef]
- Chávez, R.; Fierro, F.; García-Rico, R.O.; Vaca, I. Filamentous fungi from extreme environments as a promising source of novel bioactive secondary metabolites. Front. Microbiol. 2015, 6, 903. [Google Scholar] [CrossRef]
- Ramírez-Castillo, F.Y.; Moreno-Flores, A.C.; Avelar-González, F.J.; Márquez-Díaz, F.; Harel, J.; Guerrero-Barrera, A.L. An evaluation of multidrug-resistant Escherichia coli isolates in urinary tract infections from Aguascalientes, Mexico: Cross-sectional study. Ann. Clin. Microbiol. Antimicrob. 2018, 17, 34. [Google Scholar] [CrossRef]
- Davies, D. Understanding biofilm resistance to antibacterial agents. Nat. Rev. Drug Discov. 2003, 2, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Founou, R.C.; Founou, L.L.; Essack, S.Y. Clinical and economic impact of antibiotic resistance in developing countries: A systematic review and meta-analysis. PLoS ONE 2017, 12, e0189621. [Google Scholar] [CrossRef] [PubMed]
- Khatoon, Z.; McTiernan, C.D.; Suuronen, E.J.; Mah, T.F.; Alarcon, E.I. Bacterial biofilm formation on implantable devices and approaches to its treatment and prevention. Heliyon 2018, 4, e01067. [Google Scholar] [CrossRef]
- Hancock, R.E.W.; Alford, M.A.; Haney, E.F. Antibiofilm activity of host defence peptides: Complexity provides opportunities. Nat. Rev. Microbiol. 2021, 9, 786–797. [Google Scholar] [CrossRef]
- Chatterjee, S.; Poonawala, H.; Jain, Y. Drug-resistant tuberculosis: Is India ready for the challenge? BMJ Glob. Health 2018, 3, e000971. [Google Scholar] [CrossRef]
- Bassetti, M.; Peghin, M.; Vena, A.; Giacobbe, D.R. Treatment of infections due to MDR gram-negative bacteria. Front. Med. 2019, 6, 74. [Google Scholar] [CrossRef] [PubMed]
- Kaldalu, N.; Hauryliuk, V.; Turnbull, K.J.; La Mensa, A.; Putrinš, M.; Tenson, T. In vitro studies of persister cells. Microbiol. Mol. Biol. Rev. 2020, 84, e00070-20. [Google Scholar] [CrossRef]
- Sandoval-Motta, S.; Aldana, M. Adaptive resistance to antibiotics in bacteria: A systems biology perspective. WIREs Syst. Biol. Med. 2016, 8, 253–267. [Google Scholar] [CrossRef]
- Rizi, K.S.; Ghazvini, K.; Noghondar, M. Adaptive antibiotic resistance: Overview and perspectives. J. Infect. Dis. Ther. 2018, 6, 363. [Google Scholar] [CrossRef]
- Annavajhala, M.K.; Gomez-Simmonds, A.; Uhlemann, A.-C. Multidrug-resistant Enterobacter cloacae complex emerging as a global, diversifying threat. Front. Microbiol. 2019, 10, 44. [Google Scholar] [CrossRef]
- Llor, C.; Bjerrum, L. Antimicrobial resistance: Risk associated with antibiotic overuse and initiatives to reduce the problem. Ther. Adv. Drug Saf. 2014, 5, 229–241. [Google Scholar] [CrossRef] [PubMed]
- Chokshi, A.; Sifri, Z.; Cennimo, D.; Horng, H. Global contributors to antibiotic resistance. J. Glob. Infect. Dis. 2019, 11, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Van Boeckel, T.P.; Brower, C.; Gilbert, M.; Grenfell, B.T.; Levin, S.A.; Robinson, T.P.; Teillant, A.; Laxminarayan, R. Global trends in antimicrobial use in food animals. Proc. Natl. Acad. Sci. USA 2015, 112, 5649–5654. [Google Scholar] [CrossRef]
- Prestinaci, F.; Pezzotti, P.; Pantosti, A. Antimicrobial resistance: A global multifaceted phenomenon. Pathog. Glob. Health 2015, 109, 309–318. [Google Scholar] [CrossRef]
- Laxminarayan, R.; Duse, A.; Wattal, C.; Zaidi, A.K.; Wertheim, H.F.; Sumpradit, N.; Vlieghe, E.; Hara, G.L.; Gould, I.M.; Goossens, H.; et al. Antibiotic resistance—The need for global solutions. Lancet Infect. Dis. 2013, 13, 1057–1098. [Google Scholar] [CrossRef]
- Chaw, P.S.; Höpner, J.; Mikolajczyk, R. The knowledge, attitude and practice of health practitioners towards antibiotic prescribing and resistance in developing countries—A systematic review. J. Clin. Pharm. Ther. 2018, 43, 606–613. [Google Scholar] [CrossRef]
- Dadgostar, P. Antimicrobial resistance: Implications and costs. Infect. Drug Resist. 2019, 12, 3903–3910. [Google Scholar] [CrossRef] [PubMed]
- Bartlett, J.G.; Gilbert, D.N.; Spellberg, B. Seven ways to preserve the miracle of antibiotics. Clin. Infect. Dis. 2013, 56, 1445–1450. [Google Scholar] [CrossRef]
- Rhouma, M.; Beaudry, F.; Thériault, W.; Letellier, A. Colistin in pig production: Chemistry, mechanism of antibacterial action, microbial resistance emergence, and one health perspectives. Front. Microbiol. 2016, 7, 1789. [Google Scholar] [CrossRef]
- Klein, E.Y.; Van Boeckel, T.P.; Martinez, E.M.; Laxminarayan, R. Global increase and geographic convergence in antibiotic consumption between 2000 and 2015. Proc. Natl. Acad. Sci. USA 2018, 115, E3463–E3470. [Google Scholar] [CrossRef]
- Ruppé, E.; Armand-Lefèvre, L.; Estellat, C.; El-Mniai, A.; Boussadia, Y.; Consigny, P.H.; Girard, P.M.; Vittecoq, D.; Bouchaud, O.; Pialoux, G.; et al. Acquisition of carbapenemase-producing Enterobacteriaceae by healthy travellers to India, France, February 2012 to March 2013. Eurosurveillance 2014, 19, 20768. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Jeannot, K.; Xiao, Y.; Knapp, C.W. Editorial: Horizontal gene transfer mediated bacterial antibiotic resistance. Front. Microbiol. 2019, 10, 1933. [Google Scholar] [CrossRef] [PubMed]
- Murray, C.J.L.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Aguilar, G.R.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E.; et al. Global burden of bacterial antimicrobial resistance in 2019: A systematic analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef] [PubMed]
- Williams, P.C.M.; Jones, M.; Snelling, T.L.; Duguid, R.; Moore, N.; Dickson, B.; Wu, Y.; Saunders, J.; Wijeratne, P.; Douangnouvong, A.; et al. Coverage gaps in empiric antibiotic regimens used to treat serious bacterial infections in neonates and children in Southeast Asia and the Pacific. Lancet Reg. Health Southeast Asia 2023, 22, 100291. [Google Scholar] [CrossRef]
- Bassetti, S.; Tschudin-Sutter, S.; Egli, A.; Osthoff, M. Optimizing antibiotic therapies to reduce the risk of bacterial resistance. Eur. J. Intern. Med. 2022, 99, 7–12. [Google Scholar] [CrossRef]
- Friedrich, M.J. Drug-resistant tuberculosis predicted to increase in high-burden countries. JAMA 2017, 318, 231. [Google Scholar] [CrossRef]
- Aslam, B.; Wang, W.; Arshad, M.I.; Khurshid, M.; Muzammil, S.; Rasool, M.H.; Nisar, M.A.; Alvi, R.F.; Aslam, M.A.; Qamar, M.U.; et al. Antibiotic resistance: A rundown of a global crisis. Infect. Drug Resist. 2018, 11, 1645–1658. [Google Scholar] [CrossRef]
- Hao, H.; Cheng, G.; Iqbal, Z.; Ai, X.; Hussain, H.I.; Huang, L.; Dai, M.; Wang, Y.; Liu, Z.; Yuan, Z. Benefits and risks of antimicrobial use in food-producing animals. Front. Microbiol. 2014, 5, 288. [Google Scholar] [CrossRef]
- MacElroy, R.D. Some comments on the evolution of extremophiles. Biosystems 1974, 6, 74–75. [Google Scholar] [CrossRef]
- Arbab, S.; Ullah, H.; Khan, M.I.U.; Khattak, M.N.K.; Zhang, J.; Li, K. Diversity and distribution of thermophilic microorganisms and their applications in biotechnology. J. Basic Microbiol. 2022, 62, 95–108. [Google Scholar] [CrossRef]
- Gostinčar, C.; Stajich, J.E.; Gunde-Cimerman, N. Extremophilic and extremotolerant fungi. Curr. Biol. 2023, 33, 752–756. [Google Scholar] [CrossRef] [PubMed]
- Kanekar, P.P.; Kanekar, S.P. Psychrophilic, Psychrotrophic, and Psychrotolerant Microorganisms. In Diversity and Biotechnology of Extremophilic Microorganisms. In Diversity and Biotechnology of Extremophilic Microorganisms from India; Kanekar, P.P., Kanekar, S.P., Eds.; Springer Nature: Singapore, 2022; pp. 215–249. ISBN 978-981-19157-3-4. [Google Scholar]
- Das, T.; Ray, P.; Nandy, S.; Al-Tawaha, A.R.; Pandey, D.K.; Kumar, V.; Dey, A. Piezophilic fungi: Sources of novel natural products with preclinical and clinical significance. In Extremophilic Fungi: Ecology, Physiology and Applications; Springer: Singapore, 2022; pp. 523–545. [Google Scholar]
- Coleine, C.; Stajich, J.E.; Selbmann, L. Fungi are key players in extreme ecosystems. Trends Ecol. Evol. 2022, 37, 517–528. [Google Scholar] [CrossRef] [PubMed]
