
Coffee Leaf Rust (Hemileia vastatrix) from the Recent Invasion into Hawaii Shares a Genotypic Relationship with Latin American Populations

 ,
,  ,
,  ,
, {kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
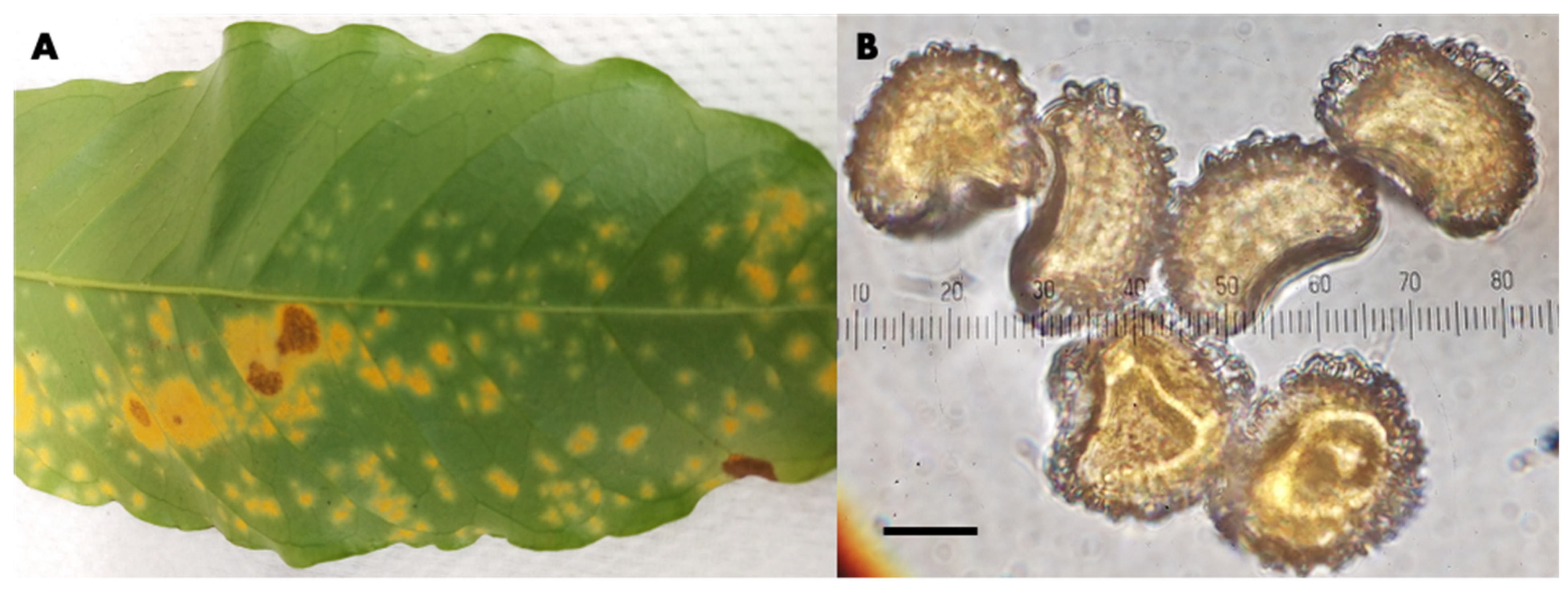
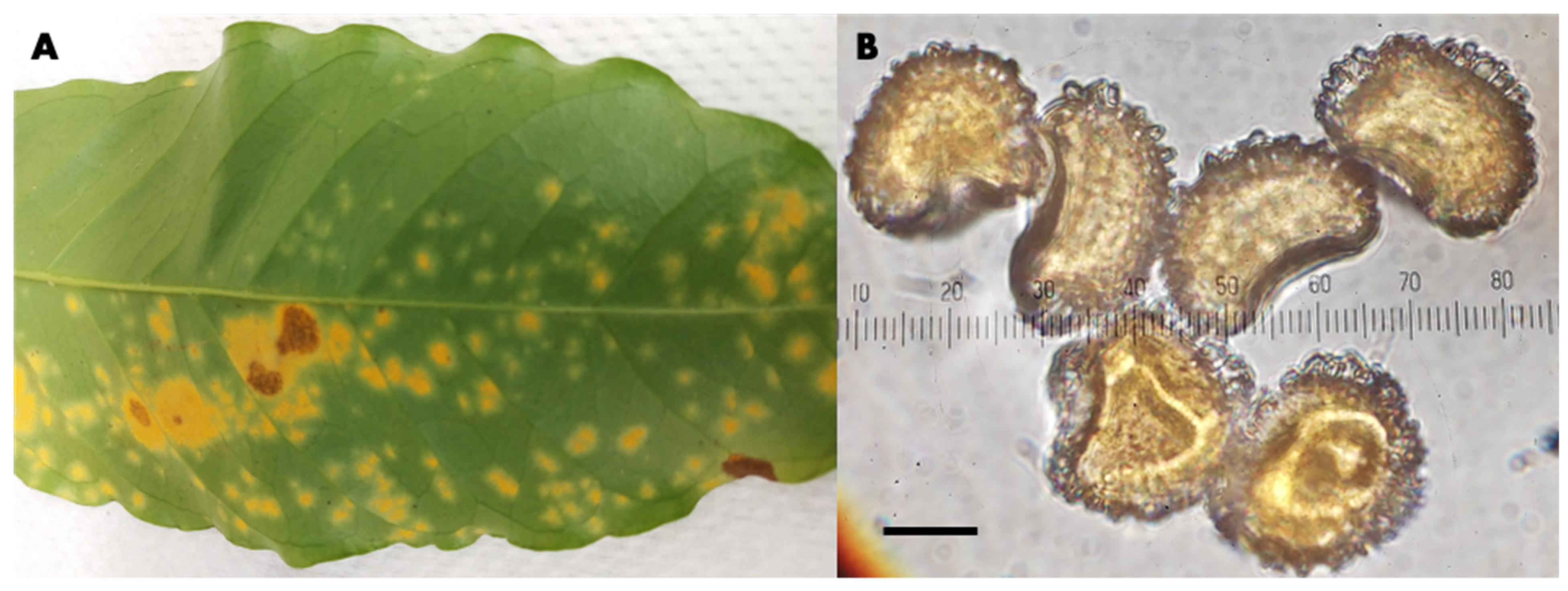
Hemileia vastatrix Specimens
3. PCR, Sequencing, and SSRs
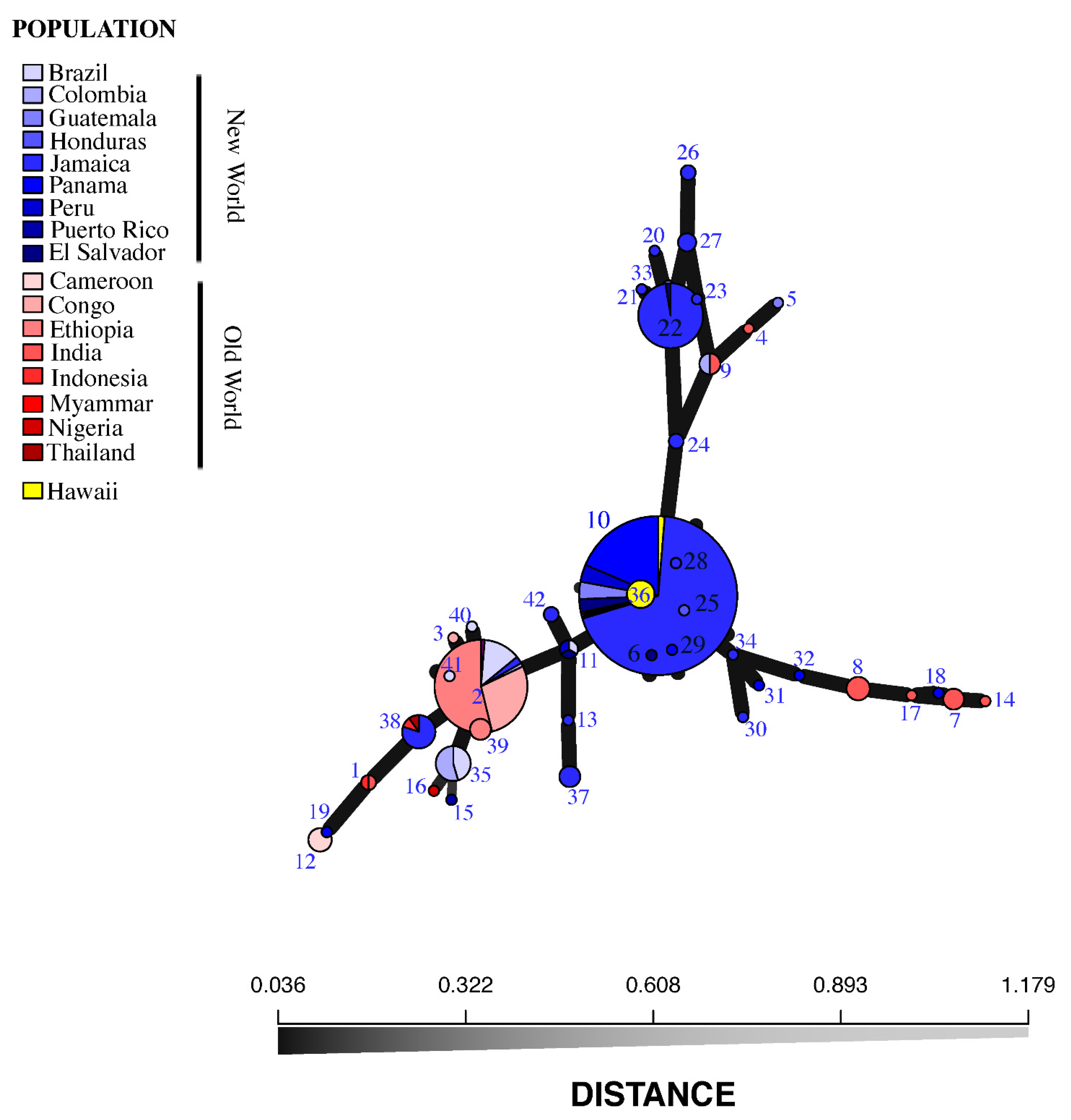
4. SSRs and Minimum Spanning Network
5. Wind Pattern Visualizations
6. Results
7. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Berkeley, M.J.; Broome, C.E. Hemileia vastatrix. Gard. Chron. 1869, 6, 1157. [Google Scholar]
- Vega, F.E.; Rosenquist, E.; Collins, W. Global project needed to tackle coffee crisis. Nature 2003, 425, 343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCook, S. Global rust belt: Hemileia vastatrix and the ecological integration of world coffee production since 1850. J. Glob. Hist. 2006, 1, 177–195. [Google Scholar] [CrossRef] [Green Version]
- Waller, J.M.; Bigger, M.; Hillocks, R.J. Coffee Pests, Diseases and Their Management; CAB Books; CABI Publishing: Wallingford, UK, 2007; ISBN 9781845932091. [Google Scholar]
