
Effects of Arbuscular Mycorrhizal Fungi and Biochar on Growth, Nutrient Absorption, and Physiological Properties of Maize (Zea mays L.)
 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Experimental Setup
2.3. Harvest and Analysis
2.4. Data Analysis
3. Results
3.1. Mycorrhizal Colonization Rate
3.2. Soil Chemical Characteristics and Phosphatase Activity
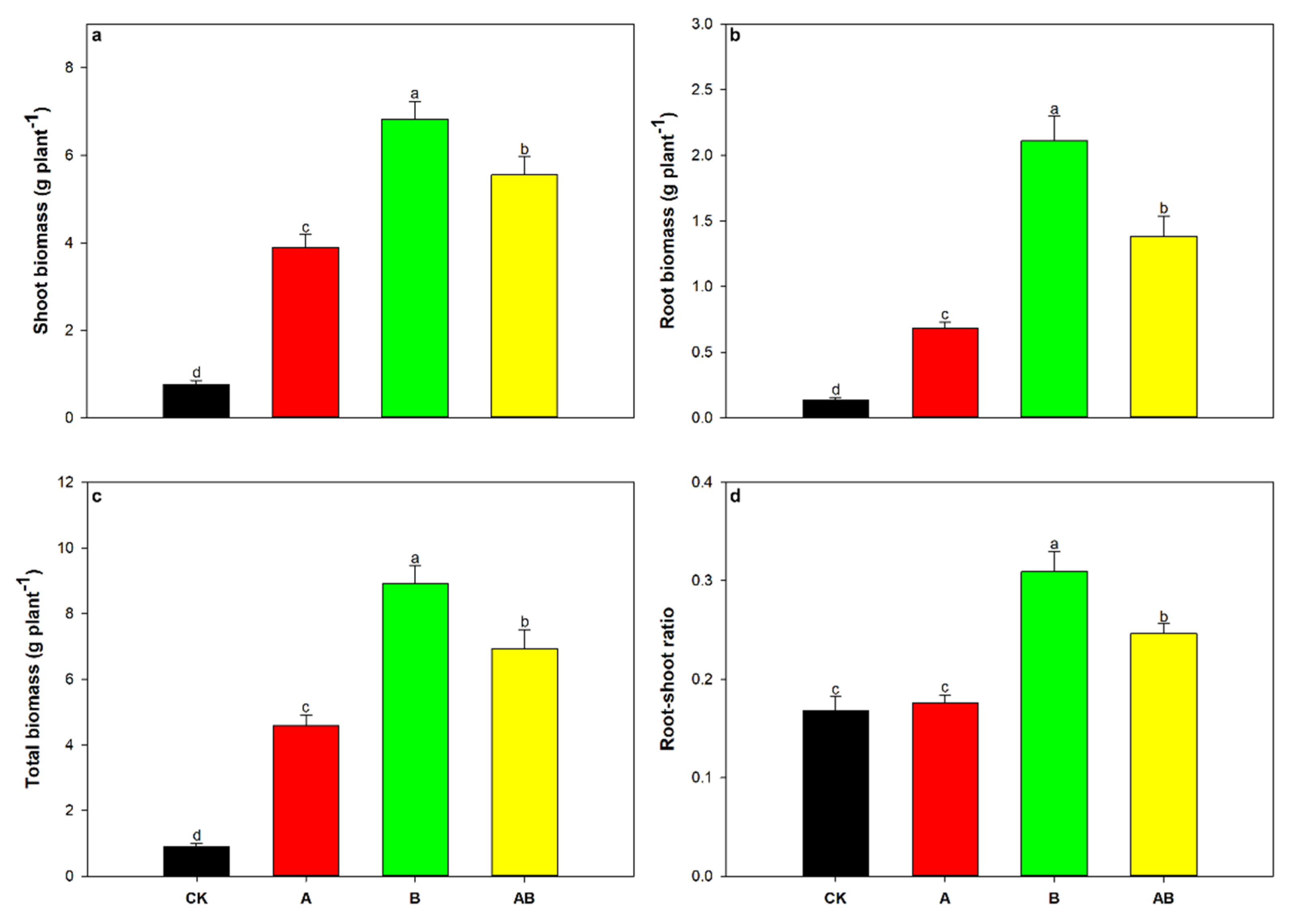
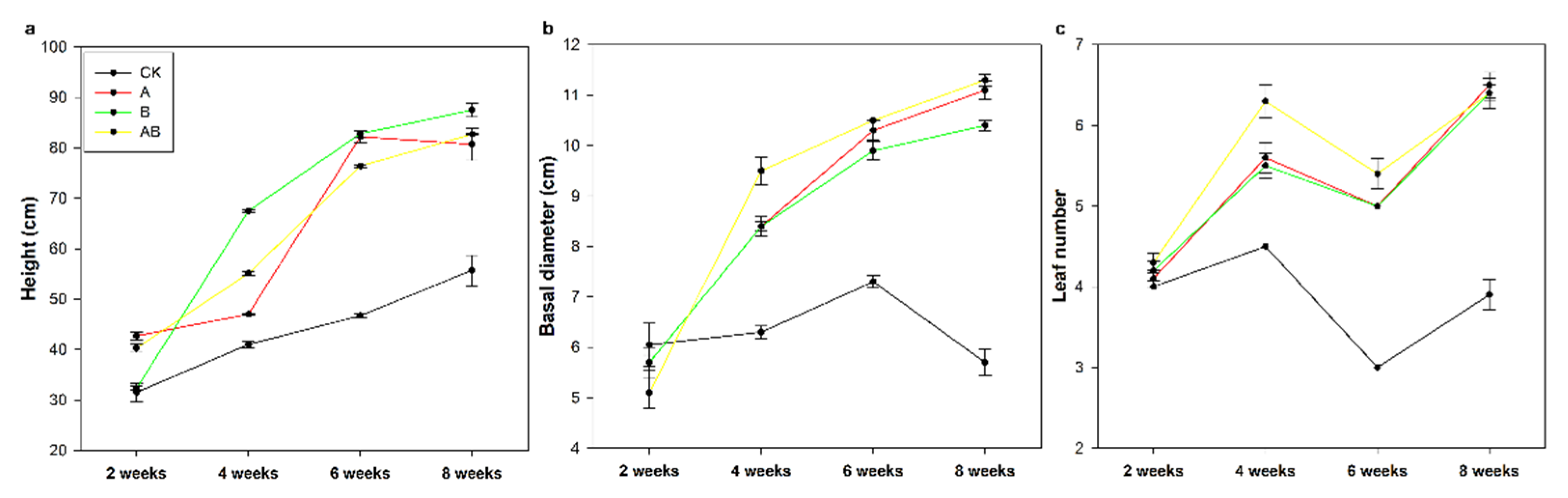
3.3. Maize Growth and Mycorrhizal Dependency
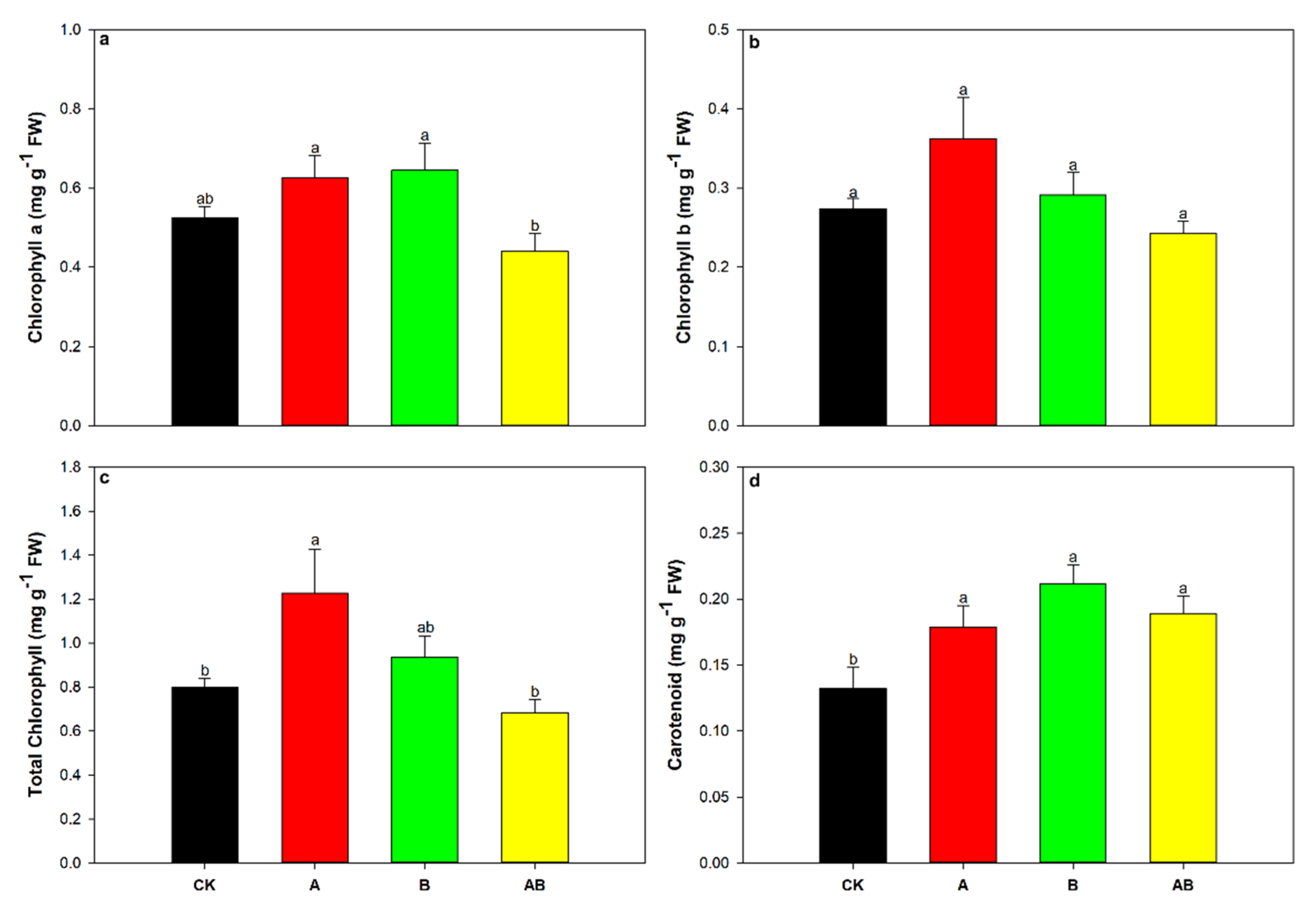
3.4. Photosynthetic Pigments
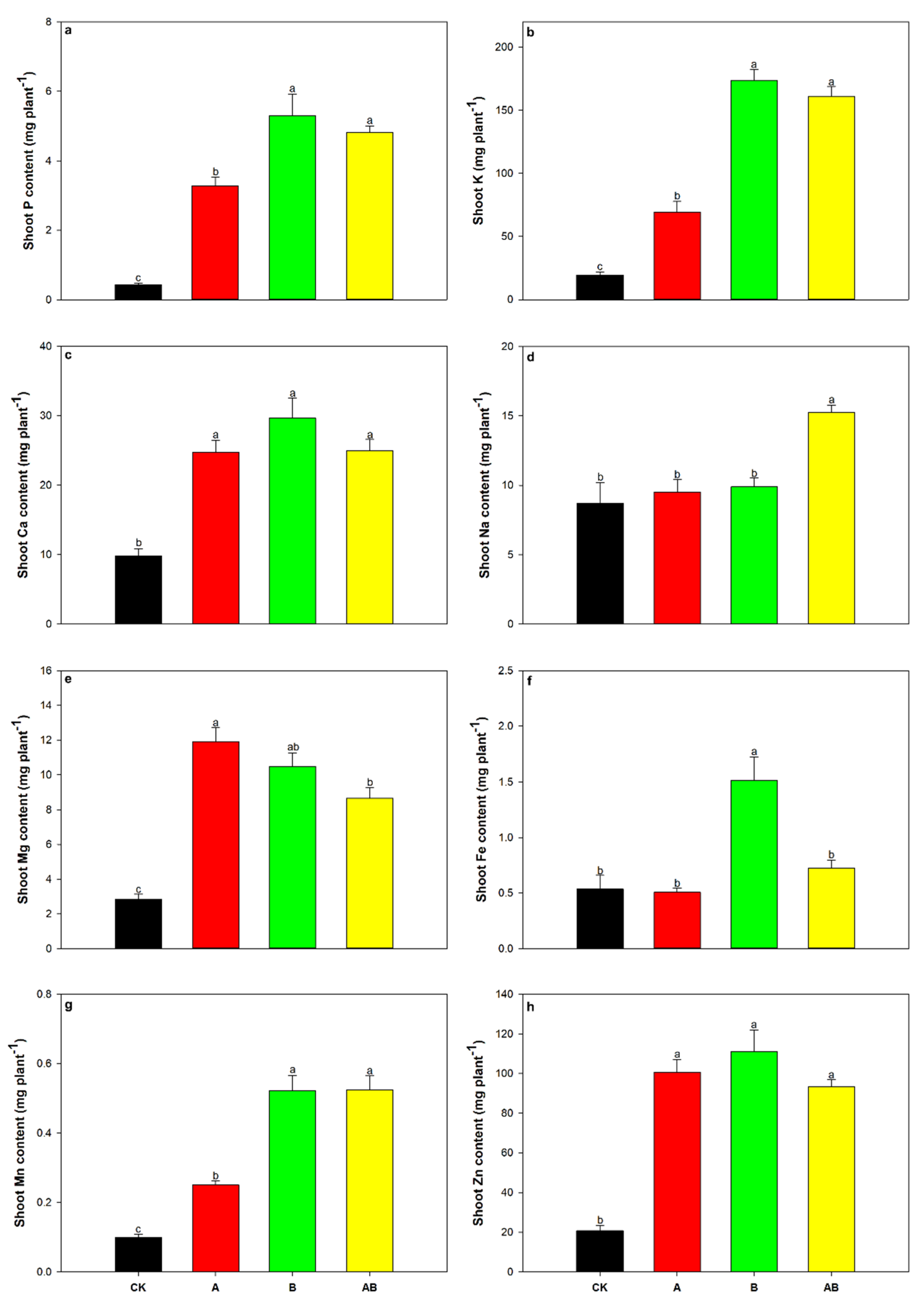
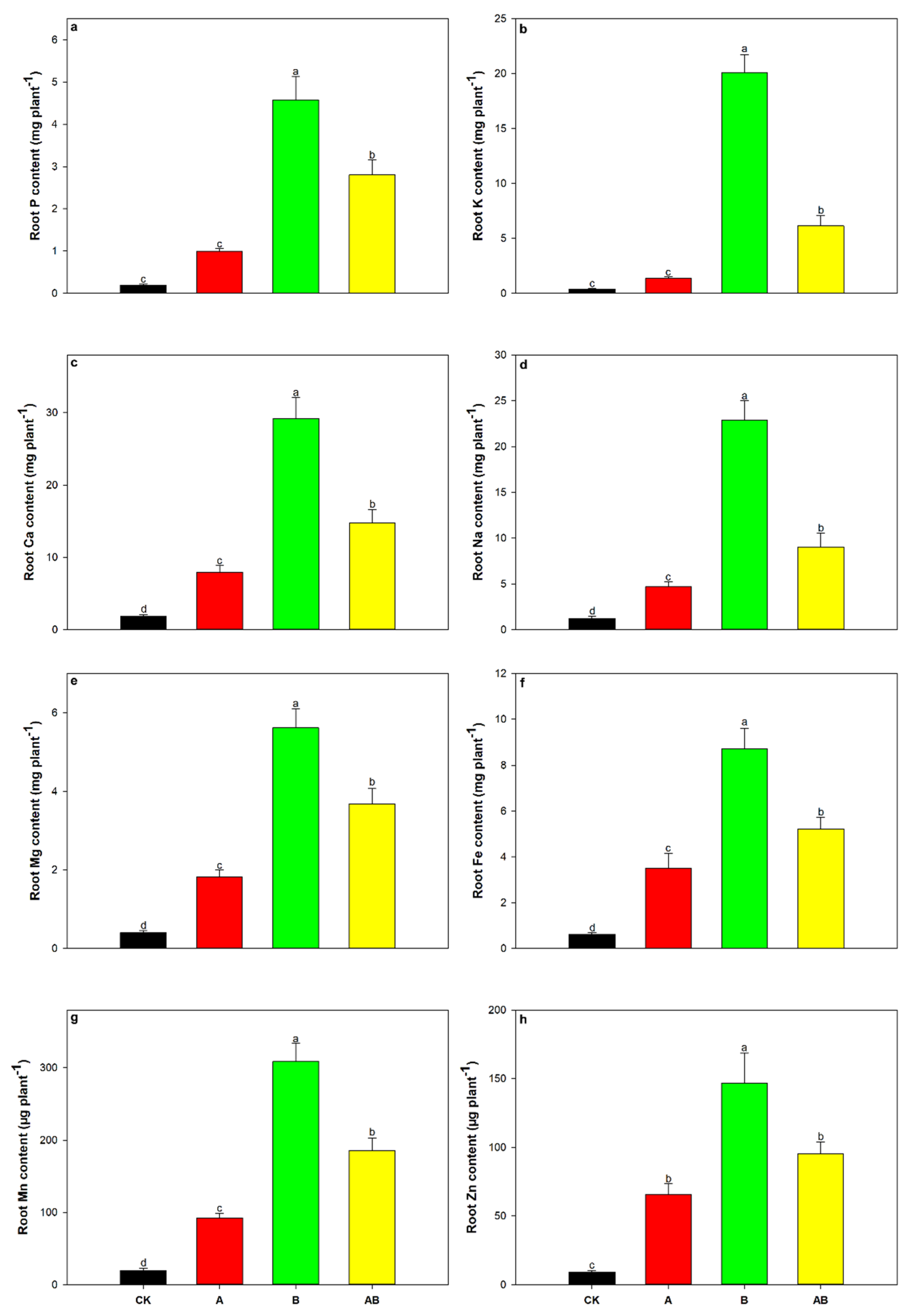
3.5. Mazie Nutrient Uptake
4. Discussion
4.1. Effects of AMFs
4.2. Effects of Biochar
4.3. Combined Effect of AMFs and Biochar
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lividini, K.; Masters, W.A. Tracing global flows of bioactive compounds from farm to fork in nutrient balance sheets can help guide intervention towards healthier food supplies. Nat. Food 2022, 3, 703–715. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.J.; Zhao, W.Q.; Li, T.; Cheng, X.Y.; Liu, Q. Balancing straw returning and chemical fertilizers in China: Role of straw nutrient resources. Renew. Sustain. Energy Rev. 2018, 81, 2695–2702. [Google Scholar] [CrossRef]
- Bai, Y.C.; Chang, Y.Y.; Hussain, M.; Lu, B.; Zhang, J.P.; Song, X.B.; Lei, X.S.; Pei, D. Soil chemical and microbiological properties are changed by long-term chemical fertilizers that limit ecosystem functioning. Microorganisms 2020, 8, 694. [Google Scholar] [CrossRef]
- Ostadi, A.; Javanmard, A.; Machiani, M.A.; Morshedloo, M.R.; Nouraein, M.; Rasouli, F.; Maggi, F. Effect of different fertilizer sources and harvesting time on the growth characteristics, nutrient uptakes, essential oil productivity and composition of Mentha x piperita L. Ind. Crops Prod. 2020, 148, 112290. [Google Scholar] [CrossRef]
- Wang, Y.F.; Chen, P.; Wang, F.H.; Han, W.X.; Qiao, M.; Dong, W.X.; Hu, C.S.; Zhu, D.; Chu, H.Y.; Zhu, Y.G. The ecological clusters of soil organisms drive the ecosystem multifunctionality under long-term fertilization. Environ. Int. 2022, 161, 107133. [Google Scholar] [CrossRef]
- Leake, J.R.; Johnson, D.; Donnelly, D.P.; Muckle, G.E.; Boddy, L.; Read, D.J. Networks of power and influence: The role of mycorrhizal mycelium in controlling plant communities and agroecosystem functioning. Can. J. Bot. 2004, 82, 1016–1045. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: Cambridge, UK, 2008; pp. 1–787. [Google Scholar]
- Foo, E.; Ross, J.J.; Jones, W.T.; Reid, J.B. Plant hormones in arbuscular mycorrhizal symbioses: An emerging role for gibberellins. Ann. Bot. 2013, 111, 769–779. [Google Scholar] [CrossRef]
