
Production, Characterization Purification, and Antitumor Activity of L-Asparaginase from Aspergillus niger

 , , , , , , and
, , , , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Comparative Study of Different Fungi in the Production of L-Asparaginase
2.2.1. Solid-State Fermentation (SsF) of L-Asparaginase
2.2.2. Submerged Fermentation (SmF) of L-Asparaginase
2.3. Fungal Culture and Isolation
2.3.1. DNA Extraction
2.3.2. Primers and Polymerase Chain Reaction (PCR)
2.4. Purification of Crude L-Asparaginase Extracts
2.5. Protein Concentration
2.6. Determination of L-Asparaginase Activity
2.7. Biochemical Characterization of Purified L-Asparaginase from Fungus Extract
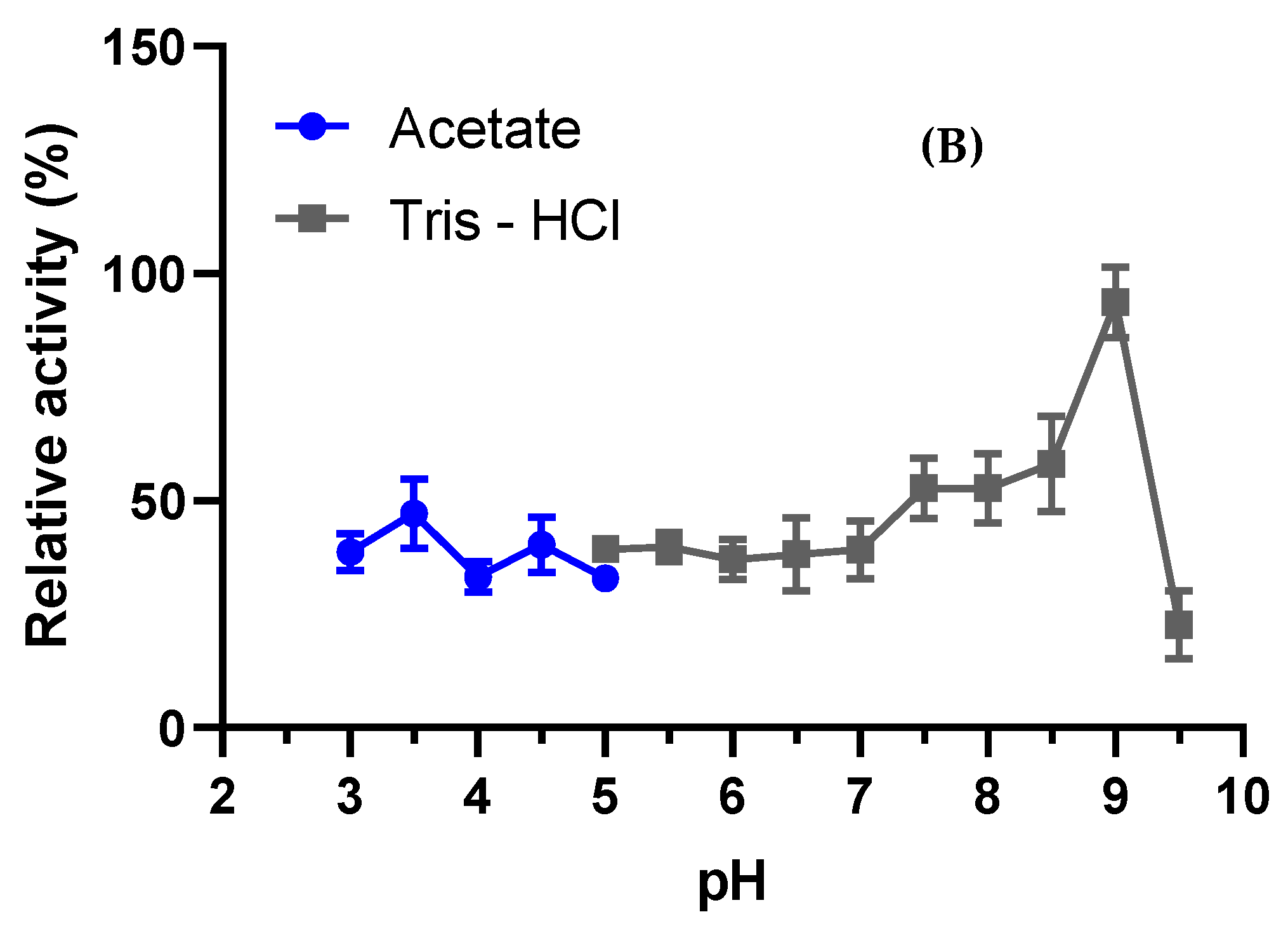
2.7.1. Fungi Optimal pH and Stability
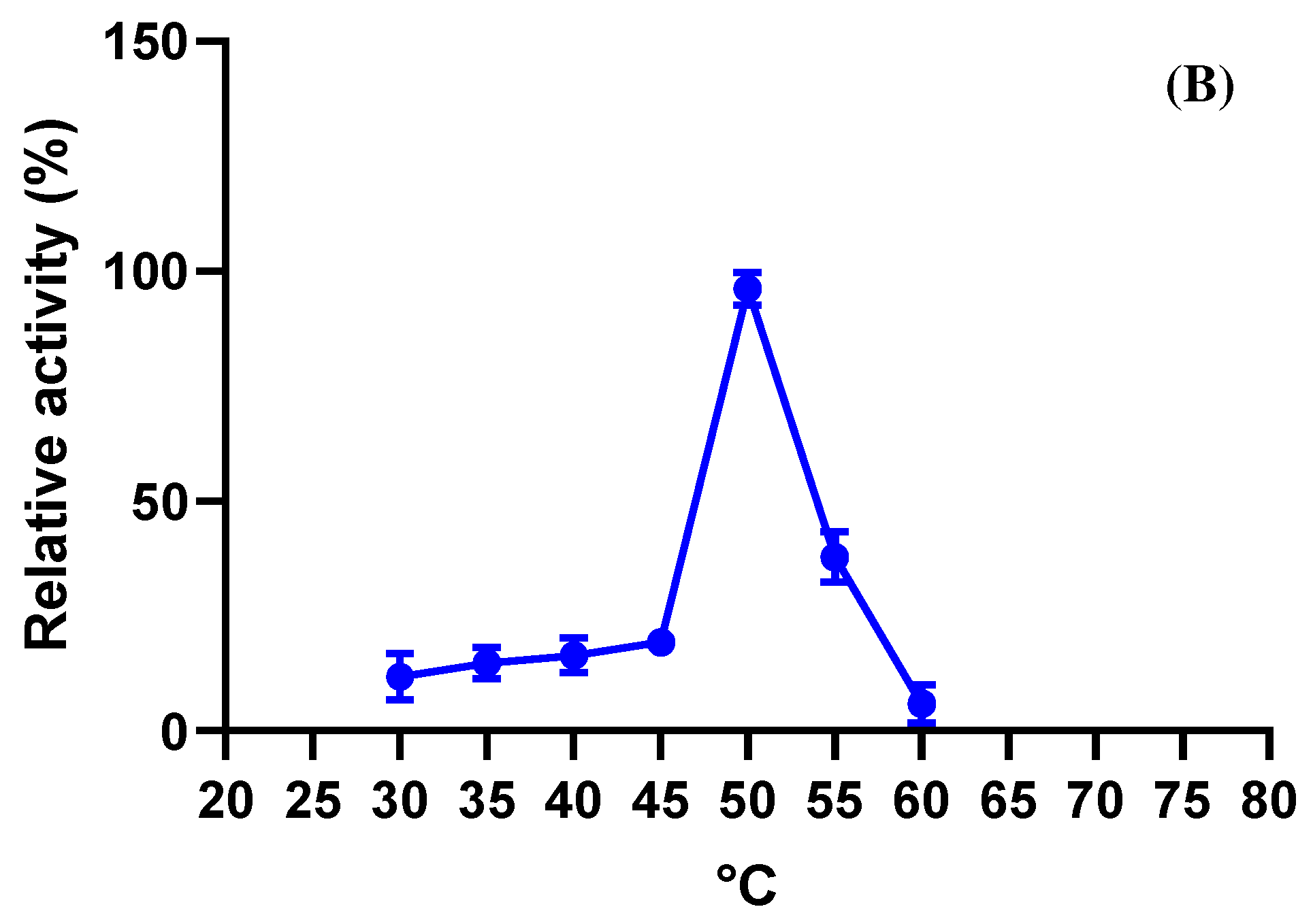
2.7.2. Optimal Temperature and Stability
2.7.3. Effect of Surfactants and Metal Ions in Salts
2.7.4. Stability in Organic Solvents
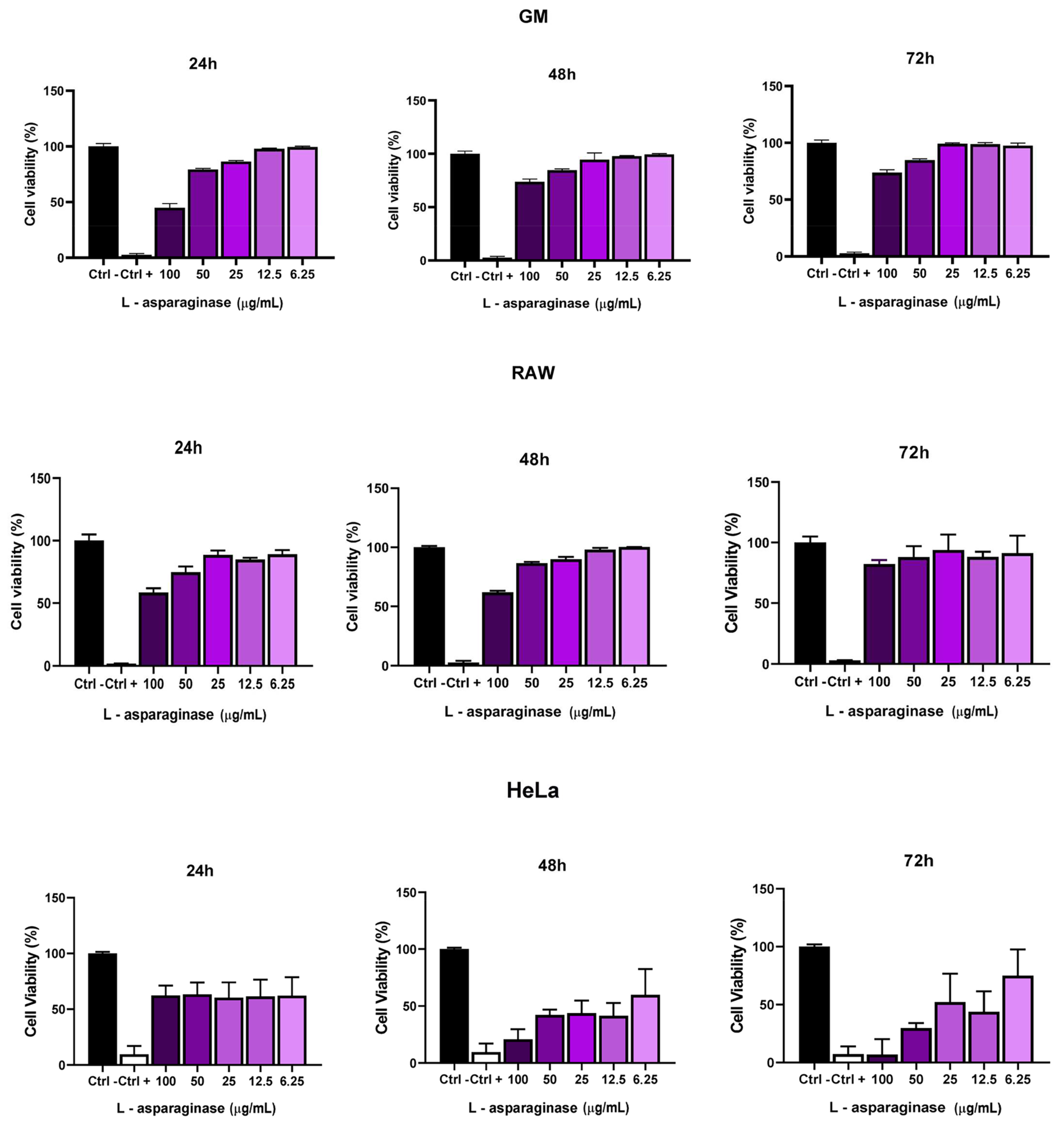
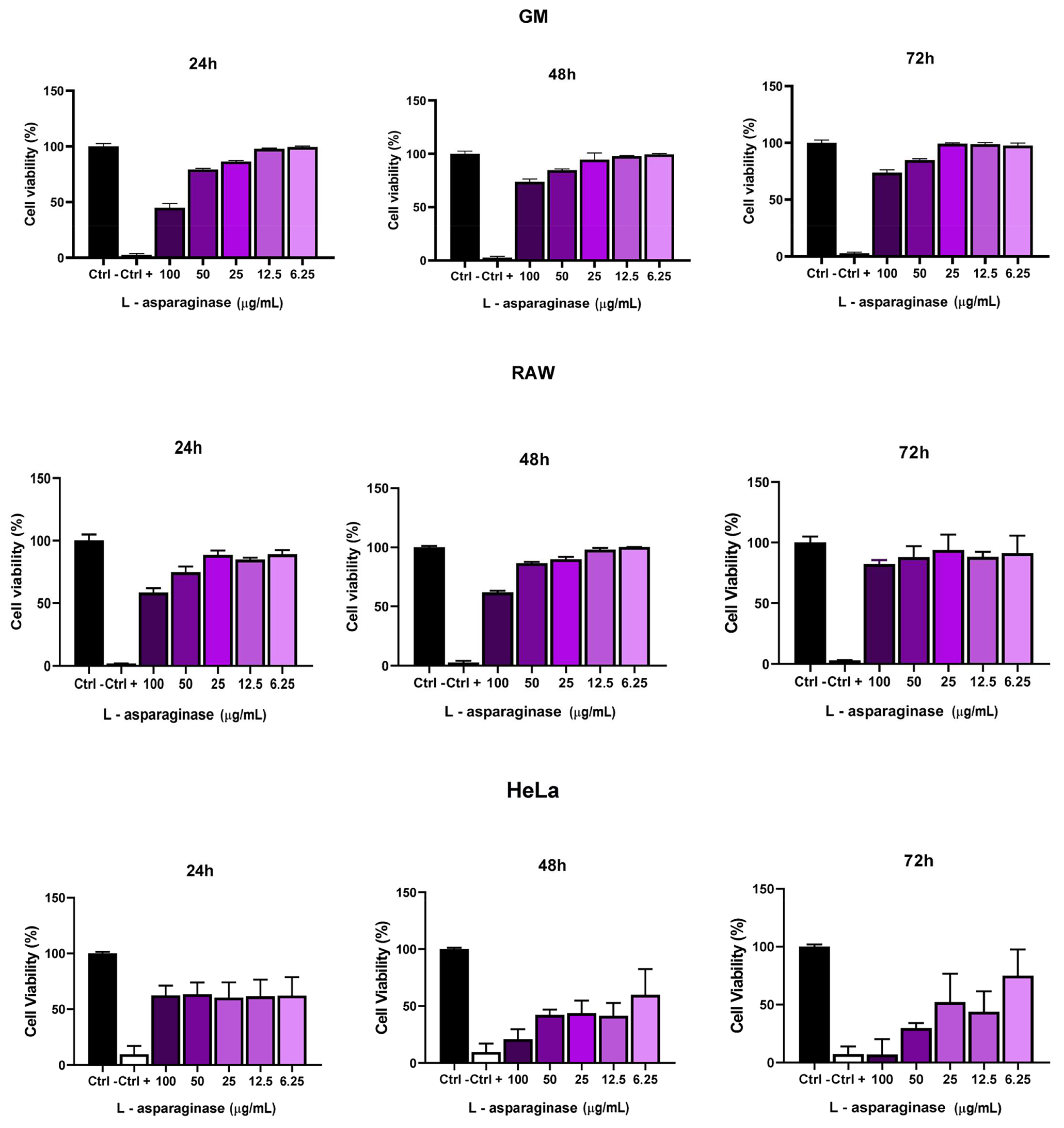
2.8. Cytotoxic Activity of L-Asparaginase
2.9. Morphological Viability in HeLa Cells by Inverted Light Microscopy
2.10. Statistical Analysis
3. Results
3.1. Tracking and Optimization of L-Asparaginase Production by Fungi
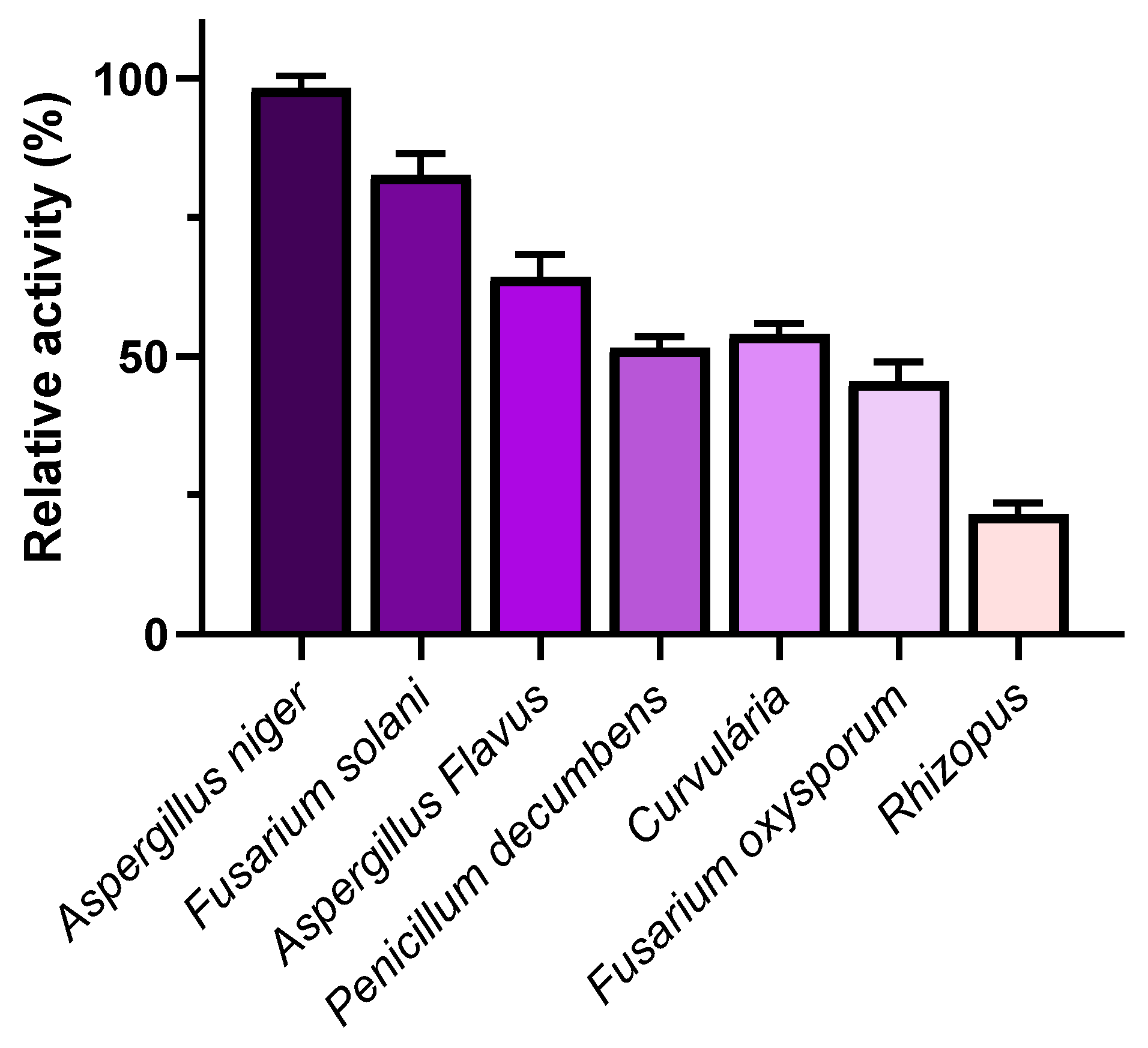
3.1.1. Screening for L-Asparaginase Producers
3.1.2. Fermentation Type for L-Asparaginase Production
3.2. Molecular Identification of A. niger
3.3. Precipitant Type for L-Asparaginase Purification
3.4. Biochemical Characterization of L-Asparaginase from A. niger
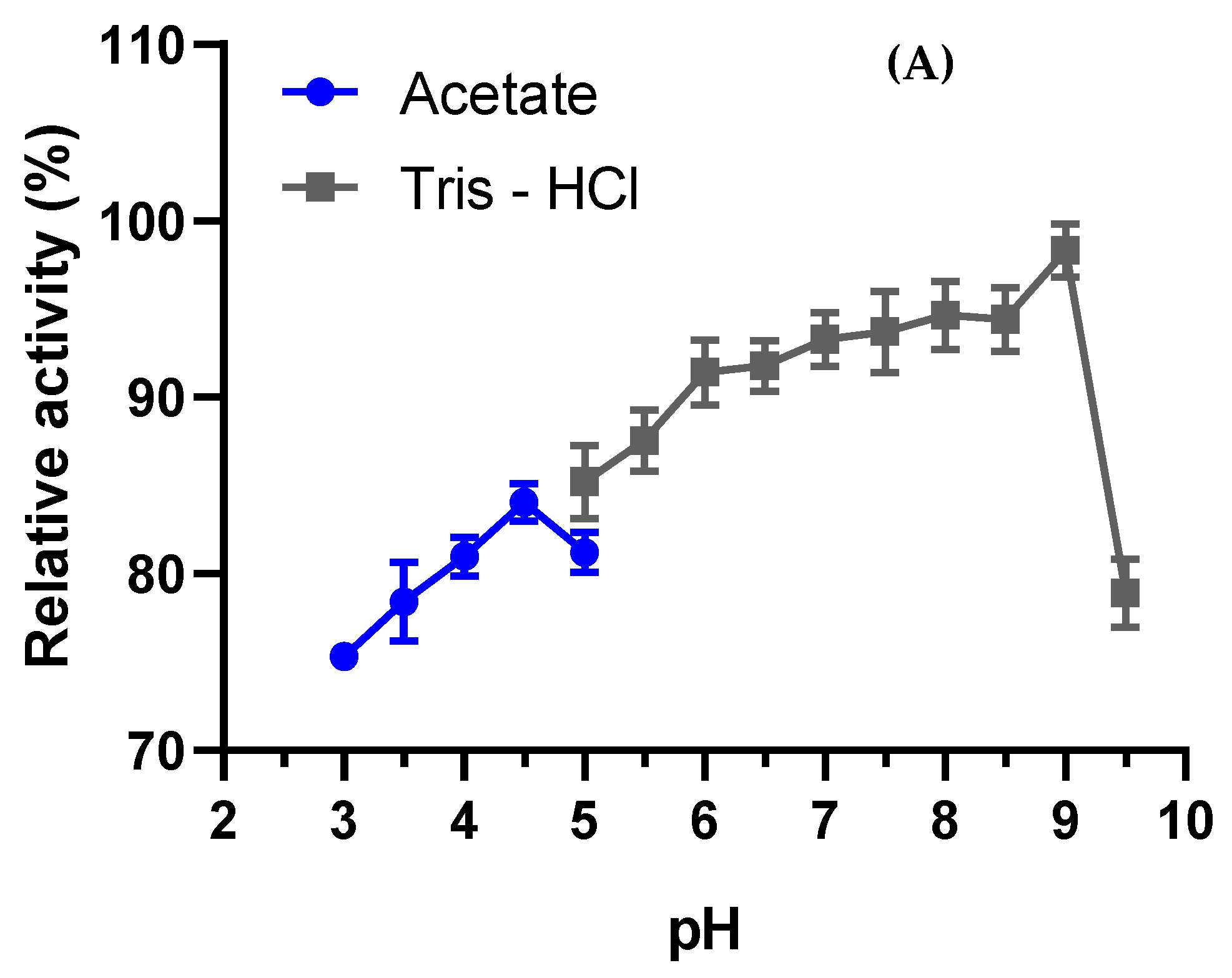
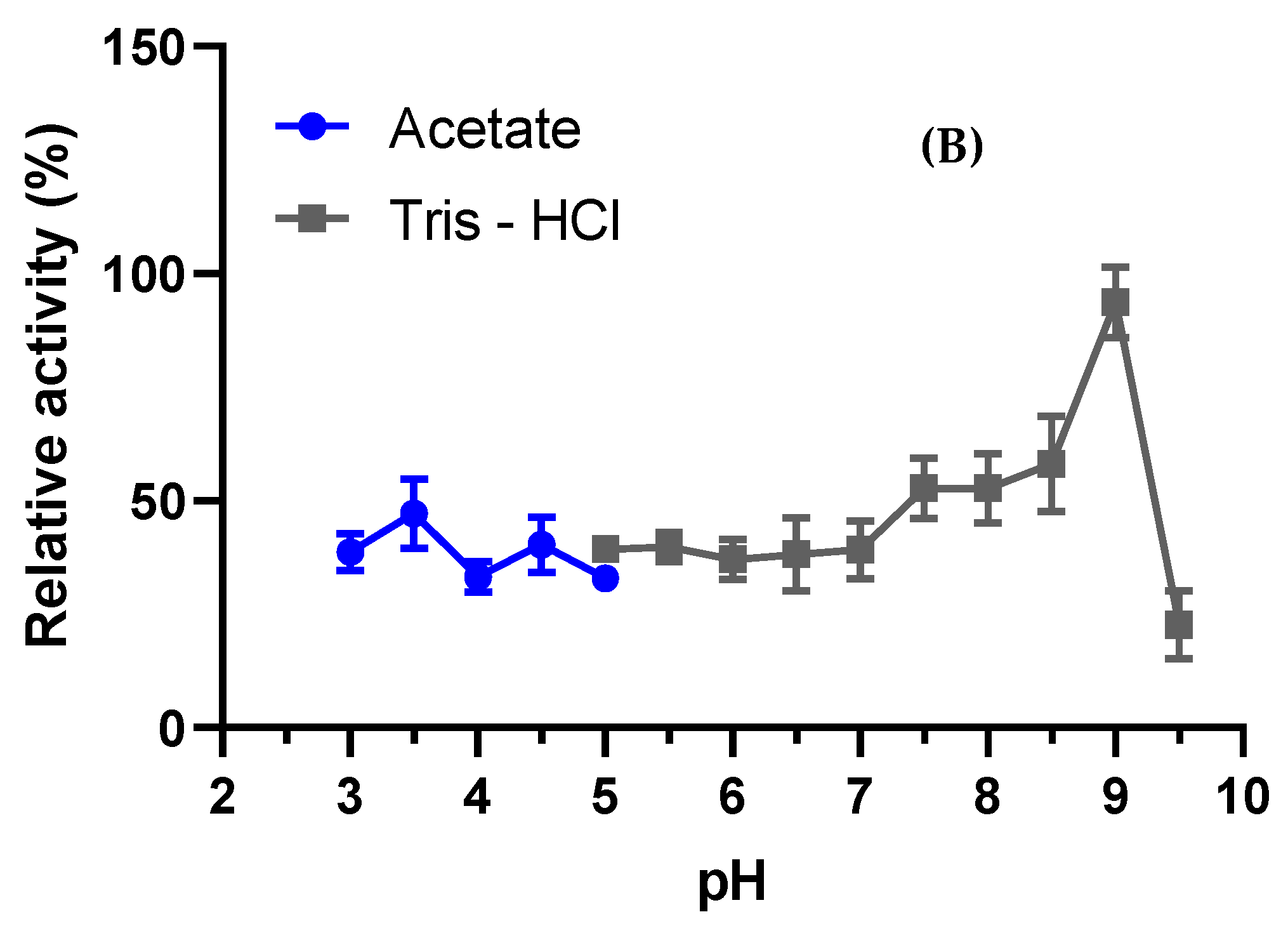
3.4.1. Optimal pH and Stability
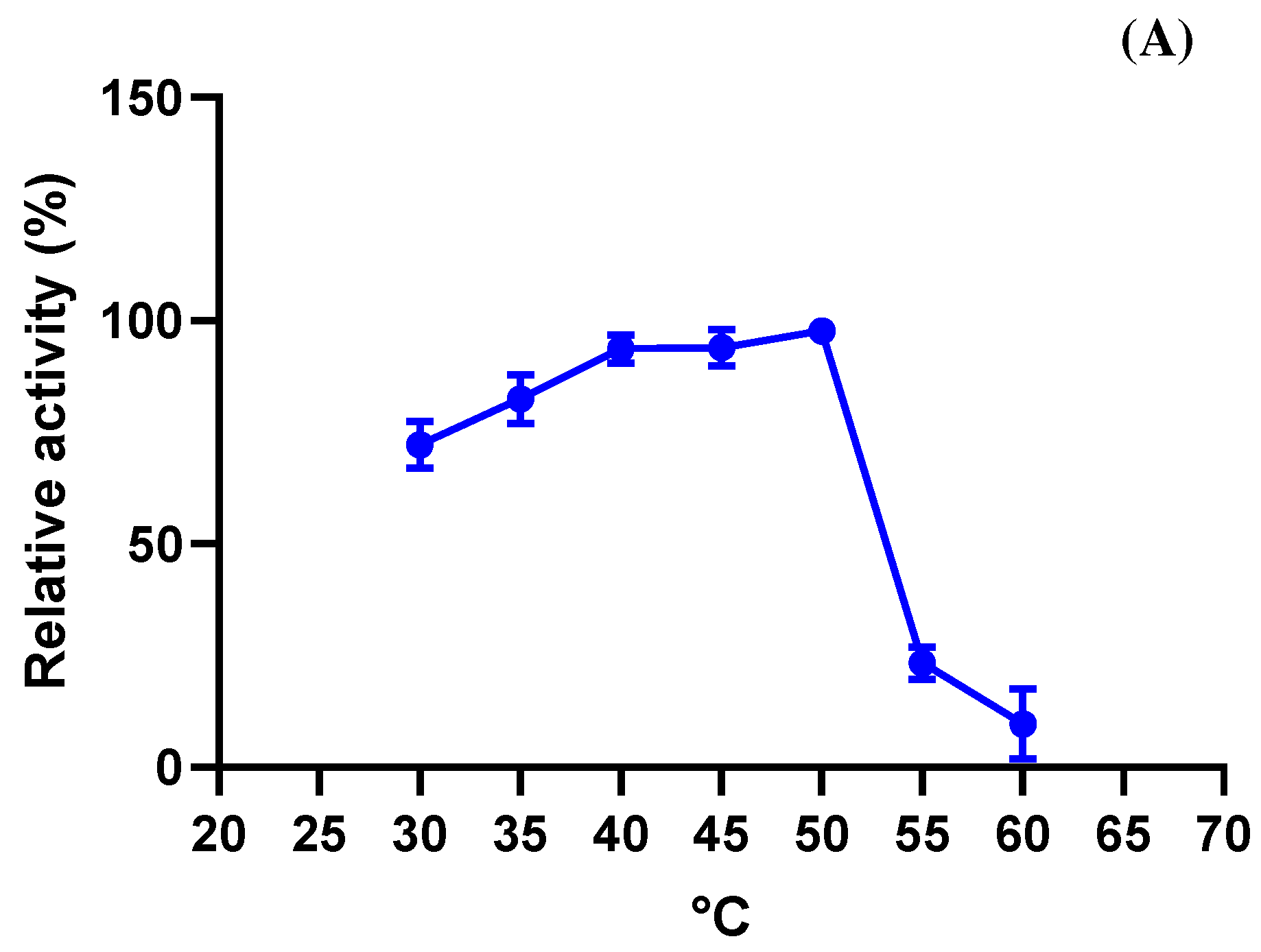
3.4.2. Optimal Temperature and Stability
3.4.3. Effect of Surfactants and Metal Ions in Salts
3.4.4. Stability in Organic Solvents
3.5. Cytotoxic Activity of L-Asparaginase in GM, RAW, and HeLa Cells
