
Elucidating Key Microbial Drivers for Methane Production during Cold Adaptation and Psychrophilic Anaerobic Digestion of Cattle Manure and Food Waste
 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Cold-Adapted Inoculum Preparation
2.2. Batch Culture Setup
2.3. Analytical Methods
2.4. Kinetics Modelling
2.5. Next Generation Sequencing of 16S rRNA V4-V5 Amplicons
2.6. Metagenomic Analyses of Sequences
3. Results and Discussion
3.1. Methane Production
3.2. Microbial Community Dynamics
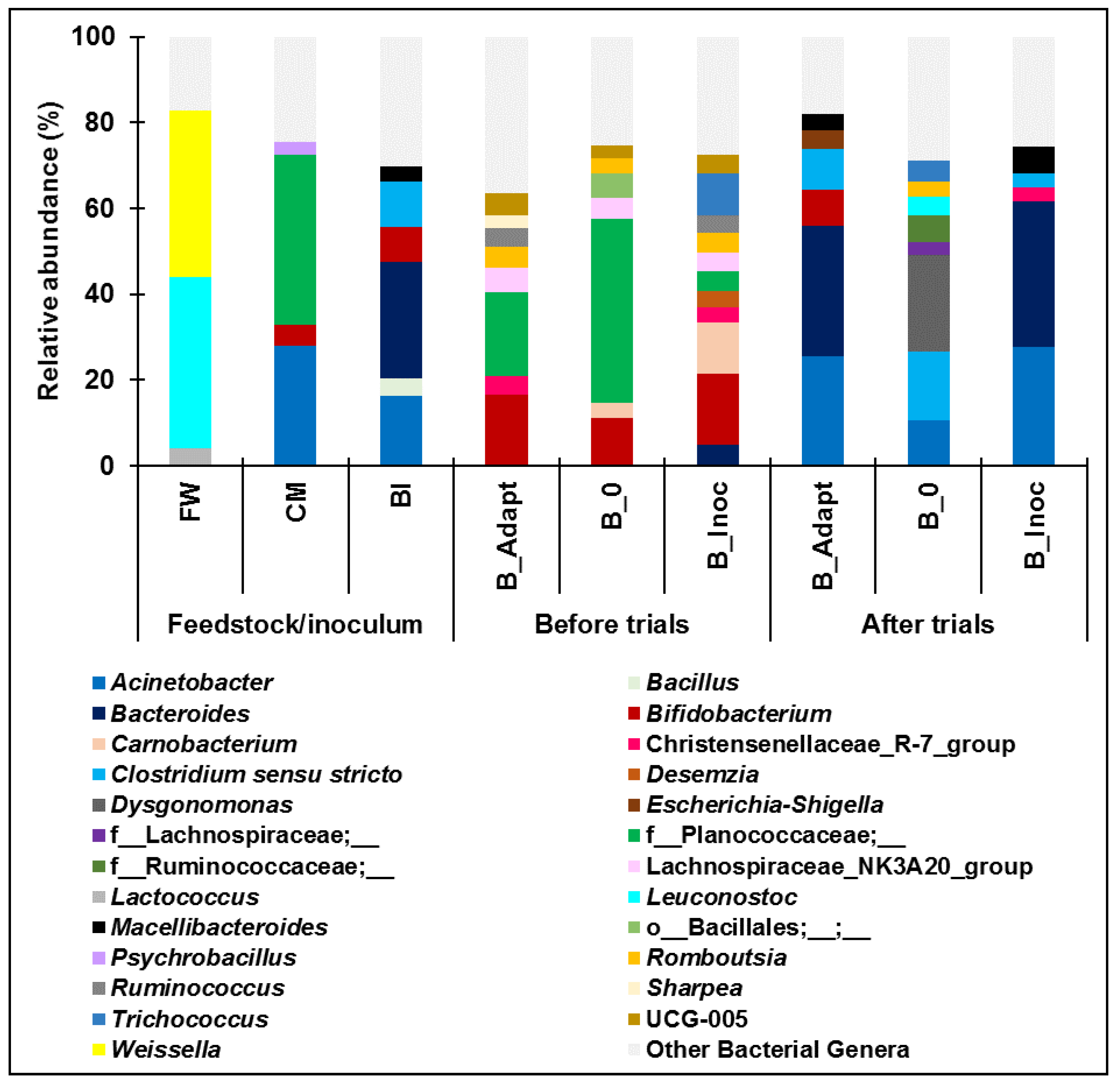
3.2.1. Bacterial Community Composition
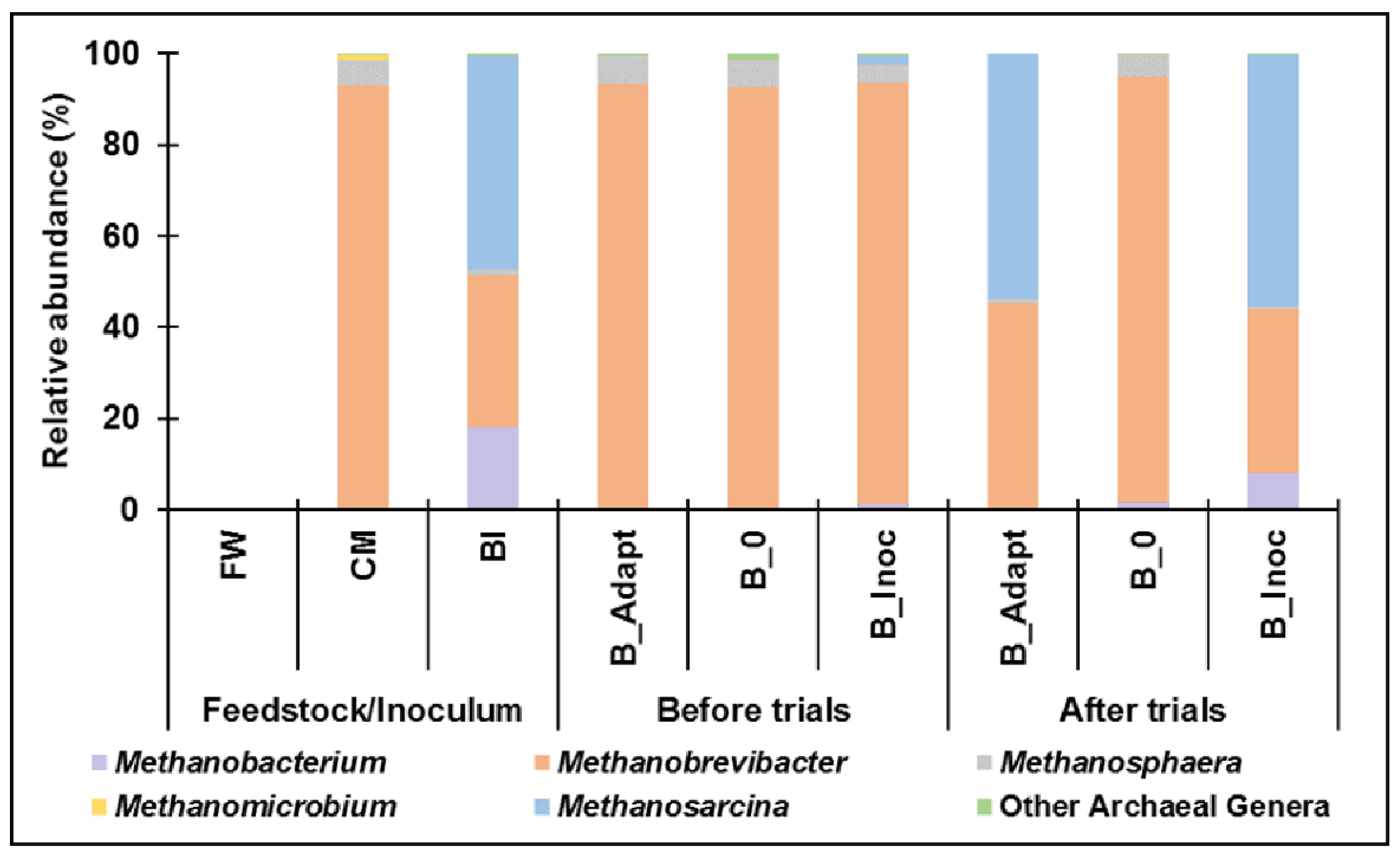
3.2.2. Archaeal Community Composition
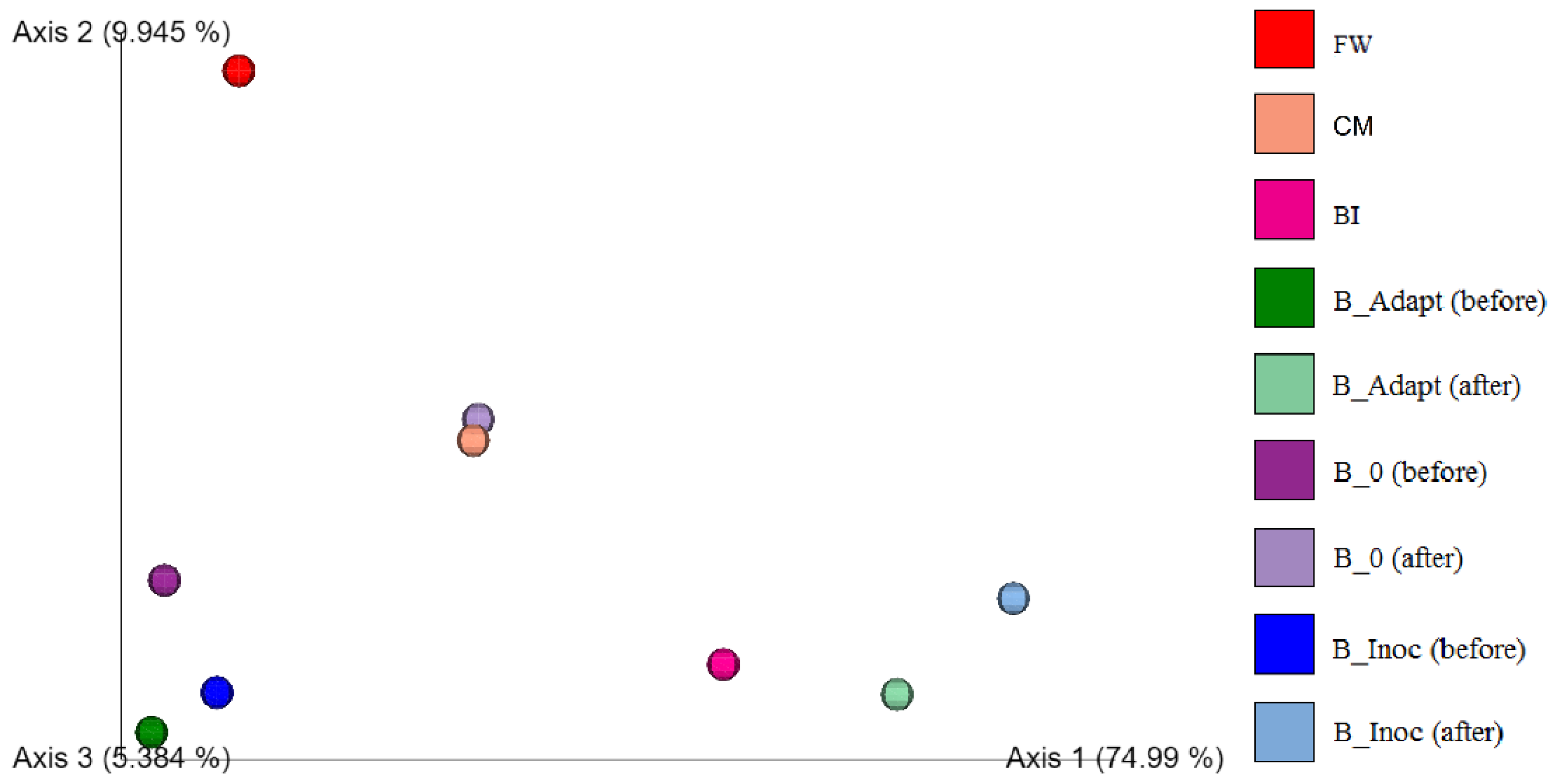
3.2.3. Microbial Diversity
3.2.4. Predicted Functional Capabilities
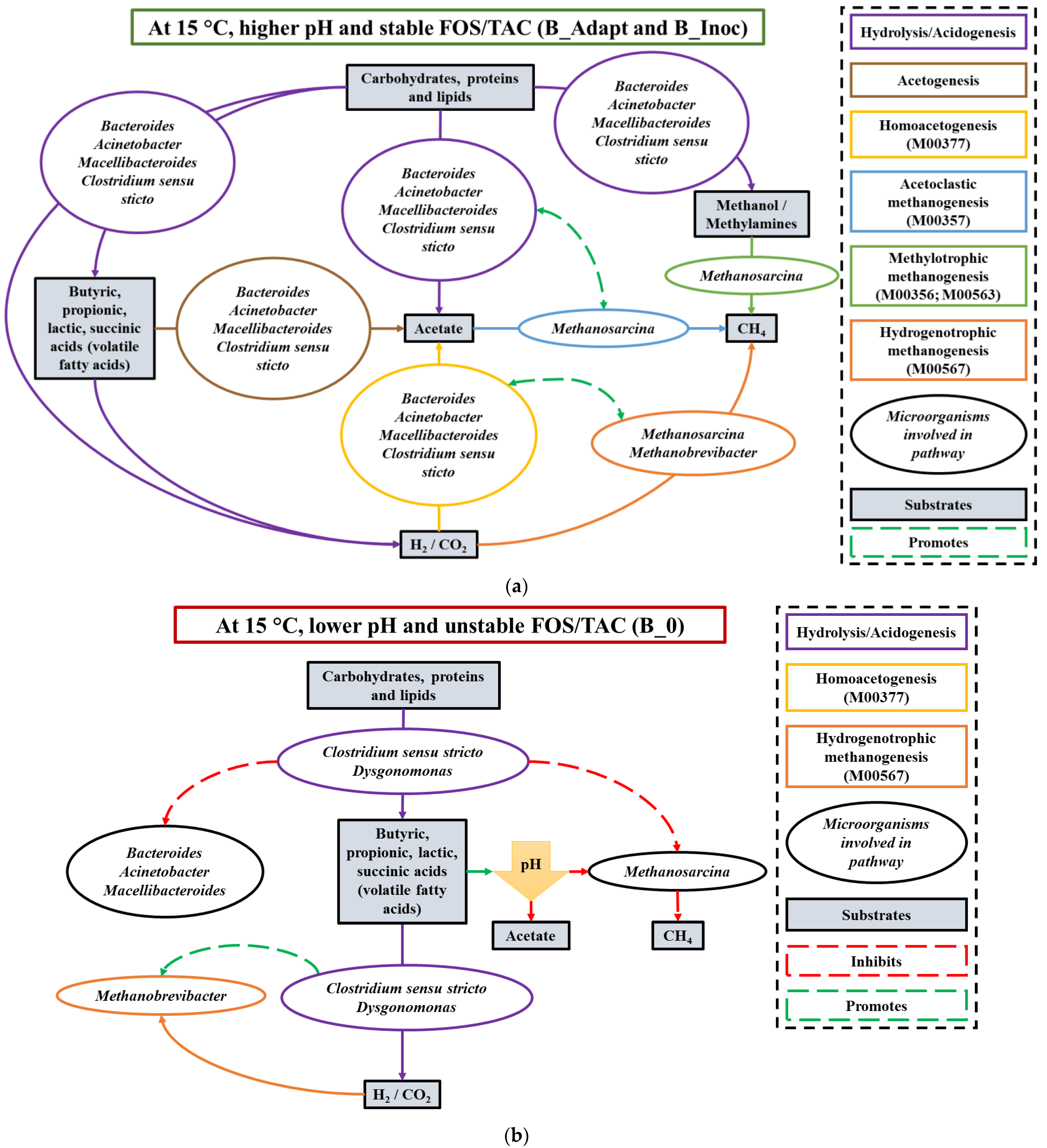
3.2.5. Possible Synergistic Dynamics between Core Microbial Groups
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rama, H.; Akindolire, M.; Obi, L.; Bello-Akinosho, M.; Ndaba, B.; Dhlamini, M.S.; Maaza, M.; Roopnarain, A. Anaerobic Digestion: Climate Change Mitigation through Sustainable Organic Waste Valorization. In Handbook of Nature-Based Solutions to Mitigation and Adaptation to Climate Change; Springer Nature: Cham, Switzerland, 2023; pp. 1–19. ISBN 978-3-030-98067-2. [Google Scholar]
- Searchinger, T.; Herrero, M.; Yan, X.; Wang, J.; Dumas, P.; Beauchemin, K.; Kebreab, E. Opportunities to Reduce Methane Emissions from Global Agriculture; Princeton University: Princeton, NJ, USA; Cornell University: Ithaca, NY, USA, 2021; pp. 1–40. [Google Scholar]
- Xu, X.; Sun, Y.; Sun, Y.; Li, Y. Bioaugmentation Improves Batch Psychrophilic Anaerobic Co-Digestion of Cattle Manure and Corn Straw. Bioresour. Technol. 2022, 343, 126118. [Google Scholar] [CrossRef] [PubMed]
- Tolessa, A.; Zantsi, S.; Louw, T.M.; Greyling, J.C.; Goosen, N.J. Estimation of Biomass Feedstock Availability for Anaerobic Digestion in Smallholder Farming Systems in South Africa. Biomass Bioenergy 2020, 142, 105798. [Google Scholar] [CrossRef]
- Abbasi, T.; Tauseef, S.M.; Abbasi, S.A. Anaerobic Digestion for Global Warming Control and Energy Generation—An Overview. Renew. Sustain. Energy Rev. 2012, 16, 3228–3242. [Google Scholar] [CrossRef]
- Roopnarain, A.; Rama, H.; Ndaba, B.; Bello-Akinosho, M.; Bamuza-Pemu, E.; Adeleke, R. Unravelling the Anaerobic Digestion ‘Black Box’: Biotechnological Approaches for Process Optimization. Renew. Sustain. Energy Rev. 2021, 152, 111717. [Google Scholar] [CrossRef]
- Ricciardi, V.; Ramankutty, N.; Mehrabi, Z.; Jarvis, L.; Chookolingo, B. How Much of the World’s Food Do Smallholders Produce? Glob. Food Secur. 2018, 17, 64–72. [Google Scholar] [CrossRef]
- Pham, C.H.; Triolo, J.M.; Sommer, S.G. Predicting Methane Production in Simple and Unheated Biogas Digesters at Low Temperatures. Appl. Energy 2014, 136, 1–6. [Google Scholar] [CrossRef]