- Goddard, M.L.; Belval, L.; Martin, I.R.; Roth, L.; Laloue, H.; Deglene-Benbrahim, L.; Valat, L.; Bertsch, C.; Chong, J.L. Arbuscular mycorrhizal symbiosis triggers major changes in primary metabolism together with modification of defense responses and signaling in both roots and leaves of Vitis vinifera. Front. Plant Sci. 2021, 12, 721614. [Google Scholar] [CrossRef] [PubMed]
- Sohi, S.P.; Krull, E.; Lopez-Capel, E.; Bol, R. A review of biochar and its use and function in soil. In Advances in Agronomy; Sparks, D.L., Ed.; Elsevier: Amsterdam, The Netherlands, 2010; Volume 105, pp. 47–82. [Google Scholar]
- Jeffery, S.; Verheijen, F.G.A.; van der Velde, M.; Bastos, A.C. A quantitative review of the effects of biochar application to soils on crop productivity using meta-analysis. Agric. Ecosyst. Environ. 2011, 144, 175–187. [Google Scholar] [CrossRef]
- Haider, F.U.; Coulter, J.A.; Cai, L.Q.; Hussain, S.; Cheema, S.A.; Wu, J.; Zhang, R.Z. An overview on biochar production, its implications, and mechanisms of biochar-induced amelioration of soil and plant characteristics. Pedosphere 2022, 32, 107–130. [Google Scholar] [CrossRef]
- Razzaghi, F.; Obour, P.B.; Arthur, E. Does biochar improve soil water retention? A systematic review and meta-analysis. Geoderma 2020, 361, 114055. [Google Scholar] [CrossRef]
- Lehmann, J.; Rillig, M.C.; Thies, J.; Masiello, C.A.; Hockaday, W.C.; Crowley, D. Biochar effects on soil biota—A review. Soil Biol. Biochem. 2011, 43, 1812–1836. [Google Scholar] [CrossRef]
- Schmidt, M.W.I.; Torn, M.S.; Abiven, S.; Dittmar, T.; Guggenberger, G.; Janssens, I.A.; Kleber, M.; Kogel-Knabner, I.; Lehmann, J.; Manning, D.A.C.; et al. Persistence of soil organic matter as an ecosystem property. Nature 2011, 478, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Vanek, S.J.; Lehmann, J. Phosphorus availability to beans via interactions between mycorrhizas and biochar. Plant Soil 2015, 395, 105–123. [Google Scholar] [CrossRef]
- Mickan, B.S.; Abbott, L.K.; Stefanova, K.; Solaiman, Z.M. Interactions between biochar and mycorrhizal fungi in a water-stressed agricultural soil. Mycorrhiza 2016, 26, 565–574. [Google Scholar] [CrossRef]
- Warnock, D.D.; Lehmann, J.; Kuyper, T.W.; Rillig, M.C. Mycorrhizal responses to biochar in soil—Concepts and mechanisms. Plant Soil 2007, 300, 9–20. [Google Scholar] [CrossRef]
- Yang, W.; Zhao, Y.; Yang, Y.; Zhang, M.; Mao, X.; Guo, Y.; Li, X.; Tao, B.; Qi, Y.; Ma, L.; et al. Co-application of biochar and microbial inoculants increases soil phosphorus and potassium fertility and improves soil health and tomato growth. J. Soils Sediments 2022. [Google Scholar] [CrossRef]
- Jabborova, D.; Annapurna, K.; Azimov, A.; Tyagi, S.; Pengani, K.R.; Sharma, P.; Vikram, K.V.; Poczai, P.; Nasif, O.; Ansari, M.J.; et al. Co-inoculation of biochar and arbuscular mycorrhizae for growth promotion and nutrient fortification in soybean under drought conditions. Front. Plant Sci. 2022, 13, 947547. [Google Scholar] [CrossRef]