3.6. Morphological Viability of the HeLa Cell Line under L-Asparaginase Treatment
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Okunade, K.S. Human papillomavirus and cervical cancer. J. Obstet. Gynaecol. 2020, 40, 602–608. [Google Scholar] [CrossRef] [PubMed]
- El-Salem, F.; Mansour, M.; Gitman, M.; Miles, B.A.; Posner, M.R.; Bakst, L.; Genden, E.M.; Westra, W.H. Real-time PCR HPV genotyping in fine needle aspirations of metastatic head and neck squamous cell carcinoma: Exposing the limitations of conventional p16 immunostaining. Oral Oncol. 2019, 90, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, M.A.A.; Mascarenhas, G.M.S.; de Sousa Lima, A.A.; Carneiro, V.M.S.; Silva, C.D.C.M.; Silva, M.V.C.M. Correlation of genetic factors of hpv 16/18 virus and cervical cancer. Adv. Stud. Health Nat. 2022, 3. [Google Scholar] [CrossRef]
- Pedreschi, F.; Mariotti, S.; Granby, K.; Risum, J. Acrylamide reduction in potato chips using commercial asparaginase in combination with conventional bleaching. LWT Food Sci. Technol. 2011, 44, 1473–1476. [Google Scholar] [CrossRef]
- Shakambari, G.; Birendranarayan, A.K.; Lincy, M.J.A.; Rai, S.K.; Ahamed, Q.T.; Ashokkumar, B.; Varalakshmi, P. Hemocompatible glutaminase free L-asparaginase from marine Bacillus tequilensis PV9W with anticancer potential modulating p53 expression. RSC Adv. 2016, 6, 25943–25951. [Google Scholar] [CrossRef]
- Nunes, J.C.; Cristóvão, R.O.; Santos-Ebinuma, V.C.; Faria, J.L.; Silva, C.G.; Neves, M.C.; Freire, M.G.; Tavares, A.P. L-Asparaginase-based biosensors. Encyclopedia 2021, 1, 848–858. [Google Scholar] [CrossRef]
- Alam, S.; Pranaw, K.; Tiwari, R.; Khare, S.K. Recent development in the uses of asparaginase as food enzyme. In Green Bio-processes: Enzymes in Industrial Food Processing; Springer: Singapore, 2019; pp. 55–81. [Google Scholar]
