3.1. Antifungal Activity of Peptides against C. albicans and C. auris
In this investigation, the antifungal activity of twenty-one peptides derived from the peptide RWQWRWQWR, named R-1-R, was evaluated against the reference strain
C. albicans SC5314 (sensitive to antifungals) and clinical isolates
C. albicans 256-PUJ-HUSI (FLC-resistant),
C. auris 435-PUJ-HUSI (FLC-sensitive), and
C. auris 537-PUJ-HUSI (FLC- and AmB-resistant) (
Table 1). The peptides used in this investigation were synthesized manually via solid-phase peptide synthesis using the Fmoc/tBu strategy, and these molecules were classified as follows:
Group I: sequences in which each amino acid is substituted sequentially by alanine (alanine scan) [
48]; these peptides were evaluated in order to determine which amino acids are relevant for antifungal activity.
Group II: Lysine (Lys) was substituted for all arginine (Arg) residues; replacing Arg residues with Lys has been shown to facilitate and reduce the cost of the synthesis [
44].
Group III: Some Arg residues were removed or added at the N-terminal and/or C-terminal ends to determine the influence of the positive charge in the antifungal activity [
46,
65,
66].
Group IV: Previous reports have shown that the substitution of Arg by Ala at position 5 increased the cytotoxic effect in breast cancer cell lines. To establish whether this position is relevant in antifungal activity, position 5 was replaced by non-natural hydrophobic amino acids [
42,
65]; additionally, the presence of non-natural amino acids in the sequence increased the proteolytic stability [
67,
68,
69].
To facilitate reading, all peptides used in this research were coded, taking into account the modification performed on central sequence WQWRWQW, which is coded as 1; hence, palindromic peptide RWQWRWQWR corresponds to R-1-R. For the peptides generated as alanine scan or modifications in sequence 1, the residue position changed is indicated in superscript, and the residue introduced is indicated in square brackets; for example, peptide RWAWRWQWR corresponds to [
3A]-R-1-R and KWQWKWQWK to K-1-K (
Table 1).
The results showed that the palindromic peptide R-1-R had significant antifungal activity against
C. albicans SC5314 and 256 (MIC and MFC values were 67 µM in all cases). Peptides where substitution of glutamine (Gln) by Ala was performed, [
3A]-R-1-R and [
7A]-R-1-R (group I), exhibited similar antifungal activity against the reference strain and greater activity against the clinical isolate than the R-1-R peptide, suggesting that the increased hydrophobicity at position 3 or 7 is relevant for antifungal activity. The Gln side chain does not contribute charge to the sequence and does not influence the amphipathic properties, so it can be considered a spacer. Additionally, peptide [
7A]-R-1-R was previously studied, and it exhibited increased antibacterial activity against
E. coli 25922 compared with R-1-R [
48]. These results suggest that the absence of Gln at the 7th position does not decrease the antimicrobial activity.
Peptides in which the tryptophan (Trp) or Arg residues were replaced by Ala or peptides in which the net positive charge was reduced exhibited lower antifungal activity against both strains of C. albicans than the R-1-R peptide, suggesting that these changes affect the amphipaticity of the sequence, which is relevant to antifungal activity. Furthermore, substitution of any Arg or Trp residue in the sequence decreased the antifungal activity, suggesting that Arg and Trp residues are relevant for antifungal activity.
Substitution of Arg or Trp by Ala in the palindromic sequence affects the net charge and amphipathic properties. Arg and Trp residues have been related to the interaction of the peptide with negatively charged molecules on the cell surface and the lipid bilayer, respectively. These results are in agreement with previous studies performed with Ala scan analogues of the R-1-R peptide against Gram-positive and Gram-negative bacteria and cancer cells [
48], where the substitution of Arg or Trp with Ala significantly reduced the biological activity, indicating that the amphipathicity generated by the cationic charge of Arg and the hydrophobicity of Trp are crucial for the antifungal activity against
C. albicans strains, thus also being consistent with the initial mechanism of antimicrobial action proposed for peptides derived from LfcinB, which is related to electrostatic and hydrophobic interactions with cell membranes [
13,
19].