- Costa, R.; Calvete, E.O.; Spengler, N.; Chiomento, J.; Paula, J.J.A.S.A. Morpho-phenological and agronomic performance of strawberry cultivars with different photoperiodic flowering responses. Sci. Hortic. 2021, 43, e45189. [Google Scholar]
- Jabborova, D.; Annapurna, K.; Choudhary, R.; Bhowmik, S.N.; Desouky, S.E.; Selim, S.; Azab, I.H.E.; Hamada, M.M.A.; Nahhas, N.E.; Elkelish, A. Interactive impact of biochar and arbuscular mycorrhizal on root morphology, physiological properties of fenugreek (Trigonella foenum-graecum L.) and soil enzymatic activities. Agronomy 2021, 11, 2341. [Google Scholar] [CrossRef]
- Cairns, J.E.; Prasanna, B.M. Developing and deploying climate-resilient maize varieties in the developing world. Curr. Opin. Plant Biol. 2018, 45, 226–230. [Google Scholar] [CrossRef]
- McGonigle, T.P.; Miller, M.H.; Evans, D.G.; Fairchild, G.L.; Swan, J.A. A new method which gives an objective-measure of colonization of roots by vesicular arbuscular mycorrhizal fungi. New Phytol. 1990, 115, 495–501. [Google Scholar] [CrossRef] [PubMed]
- Lambers, H.; Raven, J.A.; Shaver, G.R.; Smith, S.E. Plant nutrient-acquisition strategies change with soil age. Trends Ecol. Evol. 2008, 23, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Bahram, M.; Netherway, T.; Hildebrand, F.; Pritsch, K.; Drenkhan, R.; Loit, K.; Anslan, S.; Bork, P.; Tedersoo, L. Plant nutrient-acquisition strategies drive topsoil microbiome structure and function. New Phytol. 2020, 227, 1189–1199. [Google Scholar] [CrossRef] [PubMed]
- Hodge, A.; Campbell, C.D.; Fitter, A.H. An arbuscular mycorrhizal fungus accelerates decomposition and acquires nitrogen directly from organic material. Nature 2001, 413, 297–299. [Google Scholar] [CrossRef]
- Etesami, H.; Jeong, B.R.; Glick, B.R. Contribution of arbuscular mycorrhizal fungi, phosphate-solubilizing bacteria, and silicon to P uptake by plant. Front. Plant Sci. 2021, 12, 699618. [Google Scholar] [CrossRef]
- Hodge, A.; Storer, K. Arbuscular mycorrhiza and nitrogen: Implications for individual plants through to ecosystems. Plant Soil 2015, 386, 1–19. [Google Scholar] [CrossRef]
- Hodge, A.; Fitter, A.H. Substantial nitrogen acquisition by arbuscular mycorrhizal fungi from organic material has implications for N cycling. Proc. Natl. Acad. Sci. USA 2010, 107, 13754–13759. [Google Scholar] [CrossRef] [PubMed]
- Cavagnaro, T.R.; Barrios-Masias, F.H.; Jackson, L.E. Arbuscular mycorrhizas and their role in plant growth, nitrogen interception and soil gas efflux in an organic production system. Plant Soil 2012, 353, 181–194. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, G.; Luo, X.; Hou, E.; Zheng, M.; Zhang, L.; He, X.; Shen, W.; Wen, D. Mycorrhizal fungi and phosphatase involvement in rhizosphere phosphorus transformations improves plant nutrition during subtropical forest succession. Soil Biol. Biochem. 2021, 153, 108099. [Google Scholar] [CrossRef]