- Hijiya, N.; Van Der Sluis, I.M. Asparaginase-associated toxicity in children with acute lymphoblastic leukemia. Leuk. Lymphoma 2016, 57, 748–757. [Google Scholar] [CrossRef] [PubMed]
- De Melo, D.W.; Fernandez-Lafuente, R.; Rodrigues, R.C. Enhancing biotechnological applications of l-asparaginase: Immobilization on amino-epoxy-agarose for improved catalytic efficiency and stability. Biocatal. Agric. Biotechnol. 2023, 52, 102821. [Google Scholar] [CrossRef]
- Feenstra, L.R.; Gehring, R.; van Geijswijk, I.M.; König, T.; Prinsen, H.C.M.T.; Vandemeulebroecke, K.; Lammens, T.; Krupa, A.; Teske, E. Evaluation of PEG-L-asparaginase in asparagine suppression and anti-drug antibody development in healthy Beagle dogs: A multi-phase preclinical study. Vet. J. 2022, 286, 105854. [Google Scholar] [CrossRef]
- Fonseca, M.H.G.; da Silva Fiúza, T.; de Morais, S.B.; Trevizani, R. Circumventing the side effects of L-asparaginase. Biomed. Pharmacother. 2021, 139, 111616. [Google Scholar] [CrossRef]
- Khokhar, I.; Haider, M.S.; Mukhtar, I.; Mushtaq, S. Biological control of Aspergillus niger, the cause of Black-rot disease of Allium cepa L. (onion), by Penicillium species. J. Agrobiol. 2012, 29, 23. [Google Scholar] [CrossRef]
- Kim, T.; Mullaney, E.J.; Porres, J.M.; Roneker, K.R.; Crowe, S.; Rice, S.; Lei, X.G. Shifting the pH profile of Aspergillus niger PhyA phytase to match the stomach pH enhances its effectiveness as an animal feed additive. Appl. Environ. Microbiol. 2006, 72, 4397–4403. [Google Scholar] [CrossRef]
- Mustafa, G.; Arif, R.; Bukhari, S.A.; Ali, M.; Sharif, S.; Atta, A. Structural and functional annotation of citrate synthase from Aspergillus niger ANJ-120. Pak. J. Pharm. Sci. 2018, 31, 709–717. [Google Scholar] [PubMed]
- Qin, S.; Zhou, M.; Wang, Z.; Li, P.; Huang, S.; Meng, J. Effect of pulsed electric field on spore germination rate and enzyme activity of Aspergillus niger. Innov. Food Sci. Emerg. Technol. 2023, 89, 103473. [Google Scholar] [CrossRef]
