
Lauric Acid Induces Apoptosis of Rice Sheath Blight Disease Caused by Rhizoctonia solani by Affecting Fungal Fatty Acid Metabolism and Destroying the Dynamic Equilibrium of Reactive Oxygen Species


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Methods
2.2.1. Isolation, Culture, and Determination of Minimum Inhibitory Concentration and Lethal Dose of Rice Sheath Blight
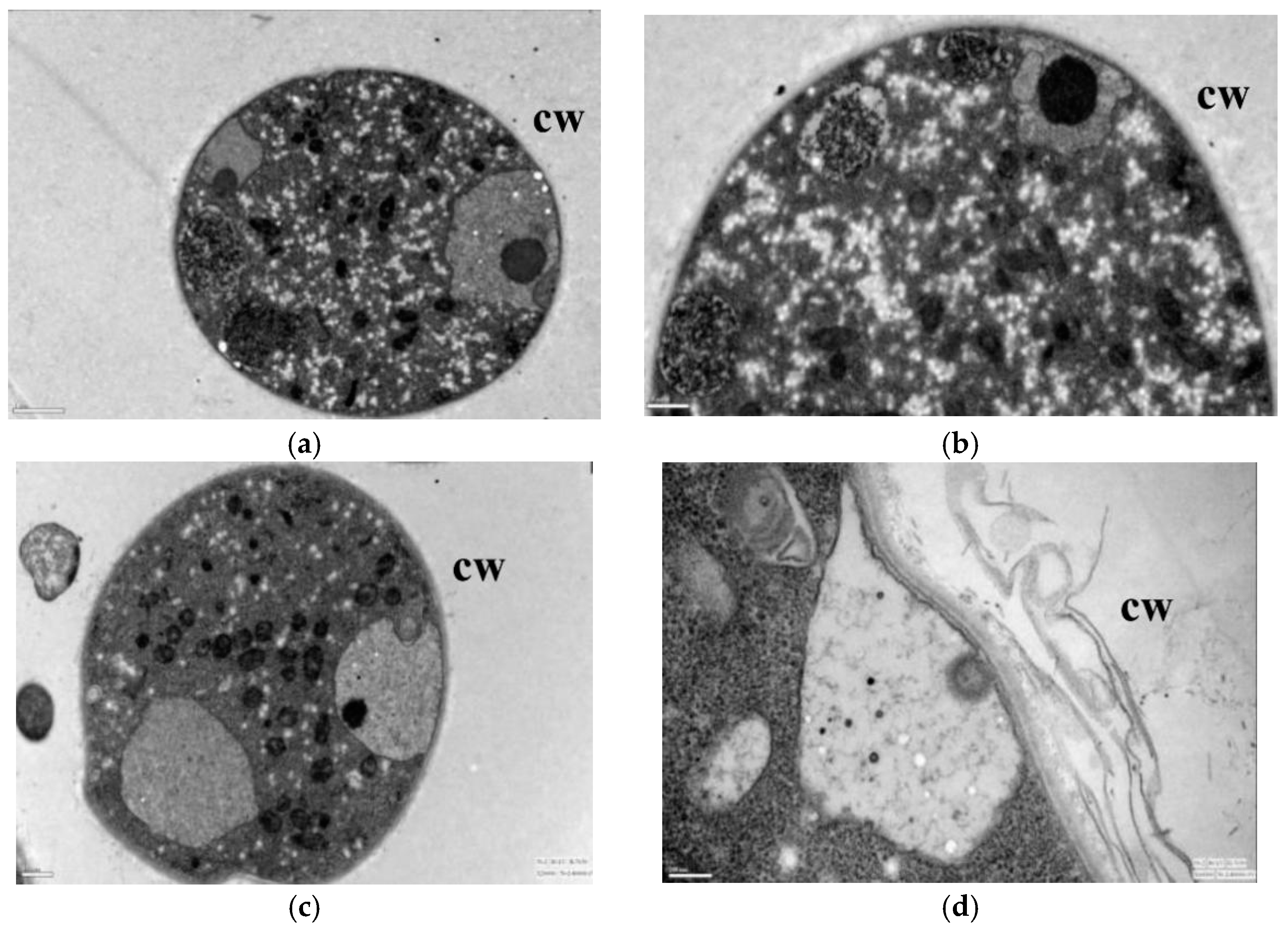
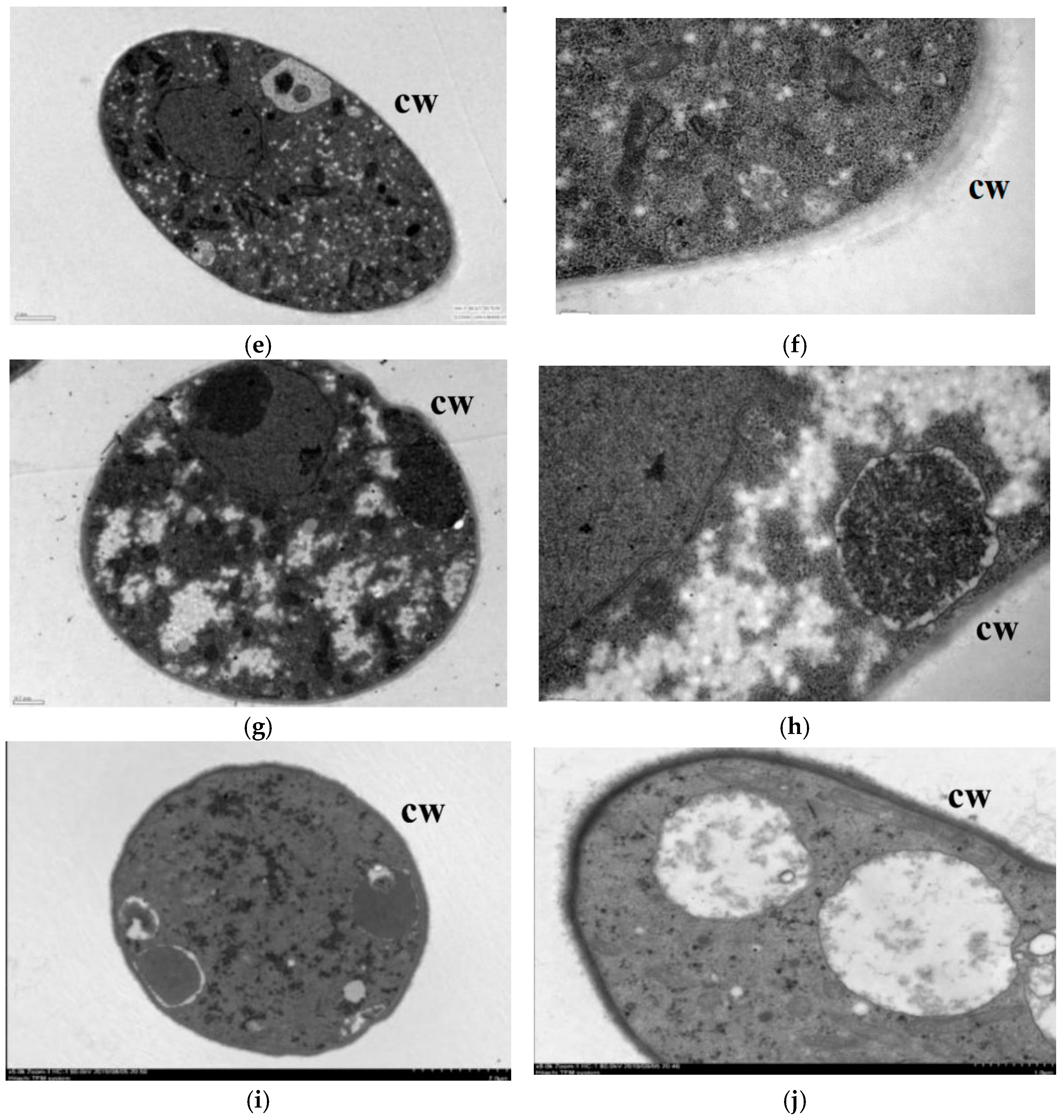
2.2.2. Scanning Electron Microscopy and Transmission Electron Microscopy Observations
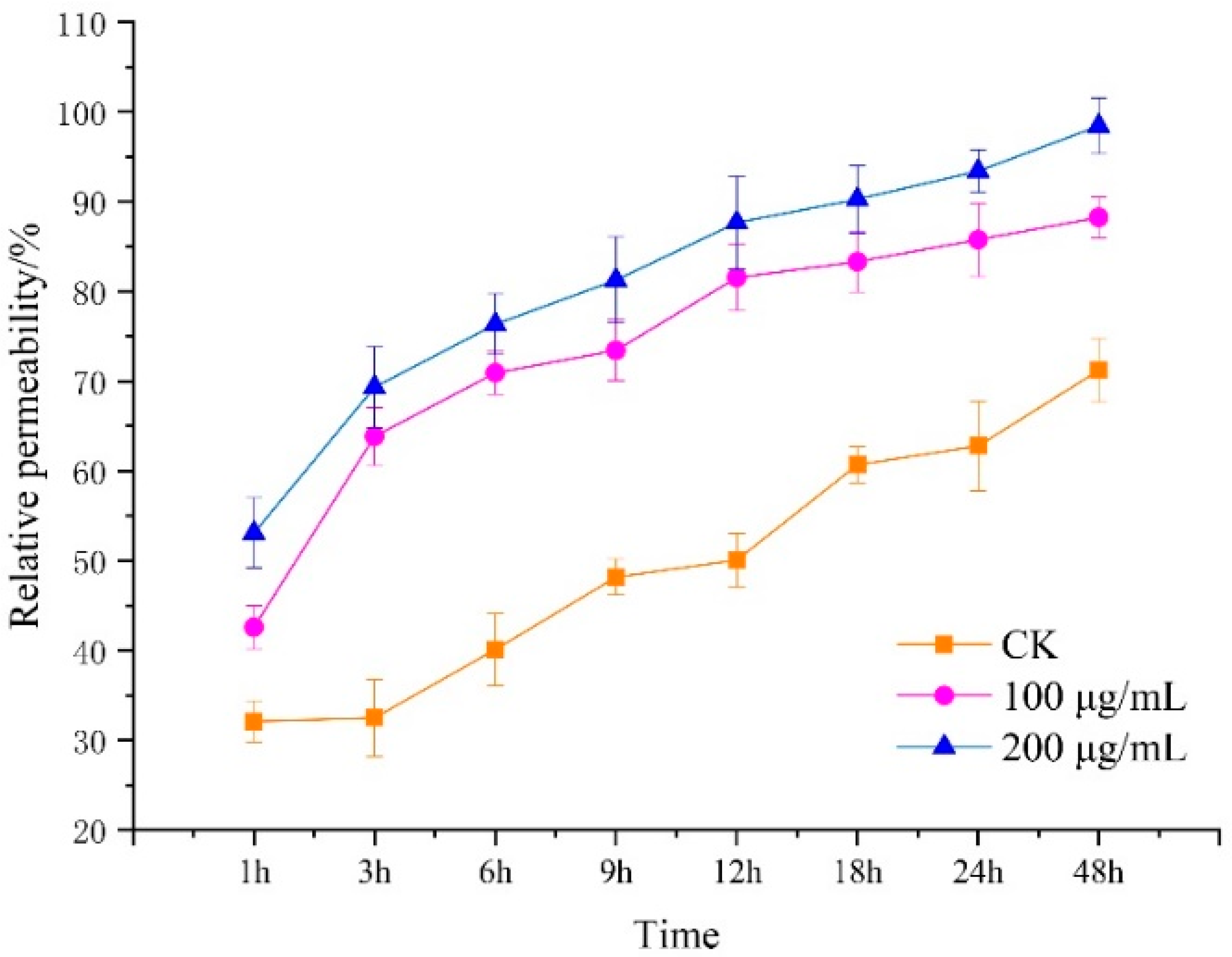
2.2.3. Determination of the Cellular Permeability of R. solani to Lauric Acid
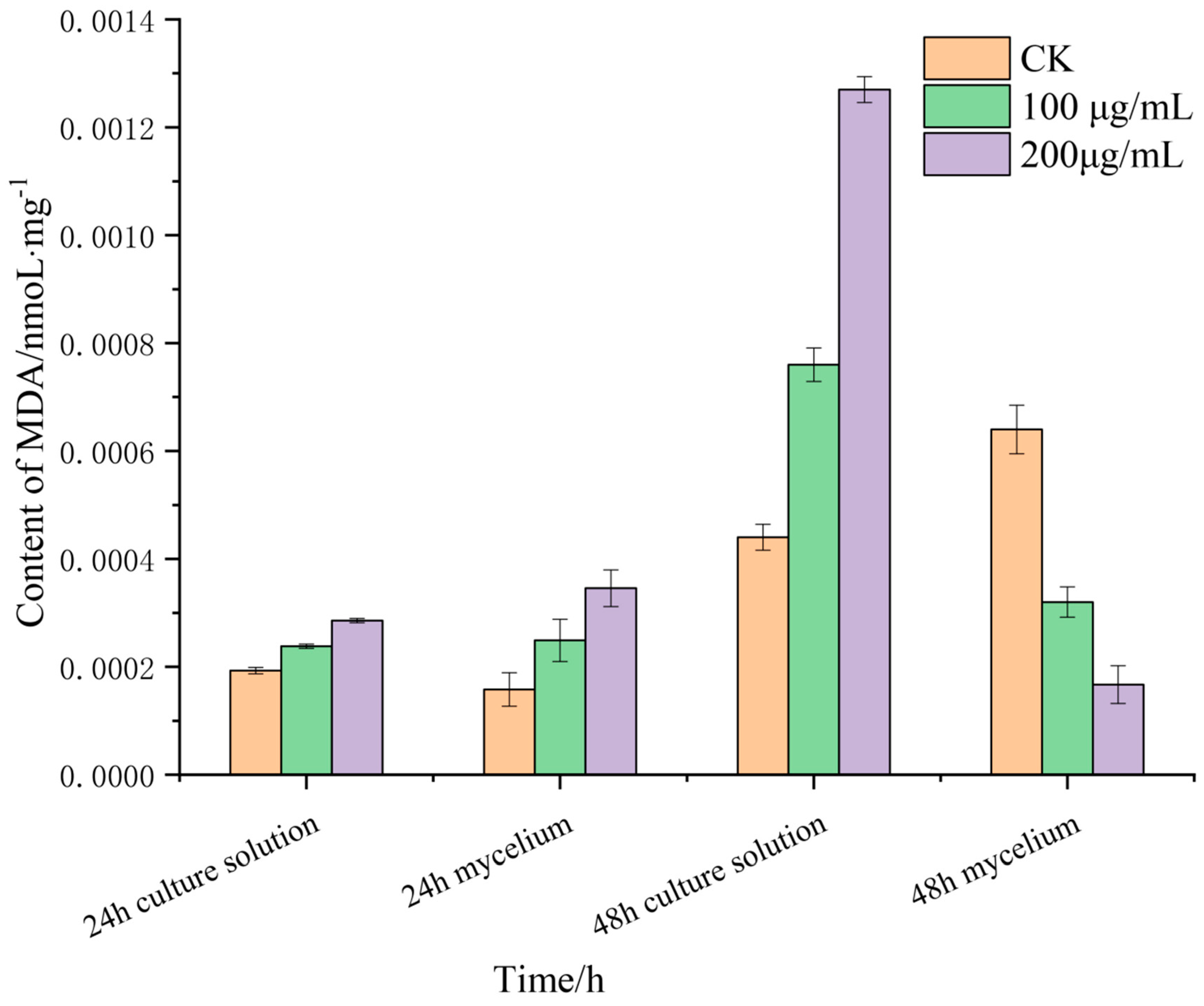
2.2.4. Determination of Malondialdehyde Activity of Lauric Acid
2.2.5. Determination of Hyphal Ergosterol Content
2.2.6. Transcriptome Analysis of Lauric Acid-Treated R. solani Mycelium
2.2.7. Bioinformatics Analysis of Sequencing Data
2.2.8. Differential Gene Expression, Gene Ontology Enrichment, and Kyoto Encyclopedia of Genes and Genomes Pathway Enrichment Analyses
3. Results
3.1. Minimum Inhibitory Concentration and Lethal Dose of Lauric Acid against R. solani Hyphae
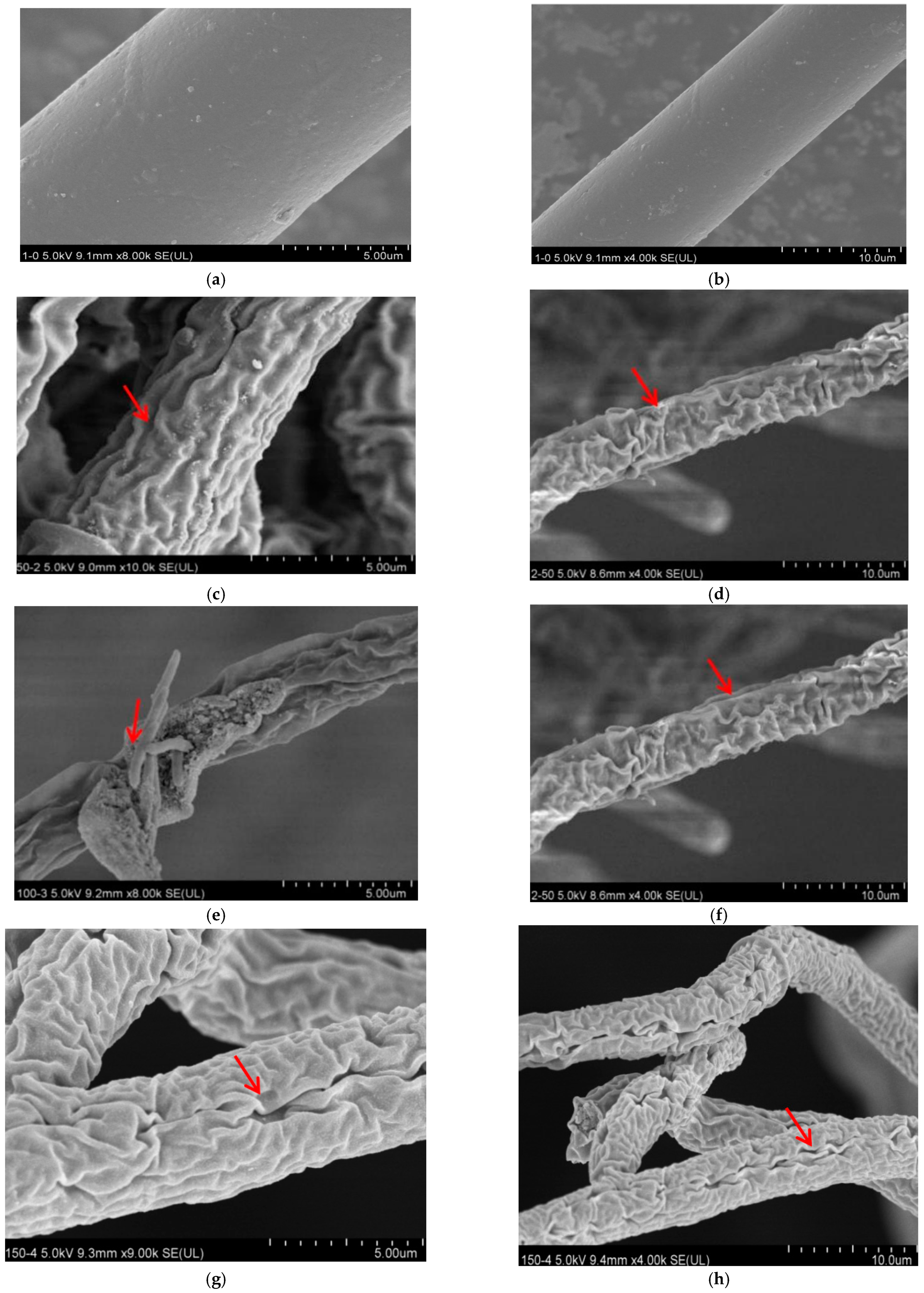
3.2. Effect of Lauric Acid on the Hyphal Morphology of R. solani
3.3. Effect of Lauric Acid on the Permeability of Hyphal Cells of R. solani
3.4. Effects of Lauric Acid on the Malondialdehyde Content R. solani Hyphae
3.5. Effect of Lauric Acid on the Ergosterol Content of R. solani Hyphae
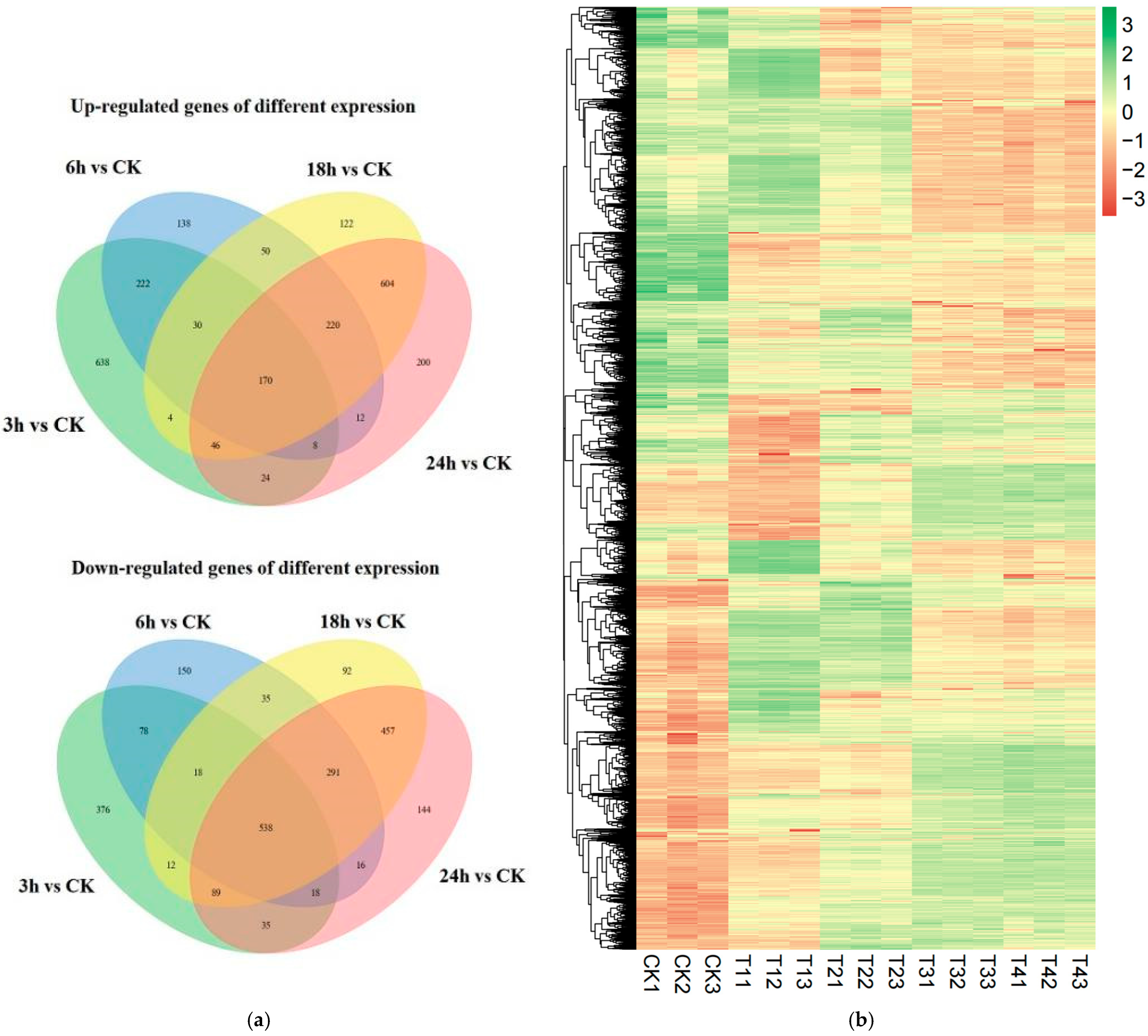
3.6. Transcriptome Analysis of R. solani Treated with Lauric Acid
3.7. Identification of Differentially Expressed Genes (DEGs)
3.8. Gene Ontology and Kyoto Encyclopedia of Genes and Genomes Analyses of Enriched Differentially Expressed Genes after Different Treatments
3.9. Analysis of Starch and Sucrose Metabolism Pathways in R. solani Induced by Lauric Acid
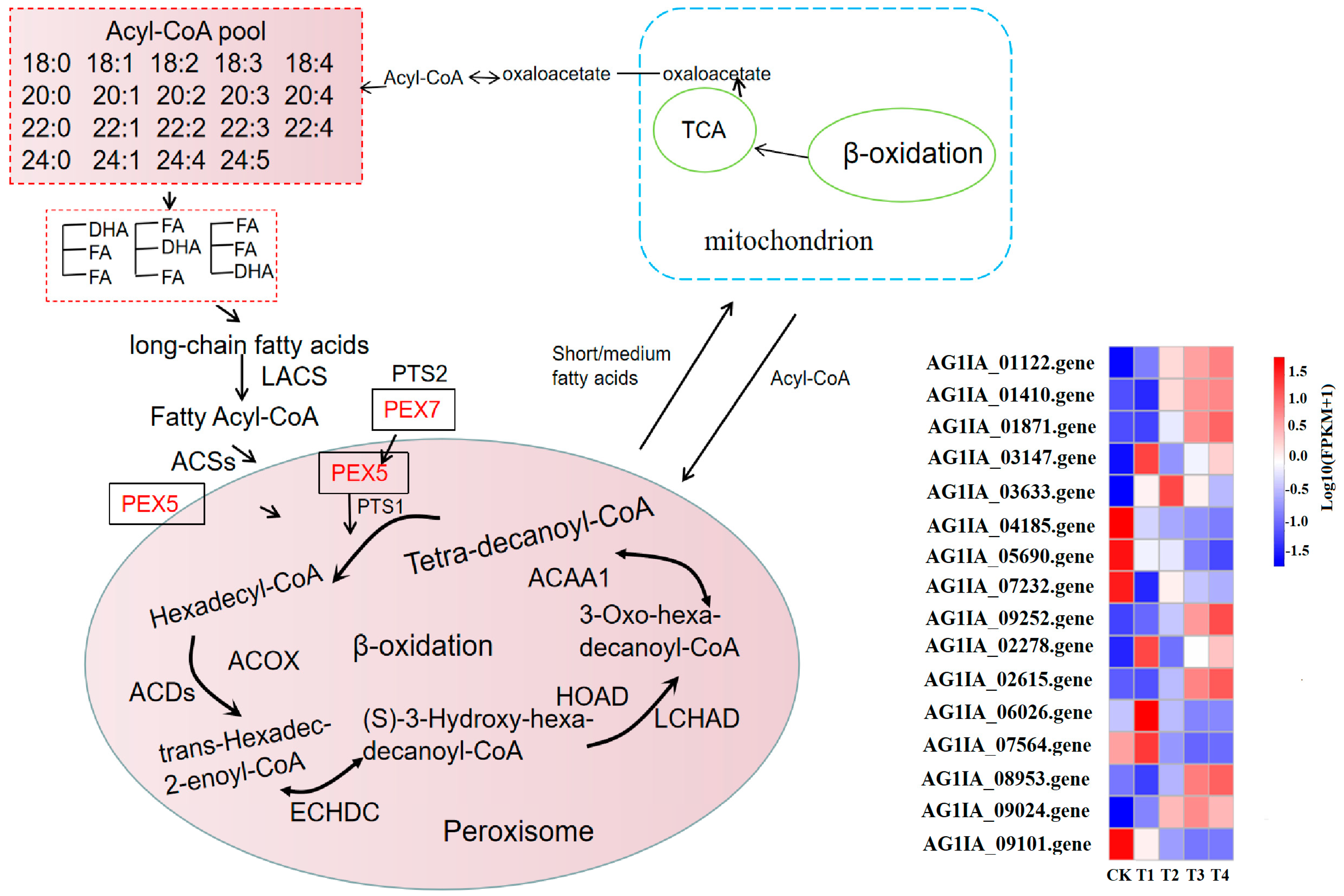
3.10. Effect of Lauric Acid on the Fatty Acid Metabolism of R. solani
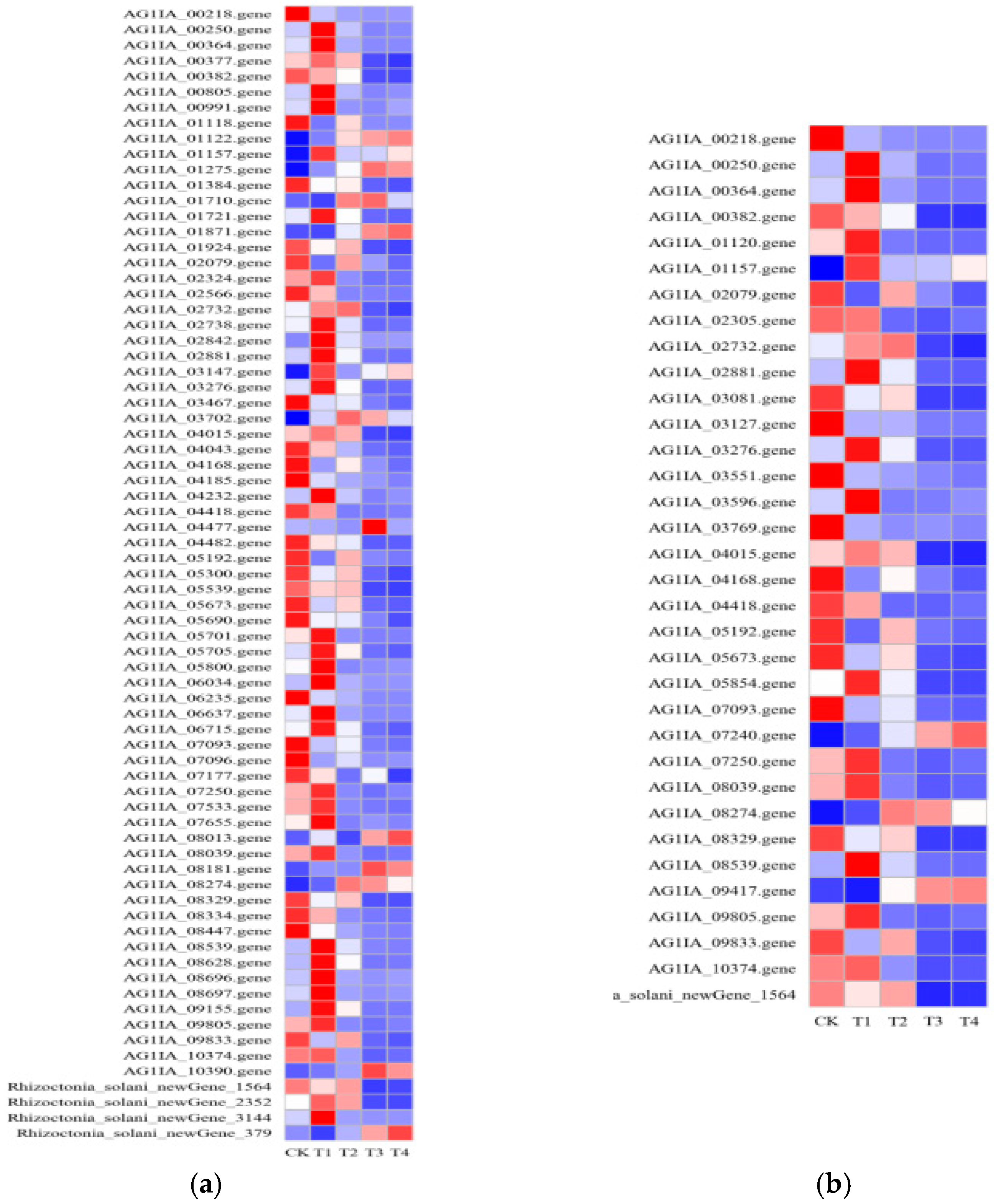
3.11. Effects of Lauric Acid on the Antibiotic Biosynthesis and Carbon Metabolism of R. solani