- SANEDI; UJ-TRCTI; UJ-PEETS; SABIA. Sector Development Plan for the Micro-Digester Sector in South Africa: Pathways to Growth and Sustainability by 2030; SANEDI: Johannesburg, South Africa, 2022; pp. 1–19. [Google Scholar]
- Kalina, M.; Ogwang, J.Ò.; Tilley, E. From Potential to Practice: Rethinking Africa’s Biogas Revolution. Humanit. Soc. Sci. Commun. 2022, 9, 374. [Google Scholar] [CrossRef]
- Nape, K.M.; Magama, P.; Moeletsi, M.E.; Tongwane, M.I.; Nakana, P.M.; Mliswa, V.K.; Motsepe, M.; Madikiza, S. Introduction of Household Biogas Digesters in Rural Farming Households of the Maluti-a-Phofung Municipality, South Africa. J. Energy South. Afr. 2019, 30, 28–37. [Google Scholar] [CrossRef]
- Martí-Herrero, J.; Castro, L.; Jaimes-Estévez, J.; Grijalva, M.; Gualatoña, M.; Aldás, M.B.; Escalante, H. Biomethane Potential Test Applied to Psychrophilic Conditions: Three Issues about Inoculum Temperature Adaptation. Bioresour. Technol. Rep. 2022, 20, 101279. [Google Scholar] [CrossRef]
- Akindolire, M.A.; Rama, H.; Roopnarain, A. Psychrophilic Anaerobic Digestion: A Critical Evaluation of Microorganisms and Enzymes to Drive the Process. Renew. Sustain. Energy Rev. 2022, 161, 112394. [Google Scholar] [CrossRef]
- Rodríguez-Jiménez, L.M.; Pérez-Vidal, A.; Torres-Lozada, P. Research Trends and Strategies for the Improvement of Anaerobic Digestion of Food Waste in Psychrophilic Temperatures Conditions. Heliyon 2022, 8, e11174. [Google Scholar] [CrossRef] [PubMed]
- Struvay, C.; Feller, G. Optimization to Low Temperature Activity in Psychrophilic Enzymes. Int. J. Mol. Sci. 2012, 13, 11643–11665. [Google Scholar] [CrossRef] [PubMed]
- Nedwell, D.B. Effect of Low Temperature on Microbial Growth: Lowered Affinity for Substrates Limits Growth at Low Temperature. FEMS Microbiol. Ecol. 1999, 30, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Alavi-Borazjani, S.A.; Capela, I.; Tarelho, L.A.C. Over-Acidification Control Strategies for Enhanced Biogas Production from Anaerobic Digestion: A Review. Biomass Bioenergy 2020, 143, 105833. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, N.; Ma, J.; Zhou, Y.; Wei, Q.; Tian, C.; Fang, Y.; Zhong, R.; Chen, G.; Zhang, S. Advances in Cold-Adapted Enzymes Derived from Microorganisms. Front. Microbiol. 2023, 14, 1152847. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.; Jha, A.K. Psychrophilic Dry Anaerobic Digestion of Cow Dung for Methane Production: Effect of Inoculum. ScienceAsia 2013, 39, 500. [Google Scholar] [CrossRef]
- Xu, X.; Yan, M.; Sun, Y.; Li, Y. Bioaugmentation with Cold-Tolerant Methanogenic Culture to Boost Methane Production from Anaerobic Co-Digestion of Cattle Manure and Corn Straw at 20 °C. Chem. Eng. J. 2023, 466, 143183. [Google Scholar] [CrossRef]