- Cairns, T.C.; Nai, C.; Meyer, V. How a fungus shapes biotechnology: 100 years of Aspergillus niger research. Fungal Biol. Biotechnol. 2018, 5, 13. [Google Scholar] [CrossRef]
- Bellaouchi, R.; Abouloifa, H.; Rokni, Y.; Hasnaoui, A.; Ghabbour, N.; Hakkou, A.; Asehraou, A. Characterization and optimization of extracellular enzymes production by Aspergillus niger strains isolated from date by-products. J. Genet. Eng. Biotechnol. 2021, 19, 50. [Google Scholar] [CrossRef]
- Dias, F.F.G.; Ruiz, A.L.T.G.; Della Torre, A.; Sato, H.H. Purification, characterization and antiproliferative activity of L-asparaginase from Aspergillus oryzae CCT 3940 with no glutaminase activity. Asian Pac. J. Trop. Biomed. 2016, 6, 785–794. [Google Scholar] [CrossRef]
- Mahajan, R.V.; Saran, S.; Saxena, R.K.; Srivastava, A.K. A rapid, efficient and sensitive plate assay for detection and screening of l-asparaginase-producing microorganisms. FEMS Microbiol. Lett. 2013, 341, 122–126. [Google Scholar] [CrossRef] [PubMed]
- Valenzuela-Lopez, N.; Sutton, D.A.; Cano-Lira, J.F.; Paredes, K.; Wiederhold, N.; Guarro, J.; Stchigel, A.M. Coelomycetous fungi in the clinical setting: Morphological convergence and cryptic diversity. J. Clin. Microbiol. 2017, 55, 552–567. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.J.W.T.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. A Guide Methods Appl. 1990, 18, 315–322. [Google Scholar]
- Alabdalall, A.H.; ALanazi, N.A.; Aldakeel, S.A.; AbdulAzeez, S.; Borgio, J.F. Molecular, physiological, and biochemical characterization of extracellular lipase production by Aspergillus niger using submerged fermentation. PeerJ 2020, 8, e9425. [Google Scholar] [CrossRef] [PubMed]