- Merino, N.; Aronson, H.S.; Bojanova, D.P.; Feyhl-Buska, J.; Wong, M.L.; Zhang, S.; Giovannelli, D. Living at the extremes: Extremophiles and the limits of life in a planetary context. Front. Microbiol. 2019, 10, 780. [Google Scholar] [CrossRef]
- Hallsworth, J.E.; Mancinelli, R.L.; Conley, C.A.; Dallas, T.D.; Rinaldi, T.; Davila, A.F.; Benison, K.C.; Rapoport, A.; Cavalazzi, B.; Selbmann, L.; et al. Astrobiology of life on Earth. Environ. Microbiol. 2021, 23, 3335–3344. [Google Scholar] [CrossRef]
- Basu, B.; Roy, C.; Saha, S.; Tirkey, A.S.; Biswas, R.; Dey, A.; Mitra, A.K. Extremophiles to polyextremophiles: Survival of the fittest. In Extremophiles: A Paradox of Nature with Biotechnological Implications; De Gruyter: Berlin, Germany; Boston, MA, USA, 2022; Volume 1, p. 221. [Google Scholar]
- Fenice, M.; Khare, S.K.; Gorrasi, S. Mining, designing, mechanisms and applications of extremophilic enzymes. Front. Microbiol. 2021, 12, 709377. [Google Scholar] [CrossRef] [PubMed]
- Naranjo-Ortiz, M.A.; Gabaldón, T. Fungal evolution: Major ecological adaptations and evolutionary transitions. Biol. Rev. Camb. Philos. Soc. 2019, 94, 1443–1476. [Google Scholar] [CrossRef]
- Selbmann, L.; Egidi, E.; Isola, D.; Onofri, S.; Zucconi, L.; de Hoog, G.S.; Varese, G.C. Biodiversity, evolution and adaptation of fungi in extreme environments. Plant Biosyst. 2013, 147, 237–246. [Google Scholar] [CrossRef]
- Drake, H.; Ivarsson, M. The role of anaerobic fungi in fundamental biogeochemical cycles in the deep biosphere. Fungal Biol. Rev. 2018, 32, 20–25. [Google Scholar] [CrossRef]
- Miteva-Staleva, J.; Krumova, E.; Stefanova, T.; Angelova, M. Age-related changes in reactive oxygen species production in the filamentous fungus Penicillium rugulosum T35 under cold stress conditions. Comptes Rendus L’académie Bulg. Sci. 2015, 68, 1123–1129. [Google Scholar]
- Gao, Y.; Fang, H.; Fang, L.; Liu, D.; Liu, J.; Su, M.; Jiao, H. The modification and design of antimicrobial peptide. Curr. Pharm. Des. 2018, 24, 904–910. [Google Scholar] [CrossRef]
- Gunde-Cimerman, N.; Plemenitaš, A.; Oren, A. Strategies of adaptation of microorganisms of the three domains of life to high salt concentrations. FEMS Microbiol. Rev. 2018, 1, 353–375. [Google Scholar] [CrossRef]
- Gostinčar, C.; Gunde-Cimerman, N.; Grube, M. Polyextremotolerance as the fungal answer to changing environments. In Microbial Evolution Under Extreme Conditions; Bakermans, C., Ed.; De Gruyter: Berlin, Germany; Boston, MA, USA, 2015; pp. 185–208. [Google Scholar] [CrossRef]
- Lindt, W. Mitteilungen über einige neue pathogene Schimmelpilze. Arch. Exp. Pathol. Pharmakol. 1886, 21, 269–298. [Google Scholar] [CrossRef]
- Tansey, M.R.; Brock, T.D. The upper temperature limit for eukaryotic organisms. Proc. Natl. Acad. Sci. USA 1972, 69, 2426–2428. [Google Scholar] [CrossRef]
- Maheshwari, R.; Bharadwaj, G.; Bhat, M.K. Thermophilic fungi: Their physiology and enzymes. Microbiol. Mol. Biol. Rev. 2000, 64, 461–488. [Google Scholar] [CrossRef] [PubMed]
- Patel, H.; Rawat, S. Thermophilic fungi: Diversity, physiology, genetics, and applications. In New and Future Developments in Microbial Biotechnology and Bioengineering; Elsevier: Amsterdam, The Netherlands, 2021; pp. 69–93. [Google Scholar]
- Cooney, D.G.; Emerson, R. Thermophilic Fungi; W.H. Freeman and Company: San Francisco, CA, USA, 1964; Volume 27. [Google Scholar]
- Massimo, N.C.; Devan, M.N.; Arendt, K.R.; Wilch, M.H.; Riddle, J.M.; Furr, S.H.; Steen, C.; U’Ren, J.M.; Sandberg, D.C.; Arnold, A.E. Fungal endophytes in aboveground tissues of desert plants: Infrequent in culture, but highly diverse and distinctive symbionts. Mycologia 2015, 70, 61–76. [Google Scholar] [CrossRef] [PubMed]
- St-Germain, G.; Robert, A.; Ishak, M.; Tremblay, C.; Claveau, S. Infection due to: Report of four cases in patients with leukemia and review. Clin. Infect. Dis. 1993, 16, 640–645. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Qiu, M.; Xu, H.; Guo, P.; Peng, Y. Application of Metagenomics Next-Generation Sequencing on Diagnosis of Disseminated Infection Caused by Rhizomucor pusillus in an Acute Lymphoblastic Leukemia Patient. Infect. Drug Resist. 2024, 17, 5707–5713. [Google Scholar] [CrossRef]
- Tedersoo, L.; Sánchez-Ramírez, S.; Kõljalg, U.; Bahram, M.; Döring, M.; Schigel, D.; May, T.; Ryberg, M.; Abarenkov, K. High-level classification of the Fungi and a tool for evolutionary ecological analyses. Fungal Divers. 2018, 90, 135–159. [Google Scholar] [CrossRef]
- Salar, R.K. Thermophilic fungi: Basic Concepts and Biotechnological Applications, 1st ed.; CRC Press: Boca Raton, FA, USA, 2018. [Google Scholar] [CrossRef]
- Kirk, P.M.; Cannon, P.F.; Minter, D.W.; Stalpers, J.A. Dictionary of the Fungi, 10th ed.; CABI: Wallingford, UK, 2008. [Google Scholar]
- Ianutsevich, E.A.; Danilova, O.A.; Groza, N.V.; Kotlova, E.R.; Tereshina, V.M. Heat shock response of thermophilic fungi: Membrane lipids and soluble carbohydrates under elevated temperatures. Microbiology 2016, 162, 989–999. [Google Scholar] [CrossRef]
- Almaguer Chávez, M. Thermotolerance and Adaptation to Climate Change. In The Impact of Climate Change. In The Impact of Climate Change on Fungal Diseases; Springer International Publishing: Cham, Switzerland, 2022; pp. 37–71. [Google Scholar] [CrossRef]
- Hassan, N.; Rafiq, M.; Hayat, M.; Shah, A.A.; Hasan, F. Psychrophilic and psychrotrophic fungi: A comprehensive review. Rev. Environ. Sci. Biotechnol. 2016, 15, 147–172. [Google Scholar] [CrossRef]
- Morita, R.Y. Psychrophilic bacteria. Bacteriol. Rev. 1975, 39, 144–167. [Google Scholar] [CrossRef] [PubMed]
- Di Menna, M.E. Three new yeasts from Antarctic soils: Candida nivalis, Candida gelida and Candida frigida spp. Antonie Van Leeuwenhoek 1966, 32, 25–28. [Google Scholar] [CrossRef] [PubMed]
- Abrashev, R.; Miteva-Staleva, J.; Gocheva, Y.; Stoyancheva, G.; Dishliyska, V.; Spasova, B.; Krumova, E.; Angelova, M. Cell Response to Oxidative Stress in Antarctic Filamentous Fungi. Appl. Sci. 2025, 15, 5149. [Google Scholar] [CrossRef]
- Ma, F.; Hanna, M.A. Biodiesel production: A review. Bioresour. Technol. 1999, 70, 1–15. [Google Scholar] [CrossRef]
- Golubev, V.I. Rhodotorula creatinovora and R. yakutica—New species of basidiomycetous yeasts, extracted from permafrost soils of eastern Siberian Arctic. Mikol. Fitopatol. 1998, 32, 8–13. [Google Scholar]
- Jodłowska, I.; Białkowska, A.M. Cold-Adapted Fungi: Goldmine of Biomolecules Applicable in Industry. Appl. Sci. 2024, 14, 11950. [Google Scholar] [CrossRef]
- Yadav, A.N.; Yadav, N.; Sachan, S.G.; Saxena, A.K. Biodiversity of psychrotrophic microbes and their biotechnological applications. J. Appl. Biol. Biotechnol. 2019, 7, 99–108. [Google Scholar] [CrossRef]
- Miteva-Staleva, J.; Krumova, E.; Stoyancheva, G.; Kostadinova, N.; Grozdanov, P.; Spassova, B.; Angelova, M. Isolation, Identification and Proteolytic Activity of Filamentous Fungi from Alaska. Acta Microbiol. Bulg. 2022, 38, 26–30. [Google Scholar]
- Tsuji, M. Survey on Fungi in Antarctica and High Arctic Regions, and Their Impact on Climate Change. Climate 2023, 11, 195. [Google Scholar] [CrossRef]
- Yusof, N.A.; Hashim, N.H.F.; Bharudin, I. Cold Adaptation Strategies and the Potential of Psychrophilic Enzymes from the Antarctic Yeast, Glaciozyma antarctica PI12. J. Fungi 2021, 7, 528. [Google Scholar] [CrossRef]
- Bowman, J.P. Genomics of Psychrophilic Bacteria and Archaea. In Psychrophiles: From Biodiversity to Biotechnology; Margesin, R., Ed.; Springer: Cham, Switzerland, 2017; pp. 201–220. [Google Scholar]
- Zalar, P.; de Hoog, G.S.; Gunde-Cimerman, N. Trimmatostroma salinum, a new species from hypersaline water. Stud. Mycol. 1999, 43, 61–67. [Google Scholar]
- Zalar, P.; de Hoog, G.S.; Gunde-Cimerman, N. Taxonomy of the endoconidial black yeast genera Phaeotheca and Hyphospora. Stud. Mycol. 1999, 43, 49–56. [Google Scholar]
- Gunde-Cimerman, N.; Zalar, P.; de Hoog, G.S.; Plemenitaš, A. Hypersaline waters in salterns—Natural ecological niches for halophilic black yeasts. FEMS Microbiol. Ecol. 2000, 32, 235–240. [Google Scholar] [CrossRef]