Peptides K-1-K, RR-1-R, and R-1-RR exhibited similar antifungal activity against both
C. albicans strains, indicating that the substitution of Arg by Lys and the incorporation of an Arg residue at the N-terminal do not reduce the antifungal activity. These results suggested that the positive charge could be relevant to antifungal activity. These results are consistent with studies that showed that Arg by Lys substitutions in the AMP sequence induced equal or less antibacterial activity [
70,
71,
72]. The action mechanism of AMPs is based on the initial interaction between the negatively charged molecules on the cell surface and the positively charged side chains of the peptide. The substitution of Arg by Lys does not change the net charge of the sequence; however, the nature of each side chain is different: the primary amine of the guanidine group interacts with the phospholipids of cell membranes and has a more distributed positive charge, so it can form a greater number of hydrogen bonds [
70,
71,
72].
Peptide RR-1-R resulted from the addition of an Arg residue at the N-terminal of the sequence of the palindromic peptide and exhibited similar antifungal activity against
C. albicans strains. When the Arg residue was incorporated into the N-terminal end, the minimal motif of the LfcinB (RRWQWR) was completed in the peptide sequence. This result is in agreement with previous reports that showed that peptides containing the minimal motif exhibited antibacterial, antifungal, antiviral, and anticancer activity [
50]. Peptides RR-1-RR and R-1-RR exhibited antifungal activity only against the SC5314 strain, while the antifungal activity against the isolated clinical
C. albicans 256 was reduced. Our results suggested that the increases in the net positive charge in the sequence did not increase the antifungal activity.
In peptides from group IV, the central Arg residue was replaced by non-natural hydrophobic amino acid. The antifungal activity against
C. albicans strains of all peptides was lost, suggesting that the Arg residue in this position is important for antifungal activity. These last results are in agreement with studies that showed that an increase in the positive charge or hydrophobicity did not improve the antibacterial activity of R-1-R; in this case, the antifungal activity was not improved either. The same thing happened in a bioinformatics modeling study with short cationic AMPs, where going from a +4 to a +7 charge also negatively affected the activity [
46,
66,
73]. Our results agree with previous studies [
21], and peptide R-1-R had antifungal activity against strains and clinical isolates of
C. albicans,
C. glabrata,
C. krusei,
C. auris, and
C. tropicalis. Additionally, it was fungistatic and fungicidal and combined with FLC exhibited a synergistic antifungal effect against
C. tropicalis 883 and
C. krusei 6258 (resistant to FLC).
The antifungal activity of peptide R-1-R against
C. albicans was comparable to that reported for LFB (MIC 200->6,400 μg/mL) and LfcinB (MIC 0.8–400 μg/mL). Additionally, the antifungal activity of the peptide R-1-R is in agreement with that previously reported for short peptides derived from LfcinB against other
C. albicans strains: Peptide 2 (FKCRRWQWRM; MIC 17.3–17.5 μM), bLF (or LFB) 17–30 (FKCRRWQWRMKKLGA; MIC 5–10 μM), LfcinB-20 (LfcinB 18–37) (FKCRRWQWRMKKLGA; MIC 8 μg/mL), Lfcin B-18-42 (MIC 100 μg/mL); Lfcin B-9 (RRWQWRMKK; MIC 25,000–32,000 μg/mL), Peptide 5 (RWQWRM; MIC 500 μM), and Peptide 3 (GAPSITCVRRAF; MIC 635 μM). The peptide R-1-R also exhibited similar antifungal activity to other palindromic sequences rich in Arg and/or Trp residues in some
C. albicans strains, as follows: IRIRIRIR, KWKWWKWK, KWKWKWKW [
21].
The foregoing supports the idea that not only are a higher cationic charge and hydrophobic residues important for antimicrobial activity but so is a balance between these two parameters, which, although they are important in the mechanism of action in AMPs, are not the only ones. For example, it is reported that the cationic nature and unique geometry of the hydrogen bonds of the Arg residue together with the complex properties of Trp can complement each other to exert an antimicrobial effect. The cationic charge of Arg provides an efficient means of attracting peptides to target membranes, and hydrogen bonds facilitate their interaction with negatively charged surfaces (effectively achieved with other cationic residues), allowing Trp (the most suitable amino acid according to studies) a prolonged association of the peptide with the cell membrane, although this interaction model depends on the AMP [
74].