- Dashwood, R.H. Chlorophylls as anticarcinogens (review). Int. J. Oncol. 1997, 10, 721–727. [Google Scholar] [CrossRef] [PubMed]
- Ghani, M.I.; Ali, A.; Atif, M.J.; Ali, M.; Amin, B.; Cheng, Z. Arbuscular mycorrhizal fungi and dry raw garlic stalk amendment alleviate continuous monocropping growth and photosynthetic declines in eggplant by bolstering its antioxidant system and accumulation of osmolytes and secondary metabolites. Front. Plant Sci. 2022, 13, 849521. [Google Scholar] [CrossRef] [PubMed]
- Malik, J.A.; AlQarawi, A.A.; Dar, B.A.; Hashem, A.; Alshahrani, T.S.; AlZain, M.N.; Habib, M.M.; Javed, M.M.; Abd-Allah, E.F. Arbuscular mycorrhizal fungi isolated from highly saline “sabkha habitat” soil alleviated the NaCl-induced stress and improved Lasiurus scindicus Henr. growth. Agriculture 2022, 12, 337. [Google Scholar] [CrossRef]
- Cortleven, A.; Schmulling, T. Regulation of chloroplast development and function by cytokinin. J. Exp. Bot. 2015, 66, 4999–5013. [Google Scholar] [CrossRef]
- Liang, J.F.; An, J.; Gao, J.Q.; Zhang, X.Y.; Song, M.H.; Yu, F.H. Interactive effects of biochar and AMF on plant growth and greenhouse gas emissions from wetland microcosms. Geoderma 2019, 346, 11–17. [Google Scholar] [CrossRef]
- Efthymiou, A.; Jensen, B.; Jakobsen, I. The roles of mycorrhiza and Penicillium inoculants in phosphorus uptake by biochar-amended wheat. Soil Biol. Biochem. 2018, 127, 168–177. [Google Scholar] [CrossRef]
- Hossain, M.Z.; Bahar, M.M.; Sarkar, B.; Donne, S.W.; Ok, Y.S.; Palansooriya, K.N.; Kirkham, M.B.; Chowdhury, S.; Bolan, N. Biochar and its importance on nutrient dynamics in soil and plant. Biochar 2020, 2, 379–420. [Google Scholar] [CrossRef]
- Laird, D.; Fleming, P.; Wang, B.; Horton, R.; Karlen, D. Biochar impact on nutrient leaching from a Midwestern agricultural soil. Geoderma 2010, 158, 436–442. [Google Scholar] [CrossRef]
- Shakeel, H.; Jahan, S.; Rafiq, K.; Iqbal, S.; Rasul, F. Efficacy of biochar-supplemented soil for modification of physio-biochemical attributes of Canola (Brassica napus L.) genotypes under different moisture regimes. J. Soil Sci. Plant Nutr. 2022, 22, 3667–3684. [Google Scholar] [CrossRef]
- Guo, L.; Yu, H.; Kharbach, M.; Wang, J. The response of nutrient uptake, photosynthesis and yield of tomato to biochar addition under reduced nitrogen application. Agronomy 2021, 11, 1598. [Google Scholar] [CrossRef]
- Rehman, M.Z.u.; Rizwan, M.; Khalid, H.; Ali, S.; Naeem, A.; Yousaf, B.; Liu, G.; Sabir, M.; Farooq, M. Farmyard manure alone and combined with immobilizing amendments reduced cadmium accumulation in wheat and rice grains grown in field irrigated with raw effluents. Chemosphere 2018, 199, 468–476. [Google Scholar] [CrossRef] [PubMed]