- Liu, Y.-C.; Ramiro-Garcia, J.; Paulo, L.M.; Braguglia, C.M.; Gagliano, M.C.; O’Flaherty, V. Psychrophilic and Mesophilic Anaerobic Treatment of Synthetic Dairy Wastewater with Long Chain Fatty Acids: Process Performances and Microbial Community Dynamics. Bioresour. Technol. 2023, 380, 129124. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, B.R.; Rouissi, T.; Brar, S.K.; Surampalli, R.Y. Critical Insights into Psychrophilic Anaerobic Digestion: Novel Strategies for Improving Biogas Production. Waste Manag. 2021, 131, 513–526. [Google Scholar] [CrossRef]
- Ariunbaatar, J.; Panico, A.; Frunzo, L.; Esposito, G.; Lens, P.N.L.; Pirozzi, F. Enhanced Anaerobic Digestion of Food Waste by Thermal and Ozonation Pretreatment Methods. J. Environ. Manag. 2014, 146, 142–149. [Google Scholar] [CrossRef]
- Mukhuba, M.; Roopnarain, A.; Moeletsi, M.E.; Adeleke, R. Metagenomic Insights into the Microbial Community and Biogas Production Pattern during Anaerobic Digestion of Cow Dung and Mixed Food Waste. J. Chem. Technol. Biotechnol. 2020, 95, 151–162. [Google Scholar] [CrossRef]
- Matsabisa, M.G.; Bala, A.; Tripathy, S.; Digashu, M.M.; Rautenbach, F.; Dassarma, B.; Erhabor, J.O.; Braga, F.C.; Mukherjee, P.K.; Tang, M.; et al. Study on South African Indigenous Teas—Antioxidant Potential, Nutritional Content, and Hypoxia-Induced Cyclooxygenase Inhibition on U87 MG Cell Line. Molecules 2022, 27, 3505. [Google Scholar] [CrossRef] [PubMed]
- Nkuna, R.; Roopnarain, A.; Adeleke, R. Effects of Organic Loading Rates on Microbial Communities and Biogas Production from Water Hyacinth: A Case of Mono- and Co-digestion. J Chem. Tech Biotech 2019, 94, 1294–1304. [Google Scholar] [CrossRef]
- SAS Institute, Inc. SAS/STAT User’s Guide, SAS 9.4 for Windows, 5th ed.; SAS Institute, Inc.: Cary, NC, USA, 2016. [Google Scholar]
- Obi, L.U.; Roopnarain, A.; Tekere, M.; Adeleke, R.A. Bioaugmentation Potential of Inoculum Derived from Anaerobic Digestion Feedstock for Enhanced Methane Production Using Water Hyacinth. World J. Microbiol. Biotechnol. 2023, 39, 153. [Google Scholar] [CrossRef] [PubMed]
- Fadeev, E.; Cardozo-Mino, M.G.; Rapp, J.Z.; Bienhold, C.; Salter, I.; Salman-Carvalho, V.; Molari, M.; Tegetmeyer, H.E.; Buttigieg, P.L.; Boetius, A. Comparison of Two 16S rRNA Primers (V3–V4 and V4–V5) for Studies of Arctic Microbial Communities. Front. Microbiol. 2021, 12, 637526. [Google Scholar] [CrossRef] [PubMed]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data 2010. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 17 July 2024).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, Interactive, Scalable and Extensible Microbiome Data Science Using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-Resolution Sample Inference from Illumina Amplicon Data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M. Toward Understanding the Origin and Evolution of Cellular Organisms. Protein Sci. 2019, 28, 1947–1951. [Google Scholar] [CrossRef]