The palindromic peptide R-1-R exhibited low antifungal activity against the two clinical isolates of
C. auris evaluated, while peptide [
5A]-R-1-R was the one peptide that exhibited the highest antifungal activity against the clinical isolate
C. auris 537. Our results suggest that the palindromic peptide exhibited less antifungal activity against
C. auris strains than those of
C. albicans. In a previous report, we showed that peptides [
5A]-R-1-R and R-1-R exhibited similar antibacterial activity against
E. coli and
S. aureus strains. Additionally, the peptide [
5A]-R-1-R exhibited a higher cytotoxic effect against colon cancer cells Caco-2 than peptide R-1-R [
48,
49].
3.4. Antifungal Activity of Mixtures of Peptide/Extract, Extract/FLC, and Peptide/FLC
The combination of new antifungal treatment alternatives (AMPs, plant extracts, or synthetic molecules) with commercial antifungal agents has been used to overcome the challenge of antifungal resistance and toxicity [
43,
60,
85,
86]. In the present study, after confirming that both the palindromic peptide R-1-R and the ethanolic extract of
B. pilosa leaves exhibited weak to moderate activity (according to those described by cut-off points suggested by [
60]) against two species of
Candida, both sensitive and resistant to antifungals, they were evaluated in combination with FLC and with each other, to see if this antifungal activity could be improved.
Initially, the antifungal activity of mixtures of R-1-R/FLC against the
C. albicans and
C. auris strains was evaluated (
Table 5). This mixture exhibited an additive effect against
C. albicans SC5314 (sensitive to FLC) and
C. albicans 256 (resistant to FLC) with FIC Index (FIC I) of 0.75 and 0.62, respectively. The MIC value of FLC in
C. albicans SC5314 was reduced by a factor of 2, while that in
C. albicans 256 was reduced by a factor of 8. Interestingly, it was found that
C. albicans 256 returned to a sensitive phenotype when FLC (4 µg/mL) was combined with R-1-R. In the case of
C. auris 435 (sensitive to FLC), a synergistic effect was obtained (FIC I = 0.28), enhancing the activity of both the peptide and the FLC, by a factor of 32 and 4, respectively. For
C. auris 537 (resistant to FLC and AmB), an indifferent effect was obtained (FIC I = 1.1) [
21].
Subsequently, the extract of
B. pilosa was combined with FLC (
Table 6), this mixture was evaluated against
C. albicans SC5314, and additivity was obtained (FIC I = 0.53), decreasing the MIC of the extract and FLC by factors of 32 and 2, respectively. Against
C. albicans 256 (FIC I = 1.25) and
C. auris 435 (FIC I = 1.03) and 537 (FIC I = 1.25), an indifferent effect was determined.
When the peptide and the extract were combined, it was possible to obtain the greatest antifungal activity. Since a synergistic effect was found for
C. albicans SC5314 (FIC I = 0.5),
C. auris 435 (FIC I = 0.37), and
C. auris 537 (FIC I = 0.12), with this combination it can be seen that the activity of both the peptide and the extract was enhanced in all strains. For
C. albicans 256, there was an additivity effect (FIC I = 0.62); however, the MIC of the R-1-R and extract was reduced by factors of between 2 and 8 (
Table 7).
Finally, it was decided to evaluate the combination of extract, peptide, and FLC (
Table 8), obtaining additivity against
C. albicans SC5314 (FIC I = 0.68),
C. albicans 256 (FIC I = 0.65), and
C. auris 435 (FIC I = 0.81) and a synergistic effect against
C. auris 537 (FIC I = 0.12). It is important to point out that
C. albicans 256 and
C. auris 537, with this combination, returned to a phenotype sensitive to FLC (MIC 4 and 8, respectively). On the other hand, it was also found that the combination of extract, peptide, and FLC against
C. albicans 5314 (E: 31.25 µg/mL, P: 50 µg/mL, FLC: 0.06) and 256 (E: 31.25 µg/mL, P: 50 µg/mL, FLC: 4) resulted in a fungicidal effect, unlike all the other combinations evaluated, where the effect was fungistatic.
MICE, MICR-1-R, and MICFLC correspond to the MIC (µg/mL) of the extract of B. pilosa, R-1-R, and FLC, respectively, and E, P, and FLC are the MIC values when combining the extract, peptide, and FLC. Minimum fractional concentration index (FIC I), MICE/E, MICR-1-R/R-1-R, and MICFLC/FLC MIC represent the factor by which the extract, peptide, or FLC are potentiated after being evaluated in combination, respectively.