- Hindorf, H.; Omondi, C.O. A review of three major fungal diseases of Coffea arabica L. in the rainforests of Ethiopia and progress in breeding for resistance in Kenya. J. Adv. Res. 2011, 2, 109–120. [Google Scholar] [CrossRef] [Green Version]
- Callan, B.E.; Carris, L.M. Fungi on living plant substrata, including fruits. In Biodiversity of Fungi; Elsevier: Boston, MA, USA, 2004; ISBN 9780080470269. [Google Scholar]
- Reuben, S.O.W.M.; Mtenga, D.J.I. Variation in resistance to coffee leaf rust (Hemileia vastatrix, Berk. and Broome) among germplasm progenitors at the Tanzanian Coffee Research Institute (TACRI). Int. J. Agric. Sci. 2012, 2, 296–300. [Google Scholar]
- Rodrigues, C.J.; Bettencourt, A.J.; Rijo, L. Races of the pathogen and resistance to coffee rust. Annu. Rev. Phytopathol. 1975, 13, 49–70. [Google Scholar] [CrossRef]
- Zambolim, L. Current status and management of coffee leaf rust in Brazil. Trop. Plant Pathol. 2016, 41, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Rhiney, K.; Guido, Z.; Knudson, C.; Avelino, J.; Bacon, C.M.; Leclerc, G.; Aime, M.C.; Bebber, D.P. Epidemics and the future of coffee production. Proc. Natl. Acad. Sci. USA 2021, 118, e2023212118. [Google Scholar] [CrossRef]
- Avelino, J.; Willocquet, L.; Savary, S. Effects of crop management patterns on coffee rust epidemics. Plant Pathol. 2004, 53, 541–547. [Google Scholar] [CrossRef]
- McCook, S. Coffee is Not Forever: A Global History of the Coffee Leaf Rust; Ohio University Press: Athens, OH, USA, 2019; ISBN 978-0-8214-2387-5. [Google Scholar]
- McCook, S.; Vandermeer, J. The Big Rust and the Red Queen: Long-term perspectives on coffee rust research. Phytopathology 2015, 105, 1164–1173. [Google Scholar] [CrossRef] [Green Version]
- De Filho, E.V.; Domian, C.A. Prevention and Control of Coffee Leaf Rust: Handbook of Best Practices for Extension Agents and Facilitators, 1st ed.; Tropical Agricultural Research and Higher Education Center (CATIE): Turrialba, Costa Rica, 2019; ISBN 9789977577043. [Google Scholar]
- Gardner, D.E. Additions to the rust fungi of Hawaii. Pacific Sci. 1997, 51, 174–182. [Google Scholar]