- Kanehisa, M.; Furumichi, M.; Sato, Y.; Kawashima, M.; Ishiguro-Watanabe, M. KEGG for Taxonomy-Based Analysis of Pathways and Genomes. Nucleic Acids Res. 2023, 51, D587–D592. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- RStudio Team. RStudio: Integrated Development for R; RStudio, PBC: Boston, MA, USA, 2020. [Google Scholar]
- Wemheuer, F.; Taylor, J.A.; Daniel, R.; Johnston, E.; Meinicke, P.; Thomas, T.; Wemheuer, B. Tax4Fun2: Prediction of Habitat-Specific Functional Profiles and Functional Redundancy Based on 16S rRNA Gene Sequences. Environ. Microbiome 2020, 15, 11. [Google Scholar] [CrossRef] [PubMed]
- Keating, C.; Hughes, D.; Mahony, T.; Cysneiros, D.; Ijaz, U.Z.; Smith, C.J.; O’Flaherty, V. Cold Adaptation and Replicable Microbial Community Development during Long-Term Low-Temperature Anaerobic Digestion Treatment of Synthetic Sewage. FEMS Microbiol. Ecol. 2018, 94, fiy095. [Google Scholar] [CrossRef] [PubMed]
- Casallas-Ojeda, M.; Cabeza, I.; Sanchez, N.; Caicedo-Concha, D.M.; Astals, S. Cheese Whey and Dairy Manure Anaerobic Co-Digestion at Psychrophilic Conditions: Technical and Environmental Evaluation. Environ. Res. 2024, 251, 118525. [Google Scholar] [CrossRef] [PubMed]
- Esparza-Soto, M.; Alcaraz-Ibarra, S.; Lucero-Chavez, M.; Jimenez-Moleon, M.D.C.; Mier-Quiroga, M.D.L.A.; Fall, C. Enhancing Kitchen Waste Minimization and Energy Generation at 20 °C: A Psychrophilic Anaerobic Co-digestion Study. J. Chem. Tech. Biotech. 2024, 99, 522–530. [Google Scholar] [CrossRef]
- Meegoda, J.; Li, B.; Patel, K.; Wang, L. A Review of the Processes, Parameters, and Optimization of Anaerobic Digestion. Int. J. Environ. Res. Public Health 2018, 15, 2224. [Google Scholar] [CrossRef] [PubMed]
- Zhou, A.; Liu, H.; Varrone, C.; Shyryn, A.; Defemur, Z.; Wang, S.; Liu, W.; Yue, X. New Insight into Waste Activated Sludge Acetogenesis Triggered by Coupling Sulfite/Ferrate Oxidation with Sulfate Reduction-Mediated Syntrophic Consortia. Chem. Eng. J. 2020, 400, 125885. [Google Scholar] [CrossRef]
- Jabari, L.; Gannoun, H.; Cayol, J.-L.; Hedi, A.; Sakamoto, M.; Falsen, E.; Ohkuma, M.; Hamdi, M.; Fauque, G.; Ollivier, B.; et al. Macellibacteroides Fermentans Gen. Nov., Sp. Nov., a Member of the Family Porphyromonadaceae Isolated from an Upflow Anaerobic Filter Treating Abattoir Wastewaters. Int. J. Syst. Evol. Microbiol. 2012, 62, 2522–2527. [Google Scholar] [CrossRef] [PubMed]
- Nishiyama, T.; Ueki, A.; Kaku, N.; Watanabe, K.; Ueki, K. Bacteroides Graminisolvens Sp. Nov., a Xylanolytic Anaerobe Isolated from a Methanogenic Reactor Treating Cattle Waste. Int. J. Syst. Evol. Microbiol. 2009, 59, 1901–1907. [Google Scholar] [CrossRef]