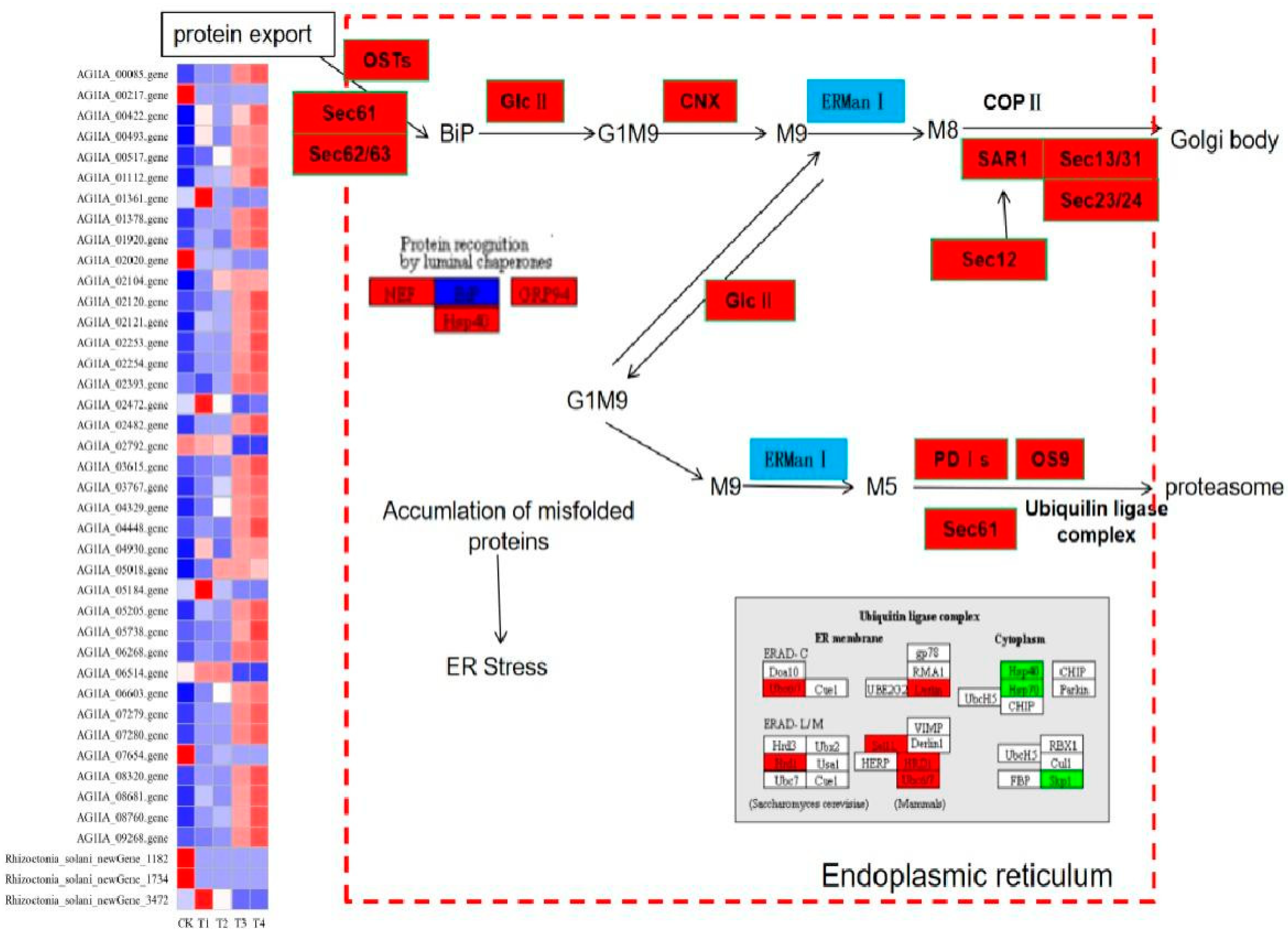
3.12. Effect of Lauric Acid on the Endoplasmic Reticulum Protein Processing Pathway of Rhizoctonia solani
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Rajendra, P.; Ayub, K.; Wendy-Ann, I.; Wayne, G.; Duraisamy, S. Plant extracts, bioagents and new generation fungicides in the control of rice sheath blight in Guyana. Crop Prot. 2019, 119, 30–37. [Google Scholar] [CrossRef]
- Prasad, B.; Eizenga, G.C. Rice sheath blight disease resistance identified in Oryza spp. accessions. Plant Dis. 2008, 92, 1503–1509. [Google Scholar] [CrossRef] [Green Version]
- Srinivasachary, L.W.; Savary, S. Resistance to rice sheath blight (Rhizoctonia solani Kühn) [(teleomorph: Thanatephorus cucumeris (A.B. Frank) Donk.] disease: Current status and perspectives. Euphytica 2011, 178, 1–22. [Google Scholar] [CrossRef]
- Zheng, L.; Liu, H.Q.; Zhang, M.L.; Cao, X.; Zhou, E.X. The comnplete genomic sequence of a novel mycovirus from Rhizoctonia solani AG-11Astrain B275. Arch. Virol. 2013, 158, 1609–1612. [Google Scholar] [CrossRef]
- Ou, S.H. A Hand Book of Rice in Tropics; The International Rice Research Institute (IRRI): Los Banos, Philippines, 1973; p. 53. [Google Scholar]
- Singh, P.; Mazumdar, P.; Harikrishna, J.A.; Babu, S. Sheath blight of rice: A review and identification of priorities for future research. Planta 2019, 250, 1387–1407. [Google Scholar] [CrossRef] [Green Version]
- Padaria, J.C.; Tarafdar, A.; Raipuria, R.; Lone, S.A.; Gahlot, P.; Shakil, N.A.; Kumar, J. Identification of phenazine-1-carboxylic acid gene (phc CD) from Bacillus pumilus MTCC7615 and its role in antagonism against Rhizoctonia solani. Basic Microbiol. 2016, 56, 999–1008. [Google Scholar] [CrossRef]
- Oloumi, H. Phytochemistry and ethno-pharmaceutics of Calotropis procera. EthnoPharmaceut. Prod. 2014, 1, 1–8. [Google Scholar]
- Yogi, B.; Gupta, S.K.; Mishra, A. Calotropis procera (Madar): A Medicinal plant of various therapeutic uses—A Review. Bull. Environ. Pharm. Life Sci. 2016, 5, 74–81. [Google Scholar]