- Vala, A.K.; Sachaniya, B.; Dudhagara, D.; Panseriya, H.Z.; Gosai, H.; Rawal, R.; Dave, B.P. Characterization of L-asparaginase from marine-derived Aspergillus niger AKV-MKBU, its antiproliferative activity and bench scale production using industrial waste. Int. J. Biol. Macromol. 2018, 108, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Warburg, O.; Christian, W. Isolation and Crystallization of Enolase. Biochem. Z. 1942, 310, 384–421. [Google Scholar]
- Drainas, C.; Kinghorn, J.R.; Pateman, J.A. Aspartic hydroxamate resistance and asparaginase regulation in the fungus Aspergillus nidulans. Microbiology 1977, 98, 493–501. [Google Scholar] [CrossRef]
- Oliveira, B.H.; Coradi, G.V.; de Oliva-Neto, P.; do Nascimento, V.M.G. Biocatalytic benefits of immobilized Fusarium sp. (GFC) lipase from solid state fermentation on free lipase from submerged fermentation. Ind. Crops Prod. 2020, 147, 112235. [Google Scholar] [CrossRef]
- El-Gendy, M.M.A.A.; Yahya, S.M.; Hamed, A.R.; Soltan, M.M.; El-Bondkly, A.M.A. Phylogenetic analysis and biological evaluation of marine endophytic fungi derived from Red Sea sponge Hyrtios erectus. Appl. Biochem. Biotechnol. 2018, 185, 755–777. [Google Scholar] [CrossRef] [PubMed]
- Elshafei, A.M.; El-Ghonemy, D.H. Screening and media optimization for enhancing L-asparaginase production, an anticancer agent, from different filamentous fungi in solid state fermentation. Br. Biotechnol. J. 2015, 9, 1–15. [Google Scholar] [CrossRef]
- Cardoso, S.L.; de Freitas, M.M.; de Souza, P.M.; Homem-de-Mello, M.; Silveira, D.; Fonseca-Bazzo, Y.M.; Magalhães, P.O. Optimization of aqueous two-phase micellar system for partial purification of L-asparaginase from Penicillium sp. grown in wheat bran as agro-industrial residue. Braz. J. Microbiol. 2020, 51, 979–988. [Google Scholar] [CrossRef] [PubMed]