- Gardner, D.E.; Hodges, C.S. The rust fungi (Uredinales) of Hawaii. Pacific Sci. 1989, 43, 41–55. [Google Scholar]
- Scholler, M.; Aime, M.C. On some rust fungi (Uredinales) collected in an Acacia koa-Metrosideros polymorpha woodland, Mauna Loa Road, Big Island, Hawaii. Mycoscience 2006, 47, 159–165. [Google Scholar] [CrossRef]
- Ocenar, J.; Kawabata, A. Coffee leaf rust. In Hawaii Department of Agriculture; 2021; pp. 1–5. Available online: https://hdoa.hawaii.gov/pi/files/2020/12/NPA-20-03-Coffee-leaf-rust-12-20.pdf (accessed on 15 January 2022).
- Keith, L.; Sugiyama, L.; Brill, E.; Adams, B.-L.; Fukada, M.; Hoffman, K.; Ocenar, J.; Kawabata, A.; Kong, A.; McKemy, J.; et al. First report of coffee leaf rust caused by Hemileia vastatrix on coffee (Coffea arabica) in Hawaii. Plant Dis. 2021. [Google Scholar] [CrossRef] [PubMed]
- Kawabata, A.M.; Nakamoto, S.T. Spraying to suppress coffee leaf rust (Hemileia vastatrix) in Hawaii. Univ. Hawaii 2021, 1–5. Available online: https://www.ctahr.hawaii.edu/oc/freepubs/pdf/PD-118.pdf (accessed on 15 January 2022).