- Ueki, A.; Abe, K.; Ohtaki, Y.; Kaku, N.; Watanabe, K.; Ueki, K. Bacteroides Paurosaccharolyticus Sp. Nov., Isolated from a Methanogenic Reactor Treating Waste from Cattle Farms. Int. J. Syst. Evol. Microbiol. 2011, 61, 448–453. [Google Scholar] [CrossRef]
- Pulami, D.; Schauss, T.; Eisenberg, T.; Blom, J.; Schwengers, O.; Bender, J.K.; Wilharm, G.; Kämpfer, P.; Glaeser, S.P. Acinetobacter Stercoris Sp. Nov. Isolated from Output Source of a Mesophilic German Biogas Plant with Anaerobic Operating Conditions. Antonie Van Leeuwenhoek 2021, 114, 235–251. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.H.; Hao, O.J.; Wang, N.S. Acinetobacter Isolates from Different Activated Sludge Processes: Characteristics and Neural Network Identification. FEMS Microbiol. Ecol. 1997, 23, 217–227. [Google Scholar] [CrossRef]
- Ryan, R.P.; Monchy, S.; Cardinale, M.; Taghavi, S.; Crossman, L.; Avison, M.B.; Berg, G.; Van Der Lelie, D.; Dow, J.M. The Versatility and Adaptation of Bacteria from the Genus Stenotrophomonas. Nat. Rev. Microbiol. 2009, 7, 514–525. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Zaiden, N.; Cao, B. The Core- and Pan-Genomic Analyses of the Genus Comamonas: From Environmental Adaptation to Potential Virulence. Front. Microbiol. 2018, 9, 3096. [Google Scholar] [CrossRef] [PubMed]
- Kelly, S.M.; Munoz-Munoz, J.; Van Sinderen, D. Plant Glycan Metabolism by Bifidobacteria. Front. Microbiol. 2021, 12, 609418. [Google Scholar] [CrossRef] [PubMed]
- Bridges, C.M.; Gage, D.J. Development and Application of Aerobic, Chemically Defined Media for Dysgonomonas. Anaerobe 2021, 67, 102302. [Google Scholar] [CrossRef]
- Hofstad, T.; Olsen, I.; Eribe, E.R.; Falsen, E.; Collins, M.D.; Lawson, P.A. Dysgonomonas Gen. Nov. to Accommodate Dysgonomonas Gadei Sp. Nov., an Organism Isolated from a Human Gall Bladder, and Dysgonomonas Capnocytophagoides (Formerly CDC Group DF-3). Int. J. Syst. Evol. Microbiol. 2000, 50, 2189–2195. [Google Scholar] [CrossRef] [PubMed]
- Alou, M.T.; Ndongo, S.; Frégère, L.; Labas, N.; Andrieu, C.; Richez, M.; Couderc, C.; Baudoin, J.-P.; Abrahão, J.; Brah, S.; et al. Taxonogenomic Description of Four New Clostridium Species Isolated from Human Gut: ‘Clostridium Amazonitimonense’, ‘Clostridium Merdae’, ‘Clostridium Massilidielmoense’ and ‘Clostridium Nigeriense’. New Microbes New Infect. 2018, 21, 128–139. [Google Scholar] [CrossRef] [PubMed]
- Liberato, V.; Benevenuti, C.; Coelho, F.; Botelho, A.; Amaral, P.; Pereira, N.; Ferreira, T. Clostridium Sp. as Bio-Catalyst for Fuels and Chemicals Production in a Biorefinery Context. Catalysts 2019, 9, 962. [Google Scholar] [CrossRef]