- Yadav, V.K.; Thrimurty, V.S. Fungitoxicity of medicinal plant extracts against Sarocladium oryzae causing sheath rot in rice. Indian J. Plant Protect. 2006, 34, 263–264. [Google Scholar]
- Abu-Seif, F.A.; Abdel-Fattah, S.M.; Sreia, Y.H.A.; Ramadan, M.M. Antifungal properties of some medicinal plants against undesirable and mycotoxin-producing fungi. J. Agric. Sci. Mansoura Univ. 2009, 34, 1745–1756. [Google Scholar] [CrossRef]
- Khaledi, N.; Taheri, P.; Tarighi, S. Antifungal activity of various essential oils against Rhizoctonia solani and Macrophomina phaseolina as major bean pathogens. J. Appl. Microbiol. 2015, 118, 704–717. [Google Scholar] [CrossRef]
- Swain, T. Secondary compound as protective agents. Ann. Rev. Plant Physiol. 1977, 28, 479–501. [Google Scholar] [CrossRef]
- Roemer, T.; Xu, D.; Singh, S.B.; Parish, C.A.; Harris, G.; Wang, H.; Davies, J.E.; Bills, G.F. Confronting the challenges of natural product-based antifungal discovery. Chem. Biol. 2011, 18, 148–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Djakpoa, O.; Xie, Y.F.; Guo, Y.H.; Yu, H.; Cheng, Y.L.; Qian, H.; Shi, R.; Yao, W.R. Anti-quorum sensing of Galla chinensis and Coptis chinensis on bacteria. Food Sci. Tech. 2019, 101, 806–811. [Google Scholar] [CrossRef]
- Widiyarti, G.; Hanafi, M.; Soewarso, W.P. Study on the synthesis of monolaurin as antibacterial agent againts Staphylococcus aureus. Indo. J. Chem. 2009, 1, 99–106. [Google Scholar] [CrossRef]
- Nitbani, F.O.; Jumina; Siswanta, D.; Solikhah, E.N. Isolation and antibacterial activity test of lauric acid from crude coconut oil (Cocos nucifera L.). Procedia Chem. 2016, 18, 132–140. [Google Scholar] [CrossRef] [Green Version]
- Isaacs, C.E.; Litov, R.E.; Thormar, H. Antimicrobial activity of lipids added to human-milk, Infant formula, and bovine-milk. J. Nutr. Biochem. 1995, 6, 362–366. [Google Scholar] [CrossRef]
- Dayrit, F.M. The properties of lauric acid and their significance in coconut oil. J. Am. Oil Chem. Soc. 2015, 92, 1–15. [Google Scholar] [CrossRef]
- Desbois, A.P.; Smith, V.J. Antibacterial free fatty acids:activities mechanisms of action and biotechnological potential. Appl. Microbiol. Biot. 2010, 85, 1629–1642. [Google Scholar] [CrossRef] [Green Version]