- He, Y.H.; Yao, Y.X.; Ji, Y.H.; Deng, J.; Zhou, G.Y.; Liu, R.Q.; Shao, J.J.; Zhou, L.Y.; Li, N.; Zhou, X.H.; et al. Biochar amendment boosts photosynthesis and biomass in C3 but not C4 plants: A global synthesis. Glob. Chang. Biol. Bioenergy 2020, 12, 605–617. [Google Scholar] [CrossRef]
- Rafique, M.; Ortas, I.; Rizwan, M.; Chaudhary, H.J.; Gurmani, A.R.; Munis, M.F.H. Residual effects of biochar and phosphorus on growth and nutrient accumulation by maize (Zea mays L.) amended with microbes in texturally different soils. Chemosphere 2020, 238, 124710. [Google Scholar] [CrossRef]
- Yang, Q.; Ravnskov, S.; Andersen, M.N. Nutrient uptake and growth of potato: Arbuscular mycorrhiza symbiosis interacts with quality and quantity of amended biochars. J. Soil Sci. Plant Nutr. 2020, 183, 220–232. [Google Scholar] [CrossRef]
- Yang, X.; Ran, Z.; Li, R.; Fang, L.; Zhou, J.; Guo, L. Effects of biochar on the growth, ginsenoside content, and soil microbial community composition of Panax quinquefolium L. J. Soil Sci. Plant Nutr. 2022, 22, 2670–2686. [Google Scholar] [CrossRef]
- Abou El Seoud, I.I.A. Effect of biochar rates on A-mycorrhizal fungi performance and maize plant growth, Phosphorus uptake, and soil P availability under calcareous soil conditions. Commun. Soil Sci. Plant Anal. 2021, 52, 815–831. [Google Scholar]
- Jabborova, D.; Annapurna, K.; Paul, S.; Kumar, S.; Saad, H.A.; Desouky, S.; Ibrahim, M.F.M.; Elkelish, A. Beneficial features of biochar and arbuscular mycorrhiza for improving spinach plant growth, root morphological traits, physiological properties, and soil enzymatic activities. J. Fungi 2021, 7, 571. [Google Scholar] [CrossRef]
- Abbott, L.K.; Robson, A.D. The effect of root density, inoculum placement and infectivity of inoculum on the development of vesicular arbuscular mycorrhizas. New Phytol. 1984, 97, 285–299. [Google Scholar] [CrossRef]
- Cavagnaro, R.A.; Oyarzabal, M.; Oesterheld, M.; Grimoldi, A.A. Species-specific trade-offs between regrowth and mycorrhizas in the face of defoliation and phosphorus addition. Fungal Ecol. 2021, 51, 101058. [Google Scholar] [CrossRef]
- Johnson, N.C.; Graham, J.H.; Smith, F.A. Functioning of mycorrhizal associations along the mutualism-parasitism continuum. New Phytol. 1997, 135, 575–586. [Google Scholar] [CrossRef]
- LeCroy, C.; Masiello, C.A.; Rudgers, J.A.; Hockaday, W.C.; Silberg, J.J. Nitrogen, biochar, and mycorrhizae: Alteration of the symbiosis and oxidation of the char surface. Soil Biol. Biochem. 2013, 58, 248–254. [Google Scholar] [CrossRef]