- Zajc, J.; Gunde-Cimerman, N. The genus Wallemia—From contamination of food to health threat. Microorganisms 2018, 6, 46. [Google Scholar] [CrossRef]
- Gunde-Cimerman, N.; Plemenitaš, A. Ecology and molecular adaptations of the halophilic black yeast Hortaea werneckii. Rev. Environ. Sci. Biotechnol. 2007, 5, 323–331. [Google Scholar] [CrossRef]
- Gunde-Cimerman, N.; Ramos, J.; Plemenitaš, A. Halotolerant and halophilic fungi. Mycol. Res. 2009, 113, 1231–1241. [Google Scholar] [CrossRef]
- Hohmann, S. Osmotic stress signaling and osmoadaptation in yeasts. Microbiol. Mol. Biol. Rev. 2002, 66, 300–372. [Google Scholar] [CrossRef] [PubMed]
- Kejžar, A.; Gobec, S.; Plemenitaš, A.; Lenassi, M. Melanin is crucial for growth of the black yeast Hortaea werneckii in its natural hypersaline environment. Fungal Biol. 2013, 117, 368–379. [Google Scholar] [CrossRef]
- Rafiq, M.; Hassan, N.; Rehman, M.; Hasan, F. Adaptation mechanisms and applications of psychrophilic fungi. In Fungi in Extreme Environments: Ecological Role and Biotechnological Significance; Tiquia-Arashiro, S., Grube, M., Eds.; Springer: Cham, Switzerland, 2019; pp. 145–160. [Google Scholar] [CrossRef]
- Kejžar, A.; Grötli, M.; Tamás, M.J.; Plemenitaš, A.; Lenassi, M. HwHog1 kinase activity is crucial for survival of Hortaea werneckii in extremely hyperosmolar environments. Fungal Genet. Biol. 2015, 74, 45–58. [Google Scholar] [CrossRef]
- Jin, Q.; Kirk, M.F. pH as a primary control in environmental microbiology: 1. Thermodynamic perspective. Front. Environ. Sci. 2018, 6, 21. [Google Scholar] [CrossRef]
- Krulwich, T.A.; Sachs, G.; Padan, E. Molecular aspects of bacterial pH sensing and homeostasis. Nat. Rev. Microbiol. 2011, 9, 330–343. [Google Scholar] [CrossRef] [PubMed]
- Baker-Austin, C.; Dopson, M. Life in acid: pH homeostasis in acidophiles. Trends Microbiol. 2007, 15, 165–171. [Google Scholar] [CrossRef]
- Matin, A. pH homeostasis in acidophiles. Novartis Found. Symp. 2007, 221, 152–166. [Google Scholar]
- Buetti-Dinh, A.; Dethlefsen, O.; Friedman, R.; Dopson, M. Transcriptomic analysis reveals how a lack of potassium ions increases Sulfolobus acidocaldarius sensitivity to pH changes. Microbiology 2016, 162, 1422–1434. [Google Scholar] [CrossRef]
- Christel, S.; Herold, M.; Bellenberg, S.; El Hajjami, M.; Buetti-Dinh, A.; Pivkin, I.V.; Sand, W.; Wilmes, P.; Poetsch, A.; Dopson, M. Multi-omics reveals the lifestyle of the acidophilic, mineral-oxidizing model species Leptospirillum ferriphilum. Appl. Environ. Microbiol. 2018, 84, e02091-17. [Google Scholar] [CrossRef]
- Sahay, S. Extremophilic fungi: Ecology, physiology, and applications. In Extremophilic Fungi; Springer: Singapore, 2022. [Google Scholar] [CrossRef]
- Goswami, S.; Das, M. Extremophiles: A clue to the origin of life and biology of other planets. Everyman’s Sci. 2016, 51, 17–25. [Google Scholar]
- Poli, A.; Finore, I.; Romano, I.; Gioiello, A.; Lama, L.; Nicolaus, B. Microbial diversity in extreme marine habitats and their biomolecules. Microorganisms 2017, 5, 25. [Google Scholar] [CrossRef] [PubMed]
- Belyagoubi, L.; Belyagoubi-Benhammou, N.; Jurado, V.; Dupont, J.; Lacoste, S.; Djebbah, F.; Saiz-Jimenez, C. Antimicrobial activities of culturable microorganisms (actinomycetes and fungi) isolated from Chaabe Cave, Algeria. Int. J. Speleol. 2018, 47, 189–199. [Google Scholar] [CrossRef]
- Meglali, A.; Ghellai, L. An overview of extremophiles: Microbial diversity, adaptive strategies, and potential applications. Microbiol. Biotechnol. Lett. 2024, 52, 233–254. [Google Scholar] [CrossRef]
- Gonçalves, N.S.; Mello, T.M.S.P.; Mizuno, C.S.; Haider, S.; Santos, R.A. Cis-trimethoxystilbene exhibits higher genotoxic and antiproliferative effects than its isomer trans-trimethoxystilbene in MCF-7 and MCF-10A cell lines. Genet. Mol. Biol. 2021, 44, e20200477. [Google Scholar] [CrossRef]
- Fredimoses, M.; Zhou, X.; Lin, X.; Tian, X.; Ai, W.; Wang, J.; Liao, S.; Liu, J.; Yang, B.; Yang, X.; et al. New prenylxanthones from the deep-sea derived fungus Emericella sp. SCSIO 05240. Mar. Drugs 2014, 12, 4218–4234. [Google Scholar] [CrossRef] [PubMed]
- Yao, Q.; Wang, J.; Zhang, X.; Nong, X.; Xu, X.; Qi, S. Cytotoxic polyketides from the deep-sea-derived fungus Engyodontium album DFFSCS021. Mar. Drugs 2014, 12, 2774–2790. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; He, W.; Huang, X.; Tian, X.; Liao, S.; Yang, B.; Wang, F.; Zhou, X.; Liu, Y. Antifungal new oxepine-containing alkaloids and xanthones from the deep-sea-derived fungus Aspergillus versicolor SCSIO 05879. J. Agric. Food Chem. 2016, 64, 2910–2916. [Google Scholar] [CrossRef]
- Wang, W.; Chen, R.; Luo, Z.; Wang, W.; Chen, J. Antimicrobial activity and molecular docking studies of a novel anthraquinone from a marine-derived fungus Aspergillus versicolor. Nat. Prod. Res. 2017, 32, 558–563. [Google Scholar] [CrossRef]
- Wang, W.; Li, S.; Chen, Z.; Li, Z.; Liao, Y.; Chen, J. Secondary metabolites produced by the deep-sea-derived fungus Engyodontium album. Chem. Nat. Compd. 2017, 53, 224–226. [Google Scholar] [CrossRef]
- Luo, X.; Lin, X.; Salendra, L.; Pang, X.; Dai, Y.; Yang, B.; Liu, J.; Wang, J.; Zhou, X.; Liu, Y. Isobenzofuranones and isochromenones from the deep-sea-derived fungus Leptosphaeria sp. SCSIO 41005. Mar. Drugs 2017, 15, 204. [Google Scholar] [CrossRef]
- Niu, S.; Liu, D.; Shao, Z.; Proksch, P.; Lin, W. Eutypellazines a-m, thiodiketopiperazine-type alkaloids from deep-sea-derived fungus Eutypella sp. MCCC 3A00281. RSC Adv. 2017, 7, 33580–33590. [Google Scholar] [CrossRef]
- Niu, S.; Liu, D.; Shao, Z.; Proksch, P.; Lin, W. Eutypellazines n–s, new thiodiketopiperazines from a deep-sea sediment-derived fungus Eutypella sp. with anti-VRE activities. Tetrahedron Lett. 2017, 58, 3695–3699. [Google Scholar] [CrossRef]
- Takahashi, K.; Sakai, K.; Nagano, Y.; Orui-Sakaguchi, S.; Lima, A.O.; Pellizari, V.H.; Iwatsuki, M.; Takishita, K.; Nonaka, K.; Fujikura, K. Cladomarine, a new anti-saprolegniasis compound isolated from the deep-sea fungus Penicillium coralligerum YK-247. J. Antibiot. 2017, 70, 911–914. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhang, Y.-X.; Zhang, J.-P.; Yu, H.-B.; Liu, X.-Y.; Lu, X.-L.; Jiao, B.-H. A new sesquiterpene lactone from fungus Eutypella sp. D-1. Nat. Prod. Res. 2017, 31, 1676–1681. [Google Scholar] [CrossRef]
- Li, Y.; Sun, B.; Liu, S.; Jiang, L.; Liu, X.; Zhang, H.; Che, Y. Bioactive asterric acid derivatives from the Antarctic ascomycete fungus Geomyces sp. J. Nat. Prod. 2008, 71, 1643–1646. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Wiese, J.; Labes, A.; Kramer, A.; Schmaljohann, R.; Imhoff, J.F. Lindgomycin, an unusual antibiotic polyketide from a marine fungus of the Lindgomycetaceae. Mar. Drugs 2015, 13, 4617–4632. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Zhu, M.-L.; Sun, G.-Y.; Li, N.; Gu, Q.-Q.; Li, D.-H.; Che, Q.; Zhu, T.-J. Exopisiod B and Farylhydrazone C, two new alkaloids from the Antarctic-derived fungus Penicillium sp. HDN14-431. J. Asian Nat. Prod. Res. 2016, 18, 959–965. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wei, X.; Qin, X.; Tian, X.; Liao, L.; Li, K.; Zhou, X.; Yang, X.; Wang, F.; Zhang, T.; et al. Antiviral merosesquiterpenoids produced by the Antarctic fungus Aspergillus ochraceopetaliformis SCSIO 05702. J. Nat. Prod. 2016, 79, 59–65. [Google Scholar] [CrossRef]
- Pan, C.; Shi, Y.; Auckloo, N.B.; Chen, X.; Chen, A.C.-T.; Tao, X.; Wu, B. An unusual conformational isomer of verrucosidin backbone from a hydrothermal vent fungus, Penicillium sp. Y-50-10. Mar. Drugs 2016, 14, 156. [Google Scholar] [CrossRef]
- Shaaban, M.; El-Metwally, M.M.; Abdel-Razek, A.A.; Laatsch, H. Terretonin M: A new meroterpenoid from the thermophilic Aspergillus terreus TM8 and revision of the absolute configuration of penisimplicins. Nat. Prod. Res. 2018, 32, 2437–2446. [Google Scholar] [CrossRef]
- Zhang, X.; Li, S.J.; Li, J.J.; Liang, Z.Z.; Zhao, C.Q. Novel natural products from extremophilic fungi. Mar. Drugs 2018, 16, 194. [Google Scholar] [CrossRef]