- Walters, D.R.; Walker, R.L.; Walker, K.C. Lauric acid exhibits antifungal activity against plant pathogenic fungi. J. Phytopathol. 2003, 151, 228–230. [Google Scholar] [CrossRef]
- Richard, J.S.; Cesar, A.S.; Mónica, Á.M. Antifungal activity of LDPE/lauric acid films against Colletotrichum tamarilloi. Food Packag. Shelf 2020, 24, 100495. [Google Scholar] [CrossRef]
- Kabara, J.J.; Vrable, R. Antimicrobial lipids: Natural and synthetic fatty acids and monoglycerides. Chem. Mater. Sci. 1977, 12, 753–759. [Google Scholar] [CrossRef]
- Růžička, J.; Velclová, K.; Janiš, R.; Krejčí, J. Antimicrobial effects of 1-monoacylglycerols prepared by catalytic reaction of glycidol with fatty acids. Eur. Food Res. Technol. 2003, 217, 329–331. [Google Scholar] [CrossRef]
- Buňková, L.; Krejčí, J.; Janiš, R.; Kašpárková, V.; Vltavská, P.; Kulendová, L.; Buňka, F. Influence of monoacylglycerols on growth inhibition of micromycetes in vitro and on bread. Eur. J. Lipid Sci. Technol. 2010, 12, 173–179. [Google Scholar] [CrossRef]
- Wang, J.; Hu, X.F.; Yang, C.L.; Wu, X.M.; Li, R.Y.; Li, M. Growth restriction of Rhizoctonia solani via breakage of intracellular organelles using crude extracts of Gallnut and Clove. Moleclues 2021, 26, 1667. [Google Scholar] [CrossRef]
- Li, R.Y.; Wu, X.M.; Yin, X.H.; Long, Y.H.; Li, M. Naturally produced citral can signifificantly inhibit normal physiology and induce cytotoxicity on Magnaporthe grisea. Pestic. Biochem. Physiol. 2015, 118, 19–25. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.D.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [Green Version]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Gene ontology analysis for RNA-seq: Accounting for selection bias. Genome Biol. 2010, 11, R14. [Google Scholar] [CrossRef] [Green Version]
- Kanehisa, M.; Araki, M.; Goto, S.; Hattori, M.; Hirakawa, M.; Itoh, M.; Katayama, T.; Kawashima, S.; Okuda, S.; Tokimatsu, T.; et al. KEGG for linking genomes to life and the environment. Nucleic Acids Res. 2008, 36, D480–D484. [Google Scholar] [CrossRef]