- Javeed, H.M.R.; Naeem, R.; Ali, M.; Qamar, R.; Sarwar, M.A.; Nawaz, F.; Atiqueur, R.; Shehzad, M.; Farooq, A.; Haseebur, R.; et al. Coupling biochar with microbial inoculants improves maize growth and nutrients acquisition under phosphorous-limited soil. Acta Physiol. Plant. 2022, 44, 110. [Google Scholar] [CrossRef]
- Ndiate, N.I.; Saeed, Q.; Haider, F.U.; Liqun, C.; Nkoh, J.N.; Mustafa, A. Co-application of biochar and arbuscular mycorrhizal fungi improves salinity tolerance, growth and lipid metabolism of maize (Zea mays L.) in an alkaline soil. Plants 2021, 10, 2490. [Google Scholar] [CrossRef]
- Li, M.Y.; Cai, L.Q. Biochar and arbuscular mycorrhizal fungi play different roles in enabling maize to uptake phosphorus. Sustainability 2021, 13, 3244. [Google Scholar] [CrossRef]
- Han, Y.; Douds, D.D., Jr.; Boateng, A.A. Effect of biochar soil-amendments on Allium porrum growth and arbuscular mycorrhizal fungus colonization. J. Plant Nutr. 2016, 39, 1654–1662. [Google Scholar] [CrossRef]
- Koide, R.T. Biochar-arbuscular mycorrhiza interaction in temperate soils. In Mycorrhizal Mediation of Soil; Johnson, N.C., Gehring, C., Jansa, J., Eds.; Elsevier: Berlin, Germany, 2017; pp. 461–477. [Google Scholar]
- Zhou, C.; Heal, K.; Tigabu, M.; Xia, L.; Hu, H.; Yin, D.; Ma, X. Biochar addition to forest plantation soil enhances phosphorus availability and soil bacterial community diversity. Forest. Ecol. Manag. 2020, 455, 117635. [Google Scholar] [CrossRef]
- Bashir, M.A.; Wang, X.K.; Naveed, M.; Mustafa, A.; Ashraf, S.; Samreen, T.; Nadeem, S.M.; Jamil, M. Biochar mediated-alleviation of chromium stress and growth improvement of different maize cultivars in tannery polluted soils. Int. J. Environ. Res. Public Health 2021, 18, 4461. [Google Scholar] [CrossRef]
- Wright, D.P.; Scholes, J.D.; Read, D.J.; Rolfe, S.A. European and African maize cultivars differ in their physiological and molecular responses to mycorrhizal infection. New Phytol. 2005, 167, 881–896. [Google Scholar] [CrossRef]
- Merlos, M.A.; Zitka, O.; Vojtech, A.; Azcon-Aguilar, C.; Ferrol, N. The arbuscular mycorrhizal fungus Rhizophagus irregularis differentially regulates the copper response of two maize cultivars differing in copper tolerance. Plant Sci. 2016, 253, 68–76. [Google Scholar] [CrossRef] [PubMed]
- Amery, F.; Debode, J.; Ommeslag, S.; Visser, R.; De Tender, C.; Vandecasteele, B. Biochar for circular horticulture: Feedstock related effects in soilless cultivation. Agronomy 2021, 11, 629. [Google Scholar] [CrossRef]
- dos Santos Trentin, T.; Dornelles, A.G.; dos Santos Trentin, N.; Huzar-Novakowiski, J.; Calvete, E.O.; Trevizan Chiomento, J.L. Addition of arbuscular mycorrhizal fungi and biochar in the cultivation substrate benefits macronutrient contents in strawberry plants. J. Soil Sci. Plant Nutr. 2022, 22, 2980–2991. [Google Scholar] [CrossRef]