- Zheng, J.; Zhu, H.; Hong, K.; Wang, Y.; Liu, P.; Wang, X.; Peng, X.; Zhu, W. Novel cyclic hexapeptides from marine-derived fungus Aspergillus sclerotiorum PT06-1. Org. Lett. 2009, 11, 5262–5265. [Google Scholar] [CrossRef]
- Bao, J.; Zhai, H.; Zhu, K.; Yu, J.-H.; Zhang, Y.; Wang, Y.; Jiang, C.-S.; Zhang, X.; Zhang, Y.; Zhang, H. Bioactive pyridone alkaloids from a deep-sea-derived fungus Arthrinium sp. UJNMF0008. Mar. Drugs 2018, 16, 174. [Google Scholar] [CrossRef]
- Elhosainy, A.M. Evaluation of Some Biological Activities of Phialosimplex asmahalo. Egypt. Acad. J. Biol. Sci. G Microbiol. 2020, 12, 9–19. [Google Scholar] [CrossRef]
- Oh, H.; Lee, Y.; Kim, Y.; Seo, Y.; Kang, J.; Park, E.; Yoon, Y. Development of antimicrobial hydrogel with edible formulations to control foodborne pathogens on food surfaces consumed raw. Innov. Food Sci. Emerg. Technol. 2021, 74, 102845. [Google Scholar] [CrossRef]
- Ballav, S.; Kerkar, S.; Thomas, S.; Augustine, N. Halophilic and halotolerant actinomycetes from a marine saltern of Goa, India producing anti-bacterial metabolites. J. Biosci. Bioeng. 2015, 119, 323–330. [Google Scholar] [CrossRef]
- Corral, P.; Esposito, F.P.; Tedesco, P.; Falco, A.; Tortorella, E.; Tartaglione, L.; de Pascale, D. Identification of a sorbicillinoid-producing Aspergillus strain with antimicrobial activity against Staphylococcus aureus: A new polyextremophilic marine fungus from Barents Sea. Mar. Biotechnol. 2018, 20, 502–511. [Google Scholar] [CrossRef]
- Wingfield, M.J.; Pham, N.Q.; Marincowitz, S.; Duong, T.A.; Wingfield, B.D.; Wilson, A.M. Blast from the past: A study of decades-old fungal cultures resolves a long-standing tree disease mystery. J. Plant Pathol. 2024, 106, 377–384. [Google Scholar] [CrossRef]
- Wang, H.; Li, Y.; Wang, Y.; Shi, T.; Wang, B. Penicillium janthinellum: A Potential Producer of Natural Products. Fermentation 2024, 10, 157. [Google Scholar] [CrossRef]
- Agrawal, S.; Chavan, P.; Dufossé, L. Hidden treasure: Halophilic fungi as a repository of bioactive lead compounds. J. Fungi 2024, 10, 290. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Bae, Y.M.; Jung, K.S.; Heu, S.; Lee, S.Y. Antimicrobial activity of natural antimicrobial substances against spoilage bacteria isolated from fresh produce. Food Control 2013, 32, 665–672. [Google Scholar] [CrossRef]
- Pócsi, I.; Dijksterhuis, J.; Houbraken, J.; de Vries, R.P. Biotechnological potential of salt-tolerant and xerophilic species of Aspergillus. Appl. Microbiol. Biotechnol. 2024, 108, 521. [Google Scholar] [CrossRef]
- Hafez Goran, S.; Taktaz, F.; Ayatollahi, S.A.; Kijjoa, A. Anthraquinones and their analogues from marine-derived fungi: Chemistry and biological activities. Mar. Drugs 2022, 20, 474. [Google Scholar] [CrossRef]
- Deng, J.; Li, Y.; Yuan, Y.; Yin, F.; Chao, J.; Huang, J.; Liu, Z.; Wang, K.; Zhu, M. Secondary metabolites from the genus Eurotium and their biological activities. Foods 2023, 12, 4452. [Google Scholar] [CrossRef]
- Zhuravleva, O.I.; Chingizova, E.A.; Oleinikova, G.K.; Starnovskaya, S.S.; Antonov, A.S.; Kirichuk, N.N.; Menshov, A.S.; Popov, R.S.; Kim, N.Y.; Berdyshev, D.V.; et al. Anthraquinone derivatives and other aromatic compounds from marine fungus Asteromyces cruciatus KMM 4696 and their effects against Staphylococcus aureus. Mar. Drugs 2023, 21, 431. [Google Scholar] [CrossRef] [PubMed]
- Pacelli, C.; Cassaro, A.; Maturilli, A.; Timperio, A.M.; Gevi, F.; Cavalazzi, B.; Onofri, S. Multidisciplinary characterization of melanin pigments from the black fungus Cryomyces antarcticus. Appl. Microbiol. Biotechnol. 2020, 104, 6385–6395. [Google Scholar] [CrossRef]
- Cavalcante, S.B.; da Silva, A.F.; Pradi, L.; Lacerda, J.W.F.; Tizziani, T.; Sandjo, L.P.; Robl, D. Antarctic fungi produce pigment with antimicrobial and antiparasitic activities. Braz. J. Microbiol. 2024, 55, 1251–1263. [Google Scholar] [CrossRef]
- Afroz, T.M.; Rahman, M.H.; Rahman, M.S.; Arif, M.; Nazir, K.H.M.N.H.; Dufossé, L. Fungal pigments: Carotenoids, riboflavin, and polyketides with diverse applications. J. Fungi 2023, 9, 454. [Google Scholar] [CrossRef]
- Chen, M.; Zheng, Y.Y.; Chen, Z.Q.; Shen, N.X.; Shen, L.; Zhang, F.M.; Wang, C.Y. NaBr-induced production of brominated azaphilones and related tricyclic polyketides by the marine-derived fungus Penicillium janthinellum HK1-6. J. Nat. Prod. 2019, 82, 368–374. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Xu, J. Fungal pigments and their roles associated with human health. J. Fungi 2020, 6, 280. [Google Scholar] [CrossRef] [PubMed]
- Giddings, L.A.; Newman, D.J. Extremophilic fungi from marine environments: Underexplored sources of antitumor, anti-infective, and other biologically active agents. Mar. Drugs 2022, 20, 62. [Google Scholar] [CrossRef] [PubMed]
- Ferrarezi, J.H.; Marin, V.R.; Vieira, G.; Ferreira, H.; Sette, L.D.; Sass, D.C. Bisdechlorogeodin from Antarctic Pseudogymnoascus sp. LAMAI 2784 for citrus canker control. J. Appl. Microbiol. 2024, 135, lxae093. [Google Scholar] [CrossRef] [PubMed]
- Fujita, K.; Ikuta, M.; Nishimura, S.; Sugiyama, R.; Yoshimura, A.; Kakeya, H. Amphiol, an antifungal fungal pigment from Pseudogymnoascus sp. PF1464. J. Nat. Prod. 2021, 84, 986–992. [Google Scholar] [CrossRef]
- Zucconi, L.; Canini, F.; Isola, D.; Caneva, G. Fungi affecting wall paintings of historical value: A worldwide meta-analysis of their detected diversity. Appl. Sci. 2022, 12, 2988. [Google Scholar] [CrossRef]
- Sułkowska-Ziaja, K.; Trepa, M.; Olechowska-Jarząb, A.; Nowak, P.; Ziaja, M.; Kała, K.; Muszyńska, B. Natural compounds of fungal origin with antimicrobial activity—Potential cosmetics applications. Pharmaceuticals 2023, 16, 1200. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Wang, Y.; Wang, J.; Liu, P.; Li, J.; Zhu, W. Antimicrobial ergosteroids and pyrrole derivatives from halotolerant Aspergillus flocculosus PT05-1 cultured in a hypersaline medium. Extremophiles 2013, 17, 963–971. [Google Scholar] [CrossRef]
- Konuray, G.; Erginkaya, Z. Antimicrobial and antioxidant properties of pigments synthesized from microorganisms. In The Battle Against Microbial Pathogens: Basic Science, Technological Advances and Educational Programs; Formatex Research Center: Norristown, PA, USA, 2015. [Google Scholar]
- Conrado, R.; Gomes, T.C.; Roque, G.S.C.; De Souza, A.O. Overview of bioactive fungal secondary metabolites: Cytotoxic and antimicrobial compounds. Antibiotics 2022, 11, 1604. [Google Scholar] [CrossRef]
- Azami, H.; Watanabe, Y.; Sakai, K.; Nakahara, H.; Kojima, H.; Tokiwa, T.; Nonaka, K.; Noguchi, Y.; Nagano, Y.; Hirose, T.; et al. Antifungal profile against Candida auris clinical isolates of tyroscherin and its new analog produced by the deep-sea-derived fungal strain Scedosporium apiospermum FKJ-0499. J. Antibiot. 2024, 77, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Wu, P.; Yang, J.; Weng, Y.; Lin, Y.; Chen, Z.; Yu, F.; Lv, X.; Ni, L. Development of a novel hybrid antimicrobial peptide for enhancing antimicrobial spectrum and potency against food-borne pathogens. J. Appl. Microbiol. 2024, 135, lxae023. [Google Scholar] [CrossRef] [PubMed]
- Cadelis, M.M.; Nipper, N.S.; Grey, A.; Geese, S.; van de Pas, S.J.; Weir, B.S.; Copp, B.R.; Wiles, S. Antimicrobial Polyketide Metabolites from Penicillium bissettii and P. glabrum. Mol. 2021, 27, 240. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Zhang, F.; Huang, Z.; Liu, H.; Xie, C.; Zhang, J.; Thacker, P.A.; Qiao, S. Effects of the antimicrobial peptide cecropin AD on performance and intestinal health in weaned piglets challenged with Escherichia coli. Peptides 2012, 35, 225–230. [Google Scholar] [CrossRef]
- Liu, J.T.; Hu, B.; Gao, Y.; Zhang, J.P.; Jiao, B.H.; Lu, X.L.; Liu, X.Y. Bioactive tyrosine-derived cytochalasins from fungus Eutypella sp. D-1. Chem. Biodivers. 2014, 11, 800–806. [Google Scholar] [CrossRef]
- Zhou, H.; Li, L.; Wang, W.; Che, Q.; Li, D.; Gu, Q.; Zhu, T. Chrodrimanins I and J from the Antarctic moss-derived fungus Penicillium funiculosum GWT2-24. J. Nat. Prod. 2015, 78, 1442–1445. [Google Scholar] [CrossRef]