- Castro, A.M.D.; Pereira, N., Jr. Production, properties and application of cellulases in the hydrolysis of agro-industrial waste. Química Nova 2010, 33, 181–188. [Google Scholar] [CrossRef]
- Babu, U.K.; Ramagopal, N.; Reddy, D.R. Optimization of L-Asparaginase production from Isolated Aspergillus niger by using Solid State Fermentation on sesame cake via application of Genetic Algorithm, and Artificial Neural Network-based design model. J. Biotechnol. 2010, 150, 538–539. [Google Scholar] [CrossRef]
- Karanam, S.K.; Medicherla, N.R. Application of Doehlert experimental design for the optimization of medium constituents for the production of L-asparaginase from Palm Kernal cake (Elaeis guineensis). J. Microbial. Biochem. Technol. 2010, 2, 1–7. [Google Scholar] [CrossRef]
- Tormo, M.A.; Gil-Exojo, I.; Romero de Tejada, A.; Campillo, J.E. White bean amylase inhibitor administered orally reduces glycaemia in type 2 diabetic rats. Br. J. Nutr. 2006, 96, 539–544. [Google Scholar] [CrossRef] [PubMed]
- Pereira, L.L.S.; Santos, C.D.D.; Pereira, C.A.; Marques, T.R.; Sátiro, L.C. Precipitation of α-amylase inhibitor from white beans: Evaluation of methods. Food Nutr. Araraquara 2010, 21, 15–20. [Google Scholar]
- El-Naggar, N.E.A.; Deraz, S.F.; Soliman, H.M.; El-Deeb, N.M.; El-Ewasy, S.M. Purification, characterization, cytotoxicity and anticancer activities of L-asparaginase, anti-colon cancer protein, from the newly isolated alkaliphilic Streptomyces fradiae NEAE-82. Sci. Rep. 2016, 6, 32926. [Google Scholar] [CrossRef] [PubMed]
- Dunn, J.B.; Mueller, S.; Wang, M.; Han, J. Energy consumption and greenhouse gas emissions from enzyme and yeast manufacture for corn and cellulosic ethanol production. Biotechnol. Lett. 2012, 34, 2259–2263. [Google Scholar] [CrossRef] [PubMed]
- Pourkhanali, K.; Khayati, G.; Mizani, F.; Raouf, F. Characterization of free and immobilized lipase from Penicillium sp. onto three modified bentonites: A comparative study. J. Biotechnol. 2022, 344, 57–69. [Google Scholar] [CrossRef] [PubMed]