- Warnock, D.D.; Mummey, D.L.; McBride, B.; Major, J.; Lehmann, J.; Rillig, M.C. Influences of non-herbaceous biochar on arbuscular mycorrhizal fungal abundances in roots and soils: Results from growth-chamber and field experiments. Appl. Soil Ecol. 2010, 46, 450–456. [Google Scholar] [CrossRef]
- Liu, M.; Zhao, Z.; Chen, L.; Wang, L.; Ji, L.; Xiao, Y. Influences of arbuscular mycorrhizae, phosphorus fertiliser and biochar on alfalfa growth, nutrient status and cadmium uptake. Ecotoxicol. Environ. Saf. 2020, 196, 110537. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Liu, F.; Ravnskov, S.; Rubæk, G.H.; Sun, Z.; Andersen, M.N. Impact of wood biochar and its interactions with mycorrhizal fungi, phosphorus fertilization and irrigation strategies on potato growth. J. Agron. Crop Sci. 2017, 203, 131–145. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Mycorrhizal Colonization Rate |
|---|---|
| A | 52.77 ± 4.74 a |
| AB | 31.52 ± 2.47 b |
| Variable Measured | A | B | A × B | |||
|---|---|---|---|---|---|---|
| F | P | F | P | F | P | |
| pH | 0.14 | 0.71 | 0.76 | 0.40 | 0.69 | 0.42 |
| NH4+-N | 2.58 | 0.13 | 12.40 | ** | 18.11 | *** |
| NO3−-N | 159.78 | *** | 200.84 | *** | 135.80 | *** |
| A-P | 6.24 | * | 108.27 | *** | 5.47 | * |
| DOC | 8.48 | * | 5.08 | * | 0.54 | 0.47 |
| MBC | 19.66 | *** | 22.15 | *** | 35.43 | *** |
| MBN | 0.22 | 0.65 | 11.31 | ** | 1.55 | 0.23 |
| Alkaline phosphatase | 0.13 | 0.72 | 1.07 | 0.32 | 5.00 | * |
| Shoot biomass | 32.80 | * | 205.00 | *** | 84.86 | *** |
| Root biomass | 3.33 | 0.09 | 194.54 | *** | 52.22 | *** |
| Total biomass | 23.66 | *** | 216.35 | *** | 81.37 | *** |
| Root–shoot ratio | 3.72 | 0.07 | 58.41 | *** | 6.25 | * |
| Chlorophyll a | 0.97 | 0.34 | 0.46 | 0.51 | 8.70 | ** |
| Chlorophyll b | 0.45 | 0.51 | 2.64 | 0.12 | 4.83 | * |
| Total chlorophyll | 0.58 | 0.46 | 3.08 | 0.10 | 8.51 | ** |
| Carotenoid | 0.88 | 0.36 | 9.47 | ** | 5.80 | * |
| Shoot P content | 11.26 | ** | 82.08 | *** | 22.29 | *** |
| Shoot K content | 23.14 | *** | 343.79 | *** | 37.92 | *** |
| Shoot Ca content | 13.18 | * | 35.70 | *** | 34.25 | *** |
| Shoot Na content | 10.14 | ** | 12.77 | ** | 5.51 | * |
| Shoot Mg content | 49.56 | *** | 25.01 | *** | 96.83 | *** |
| Shoot Fe content | 9.07 | ** | 25.71 | *** | 8.69 | ** |
| Shoot Mn content | 16.84 | *** | 193.23 | *** | 16.84 | *** |
| Shoot Zn content | 22.26 | *** | 39.69 | *** | 54.48 | *** |
| Root P content | 2.12 | 0.17 | 85.91 | *** | 14.89 | ** |
| Root K content | 48.00 | *** | 170.26 | *** | 63.18 | *** |
| Root Ca content | 5.32 | * | 89.56 | *** | 32.50 | *** |
| Root Na content | 3.46 | 0.08 | 137.64 | *** | 56.45 | *** |
| Root Mg content | 2.59 | 0.13 | 187.01 | *** | 48.65 | *** |
| Root Fe content | 2.34 | 0.15 | 96.60 | *** | 42.60 | *** |
| Root Mn content | 1.28 | 0.28 | 242.22 | *** | 69.45 | *** |