After the evaluation of the different combinations of extract, peptide, and/or FLC, several important findings were determined. First, the palindromic peptide is key to enhancing the activity of FLC, especially in resistant Candida strains. Previously, R-1-R was evaluated in combination with FLC or caspofungin (CAS) in different Candida species resistant to these antifungals, such as
C. albicans 256, C. tropicalis 883, C. krusei 6258,
C. auris 537, and C. glabrata 1875, obtaining a synergistic or additive effect [
21]. In the present study, the FLC-sensitive strains
C. albicans SC5314 and
C. auris 435 were included. According to the FIC I obtained, it can be concluded that the resistant strains were more susceptible to this combination (
Table 5), and the same occurred when the peptide was mixed with the extract and FLC; even the two resistant strains returned to an FLC-sensitive phenotype (
Table 8). These results are in agreement with previous studies carried out with LF, LfcinB, and a peptide derived from human LF (hLF) (1–11) [
20,
87].
Although the mechanism of this synergistic effect is not clear, it has been suggested after studies with the peptide hLF(1–11) [
87] that this peptide in subcandidacidal concentrations enters the yeast through interaction with surface molecules of the fungus and goes to the mitochondria, producing extracellular adenosine triphosphate (ATP) release (it has been seen that this released ATP could generate pores in the cell or the mitochondrial membrane [
88]), but this is not enough to generate cell death, for which later, when FLC is added (in subcandidacidal concentrations), it acts as an effector and finally induces cell death. This could be extrapolated to LfcinB peptides, since it has also been shown that LfcinB15 generates extracellular ATP release [
19], although more studies are needed to confirm whether, for this AMP, ATP can generate pores, release ions, and induce cell lysis.
On the other hand, it has also been suggested that strains resistant to FLC are more susceptible to LF or LfcinB, possibly due to the absence of cytochrome P-450 (important in the biosynthesis of ergosterol in the yeast membrane), which could produce disturbances in the membrane due to alterations in the sterols [
20,
89]. This makes resistant strains more susceptible to the action of LF, LfcinB, or derived peptides, which, as mentioned before, initially generates membrane damage. Additionally, it has been proposed that LfcinB could inhibit efflux pumps [
20,
90] (another mechanism of resistance to azoles), which could explain, in this study, the return of strains resistant to FLC to sensitivity, but complementary studies are needed to confirm it.
Second, when the extract combined with FLC was evaluated (
Table 6), an indifferent effect was observed in three strains and only synergy was observed for
C. albicans SC5314, suggesting that this extract is not of great importance for enhancing FLC activity in all strains. However, when the extract was combined with peptide and FLC (
Table 8), it was observed that
C. auris 537 returned to FLC sensitivity, an effect that was not found only by combining R-1-R and FLC.
When the extract was combined with the peptide (
Table 7), it was possible to find the lowest FIC I (0.12–0.62) of all the combinations evaluated against the four study strains. It should even be noted that
C. auris 537, resistant to FLC and AmB, was the most susceptible strain to this mixture, where both the peptide and extract activity were enhanced by a factor of 16. This indicates that in some way the extract is contributing to the mechanism of action of the peptide, but since there is little information on the mechanism of action of
B. pilosa extracts, little can be inferred about how the extract acts, with FLC or with the peptide. According to what was described by Angelini [
28], it is possible that the extract is helping the peptide in the membrane disruption process, but more studies are needed in this regard. This was corroborated by combining the extract with peptides derived from R-1-R: two ([
3A]-R-1-R, RR-1-R) presented MICs similar to the palindromic peptide and two peptides ([
2A]-R-1-R, [
5Bpa]-R-1-R) that did not exhibit activity. The results showed that with the combination peptide/
B. pilosa, a synergistic effect is achieved in all cases, except for [
3A]-R-1-R, which was additive, enhancing the activity of the peptides and extract by a factor of between 2 and 16 (
Table S1).
Third, according to the results obtained, it is considered that the combinations of peptide/FLC, peptide/extract, and peptide and extract/FLC, depending on the yeast, could be a promising strategy for enhancing the activity against Candida and could be considered for developing new treatments to mitigate the problem of resistance to antifungals.