- Sun, M.; Yanagawa, K.; Prasitwuttisak, W.; Goel, R.; Watanabe, R.; Harada, H.; Liu, B.; Terashima, M.; Yasui, H. Kinetics for the Methanogen’s Death in the Acidic Environments. J. Wat. Environ. Tech. 2023, 21, 59–75. [Google Scholar] [CrossRef]
- Roopnarain, A.; Akindolire, M.A.; Rama, H.; Ndaba, B. Casting Light on the Micro-Organisms in Digestate: Diversity and Untapped Potential. Fermentation 2023, 9, 160. [Google Scholar] [CrossRef]
- Jetten, M.S.M.; Stams, A.J.M.; Zehnder, A.J.B. Acetate Threshold Values and Acetate Activating Enzymes in Methanogenic Bacteria. FEMS Microbiol. Lett. 1990, 73, 339–344. [Google Scholar] [CrossRef]
- Dev, S.; Saha, S.; Kurade, M.B.; Salama, E.-S.; El-Dalatony, M.M.; Ha, G.-S.; Chang, S.W.; Jeon, B.-H. Perspective on Anaerobic Digestion for Biomethanation in Cold Environments. Renew. Sustain. Energy Rev. 2019, 103, 85–95. [Google Scholar] [CrossRef]
- Kurth, J.M.; Op Den Camp, H.J.M.; Welte, C.U. Several Ways One Goal—Methanogenesis from Unconventional Substrates. Appl. Microbiol. Biotechnol. 2020, 104, 6839–6854. [Google Scholar] [CrossRef] [PubMed]
- Hagen, L.H.; Vivekanand, V.; Pope, P.B.; Eijsink, V.G.H.; Horn, S.J. The Effect of Storage Conditions on Microbial Community Composition and Biomethane Potential in a Biogas Starter Culture. Appl. Microbiol. Biotechnol. 2015, 99, 5749–5761. [Google Scholar] [CrossRef]
- Jost, L. The Relation between Evenness and Diversity. Diversity 2010, 2, 207–232. [Google Scholar] [CrossRef]
- Kim, B.-R.; Shin, J.; Guevarra, R.B.; Lee, J.H.; Kim, D.W.; Seol, K.-H.; Lee, J.-H.; Kim, H.B.; Isaacson, R.E. Deciphering Diversity Indices for a Better Understanding of Microbial Communities. J. Microbiol. Biotechnol. 2017, 27, 2089–2093. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Ma, F.; Ma, W.; Wang, P.; Zhao, G.; Lu, X. Influence of Temperature on Biogas Production Efficiency and Microbial Community in a Two-Phase Anaerobic Digestion System. Water 2019, 11, 133. [Google Scholar] [CrossRef]
- Yu, X.; Polz, M.F.; Alm, E.J. Interactions in Self-Assembled Microbial Communities Saturate with Diversity. ISME J. 2019, 13, 1602–1617. [Google Scholar] [CrossRef]
- Lozupone, C.; Lladser, M.E.; Knights, D.; Stombaugh, J.; Knight, R. UniFrac: An Effective Distance Metric for Microbial Community Comparison. ISME J. 2011, 5, 169–172. [Google Scholar] [CrossRef]
- Singh, S.; Keating, C.; Ijaz, U.Z.; Hassard, F. Molecular Insights Informing Factors Affecting Low Temperature Anaerobic Applications: Diversity, Collated Core Microbiomes and Complexity Stability Relationships in LCFA-Fed Systems. Sci. Total Environ. 2023, 874, 162420. [Google Scholar] [CrossRef] [PubMed]
- McKeown, R.M.; Scully, C.; Enright, A.-M.; Chinalia, F.A.; Lee, C.; Mahony, T.; Collins, G.; O’Flaherty, V. Psychrophilic Methanogenic Community Development during Long-Term Cultivation of Anaerobic Granular Biofilms. ISME J. 2009, 3, 1231–1242. [Google Scholar] [CrossRef] [PubMed]