- Mao, X.Z.; Cai, T.; Olyarchuk, J.G.; Wei, L.P. Automated genome annotation and pathway identification using the KEGG Orthology (KO) as a controlled vocabulary. Bioinformatics 2005, 21, 3787–3793. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.W.; Bao, J.Q.; Zhang, D.H.; Li, Y.; Li, H.S.; He, H.Z. Effect of heterocystous nitrogen-fixing cyanobacteria against rice sheath blight and the underlying mechanism. Appl. Soil Ecol. 2020, 153, 103580. [Google Scholar] [CrossRef]
- Peng, D.; Li, S.D.; Wang, J.X.; Chen, C.J.; Zhou, M.G. Integrated biological and chemical control of rice sheath blight by Bacillus subtilis NJ-18 and Jinggangmycin. Pest Manag. Sci. 2014, 70, 258–263. [Google Scholar] [CrossRef]
- Mosquera-Espinosa, A.T.; Bayman, P.; Prado, G.A.; Gómez-Carabalí, A.; Otero, J.T. The double life of Ceratobasidium: Orchid mycorrhizal fungi and their potential for bio-control of Rhizoctonia solani sheath blight of rice. Mycologia 2013, 105, 141–150. [Google Scholar] [CrossRef]
- Bhattacharyya, P.N.; Jha, D.K. Plant growth-promoting rhizobacteria (PGPR): Emergence in agriculture. World J. Microbiol. Biotechnol. 2012, 28, 1327–1350. [Google Scholar] [CrossRef] [PubMed]
- Nobel, D.H.; Ende, H.V.D.; Klis, F.M. Cell wall maintenance in fungi. Trend Microbiol. 2000, 8, 344–345. [Google Scholar] [CrossRef]
- Møller, I.M.; Jensen, P.E.; Hansson, A. Oxidative modificatlons to cellular components in plants. Annu. Rev. Plant Biol. 2007, 58, 459–481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahn, I.P.; Lee, S.W.; Suh, S.C. Rhizobacteria-induced priming in Arabidopsis is dependent on ethylene, jasmonic acid, and NPR1. Mol. Plant Microbe Interact. 2007, 20, 759–768. [Google Scholar] [CrossRef] [Green Version]
- Gaspar, Y.M.; Mckenna, J.A.; Mcginness, B.S.; Hinch, J.; Poon, S.; Connelly, A.A.; Anderson, M.A.; Heath, R.L. Field resistance to Fusarium oxysporum and Verticillium dahliae in transgenic cotton expressing the plant defensin NaD1. J. Exp. Bot. 2014, 65, 1541–1550. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhao, J.; Ding, L.Y.; Zou, L.F.; Li, Y.R.; Chen, G.Y.; Zhang, T.Z. Constitutive expression of a novel antimicrobial protein, Hcm1, confers resistance to both Verticillium and Fusarium wilts in cotton. Sci. Rep. 2016, 6, 20773. [Google Scholar] [CrossRef] [Green Version]