3.5. Microscopy
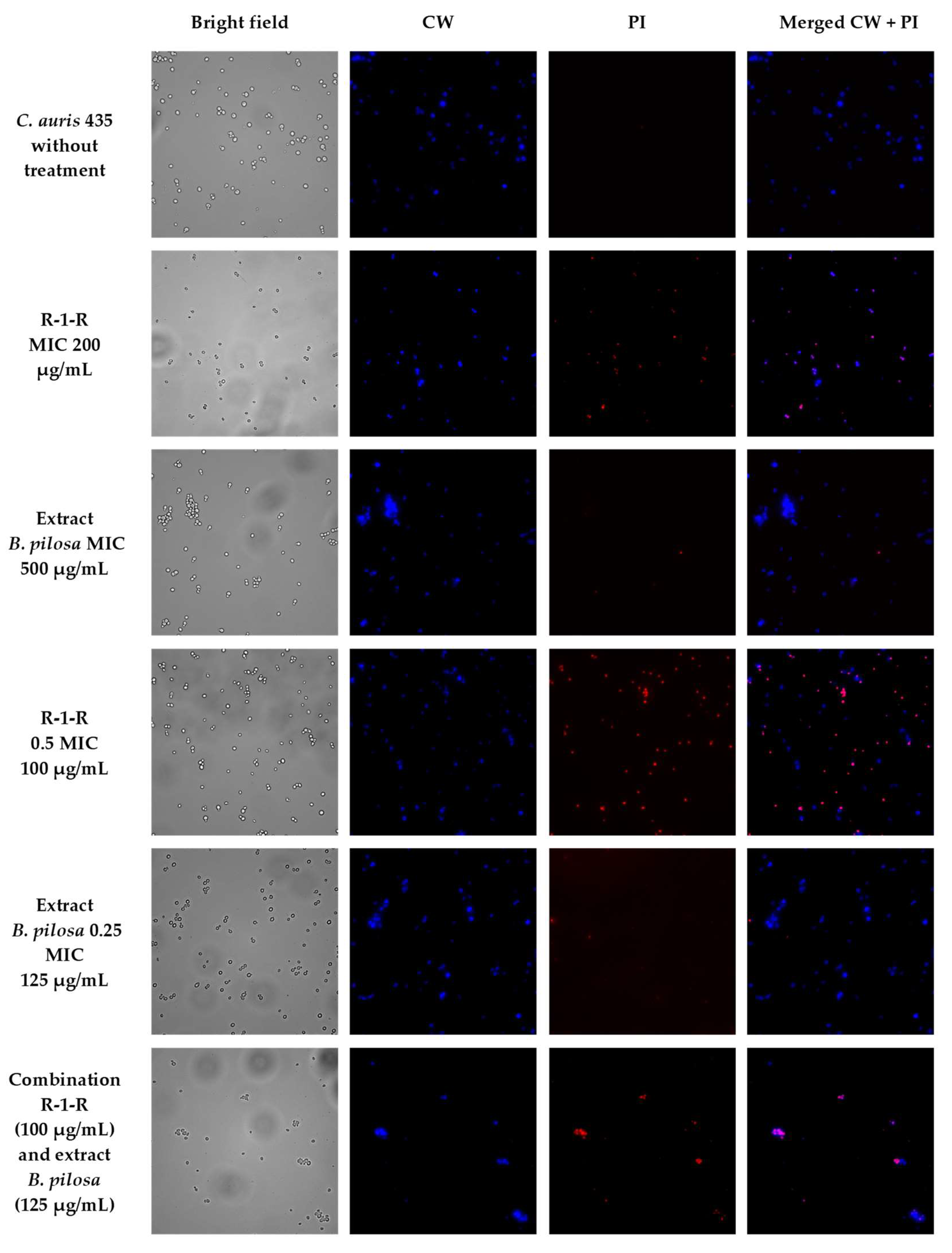
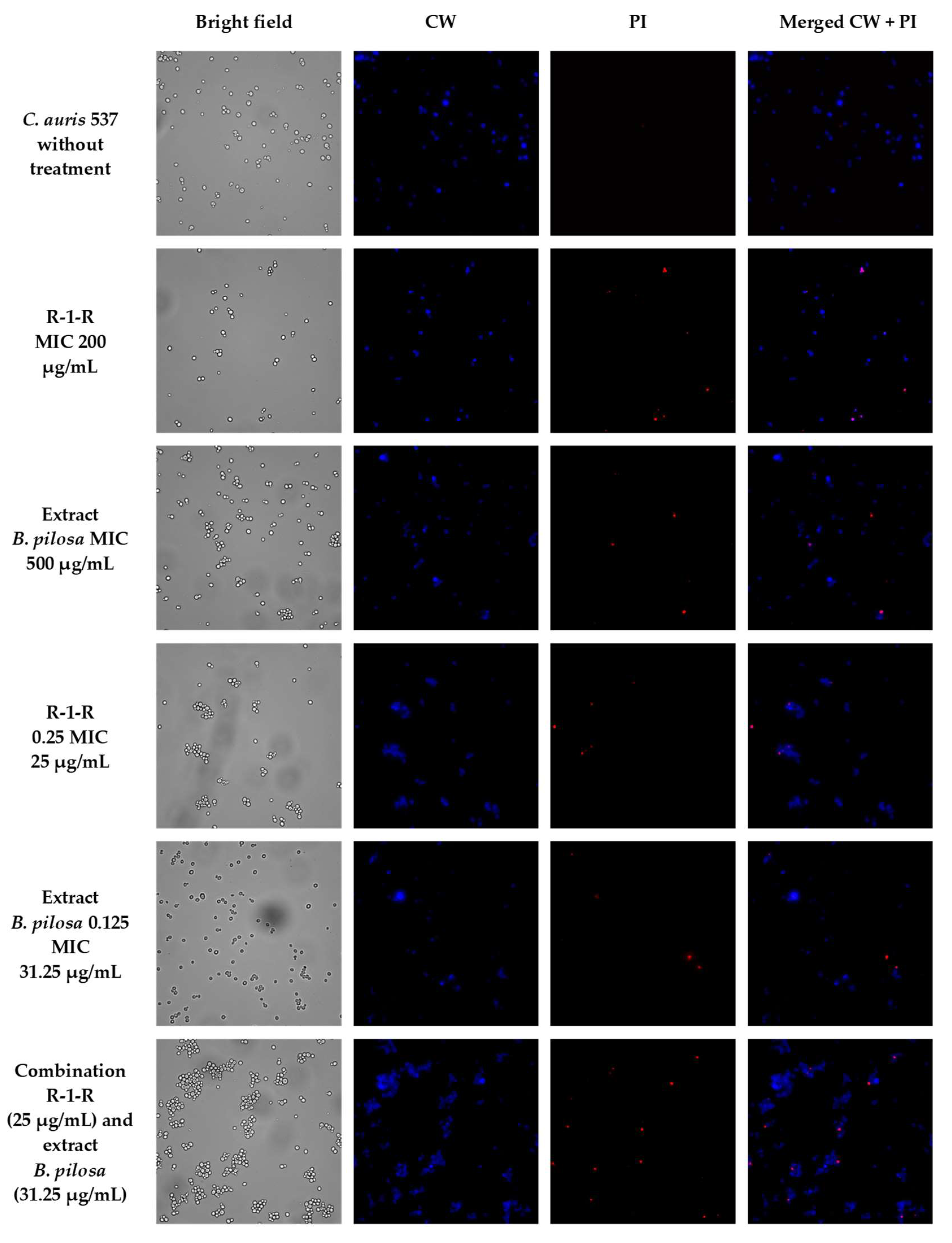
To visually capture the antifungal effect obtained by the peptide, the extract, and the combination with the greatest activity in the four study strains, confocal laser scanning microscopy (CLSM) (LIVE/DEAD Yeast commercial kit), and scanning electron microscopy (SEM) were performed.
The effect after 48 h of incubation with R-1-R, extract, or the mixture R-1-R/extract (at MICs and sub-MICs where there was a synergistic or additive effect) was evaluated for the four strains using the LIVE/DEAD™ Yeast Viability kit (
Figures S12–S15). Live control without treatment was used with a control of hypochlorite-treated dead yeast cells (results not shown). Live control cells were labeled with a blue fluorescence (Calcofluor White M2R), highlighting cell wall chitin regardless of metabolic state; yeasts were also labeled with FUN1 using two different excitation filters (488 and 532 nm), revealing yeast with a diffusely distributed green and red color, indicating plasma membrane integrity and the yeast cells’ metabolic capability to convert this intracellular staining to a red-orange fluorescence.