- Yun, H.; Liang, B.; He, Z.; Li, M.; Zong, S.; Wang, Z.; Ge, B.; Zhang, P.; Li, X.; Wang, A. Insights into Methanogenesis of Mesophilic-Psychrophilic Varied Anaerobic Digestion of Municipal Sludge with Antibiotic Stress. J. Environ. Manag. 2023, 331, 117278. [Google Scholar] [CrossRef] [PubMed]
- Catlett, J.L.; Carr, S.; Cashman, M.; Smith, M.D.; Walter, M.; Sakkaff, Z.; Kelley, C.; Pierobon, M.; Cohen, M.B.; Buan, N.R. Metabolic Synergy between Human Symbionts Bacteroides and Methanobrevibacter. Microbiol. Spectr. 2022, 10, e01067-22. [Google Scholar] [CrossRef] [PubMed]
- Catlett, J.L.; Catazaro, J.; Cashman, M.; Carr, S.; Powers, R.; Cohen, M.B.; Buan, N.R. Metabolic Feedback Inhibition Influences Metabolite Secretion by the Human Gut Symbiont Bacteroides Thetaiotaomicron. mSystems 2020, 5, e00252-20. [Google Scholar] [CrossRef] [PubMed]
- Samuel, B.S.; Hansen, E.E.; Manchester, J.K.; Coutinho, P.M.; Henrissat, B.; Fulton, R.; Latreille, P.; Kim, K.; Wilson, R.K.; Gordon, J.I. Genomic and Metabolic Adaptations of Methanobrevibacter Smithii to the Human Gut. Proc. Natl. Acad. Sci. USA 2007, 104, 10643–10648. [Google Scholar] [CrossRef] [PubMed]
- Akila, G.; Chandra, T.S. Stimulation of Biomethanation by Clostridium Sp. PXYL1 in Coculture with a Methanosarcina Strain PMET1 at Psychrophilic Temperatures: Stimulation of Biomethanation by Coculture. J. Appl. Microbiol. 2010, 108, 204–213. [Google Scholar] [CrossRef]
- Lu, H.; Ng, S.-K.; Jia, Y.; Cai, M.; Lee, P.K.H. Physiological and Molecular Characterizations of the Interactions in Two Cellulose-to-Methane Cocultures. Biotechnol. Biofuels 2017, 10, 37. [Google Scholar] [CrossRef]
- Tomita, H.; Okazaki, F.; Tamaru, Y. Biomethane Production from Sugar Beet Pulp under Cocultivation with Clostridium Cellulovorans and Methanogens. AMB Express 2019, 9, 28. [Google Scholar] [CrossRef]
- Kotsyurbenko, O.R. Trophic Interactions in the Methanogenic Microbial Community of Low-Temperature Terrestrial Ecosystems. FEMS Microbiol. Ecol. 2005, 53, 3–13. [Google Scholar] [CrossRef]
- Dyksma, S.; Jansen, L.; Gallert, C. Syntrophic Acetate Oxidation Replaces Acetoclastic Methanogenesis during Thermophilic Digestion of Biowaste. Microbiome 2020, 8, 105. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Martinez, A.; Sihvonen, M.; Muñoz-Palazon, B.; Rodriguez-Sanchez, A.; Mikola, A.; Vahala, R. Microbial Ecology of Full-Scale Wastewater Treatment Systems in the Polar Arctic Circle: Archaea, Bacteria and Fungi. Sci. Rep. 2018, 8, 2208. [Google Scholar] [CrossRef] [PubMed]