- El-Bessoumy, A.A.; Sarhan, M.; Mansour, J. Production, isolation, and purification of L-asparaginase from Pseudomonas aeruginosa 50071 using solid-state fermentation. BMB Rep. 2004, 37, 387–393. [Google Scholar] [CrossRef]
- Mukherjee, R.; Bera, D. Biochemical characterization and thermodynamic principles of purified L-asparaginase from novel Brevibacillus borstelensis ML12. Biocatal. Agric. Biotechnol. 2022, 39, 102260. [Google Scholar] [CrossRef]
- Singh, Y.; Gundampati, R.K.; Jagannadham, M.V.; Srivastava, S.K. Extracellular L-asparaginase from a protease-deficient Bacillus aryabhattai ITBHU02: Purification, biochemical characterization, and evaluation of antineoplastic activity in vitro. Appl. Biochem. Biotechnol. 2013, 171, 1759–1774. [Google Scholar] [CrossRef]
- Moreno-Enríquez, A.; Evangelista-Martínez, Z.; González-Mondragón, E.G.; Calderón-Flores, A.; Arreguín, R.; Pérez-Rueda, E.; Huerta-Saquero, A. Biochemical characterization of recombinant L-asparaginase (AnsA) from Rhizobium etli, a member of an increasing rhizobial-type family of L-asparaginases. J. Microbiol. Biotechnol. 2012, 22, 292–300. [Google Scholar] [CrossRef]
- Krishnapura, P.R.; Belur, P.D. Partial purification and characterization of L-asparaginase from an endophytic Talaromyces pinophilus isolated from the rhizomes of Curcuma amada. J. Mol. Catal. B Enzym. 2016, 124, 83–91. [Google Scholar] [CrossRef]
- Meghavarnam, A.K.; Janakiraman, S. Evaluation of acrylamide reduction potential of l-asparaginase from Fusarium culmorum (ASP-87) in starchy products. LWT 2018, 89, 32–37. [Google Scholar] [CrossRef]
- Asthana, N.; Azmi, W. Microbial L-asparaginase: A Potent Antitumour Enzyme. 2003. Available online: https://api.semanticscholar.org/CorpusID:33108832 (accessed on 14 November 2023).