- Zhang, S.D.; Barbe, V.; Garel, M.; Zhang, W.-J.; Chen, H.; Santini, C.-L.; Murat, D.; Jing, H.; Zhao, Y.; Lajus, A.; et al. Genome sequence of luminous piezophile Photobacterium phosphoreum ANT-2200. Genome Announc. 2014, 2, e00096-14. [Google Scholar] [CrossRef]
- Zhang, S.D.; Petersen, N.; Zhang, W.J.; Cargou, S.; Ruan, J.; Murat, D.; Santini, C.-L.; Song, T.; Kato, T.; Notareschi, P.; et al. Swimming behaviour and magnetotaxis function of the marine bacterium strain MO-1. Environ. Microbiol. Rep. 2014, 6, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Dickinson, I.; Goodall-Copestake, W.; Thorne, M.A.; Schlitt, T.; Ávila-Jiménez, M.L.; Pearce, D.A. Extremophiles in an Antarctic marine ecosystem. Microorganisms 2016, 4, 8. [Google Scholar] [CrossRef]
- Gonçalves, V.N.; Oliveira, F.S.; Carvalho, C.R.; Schaefer, C.E.; Rosa, C.A.; Rosa, L.H. Antarctic rocks from continental Antarctica as source of potential human opportunistic fungi. Extremophiles 2017, 21, 851–860. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Wu, G.; Zhan, T.; Shao, Z.; Liu, Z. Characterization of a cold-adapted and salt-tolerant esterase from a psychrotrophic bacterium Psychrobacter pacificensis. Extremophiles 2013, 17, 809–819. [Google Scholar] [CrossRef]
- Lin, A.; Wu, G.; Gu, Q.; Zhu, T.; Li, D. New eremophilane-type sesquiterpenes from an Antarctic deep-sea-derived fungus, Penicillium sp. PR19 N-1. Arch. Pharm. Res. 2014, 37, 839–844. [Google Scholar] [CrossRef]
- Boysen, J.M.; Saeed, N.; Hillmann, F. Natural products in the predatory defense of the filamentous fungal pathogen Aspergillus fumigatus. Beilstein J. Org. Chem. 2021, 17, 1814–1827. [Google Scholar] [CrossRef]
- Cen, Q.W.; Wang, Z.Y.; Tang, Z.X.; Zhang, Y.; Chen, T.; Xue, D.W.; Xu, M.-F.; Bai, X.-L.; Zhou, T.; Shi, L.E. Initial purification of antimicrobial fermentation metabolites from Paecilomyces cicadae and its antimicrobial mechanism. LWT 2021, 148, 111785. [Google Scholar] [CrossRef]
- Anwar, A.; Kanwal, Q.; Sadiqa, A.; Razaq, T.; Khan, I.H.; Javaid, A.; Khan, S.; Tag-Eldin, E.; Ouladsmane, M. Synthesis and antimicrobial analysis of high surface area strontium-substituted calcium phosphate nanostructures for bone regeneration. Int. J. Mol. Sci. 2023, 24, 14527. [Google Scholar] [CrossRef]
- Guo, Y.; Zhou, H.; Qin, L.; Pang, Z.; Qin, T.; Ren, H.; Pan, Z.; Zhou, J. Frequency, antimicrobial resistance, and genetic diversity of Klebsiella pneumoniae in food samples. PLoS ONE 2016, 11, e0153561. [Google Scholar] [CrossRef]
- Mo, Y.; Tan, W.C.; Cooper, B.S. Antibiotic duration for common bacterial infections—A systematic review. JAC Antimicrob. Resist. 2025, 7, dlae215. [Google Scholar] [CrossRef]
- Zucconi, L.; Canini, F.; Temporiti, M.E.; Tosi, S. Extracellular enzymes and bioactive compounds from Antarctic terrestrial fungi for bioprospecting. Int. J. Environ. Res. Public Health 2020, 17, 6459. [Google Scholar] [CrossRef] [PubMed]
- Willems, T.; De Mol, M.L.; De Bruycker, A.; De Maeseneire, S.L.; Soetaert, W.K. Alkaloids from marine fungi: Promising antimicrobials. Antibiotics 2020, 9, 340. [Google Scholar] [CrossRef]
- Liao, J.; Wang, L.; Ding, S.; Tian, G.; Hu, H.; Wang, Q.; Yin, W. Molybdenum-based antimicrobial nanomaterials: A comprehensive review. Nano Today 2023, 50, 101875. [Google Scholar] [CrossRef]
- Christiansen, J.V.; Isbrandt, T.; Petersen, C.; Sondergaard, T.E.; Nielsen, M.R.; Pedersen, T.B.; Sørensen, J.L.; Larsen, T.O.; Frisvad, J.C. Fungal quinones: Diversity, producers, and applications of quinones from Aspergillus, Penicillium, Talaromyces, Fusarium, and Arthrinium. Appl. Microbiol. Biotechnol. 2021, 105, 8157–8193. [Google Scholar] [CrossRef]
- Dahlem Junior, M.A.; Nguema Edzang, R.W.; Catto, A.L.; Raimundo, J.M. Quinones as an efficient molecular scaffold in the antibacterial/antifungal or antitumoral arsenal. Int. J. Mol. Sci. 2022, 23, 14108. [Google Scholar] [CrossRef] [PubMed]
- Rogozhin, E.A.; Sadykova, V.S.; Baranova, A.A.; Vasilchenko, A.S.; Lushpa, V.A.; Mineev, K.S.; Georgieva, M.L.; Kul’ko, A.B.; Krasheninnikov, M.E.; Lyundup, A.V.; et al. A novel lipopeptaibol emericellipsin A with antimicrobial and antitumor activity produced by the extremophilic fungus Emericellopsis alkalina. Molecules 2018, 23, 2785. [Google Scholar] [CrossRef]
- Baranova, A.A.; Alferova, V.A.; Korshun, V.A.; Tyurin, A.P. Antibiotics from extremophilic micromycetes. Russ. J. Bioorganic Chem. 2020, 46, 903–971. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Zhao, S.; Yu, Y.; Chen, Z.; Shen, H.; Zhou, L. Beauvericin K, a new antifungal beauvericin analogue from a marine-derived Fusarium sp. Nat. Prod. Commun. 2016, 11, 1825–1826. [Google Scholar]
- Liu, M.; Sun, W.; Wang, J.; He, Y.; Zhang, J.; Li, F.; Qi, C.; Zhu, H.; Xue, Y.; Hu, Z.; et al. Bioactive secondary metabolites from the marine-associated fungus Aspergillus terreus. Bioorg. Chem. 2018, 80, 525–530. [Google Scholar] [CrossRef]
- Scharf, D.H.; Heinekamp, T.; Brakhage, A.A. Gliotoxin—A fungal virulence factor and its role in Aspergillus fumigatus pathogenicity. Curr. Pharm. Design 2012, 18, 4859–4873. [Google Scholar]
- Singh, S.P.; Qureshi, A.; Hassan, W. Mechanisms of action by antimicrobial agents: A review. McGill J. Med. 2021, 19. [Google Scholar] [CrossRef]
- Denning, D.W. Echinocandins: A new class of antifungal. J. Antimicrob. Chemother. 2002, 49, 889–891. [Google Scholar] [CrossRef] [PubMed]
- Roy, A.; Ahuja, S.; Garg, S. Fungal secondary metabolites: Biological activity and potential applications. In Recent Trends in Mycological Research: Volume 1: Agricultural and Medical Perspective; Springer: Cham, Switzerland, 2021; pp. 159–188. [Google Scholar]
- Waditee-Sirisattha, R.; Kageyama, H.; Takabe, T. Halophilic microorganism resources and their applications in industrial and environmental biotechnology. AIMS Microbiol. 2016, 2, 42–54. [Google Scholar] [CrossRef]
- Kour, D.; Kaur, T.; Devi, R.; Rana, K.L.; Yadav, N.; Rastegari, A.A.; Yadav, A.N. Biotechnological applications of beneficial microbiomes for evergreen agriculture and human health. In Trends of Microbial Biotechnology for Sustainable Agriculture and Biomedicine Systems: Perspectives for Human Health; Rastegari, A.A., Yadav, A.N., Yadav, N., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 255–279. [Google Scholar]
- Yadav, A.N.; Kour, D.; Kaur, T.; Devi, R.; Guleria, G.; Rana, K.L.; Yadav, N.; Rastegari, A.A. Microbial biotechnology for sustainable biomedicine systems: Current research and future challenges. In Trends of Microbial Biotechnology for Sustainable Agriculture and Biomedicine Systems: Perspectives for Human Health; Rastegari, A.A., Yadav, A.N., Yadav, N., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 281–292. [Google Scholar]
- Arifeen, M.Z.U.; Ma, Y.-N.; Xue, Y.-R.; Liu, C. Deep-Sea Fungi Could Be the New Arsenal for Bioactive Molecules. Mar. Drugs 2019, 18, 9. [Google Scholar] [CrossRef]
- Petersen, L.E.; Kellermann, M.Y.; Schupp, P.J. Secondary Metabolites of Marine Microbes: From Natural Products Chemistry to Chemical Ecology. In YOUMARES 9—The Oceans: Our Research, Our Future: Proceedings of the 2018 Conference for YOUng MArine RESearchers in Oldenburg, Germany; Jungblut, S., Liebich, V., Bode-Dalby, M., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 159–180. [Google Scholar]
- Yadav, A.N.; Kumar, R.; Kumar, S.; Kumar, V.; Sugitha, T.; Singh, B.; Chauhan, V.; Dhaliwal, H.S.; Saxena, A.K. Beneficial microbiomes: Biodiversity and potential biotechnological applications for sustainable agriculture and human health. J. Appl. Biol. Biotechnol. 2017, 5, 45–57. [Google Scholar] [CrossRef]
- Yadav, A.N.; Saxena, A.K. Biodiversity and biotechnological applications of halophilic microbes for sustainable agriculture. J. Appl. Biol. Biotechnol. 2018, 6, 48–55. [Google Scholar] [CrossRef]
- Gómez-Silva, B. Lithobiontic life: “Atacama rocks are well and alive”. Antonie Van Leeuwenhoek 2018, 111, 1333–1343. [Google Scholar] [CrossRef]
- Okoro, C.K.; Brown, R.; Jones, A.L.; Andrews, B.A.; Asenjo, J.A.; Goodfellow, M.; Bull, A.T. Diversity of culturable actinomycetes in hyper-arid soils of the Atacama Desert, Chile. Antonie Van Leeuwenhoek 2009, 95, 121–133. [Google Scholar] [CrossRef] [PubMed]