| Root Zn content | 0.05 | 0.83 | 45.92 | *** | 19.15 | *** |
| Treatment | pH | NH4+-N (mg kg−1) | NO3−-N (mg kg−1) | A-P (mg kg−1) | DOC (mg kg−1) | MBC (mg kg−1) | MBN (mg kg−1) | Alkaline Phosphatase (nmol·g−1·d−1) |
|---|---|---|---|---|---|---|---|---|
| CK | 8.39 ± 0.06 a | 5.51 ± 0.49 a | 23.93 ± 1.3 a | 7.12 ± 0.69 c | 71.41 ± 6.84 b | 299.01 ± 18.18 a | 18.6 ± 1.76 a | 341.23 ± 59.56 b |
| A | 8.49 ± 0.04 a | 3.59 ± 0.35 b | 4.48 ± 0.75 b | 6.14 ± 0.24 c | 86.44 ± 9.04 ab | 147.43 ± 12.9 c | 17.56 ± 1.09 a | 2022.99 ± 253.72 a |
| B | 8.37 ± 0.18 a | 2.97 ± 0.04 b | 3.25 ± 0.37 b | 85.88 ± 12.21 a | 81.9 ± 3.29 b | 143.45 ± 11.88 c | 12.4 ± 1.42 b | 2052.78 ± 792.99 a |
| AB | 8.32 ± 0.05 a | 3.83 ± 0.26 b | 2.46 ± 0.41 b | 55.99 ± 1.74 b | 107.06 ± 7.16 a | 165.61 ± 14.63 c | 14.7 ± 0.96 ab | 1162.71 ± 310.19 ab |
| Treatment | Mycorrhizal Dependency |
|---|---|
| Without biochar | 80.57 ± 1.63a |
| With biochar | −28.67 ± 3.93b |
| Variable Measured | A | B | T | A × B | A × T | B × T | A × B × T | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| F | P | F | P | F | P | F | P | F | P | F | P | F | P | |
| Maize height | 135.17 | *** | 330.47 | *** | 761.03 | *** | 305.11 | *** | 32.73 | *** | 42.21 | *** | 38.15 | *** |
| Basal diameter | 202.22 | *** | 135.62 | *** | 300.61 | *** | 90.99 | *** | 49.11 | *** | 33.32 | *** | 19.12 | *** |
| Leaf number | 180.04 | *** | 170.04 | *** | 133.18 | *** | 72.32 | *** | 16.99 | *** | 12.70 | *** | 20.70 | *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, J.; Jia, Q.; Li, Y.; Zhang, T.; Chen, J.; Ren, Y.; Dong, K.; Xu, S.; Shi, N.-N.; Fu, S. Effects of Arbuscular Mycorrhizal Fungi and Biochar on Growth, Nutrient Absorption, and Physiological Properties of Maize (Zea mays L.). J. Fungi 2022, 8, 1275. https://doi.org/10.3390/jof8121275
Sun J, Jia Q, Li Y, Zhang T, Chen J, Ren Y, Dong K, Xu S, Shi N-N, Fu S. Effects of Arbuscular Mycorrhizal Fungi and Biochar on Growth, Nutrient Absorption, and Physiological Properties of Maize (Zea mays L.). Journal of Fungi. 2022; 8(12):1275. https://doi.org/10.3390/jof8121275
Chicago/Turabian StyleSun, Jiahua, Qiong Jia, Yi Li, Ting Zhang, Jiayuan Chen, Yanan Ren, Kanglong Dong, Shuai Xu, Nan-Nan Shi, and Shenglei Fu. 2022. "Effects of Arbuscular Mycorrhizal Fungi and Biochar on Growth, Nutrient Absorption, and Physiological Properties of Maize (Zea mays L.)" Journal of Fungi 8, no. 12: 1275. https://doi.org/10.3390/jof8121275
APA StyleSun, J., Jia, Q., Li, Y., Zhang, T., Chen, J., Ren, Y., Dong, K., Xu, S., Shi, N.-N., & Fu, S. (2022). Effects of Arbuscular Mycorrhizal Fungi and Biochar on Growth, Nutrient Absorption, and Physiological Properties of Maize (Zea mays L.). Journal of Fungi, 8(12), 1275. https://doi.org/10.3390/jof8121275