- Kannan, A.D.; Evans, P.; Parameswaran, P. Long-Term Microbial Community Dynamics in a Pilot-Scale Gas Sparged Anaerobic Membrane Bioreactor Treating Municipal Wastewater under Seasonal Variations. Bioresour. Technol. 2020, 310, 123425. [Google Scholar] [CrossRef]
- Ciotola, R.; Martin, J.; Castańo, J.; Lee, J.; Michel, F. Microbial Community Response to Seasonal Temperature Variation in a Small-Scale Anaerobic Digester. Energies 2013, 6, 5182–5199. [Google Scholar] [CrossRef]
- Karekar, S.; Stefanini, R.; Ahring, B. Homo-Acetogens: Their Metabolism and Competitive Relationship with Hydrogenotrophic Methanogens. Microorganisms 2022, 10, 397. [Google Scholar] [CrossRef]
- Pan, X.; Zhao, L.; Li, C.; Angelidaki, I.; Lv, N.; Ning, J.; Cai, G.; Zhu, G. Deep Insights into the Network of Acetate Metabolism in Anaerobic Digestion: Focusing on Syntrophic Acetate Oxidation and Homoacetogenesis. Water Res. 2021, 190, 116774. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Food Waste | Cattle Manure | Cold-Adapted Inoculum 1 |
|---|---|---|---|
| TS (%) | 6.62 | 16.11 | 3.19 |
| VS (%) | 79.83 | 80.99 | 90.70 |
| pH 2 | 4.25 | 8.11 | 7.05 |
| C/N ratio | 17.11 | 19.66 | 20.90 |
| Lipids (%) | 1.08 | 2.44 | ND |
| Proteins (%) | 13.95 | 11.22 | ND |
| Carbohydrates (%) | 7.71 | 11.86 | ND |
| Substrate Composition | P (mL CH4 g−1 VS) | Rm (mL CH4 g−1 VS d−1) | λ (Days) | R2 |
|---|---|---|---|---|
| B_Adapt (cattle manure and food waste) | 160.538 | 3.587 | 42 | 0.98 |
| B_Inoc (cattle manure, food waste and cold-adapted inoculum) | 80.803 | 2.538 | 35 | 0.98 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rama, H.; Ndaba, B.; Dhlamini, M.S.; Cochrane, N.; Maaza, M.; Roopnarain, A. Elucidating Key Microbial Drivers for Methane Production during Cold Adaptation and Psychrophilic Anaerobic Digestion of Cattle Manure and Food Waste. Fermentation 2024, 10, 370. https://doi.org/10.3390/fermentation10070370
Rama H, Ndaba B, Dhlamini MS, Cochrane N, Maaza M, Roopnarain A. Elucidating Key Microbial Drivers for Methane Production during Cold Adaptation and Psychrophilic Anaerobic Digestion of Cattle Manure and Food Waste. Fermentation. 2024; 10(7):370. https://doi.org/10.3390/fermentation10070370
Chicago/Turabian StyleRama, Haripriya, Busiswa Ndaba, Mokhotjwa Simon Dhlamini, Nicolene Cochrane, Malik Maaza, and Ashira Roopnarain. 2024. "Elucidating Key Microbial Drivers for Methane Production during Cold Adaptation and Psychrophilic Anaerobic Digestion of Cattle Manure and Food Waste" Fermentation 10, no. 7: 370. https://doi.org/10.3390/fermentation10070370
APA StyleRama, H., Ndaba, B., Dhlamini, M. S., Cochrane, N., Maaza, M., & Roopnarain, A. (2024). Elucidating Key Microbial Drivers for Methane Production during Cold Adaptation and Psychrophilic Anaerobic Digestion of Cattle Manure and Food Waste. Fermentation, 10(7), 370. https://doi.org/10.3390/fermentation10070370