- Kawabata, A.; Wages, S.; Nakamoto, S. Pruning methods for the management of coffee leaf rust and coffee berry borer in Hawaii. Univ. Hawaii Coop. Ext. Serv. 2021, 1–9. [Google Scholar]
- Chapman, E.G.; Messing, R.H.; Harwood, J.D. Determining the origin of the coffee berry borer invasion of Hawaii. Ann. Entomol. Soc. Am. 2015, 108, 585–592. [Google Scholar] [CrossRef]
- Grebner, D.L.; Jacobson, M.G.; Fajvan, M.A. Forest ecosystem services and the scourge of invasive species. J. For. 2014, 112, 620–622. [Google Scholar] [CrossRef]
- Kilambo, D.L.; Reuben, S.O.W.M.; Mamiro, D. Races of Hemileia vastatrix and variation in pathogenicity of Colletotrichum kahawae isolates to compact coffee genotypes in Tanzania. J. Plant Stud. 2013, 2, 95–104. [Google Scholar] [CrossRef] [Green Version]
- Cabral, P.G.C.; Maciel-Zambolim, E.; Oliveira, S.A.S.; Caixeta, E.T.; Zambolim, L. Genetic diversity and structure of Hemileia vastatrix populations on Coffea spp. Plant Pathol. 2016, 65, 196–204. [Google Scholar] [CrossRef] [Green Version]
- Gouveia, M.M.C.; Ribeiro, A.; Várzea, V.M.P.; Rodrigues, C.J. Genetic diversity in Hemileia vastatrix based on RAPD markers. Mycologia 2005, 97, 396–404. [Google Scholar] [CrossRef]
- Nunes, C.C.; Maffia, L.A.; Mizubuti, E.S.G.; Brommonschenkel, S.H.; Silva, J.C. Genetic diversity of populations of Hemileia vastatrix from organic and conventional coffee plantations in Brazil. Australas. Plant Pathol. 2009, 38, 445–452. [Google Scholar] [CrossRef]
- Rozo, Y.; Escobar, C.; Gaitán, Á.; Cristancho, M. Aggressiveness and genetic diversity of Hemileia vastatrix during an epidemic in Colombia. J. Phytopathol. 2012, 160, 732–740. [Google Scholar] [CrossRef]
- Ramírez-Camejo, L.A.; Eamvijarn, A.; Díaz-Valderrama, J.R.; Karlsen-Ayala, E.; Koch, R.; Johnson, E.; Pruvot-Woehl, S.; Mejía, L.C.; Montagnon, C.; Maldonado-Fuentes, C.; et al. Global analysis of Hemileia vastatrix populations shows clonal reproduction for the coffee leaf rust pathogen throughout most of its range. Phytopathology 2021. [Google Scholar] [CrossRef] [PubMed]
- Batista, D.; Guerra-guimarães, L.; Talhinhas, P.; Loureiro, A.; Silva, D.N.; Gonzalez, L.; Pereira, A.P.; Vieira, A.; Azinheira, H.G.; Struck, C.; et al. Analysis of population genetic diversity and differentiation in Hemileia vastatrix by molecular markers. In Proceedings of the 23rd International Conference on Coffee Science, Bali, Indonesia, 3–8 October 2010; pp. 738–742. [Google Scholar]
- Aime, M.C. Toward resolving family-level relationships in rust fungi (Uredinales). Mycoscience 2006, 47, 112–122. [Google Scholar] [CrossRef]
- Karaoglu, H.; Lee, C.M.Y.; Meyer, W. Survey of simple sequence repeats in completed fungal genomes. Mol. Biol. Evol. 2005, 22, 639–649. [Google Scholar] [CrossRef]
- Schuelke, M. An economic method for the fluorescent labeling of PCR fragments—A poor man’ s approach to genotyping for research and high-throughput diagnostics. Nat. Biotechnol. 2000, 18, 233–234. [Google Scholar] [CrossRef]
- Díaz-Valderrama, J.R.; Aime, M.C. The cacao pathogen Moniliophthora roreri (Marasmiaceae) possesses biallelic A and B mating loci but reproduces clonally. Heredity 2016, 116, 491–501. [Google Scholar] [CrossRef]