- Rosa, L.H.; Zani, C.L.; Cantrell, C.L.; Duke, S.O.; Van Dijck, P.; Desideri, A.; Rosa, C.A. Fungi in Antarctica: Diversity, ecology, effects of climate change, and bioprospection for bioactive compounds. In Fungi of Antarctica: Diversity, Ecology and Biotechnological Applications; Springer: Berlin/Heidelberg, Germany, 2019; pp. 1–17. [Google Scholar]
- Bakermans, C.; Bergholz, P.W.; Ayala-del-Río, H.; Tiedje, J. Genomic insights into cold adaptation of permafrost bacteria. In Permafrost Soils; Springer: Berlin/Heidelberg, Germany, 2009; pp. 159–168. [Google Scholar]
- Bowman, J.P. Genomic analysis of psychrophilic prokaryotes. In Psychrophiles: From Biodiversity to Biotechnology; Springer: Berlin, Germany, 2008; pp. 265–284. [Google Scholar]
- Unell, M. Physiological, Genetic and Proteomic Characterization of Arthrobacter Chlorophenolicus During Growth on Different Phenolic Substrates or Temperatures; Swedish University of Agricultural Sciences: Uppsala, Sweden, 2008; No. 2008: 26. [Google Scholar]
- Gocheva, Y.; Krumova, E.; Slokoska, L.; Miteva, J.; Angelova, M. Cell response of Antarctic and temperate strains of Penicillium spp. to different growth temperatures. Mycol. Res. 2006, 110, 1347–1354. [Google Scholar] [CrossRef]
- Ruisi, S.; Barreca, D.; Selbmann, L.; Zucconi, L.; Onofri, S. Fungi in Antarctica. Rev. Environ. Sci. Biotechnol. 2007, 6, 127–141. [Google Scholar] [CrossRef]
- Tosi, S.; Kostadinova, N.; Krumova, E.; Pashova, S.; Dishliiska, V.; Spassova, B.; Vassilev, S.; Angelova, M. Antioxidant enzyme activity of filamentous fungi isolated from Livingston Island, Maritime Antarctica. Polar Biol. 2010, 33, 1227–1237. [Google Scholar] [CrossRef]
- Feller, G.; Gerday, C. Psychrophilic enzymes: Hot topics in cold adaptation. Nat. Rev. Microbiol. 2003, 1, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Collins, T.; Roulling, F.; Piette, F.; Marx, J.C.; Feller, G.; Gerday, C.; D’Amico, S. Fundamentals of cold-adapted enzymes. In Psychrophiles: From Biodiversity to Biotechnology; Margesin, R., Schinner, F., Gerday, C., Marx, J.C., Eds.; Springer: Berlin, Germany, 2007; pp. 211–227. [Google Scholar]
- Krumova, E.; Abrashev, R.; Dishliyska, V.; Stoyancheva, G.; Kostadinova, N.; Miteva-Staleva, J.; Spasova, B.; Angelova, M. Cold-active catalase from the psychrotolerant fungus Penicillium griseofulvum. J. Biol. Microbiol. 2021, 61, 782–794. [Google Scholar] [CrossRef]
- Giordano, D. Bioactive Molecules from Extreme Environments. Mar. Drugs 2020, 18, 640. [Google Scholar] [CrossRef]
- Gittel, A.; Bárta, J.; Kohoutová, I.; Mikutta, R.; Owens, S.; Gilbert, J.; Urich, T. Distinct microbial communities associated with buried soils in the Siberian tundra. ISME J. 2014, 8, 841–853. [Google Scholar] [CrossRef] [PubMed]
- Deslippe, J.R.; Hartmann, M.; Grayston, S.J.; Simard, S.W.; Mohn, W.W. Stable isotope probing implicates a species of Cortinarius in carbon transfer through ectomycorrhizal fungal mycelial networks in Arctic tundra. New Phytol. 2016, 210, 383–390. [Google Scholar] [CrossRef]
- Fujiyoshi, M.; Yoshitake, S.; Watanabe, K.; Murota, K.; Tsuchiya, Y.; Uchida, M.; Nakatsubo, T. Successional changes in ectomycorrhizal fungi associated with the polar willow Salix polaris in a deglaciated area in the High Arctic, Svalbard. Polar Biol. 2011, 34, 667–673. [Google Scholar] [CrossRef]
- Arenz, B.E.; Blanchette, R.A. Distribution and abundance of soil fungi in Antarctica at sites on the Peninsula, Ross Sea Region, and McMurdo Dry Valleys. Soil Biol. Biochem. 2011, 43, 308–315. [Google Scholar] [CrossRef]
- Park, C.H.; Kim, K.M.; Elvebakk, A.; Kim, O.S.; Jeong, G.; Hong, S.G. Algal and fungal diversity in Antarctic lichens. J. Eukaryot. Microbiol. 2015, 62, 196–205. [Google Scholar] [CrossRef]
- Nagano, Y.; Nagahama, T.; Abe, F. Cold-adapted yeasts in deep-sea environments. In Cold-Adapted Yeasts; Buzzini, P., Margesin, R., Eds.; Springer: Berlin, Germany, 2013; pp. 149–171. [Google Scholar]
- Singh, P.; Raghukumar, C.; Verma, P.; Shouche, Y. Phylogenetic diversity of culturable fungi from the deep-sea sediments of the Central Indian Basin and their growth characteristics. Fungal Divers. 2010, 40, 89–102. [Google Scholar] [CrossRef]
- Singh, P.; Raghukumar, C.; Meena, R.M.; Verma, P.; Shouche, Y. Fungal diversity in deep-sea sediments revealed by culture-dependent and culture-independent approaches. Fungal Ecol. 2012, 5, 543–553. [Google Scholar] [CrossRef]
- Che Zain, M.S.; Lee, S.Y.; Nasir, N.M.; Fakurazi, S.; Shaari, K. Metabolite characterization and correlations with antioxidant and wound healing properties of oil palm (Elaeis guineensis Jacq.) leaflets via 1H-NMR-based metabolomics approach. Molecules 2020, 25, 5636. [Google Scholar] [CrossRef] [PubMed]
- Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2019, 36, 122–173. [Google Scholar] [CrossRef]
- Jansson, J.K.; Tas, N. The microbial ecology of permafrost. Nat. Rev. Microbiol. 2014, 12, 414–425. [Google Scholar] [CrossRef]
- Bakermans, C.; Skidmore, M.L.; Douglas, S.; McKay, C.P. Molecular characterization of bacteria from permafrost of the Taylor Valley, Antarctica. FEMS Microbiol. Ecol. 2014, 89, 331–346. [Google Scholar] [CrossRef]
- De Maayer, P.; Anderson, D.; Cary, C.; Cowan, D.A. Some like it cold: Understanding the survival strategies of psychrophiles. EMBO Rep. 2014, 15, 508–517. [Google Scholar] [CrossRef]
- Hu, W.; Zhang, Q.; Tian, T.; Cheng, G.; An, L.; Feng, H. The microbial diversity, distribution, and ecology of permafrost in China: A review. Extremophiles 2015, 19, 693–705. [Google Scholar] [CrossRef]
- Xing, C.; Chen, D.; Xie, C.; Liu, Q.; Zhong, T.; Shao, Z.; Liu, G.; Luo, L.; Yang, X. Anti-food allergic compounds from Penicillium griseofulvum MCCC 3A00225, a deep-sea-derived fungus. Mar. Drugs 2021, 19, 224. [Google Scholar] [CrossRef] [PubMed]
- Rastogi, G.; Osman, S.; Kukkadapu, R.; Engelhard, M.; Vaishampayan, P.A.; Andersen, G.L.; Sani, R.K. Microbial and mineralogical characterizations of soils collected from the deep biosphere of the former Homestake gold mine, South Dakota. Microb. Ecol. 2010, 60, 539–550. [Google Scholar] [CrossRef]
- Urbieta, M.S.; Donati, E.R.; Chan, K.G.; Shahar, S.; Sin, L.L.; Goh, K.M. Thermophiles in the genomic era: Biodiversity, science, and applications. Biotechnol. Adv. 2015, 33, 633–647. [Google Scholar] [CrossRef]
- Bajpai, P. Developments and Applications of Enzymes from Thermophilic Microorganisms; Elsevier: Amsterdam, The Netherlands, 2023. [Google Scholar]
- Hidese, R.; Fukuda, W.; Niitsu, M.; Fujiwara, S. Identification of branched-chain polyamines in hyperthermophiles. In Polyamines; Alcazar, R., Tiburcio, A.F., Eds.; Humana Press: New York, NY, USA, 2018; pp. 81–94. [Google Scholar]
- Goh, K.M.; Kahar, U.M.; Chai, Y.Y.; Chong, C.S.; Chai, K.P.; Ranjani, V.; Illias, R.M.; Chan, K.G. Recent discoveries and applications of Anoxybacillus. Appl. Microbiol. Biotechnol. 2013, 97, 1475–1488. [Google Scholar] [CrossRef] [PubMed]
- Zeldes, B.M.; Keller, M.W.; Loder, A.J.; Straub, C.T.; Adams, M.W.W.; Kelly, R.M. Extremely thermophilic microorganisms as metabolic engineering platforms for production of fuels and industrial chemicals. Front. Microbiol. 2015, 6, 1209. [Google Scholar] [CrossRef] [PubMed]
- Gessler, N.N.; Egorova, A.S.; Belozerskaya, T.A. Melanin pigments of fungi under extreme environmental conditions. Appl. Biochem. Microbiol. 2014, 50, 105–113. [Google Scholar] [CrossRef]
- Conley, C.A.; Ishkhanova, G.; McKay, C.P.; Cullings, K. A preliminary survey of non-lichenized fungi cultured from the hyperarid Atacama Desert of Chile. Astrobiology 2006, 6, 521–526. [Google Scholar] [CrossRef]
- Santiago, I.F.; Goncalves, V.N.; Gomez-Silva, B.; Galetovic, A.; Rosa, L.H. Fungal diversity in the Atacama Desert. Antonie Van Leeuwenhoek 2018, 111, 1345–1360. [Google Scholar] [CrossRef]
- Grishkan, I.; Nevo, E. Spatiotemporal distribution of soil microfungi in the Makhtesh Ramon area, central Negev desert, Israel. Fungal Ecol. 2010, 3, 326–337. [Google Scholar] [CrossRef]