- Liu, N.; Sun, Y.; Pei, Y.K.; Zhang, X.Y.; Wang, P.; Li, X.C.; Li, F.G.; Hou, Y.X. A pectin methylesterase inhibitor enhances resistance to Verticillium wilt. Plant Physiol. 2018, 176, 2202–2220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Yu, X.R.; Zhang, C.J.; Zhang, Q.; Sun, Y.; Zhu, H.Q.; Tang, C.M. Pectin lyase enhances cotton resistance to Verticillium wilt by inducing cell apoptosis of Verticillium dahliae. J. Hazard. Mater. 2021, 404, 124029. [Google Scholar] [CrossRef] [PubMed]
- Chavan, S.; Bray, F.; Lortet-Tieulent, J.; Goodman, M.; Jemal, A. International variations in bladder cancer incidence and mortality. Eur. Urol. 2014, 66, 59–73. [Google Scholar] [CrossRef]
- Chauhan, A.K.; Jakhar, R.; Paul, S.; Kang, S.C. Potentiation of macrophage activity by thymol through augmenting phagocytosis. Int. Immunopharmacol. 2014, 18, 340–346. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, R.; Redman, R. Balancing the generation and elimination of reactive oxygen species. Proc. Natl. Acad. Sci. USA 2005, 102, 3175–3176. [Google Scholar] [CrossRef] [Green Version]
- Rehman, L.; Su, X.; Li, X.; Qi, X.; Guo, H.; Cheng, H. FreB is involved in the ferric metabolism and multiple pathogenicity-related traits of Verticillium dahliae. Curr. Genet. 2017, 64, 645–659. [Google Scholar] [CrossRef]
- Zorov, D.B.; Juhaszova, M.; Sollott, S.J. Mitochondrial ROS-induced ROS release: An update and review. Biochim. Biophys. Acta 2006, 1757, 509–517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.J.; Jiang, Z.Y.; Zhu, Q.; Zhong, G.H. Discovery of β-Carboline oxadiazole derivatives as fungicidal agents against rice sheath blight. J. Agric. Food. Chem. 2018, 22, 1–38. [Google Scholar] [CrossRef]
- Mukhopadhyay, K.; Prasad, T.; Saini, P.; Pucadyil, T.J.; Chattopadhyay, A.; Prasad, R. Membrane sphingolipid-ergosterol interactions are important determinants of ultidrug resistance in Candida albicans. Antimicrob. Agents Chemother. 2004, 48, 778–787. [Google Scholar] [CrossRef] [Green Version]