For
C. albicans SC5314 (
Figure S12), it was observed that although the metabolic viability was not affected, the number of cells was reduced compared to the live control. For example, with the peptide at 100 µg/mL, no presence of yeasts was observed. This concentration corresponds to the value of the MFC according to the technique of microdilution in broth (
Table 1). With the extract at 500 µg/mL (MIC), a decrease of more than ~50% in the cell population was seen. The peptide at 25 µg/mL (0.25 MIC) and the extract at 250 µg/mL (0.5 MIC) (concentrations where there was a synergic effect) also generated a reduction in the number of cells, but less than ~50%. Finally, when peptide (0.25 MIC) and extract (0.5 MIC) were evaluated in combination, it was observed that there was a reduction of approximately ~90% in the cell growth.
A similar effect was seen in the other study strains (
Figures S13–S15), with the difference that for
C. albicans 256 and
C. auris 435, in the case of combinations of peptide and extract, the number of cells was not reduced by more than ~50%, but rather the metabolic viability; this is inferred from the decreased red color staining in FUN1.
After exposing the yeasts for 48 h to 0.5 MIC, concentrations of peptide and extracts alone and in combination, by SEM, it was observed that compared to untreated yeast in
C. albicans SC5314 (
Figure 3a), the peptide at 50 µg/mL generates an alteration in the cell wall, forming protuberances in the cell in the form of yeast and indentations in the hyphae. On increasing the resolution of the image (500 nm), it was seen as a veil on the cell surface, which could correspond to leakage of the cytoplasmic content. With the extract at 250 µg/mL, it was observed that yeast has a more rounded appearance and there are cracks in the cell wall and blisters in the hyphae. In the case of the combination, a greater disturbance in the cell surface and inhibition of filamentation were observed.
For
C. albicans 256 (
Figure 3b), it was observed that there is also a disturbance in the membrane with peptide (50 µg/mL) and extract (250 µg/mL) alone; specifically, blisters were observed on the surface of the yeast, but in the combination (P: 50 µg/mL and E: 62.5 µg/mL), where there was an additive effect, the damage was more significant, with a rupture of the cell wall being observed. Another important aspect to point out is that compared to FLC-sensitive
C. albicans SC5314, for
C. albicans 256, which is resistant to FLC, the presence of hyphae was not observed in the untreated control.
When
C. auris 435 (
Figure 4a) and 537 (
Figure 4b) were treated with peptide (100 µg/mL) and extract (250 µg/mL) alone, damage to the cell wall with indentations and protuberances was also observed. With the combinations of peptide and extract there was rupture of the cell wall, and in
C. auris 537, even leakage of intracellular content is seen.
The results obtained are consistent with previous studies, carried out with peptides derived from LfcinB [
18,
19], where damage was also seen on the cell surface, and they are also consistent with the mechanism of action proposed for this AMP. Regarding the extract, this is the first study where SEM was performed and damage to the surface of the yeast was also observed. This could contribute to the results obtained in silico, where it has been proposed that the mechanism of action for extracts of
B. pilosa is directed to the cell membrane [
28].
To corroborate that the cell surface damage seen in SEM was related to the increased permeability in the cell membrane, the PI assay was performed at the value of the MICs, subinhibitory concentrations, and combinations of peptide and extract, where the synergistic or additive effect against the four study strains was seen (
Figure 5,
Figure 6,
Figure 7 and
Figure 8). After 4 h of incubation and in the case of peptide, combinations, and, to a lesser extent, for the extract (according to the strain), the presence of positive PI cells was found (red fluorescence) compared to untreated controls, highlighting a greater effect of permeability of the combinations in strains resistant to FLC.
The hemolytic activity of R-1-R,
B. Pilosa leaf extract, and their combinations was assessed against human erythrocytes in order to determine their effect on erythrocyte membrane integrity. The results showed that peptide and extract alone are safe and non-toxic at a wide range of concentrations, including the MIC and MFC, 1.56–200 and 15.62–2000 μg/mL, respectively. At high concentrations of 200 (peptide) and 2000 μg/mL (extract), a slight hemolytic effect on human red blood cells was observed, with percentages of 8% and 4%, respectively (
Figure 9); however, they are not considered hemolytic, because they have percentages of less than 10% [
91].