- Koch, R.A.; Aime, M.C. Population structure of Guyanagaster necrorhizus supports termite dispersal for this enigmatic fungus. Mol. Ecol. 2018, 27, 2667–2679. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GENALEX 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GenALEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research—An update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamvar, Z.N.; Brooks, J.C.; Grünwald, N.J. Novel R tools for analysis of genome-wide population genetic data with emphasis on clonality. Front. Genet. 2015, 6, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamvar, Z.N.; Tabima, J.F.; Grünwald, N.J. Poppr: An R package for genetic analysis of populations with clonal, partially clonal, and/or sexual reproduction. PeerJ 2014, 2, e281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olivera, P.; Newcomb, M.; Szabo, L.J.; Rouse, M.; Johnson, J.; Gale, S.; Luster, D.G.; Hodson, D.; Cox, J.A.; Burgin, L.; et al. Phenotypic and genotypic characterization of race TKTTF of Puccinia graminis f. sp. tritici that caused a wheat stem rust epidemic in southern Ethiopia in 2013–14. Phytopathology 2015, 105, 917–928. [Google Scholar] [CrossRef] [Green Version]
- Aime, M.C.; Bell, C.D.; Wilson, A.W. Deconstructing the evolutionary complexity between rust fungi (Pucciniales) and their plant hosts. Stud. Mycol. 2018, 89, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Cheng, P.; Chen, X.M. Virulence and molecular analyses support asexual reproduction of Puccinia striiformis f. sp. tritici in the U.S. Pacific Northwest. Phytopathology 2014, 104, 1208–1220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drenth, A.; McTaggart, A.R.; Wingfield, B.D. Fungal clones win the battle, but recombination wins the war. IMA Fungus 2019, 10, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McTaggart, A.R.; du Plessis, E.; Roux, J.; Barnes, I.; Fraser, S.; Granados, G.M.; Ho, W.W.H.; Shuey, L.S.; Drenth, A. Sexual reproduction in populations of Austropuccinia psidii. Eur. J. Plant Pathol. 2020, 156, 537–545. [Google Scholar] [CrossRef]
- Purdy, L.H.; Krupa, S.V.; Dean, J.L. Introduction of sugarcane rust into the Americas and its spread to Florida. Plant Dis. 1985, 69, 689–693. [Google Scholar] [CrossRef]
- Wyrtki, K.; Meyers, G. The trade wind field over the Pacific Ocean. J. Appl. Meteorol. 1976, 15, 698–704. [Google Scholar] [CrossRef]
- Garza, J.A.; Chu, P.S.; Norton, C.W.; Schroeder, T.A. Changes of the prevailing trade winds over the islands of Hawaii and the North Pacific. J. Geophys. Res. Atmos. 2012, 117, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Berg, R.; Kimberlain, T. The 2014 Eastern North Pacific hurricane season: A very active season brings devastation. Weatherwise 2015, 68, 36–45. [Google Scholar] [CrossRef]
- Tipton, L.; Zahn, G.; Datlof, E.; Kivlin, S.N.; Sheridan, P.; Amend, A.S.; Hynson, N.A. Fungal aerobiota are not affected by time nor environment over a 13-y time series at the Mauna Loa Observatory. Proc. Natl. Acad. Sci. USA 2019, 116, 25728–25733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowden, J.; Gregory, P.H.; Johnson, C.G. Possible wind transport of coffee leaf rust across the Atlantic Ocean. Nature 1971, 229, 500–501. [Google Scholar] [CrossRef]
- Kellogg, C.A.; Griffin, D.W. Aerobiology and the global transport of desert dust. Trends Ecol. Evol. 2006, 21, 638–644. [Google Scholar] [CrossRef]
- Maddison, A.C.; Manners, J.G. Lethal effects of artificial ultraviolet radiation on cereal rust uredospores. Trans. Br. Mycol. Soc. 1973, 60, 471–494. [Google Scholar] [CrossRef]
- Philipp, P. Diversified Agriculture of Hawaii: An Economist’s View of Its History, Present Status, and Future Prospects; University of Hawaii Press: Honolulu, HI, USA, 1953. [Google Scholar]
- Goto, B. Ethnic groups and the coffee industry in Hawai’i. Hawaii J. Hist. 1825, 16, 112–124. [Google Scholar]
- Higa, S. Plant Importation Rules and Regulations. 1992, pp. 707–711. Available online: http://hear.org/books/apineh1992/pdfs/apineh1992vii1higa.pdf (accessed on 15 January 2022).