- Ghannoum, M.A.; Rice, L.B. Antifungal agents, mode of action, mechanisms of resistance, and correlation of these mechanisms with bacterial resistance. Clin. Microbiol. Rev. 1999, 12, 501–517. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Kaufman, R. Protein misfolding in the endoplasmic reticulum as a conduit to human disease. Nature 2016, 529, 326–335. [Google Scholar] [CrossRef] [PubMed]
- Ron, D.; Walter, P. Signal integration in the endoplasmic reticulum unfolded protein response. Nat. Rev. Mol. Cell Biol. 2007, 8, 519–529. [Google Scholar] [CrossRef] [PubMed]
- Hishiya, A.; Takayama, S. Molecular chaperones as regulators of cell death. Oncogene 2008, 27, 6489–6506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertolotti, A.; Zhang, Y.; Hendershot, L.M.; Harding, H.P.; Ron, D. Dynamic interaction of BiP and ER stress transducers in the unfolded-protein response. Nat. Cell Biol. 2000, 2, 326–332. [Google Scholar] [CrossRef]
- Hetz, C.; Saxena, S. ER stress and the unfolded protein response in neurodegeneration. Nat. Rev. Neurol. 2017, 13, 477–491. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lauric Acid Treatment (µg/mL) | 1d | 2d | 3d |
|---|---|---|---|
| 0 | ++ | +++ | ++++ |
| 50 | ++ | ++ | ++ |
| 100 | + | + | + |
| 150 | − | + | + |
| 200 | − | − | + |
| 250 | − | − | + |
| 300 | − | − | − |
| 350 | − | − | − |
| Samples | Total Reads | Clean Reads | Clean Bases | GC Content (%) | % ≥Q30 (%) | Total Mapped |
|---|---|---|---|---|---|---|
| CK1 | 64,784,226 | 32,392,113 | 9,678,905,954 | 53.27 | 94.59 | 58,992,337(91.06) |
| CK2 | 61,435,340 | 30,717,670 | 9,186,324,764 | 53.33 | 94.28 | 56,073,649(91.27) |
| CK3 | 64,967,078 | 32,483,539 | 9,712,848,232 | 53.25 | 94.7 | 59,354,850(91.36) |
| 3h-1 | 65,541,784 | 32,770,892 | 9,803,952,030 | 53.11 | 94.28 | 59,711,907(91.11) |
| 3h-2 | 69,943,128 | 34,971,564 | 10,457,950,356 | 53.12 | 94.42 | 63,761,674(91.16) |
| 3h-3 | 60,559,660 | 30,279,830 | 9,059,999,778 | 53.13 | 94.48 | 55,045,427(90.89) |
| 6h-1 | 62,595,230 | 31,297,615 | 9,357,641,672 | 53.23 | 94.37 | 56,786,050(90.72) |