- Rani, S.A.; Sundaram, L.; Vasantha, P.B. A study on in vitro antioxidant and anticancer activity of l-asparaginase. J. Pharm. Res. 2012, 5, 1463–1466. [Google Scholar]
- Sudarkodi, C.; Sundar, S. Anticancer activity of L-asparaginase from Aspergillus oryzae against HEP-G2 and Hela cell lines. Int. J. Recent Sci. Res. 2018, 9, 25328–25330. [Google Scholar]
- Fatima, N.; Khan, M.M.; Khan, I.A. L-asparaginase produced from soil isolates of Pseudomonas aeruginosa shows potent anti-cancer activity on HeLa cells. Saudi J. Biol. Sci. 2019, 26, 1146–1153. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Fermentation | Purification Stage | Total Activity (U/mL) | Total Protein (mg) | Specific Activity (U/mg) | Yield (%) | Purification |
|---|---|---|---|---|---|---|
| SmF | Crude Extract | 3711.50 | 480.00 | 7.73 | 100 | 1 |
| Fraction 1 (0–40%) | 363.96 | 18.95 | 19.21 | 9.81 | 2.48 | |
| Fraction 2 (40–80%) | 767.20 | 44.93 | 17.07 | 20.67 | 2.21 | |
| SsF | Crude Extract | 5261 | 480 | 10.96 | 100.00 | 1.00 |
| Fraction 1 (0–40%) | 888.58 | 84.63 | 10.50 | 16.89 | 0.96 | |
| Fraction 2 (40–80%) | 2748.9 | 14.685 | 187.19 | 52.25 | 17.08 |
| Type of Precipitation | Purification Stage | Total Activity (U/mL) | Total Protein (mg) | Specific Activity (U/mg) | Yield | Purification (%) |
|---|---|---|---|---|---|---|
| Crude Extract | 5520.50 | 3367.5 | 1.64 | 100.00 | 1.00 | |
| Ethanol | Fraction 1 (0–40%) | 1205.68 | 84.63 | 14.25 | 21.84 | 8.69 |
| Fraction 2 (40–80%) | 2671.90 | 114.18 | 23.40 | 48.40 | 14.27 | |
| DEAE Cellulose Column | 1092.30 | 31.8 | 34.35 | 19.79 | 20.95 | |
| Ammonium Sulfate | Fraction 1 (0–40%) | 182.16 | 57.09 | 3.19 | 3.30 | 1.95 |
| Fraction 2 (40–80%) | 826.20 | 152.01 | 5.44 | 14.97 | 3.32 | |
| DEAE Cellulose Column | 265.00 | 21.5 | 12.33 | 4.80 | 7.52 | |
| Isopropanol | Fraction 1 (0–40%) | 113.40 | 64.35 | 1.76 | 2.05 | 1.07 |
| Fraction 2 (40–80%) | 306.60 | 143.955 | 2.13 | 5.55 | 1.30 | |
| DEAE Cellulose Column | 104.10 | 16.3 | 6.39 | 1.89 | 3.90 |
| Type | Concentrations | U/mL | Relative Activity (%) ± SD |
|---|---|---|---|
| Control | 47.48 | 100 ± 0.014 | |
| Tween 20 | 0.01% | 44.98 | 94.73 ± 0.035 |
| 0.10% | 55.27 | 116.41 ± 0.084 | |
| 0.50% | 51.35 | 108.15 ± 0.028 | |
| Triton X-100 | 0.01% | 45.91 | 96.69 ± 0.013 |
| 0.10% | 36.56 | 77.00 ± 0.018 | |
| 0.50% | 33.92 | 71.44 ± 0.020 | |
| Tween 80 | 0.01% | 41.61 | 87.64 ± 0.011 |
| 0.10% | 42.45 | 89.41 ± 0.006 | |
| 0.50% | 38.01 | 80.05 ± 0.027 | |
| SDS | 0.01% | 35.02 | 73.76 ± 0.012 |
| 0.10% | 24.13 | 50.82 ± 0.009 | |
| 0.50% | 24.43 | 51.45 ± 0.025 | |
| E.D.T. A | 0.01% | 31.1 | 65.50 ± 0.005 |
| 0.10% | 22.12 | 46.59 ± 0.004 | |
| 0.50% | 19.13 | 40.29 ± 0.003 | |
| Type | U/mL | Relative Activity (%) ± SD |
|---|---|---|
| Control | 31.31 | 100 ± 0.04 |
| Mg2+ | 46.4 | 148.20 ± 0.02 |
| Co2+ | 16.89 | 153.94 ± 0.01 |
| SO42− | 15.05 | 48.07 ± 0.03 |
| Zn+2 | 16.21 | 51.77 ± 0.01 |
| Ca2+ | 46.13 | 47.33 ± 0.01 |
| Fe3+ | 27.63 | 88.25 ± 0.01 |
| Na+ | 28.47 | 90.93 ± 0.02 |
| Type | Concentrations | U/mL | Relative Activity (%) ± SD |
|---|---|---|---|
| Control | 32.48 | 100 ± 0.014 | |
| Acetone | 25 | 16.79 | 51.72 ± 0.05 |
| 50 | 14.53 | 44.72 ± 0.02 | |
| 80 | 5.76 | 17.73 ± 0.03 | |
| 100 | 2.49 | 7.67 ± 0.04 | |
| Ethanol | 25 | 24.15 | 74.38 ± 0.04 |
| 50 | 15.02 | 46.18 ± 0.01 | |
| 80 | 9.54 | 29.34 ± 0.02 | |
| 100 | 6.28 | 19.33 ± 0.02 | |
| Isopropanol | 25 | 13.45 | 41.37 ± 0.03 |
| 50 | 10.26 | 31.59 ± 0.01 | |
| 80 | 6.75 | 20.78 ± 0.03 | |
| 100 | 4.14 | 12.75 ± 0.01 | |
| Methanol | 25 | 8.55 | 26.32 ± 0.09 |
| 50 | 7.82 | 24.05 ± 0.03 | |
| 80 | 5.19 | 16.00 ± 0.02 | |
| 100 | 2.34 | 7.20 ± 0.07 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
da Silva Duarte, S.M.; de Carvalho Silva, A.K.; Assunção Borges, K.R.; Borges Cordeiro, C.; Lindoso Lima, F.J.; da Silva, M.A.C.N.; de Souza Andrade, M.; Nascimento, M.d.D.S.B. Production, Characterization Purification, and Antitumor Activity of L-Asparaginase from Aspergillus niger. Fermentation 2024, 10, 226. https://doi.org/10.3390/fermentation10050226
da Silva Duarte SM, de Carvalho Silva AK, Assunção Borges KR, Borges Cordeiro C, Lindoso Lima FJ, da Silva MACN, de Souza Andrade M, Nascimento MdDSB. Production, Characterization Purification, and Antitumor Activity of L-Asparaginase from Aspergillus niger. Fermentation. 2024; 10(5):226. https://doi.org/10.3390/fermentation10050226
Chicago/Turabian Styleda Silva Duarte, Suzane Meriely, Allysson Kayron de Carvalho Silva, Katia Regina Assunção Borges, Carolina Borges Cordeiro, Fernanda Jeniffer Lindoso Lima, Marcos Antônio Custódio Neto da Silva, Marcelo de Souza Andrade, and Maria do Desterro Soares Brandão Nascimento. 2024. "Production, Characterization Purification, and Antitumor Activity of L-Asparaginase from Aspergillus niger" Fermentation 10, no. 5: 226. https://doi.org/10.3390/fermentation10050226
APA Styleda Silva Duarte, S. M., de Carvalho Silva, A. K., Assunção Borges, K. R., Borges Cordeiro, C., Lindoso Lima, F. J., da Silva, M. A. C. N., de Souza Andrade, M., & Nascimento, M. d. D. S. B. (2024). Production, Characterization Purification, and Antitumor Activity of L-Asparaginase from Aspergillus niger. Fermentation, 10(5), 226. https://doi.org/10.3390/fermentation10050226