- Vargas Castillo, R.; Beck, A. Photobiont selectivity and specificity in Caloplaca species in a fog-induced community in the Atacama Desert, northern Chile. Fungal Biol. 2012, 116, 665–676. [Google Scholar] [CrossRef]
- Gonçalves, V.N.; Cantrell, C.L.; Wedge, D.E.; Ferreira, M.C.; Soares, M.A.; Jacob, M.R.; Rosa, L.H. Fungi associated with rocks of the A tacama D esert: Taxonomy, distribution, diversity, ecology and bioprospection for bioactive compounds. Environ. Microbiol. 2016, 18, 232–245. [Google Scholar] [CrossRef]
- Martinelli, L.; Zalar, P.; Gunde-Cimerman, N.; Azua-Bustos, A.; Sterflinger, K.; Piñar, G. Aspergillus atacamensis and A. salisburgensis: Two new halophilic species from hypersaline/arid habitats with a Phialosimplex-like morphology. Extremophiles 2017, 21, 755–773. [Google Scholar] [CrossRef]
- Symanczik, S.; Błaszkowski, J.; Chwat, G.; Boller, T.; Wiemken, A.; Al-Yahyaei, N.; Al-Yahya’ei, M.N. Three new species of arbuscular mycorrhizal fungi discovered at one location in a desert of Oman: Diversispora omaniana, Septoglomus nakheelum and Rhizophagus arabicus. Mycology 2014, 106, 243–259. [Google Scholar] [CrossRef]
- Zalar, P.; de Hoog, G.S.; Schroers, H.J.; Frank, J.M.; Gunde-Cimerman, N. Taxonomy and phylogeny of the xerophilic genus Wallemia (Wallemiomycetes and Wallemiales, cl. et ord. nov.). Antonie Van Leeuwenhoek 2005, 87, 311–328. [Google Scholar] [CrossRef] [PubMed]
- Jancic, S.; Frisvad, J.C.; Kocev, D.; Gostincar, C.; Dzeroski, S.; Gunde-Cimerman, N. Production of secondary metabolites in extreme environments: Food- and airborne Wallemia spp. produce toxic metabolites at hypersaline conditions. PLoS ONE 2016, 11, e0169116. [Google Scholar] [CrossRef]
- Peng, X.P.; Wang, Y.; Liu, P.P.; Hong, K.; Chen, H.; Yin, X.; Zhu, W.-M. Aromatic compounds from the halotolerant fungal strain of Wallemia sebi PXP-89 in a hypersaline medium. Arch. Pharm. Res. 2011, 34, 907–912. [Google Scholar] [CrossRef]
- Jiang, W.; Ye, P.; Chen, C.T.A.; Wang, K.; Liu, P.; He, S.; Wu, B. Two novel hepatocellular carcinoma cycle inhibitory cyclodepsipeptides from a hydrothermal vent crab-associated fungus Aspergillus clavatus C2WU. Mar. Drugs 2013, 11, 4761–4772. [Google Scholar] [CrossRef] [PubMed]
- Chu, Y.S.; Niu, X.M.; Wang, Y.L.; Guo, J.P.; Pan, W.Z.; Huang, X.W.; Zhang, K.Q. Isolation of putative biosynthetic intermediates of prenylated indole alkaloids from a thermophilic fungus Talaromyces thermophilus. Org. Lett. 2010, 12, 4356–4359. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.P.; Tan, J.L.; Wang, Y.L.; Wu, H.Y.; Zhang, C.P.; Niu, X.-M.; Pan, W.Z.; Huang, X.W.; Zhang, K.-Q. Isolation of talathermophilins from the thermophilic fungus Talaromyces thermophilus YM3-4. J. Nat. Prod. 2011, 74, 2278–2281. [Google Scholar] [CrossRef]
- Azua-Bustos, A.; Urrejola, C.; Vicuña, R. Life at the dry edge: Microorganisms of the Atacama Desert. FEBS Lett. 2012, 586, 2939–2945. [Google Scholar] [CrossRef]
- Cordero, R.R.; Damiani, A.; Jorquera, J.; Sepulveda, E.; Caballero, M.; Fernandez, S.; Feron, S.; Llanillo, P.J.; Carrasco, J.; Laroze, D.; et al. Ultraviolet radiation in the Atacama Desert. Antonie Van Leeuwenhoek 2018, 111, 1301–1313. [Google Scholar] [CrossRef]
- Bull, A.T.; Asenjo, J.A.; Goodfellow, M.; Gómez-Silva, B. The Atacama Desert: Technical resources and the growing importance of novel microbial diversity. Annu. Rev. Microbiol. 2016, 70, 215–234. [Google Scholar] [CrossRef]
- Marchetta, A. Assessment of fungal diversity present in Arctic and Antarctic lakes and selection of Heavy Metal tolerant fungal isolates. Ph.D. Thesis, Università degli Studi di Messina, Messina, Italy, 2023. [Google Scholar]
- Navarro-Gonzalez, R.; Rainey, F.A.; Molina, P.; Bagaley, D.R.; Hollen, B.J.; de la Rosa, J.; Small, A.M.; Quinn, R.C.; Grunthaner, F.J.; Cáceres, L.; et al. Mars-like soils in the Atacama Desert, Chile, and the dry limit of microbial life. Science 2003, 302, 1018–1021. [Google Scholar] [CrossRef]
- Schulze-Makuch, D.; Wagner, D.; Kounaves, S.P.; Mangelsdorf, K.; Devine, K.G.; de Vera, J.P.; Zamorano, P.; Schmitt-Kopplin, P.; Grossart, H.-P.; Parro, V.; et al. Transitory microbial habitat in the hyperarid Atacama Desert. Proc. Natl. Acad. Sci. USA 2018, 115, 2670–2675. [Google Scholar] [CrossRef] [PubMed]
- Azua-Bustos, A.; Fairen, A.G.; Gonzalez-Silva, C.; Ascaso, C.; Carrizo, D.; Fernandez-Martínez, M.A.; Fernandez-Sampedro, M.; Garcia-Descalzo, L.; García-Villadangos, M.; Martin-Redondo, M.P.; et al. Unprecedented rains decimate surface microbial communities in the hyperarid core of the Atacama Desert. Sci. Rep. 2018, 8, 16706. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.B.; Quatrini, R. Acidophile microbiology in space and time. In Acidophiles: Life in Extremely Acidic Environments; Quatrini, R., Johnson, D.B., Eds.; Caister Academic Press: Norfolk, UK, 2016; pp. 3–16. [Google Scholar]
- Druschel, G.K.; Baker, B.J.; Gihring, T.M.; Banfield, J.F. Acid mine drainage biogeochemistry at Iron Mountain, California. Geochem. Trans. 2004, 5, 13. [Google Scholar] [CrossRef] [PubMed]
- Mirete, S.; Morgante, V.; Gonzalez-Pastor, J.E. Acidophiles: Diversity and mechanisms of adaptation to acidic environments. In Adaption of Microbial Life to Environmental Extremes; Stan-Lotter, H., Fendrihan, S., Eds.; Springer: Cham, Switzerland, 2017; pp. 227–251. [Google Scholar]
- Denef, V.J.; Mueller, R.S.; Banfield, J.F. AMD biofilms: Using model communities to study microbial evolution and ecological complexity in nature. ISME J. 2010, 4, 599–610. [Google Scholar] [CrossRef]
- Konings, W.N.; Albers, S.V.; Koning, S.; Driessen, A.J. The cell membrane plays a crucial role in survival of bacteria and archaea in extreme environments. Anton. Leeuw. 2002, 81, 61–72. [Google Scholar] [CrossRef]
- Falteisek, L.; Cepicka, I. Microbiology of diverse acidic and non-acidic microhabitats within a sulfidic ore mine. Extremophiles 2012, 16, 911–922. [Google Scholar] [CrossRef]
- Stierle, D.B.; Stierle, A.A.; Patacini, B.; McIntyre, K.; Girtsman, T.; Bolstad, E. Berkeleyones and related meroterpenes from a deep water acid mine waste fungus that inhibit the production of interleukin 1-b from induced inflammasomes. J. Nat. Prod. 2011, 74, 2273–2277. [Google Scholar] [CrossRef]
- Stierle, D.B.; Stierle, A.A.; Girtsman, T.; McIntyre, K.; Nichols, J. Caspase-1 and-3 inhibiting drimane sesquiterpenoids from the extremophilic fungus Penicillium solitum. J. Nat. Prod. 2012, 75, 262–266. [Google Scholar] [CrossRef]
- Díaz-Cardenas, C.; Cantillo, A.; Rojas, L.Y.; Sandoval, T.; Fiorentino, S.; Robles, J.; Ramos, F.A.; Zambrano, M.M.; Baena, S. Microbial diversity of saline environments: Searching for cytotoxic activities. AMB Express 2017, 7, 223. [Google Scholar] [CrossRef]
- Enache, M.; Teodosiu, G.; Itoh, T.; Kamekura, M.; Stan-Lotter, H. Halophilic microorganisms from manmade and natural hypersaline environments: Physiology, ecology, and biotechnological potential. In Adaption of Microbial Life to Environmental Extremes; Stan-Lotter, H., Fendrihan, S., Eds.; Springer: Cham, Switzerland, 2017; pp. 201–226. [Google Scholar]
- Oren, A. Bioenergetic aspects of halophilism. Microbiol. Mol. Biol. Rev. 1999, 63, 334–348. [Google Scholar] [CrossRef]
- Gunde-Cimerman, N.; Zalar, P. Extremely halotolerant and halophilic fungi inhabit brine in solar salterns around the globe. Food Technol. Biotechnol. 2014, 52, 170–179. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
| Fungal Species | Habitat | Antibacterial Activity | Antifungal Activity | Antiviral Activity | References |
|---|---|---|---|---|---|
| Emericella sp. | depth 3258 m, the South China Sea | E. coli (ATCC 29922), Klebsiella pneumoniae (ATCC 13883), S. aureus (ATCC 29213), Enterococcus faecalis (ATCC 29212), Acinetobacter baumannii (ATCC 19606), Aeromonas hydrophila (ATCC 7966) | Fusarium sp., Penicillium sp., A. niger, Rhizoctonia solani, Fusarium oxysporum f. sp. niveum, F. oxysporum f. sp. cucumeris | [103] | |