| 6h-2 | 58,713,586 | 29,356,793 | 8,781,141,820 | 53.22 | 94.34 | 53,231,872(90.66) |
| 6h-3 | 63,140,638 | 31,570,319 | 9,437,986,814 | 53.21 | 94.05 | 57,316,763(90.78) |
| 18h-1 | 68,915,458 | 34,457,729 | 10,297,492,078 | 53.02 | 94.29 | 62,304,463(90.41) |
| 18h-2 | 72,425,996 | 36,212,998 | 10,835,409,898 | 53.14 | 94.41 | 65,386,591(90.28) |
| 18h-3 | 72,620,234 | 36,310,117 | 10,851,217,652 | 53.13 | 94.15 | 65,585,306(90.31) |
| 24h-1 | 67,231,244 | 33,615,622 | 10,054,713,440 | 53.11 | 94.07 | 60,647,299(90.21) |
| 24h-2 | 71,866,228 | 35,933,114 | 10,750,769,100 | 53.14 | 94.09 | 64,791,542(90.16) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Yang, C.; Hu, X.; Yao, X.; Han, L.; Wu, X.; Li, R.; Wen, T.; Ming, L. Lauric Acid Induces Apoptosis of Rice Sheath Blight Disease Caused by Rhizoctonia solani by Affecting Fungal Fatty Acid Metabolism and Destroying the Dynamic Equilibrium of Reactive Oxygen Species. J. Fungi 2022, 8, 153. https://doi.org/10.3390/jof8020153
Wang J, Yang C, Hu X, Yao X, Han L, Wu X, Li R, Wen T, Ming L. Lauric Acid Induces Apoptosis of Rice Sheath Blight Disease Caused by Rhizoctonia solani by Affecting Fungal Fatty Acid Metabolism and Destroying the Dynamic Equilibrium of Reactive Oxygen Species. Journal of Fungi. 2022; 8(2):153. https://doi.org/10.3390/jof8020153
Chicago/Turabian StyleWang, Jian, Chenglong Yang, Xianfeng Hu, Xiaolong Yao, Lei Han, Xiaomao Wu, Rongyu Li, Tingchi Wen, and Li Ming. 2022. "Lauric Acid Induces Apoptosis of Rice Sheath Blight Disease Caused by Rhizoctonia solani by Affecting Fungal Fatty Acid Metabolism and Destroying the Dynamic Equilibrium of Reactive Oxygen Species" Journal of Fungi 8, no. 2: 153. https://doi.org/10.3390/jof8020153
APA StyleWang, J., Yang, C., Hu, X., Yao, X., Han, L., Wu, X., Li, R., Wen, T., & Ming, L. (2022). Lauric Acid Induces Apoptosis of Rice Sheath Blight Disease Caused by Rhizoctonia solani by Affecting Fungal Fatty Acid Metabolism and Destroying the Dynamic Equilibrium of Reactive Oxygen Species. Journal of Fungi, 8(2), 153. https://doi.org/10.3390/jof8020153