- Krishnan, S.; Matsumoto, T.; Nagai, C.; Falconer, J.; Shriner, S.; Long, J.; Medrano, J.F.; Vega, F.E. Vulnerability of coffee (Coffea spp.) genetic resources in the United States. Genet. Resour. Crop Evol. 2021, 68, 2691–2710. [Google Scholar] [CrossRef]
- HDOA. Imported Coffee-Growing Kits Confiscated; HDOA: Honolulu, HI, USA, 2017.
- Loope, L. A Summary of Information on the Rust Puccinia Psidii Winter (Guava Rust) with Emphasis on Means to Prevent Introduction of Additional Strains to Hawaii; US Geological Survey: Reston, VA, USA, 2010.
- Uchida, J.; Zhong, S.; Killgore, E. First report of a rust disease on Ohia caused by Puccinia psidii in Hawaii. Plant Dis. 2006, 90, 524. [Google Scholar] [CrossRef]
- Rayner, R.W. Germination and penetration studies on coffee rust (Hemileia vastatrix B. & Br.). Ann. Appl. Biol. 1961, 49, 497–505. [Google Scholar] [CrossRef]
- Pemberton, C.E. Highlights in the history of entomology in Hawaii 1778–1963. Pac. Insects 1964, 6, 689–729. [Google Scholar]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramírez-Camejo, L.A.; Keith, L.M.; Matsumoto, T.; Sugiyama, L.; Fukada, M.; Brann, M.; Moffitt, A.; Liu, J.; Aime, M.C. Coffee Leaf Rust (Hemileia vastatrix) from the Recent Invasion into Hawaii Shares a Genotypic Relationship with Latin American Populations. J. Fungi 2022, 8, 189. https://doi.org/10.3390/jof8020189
Ramírez-Camejo LA, Keith LM, Matsumoto T, Sugiyama L, Fukada M, Brann M, Moffitt A, Liu J, Aime MC. Coffee Leaf Rust (Hemileia vastatrix) from the Recent Invasion into Hawaii Shares a Genotypic Relationship with Latin American Populations. Journal of Fungi. 2022; 8(2):189. https://doi.org/10.3390/jof8020189
Chicago/Turabian StyleRamírez-Camejo, Luis A., Lisa M. Keith, Tracie Matsumoto, Lionel Sugiyama, Mach Fukada, Mia Brann, Ariana Moffitt, Jingyu Liu, and M. Catherine Aime. 2022. "Coffee Leaf Rust (Hemileia vastatrix) from the Recent Invasion into Hawaii Shares a Genotypic Relationship with Latin American Populations" Journal of Fungi 8, no. 2: 189. https://doi.org/10.3390/jof8020189
APA StyleRamírez-Camejo, L. A., Keith, L. M., Matsumoto, T., Sugiyama, L., Fukada, M., Brann, M., Moffitt, A., Liu, J., & Aime, M. C. (2022). Coffee Leaf Rust (Hemileia vastatrix) from the Recent Invasion into Hawaii Shares a Genotypic Relationship with Latin American Populations. Journal of Fungi, 8(2), 189. https://doi.org/10.3390/jof8020189