| Engyodontium album DFFSCS021 | 3739 m deep-sea sediment sample in the South China Sea | E. coli and Bacillus subtilis | [104] | ||
| Aspergillus versicolor SCSIO 05879 | depth 3972 m, the Indian Ocean | Colletotrichum acutatum | [105] | ||
| A. versicolor | depth of 2869 m in the West Pacific Ocean | multidrug-resistant strains of S. aureus ATCC 43300 and CGMCC 1.12409 Vibrio | [106] | ||
| Engyodontium album | depth 3542 m, the Atlantic Ocean | Vibrio vulnificus, V. rotiferianus, V. campbellii | A. niger | [107] | |
| Leptosphaeria sp. SCSIO 41005 | depth 3614 m, the Indian Ocean | H3N2, EV71, and HIV viruses | [108] | ||
| Eutypella sp. MCCC 3A00281 | depth 5610 m, the South Atlantic Ocean | S. aureus ATCC 25923 and vancomycin-resistant enterococci | HIV-1 | [109,110] | |
| Penicillium coralligerum YK-247 | depth 3064 m, São Paulo Plateau, off Brazil | Saprolegnia parasitica, Pythium sp. sakari1 | [111] | ||
| Eutypella sp. D-1 | London Island of Kongsfjorden of Ny-Ålesund District (altitude of 100 m), Arctic | E. coli, Bacillus subtilis, S. aureus | [112] | ||
| Geomyces species | Antarctic | Gram-positive and Gram-negative bacteria | A. fumigatus | [113] | |
| Pseudogymnoascus sp. F09-T18-1, | King George Island of Antarctic | antimicrobial activities | [8] | ||
| Lindgomycetaceae strains KF970 and LF327 | Antarctic and the Kiel Fjord, Baltic Sea | B. subtilis, S. aureus, methicillin-resistant S. aureus | [114] | ||
| Penicillium sp. HDN14-431 | mesolittoral zone in Antarctic | Proteusbacillus vulgaris | [115] | ||
| Aspergillus ochraceopetaliformis SCSIO 05702 | the Great Wall station (Chinese Antarctic station) | H1N1 and H3N2 influenza viruses anti-H1N1 activity | [116] | ||
| Penicillium sp. Y-50-10 | sulfur rich sediment (Kueishantao, Taiwan) | B. subtilis | [117] | ||
| Aspergillus terreus TM8 | hot desert place (~50 °C) in South Egypt | Proteus, sp., Streptococcus pyogenes | C. albicans | [118] | |
| Alternaria raphani THW-18 | sediment sample in the Hongdao sea salt field, China | E. coli, B. subtilis | C. albicans | [119] | |
| Aspergillus sclerotiorum PT06-1 | the Putian Sea Salt Field, China | P. aeruginosa | C. albicans | [120] | |
| Arthrinium sp. UJNMF0008 | South China Sea at a depth of 3858 m | Mycobacterium smegmatis, S. aureus | [121] | ||
| Phialosimplex asmahalo | obligate halophilic fungus | Hepatitis A virus. Herpes Simplex type 1 | [122] | ||
| A. affinis, E. cladophorae, Pen. lusitanum and T. aestuarinum | marine fungal strains | K. pneumoniae | [102,123] | ||
| Micromonospora sp., Kocuria sp., Actinomycetes sp. | halophilic and halotolerant | V. cholera, S. aureus, S. citreus | [124] | ||
| Aspergillus protuberus MUT 3638 | cold seawater | S. aureus, K. pneumoniae, B. metallica, A. baumanii | [125] | ||
| Pen. lusitanum | halophilic | C. albicans | [102] | ||
| Penecilium citrinum NM-3 Aspergilus subalbidus | soil of solar salt flats | E. faecalis, Micrococcus luteus, S. aureus, methicillin-resistant S. aureus, E. coli, Salmonella typhi | C. albicans, A. fumigatus | [126] | |
| Penicillium janthinellum | Gram-positive and Gram-negative bacteria, including multidrug-resistant S. aureus (MRSA) | antifungal drugs | [127] |
| Name of Active Substances | Species | Extremophilic Conditions | Antimicrobial Activity Against | References |
|---|---|---|---|---|
| Ergosteroids | Aspergillus flocculosus | Halotolerant | Enterobacter aerogenes, Pseudomonas aeruginosa, and Candida albicans | [129] |
| Terremides | A. terreus | Halophilic | E. aerogenes, P. aeruginosa Staphylococcus aureus and C. albicans | [130] |
| Anthraquinones | Halorosellinia sp. | Halophilic | antimicrobial activity | [131] |
| Eurotium rubrum | [132] | |||
| Asteromyces cruciatus KMM 4696. | Halotolerant | S. aureus | [133] | |
| Diketopiperazine alkaloid, alternarosin A | Alternaria raphanin THW-18 | Halotolerant | antimicrobial activity against E. coli, B. subtilis, and C. albicans | [128] |
| Carotenoids | Cold-adapted | antimicrobial activity | [75] | |
| Melanins | Cryomyces minterii | Cold-adapted | antimicrobial activity | [134] |
| Pseudogymnoascus spp. Talaromyces cnidii. All Pseudogymnoascus | Antarctic fungi, cold-adapted | antimicrobial activity | [135] | |
| Trimmatostroma salinum and Phaeotheca triangularis, | Halophiles | antimicrobial activity | [136] | |
| Penicilone H | Penicillium janthinellum HK1-6 | Halotoreant fundus isolated from mangrove | antimicrobial activity | [137] |
| Penicilones B–D | Penicillium janthinellum HK1-6 | Halotoreant fundus isolated from mangrove | S. aureus | [138] |
| Torulene | Rhodotorula mucilaginosa | Cold-adapted | antimicrobial activity | [75] |
| Eutypellazines P–S | deep-sea-derived fungus Eutypella sp | halotolerant | Enterococci S. aureus | [139] |
| Bisvertinolone | Aspergillus versicolor | halophilic | S. aureus | [125] |
| Bisdechlorogeodin | Pseudogymnoascus sp. LAMAI 2784 isolated from Antarctic soil | Cold-adapted Antarctic soil fungi | Xanthomonas citri subsp. citri | [140] |
| Pseudogymnoascus sp. | [135] | |||
| Amphiol | Pseudogymnoascus sp. | Cold-adapted Antarctic soil fungi | antifungal effect | [141] |
| Exopolysaccharides | Lecanicillium muscarium and Thelebolus microsporus | Antarctic fungi, cold-adapted | antibacterial and antifungal effect | [142] |
| Characteristic | Antibiotics | Other Antimicrobial Compounds from Filamentous Fungi |
|---|---|---|
| Application | Clinical | Clinical, Agricultural, Industrial |
| Synthesis | Secondary metabolites | Secondary metabolites |
| Bioactivity spectra | Mostly broad | Narrow to broad, depending on the compound |
| Intensity of bioactivity | Active at micro-to-milli molar range | Often active at nano-to-micro molar range |
| Proteolytic enzyme degradability | Moderate to none | Highly variable (some highly stable, others degradable) |
| Thermal stability | Low | Compound-dependent; often moderate to high |
| Active pH range | Narrow | Wide (many remain active across broader pH ranges) |
| Color/taste/odor | Yes | Often distinct and compound-specific |
| Amenability to bioengineering | No | Limited, though possible via synthetic biology approaches |
| Possible mechanism of target cell developing resistance | Genetically transferable determinant that inactivates the active compound | Membrane modification, efflux pumps, enzymatic degradation, oxidative stress adaptation |
| Mode of action | Cell membrane or intercellular targets | Diverse: membrane disruption, enzyme inhibition, ROS generation, DNA/protein synthesis inhibition |
| Toxicity towards eukaryotic cells | Yes | Variable—some are highly toxic (e.g., mycotoxins), others have selective activity |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dishliyska, V.; Miteva-Staleva, J.; Gocheva, Y.; Stoyancheva, G.; Yovchevska, L.; Abrashev, R.; Spasova, B.; Angelova, M.; Krumova, E. Biological Potential of Extremophilic Filamentous Fungi for the Production of New Compounds with Antimicrobial Effect. Fermentation 2025, 11, 347. https://doi.org/10.3390/fermentation11060347
Dishliyska V, Miteva-Staleva J, Gocheva Y, Stoyancheva G, Yovchevska L, Abrashev R, Spasova B, Angelova M, Krumova E. Biological Potential of Extremophilic Filamentous Fungi for the Production of New Compounds with Antimicrobial Effect. Fermentation. 2025; 11(6):347. https://doi.org/10.3390/fermentation11060347
Chicago/Turabian StyleDishliyska, Vladislava, Jeny Miteva-Staleva, Yana Gocheva, Galina Stoyancheva, Lyudmila Yovchevska, Radoslav Abrashev, Boryana Spasova, Maria Angelova, and Ekaterina Krumova. 2025. "Biological Potential of Extremophilic Filamentous Fungi for the Production of New Compounds with Antimicrobial Effect" Fermentation 11, no. 6: 347. https://doi.org/10.3390/fermentation11060347
APA StyleDishliyska, V., Miteva-Staleva, J., Gocheva, Y., Stoyancheva, G., Yovchevska, L., Abrashev, R., Spasova, B., Angelova, M., & Krumova, E. (2025). Biological Potential of Extremophilic Filamentous Fungi for the Production of New Compounds with Antimicrobial Effect. Fermentation, 11(6), 347. https://doi.org/10.3390/fermentation11060347