When R-1-R and B. pilosa leaf extract were evaluated in combination at concentrations where there was synergy or additivity, they did not exhibit significant hemolytic activity (0–2%); on the contrary, it was observed that all combinations decreased the hemolysis percentage by a factor of 2 in comparison with the peptide or the extract alone, while the positive control Tween-20 completely lysed the red blood cells.
In vitro toxicity testing can reveal the risk that may be associated with the use of AMPs and metabolites from plant extracts. In previous studies, Lys/Arg substitutions decreased the hemolytic activity of peptide R-1-R, preserving the antibacterial activity [
70]. In the present study, it was demonstrated that the combination of R-1-R and the extract of
B. pilosa is also an option for reducing the percentage of hemolysis, but it also increases the antifungal activity. This is mainly due to the fact that a greater antifungal activity is achieved with lower concentrations of peptide and extract.
On the other hand, many plants contain chemical substances that might have a hemolytic effect on erythrocytes. The ability of the plant extracts to lyse red blood cells could be dependent on different tannins and saponins that can cause cell disruption [
92,
93]; however, neither of these two metabolites was found in the extract from
B. pilosa leaves. In contrast to the metabolites found (
Table 2), it has not been found to exhibit considerable hemolytic activity at high concentrations [
78,
94,
95]. Additionally, a study showed that both ethanol and ethylacetate/ethanol extracts from
B. pilosa plants had a protective effect in normal human erythrocytes against oxidative hemolysis in vitro [
96].
Peptide, extract, and their synergistic combinations were subjected to cytotoxicity analysis on a primary cell culture of fibroblasts. The IC
50 and R
2 values were 175.9 μg/mL and 0.69 μg/mL, respectively, for R-1-R, while for
B. pilosa leaf extracts, the IC
50 and R
2 values were 657.6 μg/mL and 0.96 μg/mL, respectively. The peptide showed no appreciable toxicity against fibroblasts at 1.56–150 μg/mL, with percentages of cell viability above 50%. However, the extract at concentrations above 1000 μg/mL decreased the percentage of cell viability by more than 50% (
Figure 10). Interestingly, when they were evaluated in combination concentrations where there was synergy or additivity, no toxic effect was observed at the tested concentrations of peptide and extract (C1: 25/250 µg/mL, C2: 50/62.5 µg/mL, C3: 100/125 µg/mL, and C4: 25/31.25 µg/mL), with percentages of cell viability above 80%.
Similarly, Barragán-Cárdenas et al. have reported that this palindromic peptide exhibits a rapid cytotoxic effect on breast cancer cells. They also tested its selectivity on primary fibroblasts, non-cancerous MCF-12 cells, and BMEC bovine mammary cells, with minimal effect on them [
47,
48].
Phytochemical analysis has revealed that
B. pilosa leaf extracts contain a broad chemical constitution, suggesting that the cytotoxicity activity could be related to secondary metabolites such as terpenes, flavonoids, and alkaloids [
97]. In previous studies, it was suggested that the strong in vitro activity at high concentrations is related to other mechanisms, such as interference with mitochondrial or lysosomal function [
98].
However, the combination did not induce any adverse effects on the fibroblast cells and erythrocytes. Several studies have demonstrated that the combination of drugs offers numerous advantages over monotherapy, which include reduced dosage, synergistic response, decreased possibility of resistance development, and reduced toxicity [
99]. Combinations of antimicrobials and NPs that exhibit synergism will possibly show effective pharmacological results [
49].
It is possible that secondary metabolites such as hydroxycinnamic acid (HCA) derivatives such as caffeoylquinic acids, flavonoid glycosides, and the di-caffeoylquinic acids and their 3,4- and 3,5-geometric isomers that have been found in
B. pilosa may improve or facilitate the interaction with the peptide. This strategy has helped prevent the development of resistance while also minimizing toxicity, by allowing the use of lower concentrations of both agents [
100]. In order to comprehensively comprehend the potential health advantages and associated risks related to their consumption, additional research is required.


 ,
, 



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}









