

FgLEU1 Is Involved in Leucine Biosynthesis, Sexual Reproduction, and Full Virulence in Fusarium graminearum


Abstract
1. Introduction
2. Materials and Methods
2.1. Fungal Strains and Culture Conditions
2.2. Generation of FgLEU1 Deletion Mutants
2.3. Gene Complementation
2.4. Asexual and Sexual Reproduction Assays
2.5. RNA Extraction and Quantitative Real-Time PCR (qRT-PCR)
2.6. Plant Infection and DON Production Assays
2.7. Statistical Analysis
3. Results
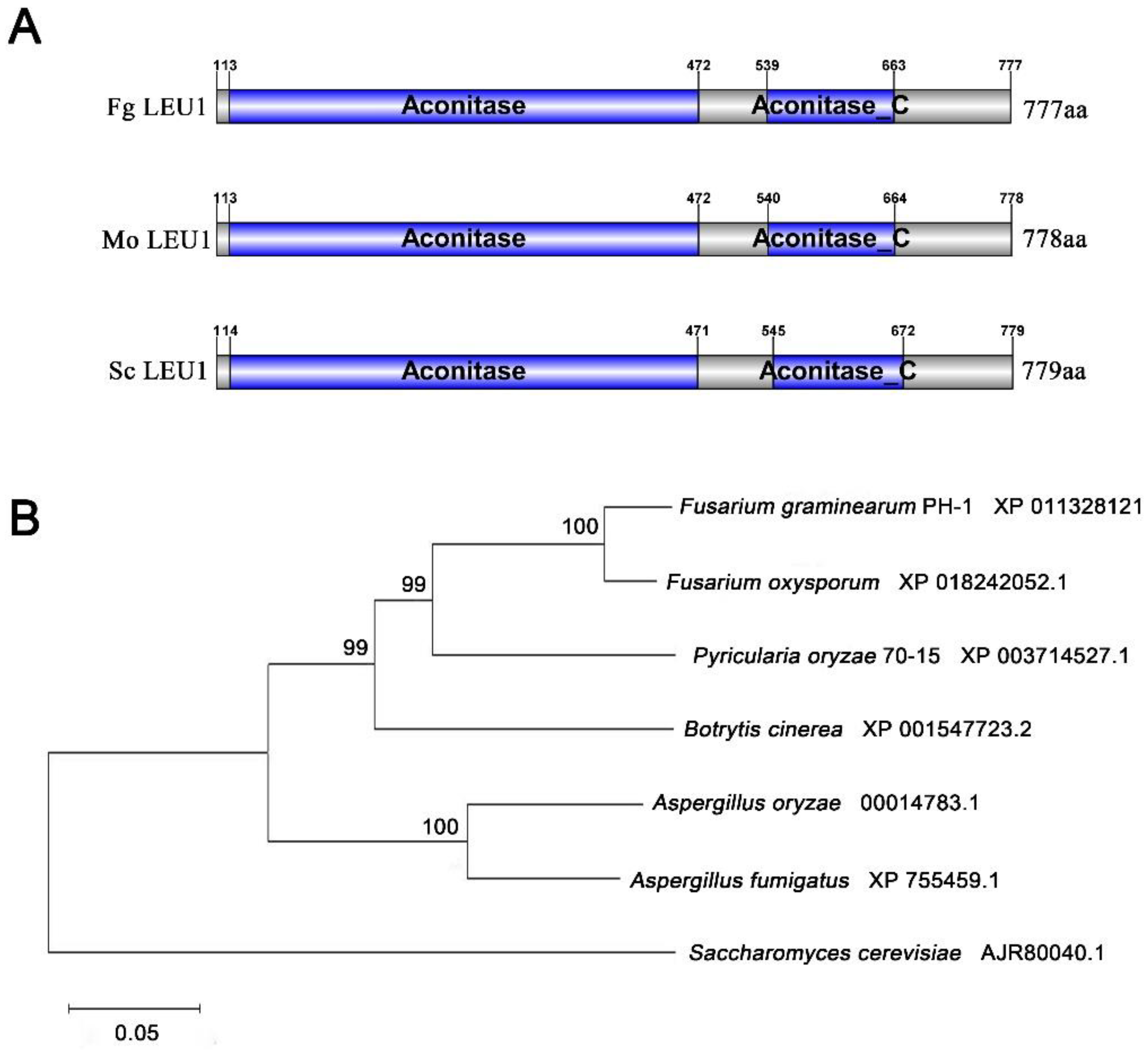
3.1. Identification of FgLEU1 in F. graminearum
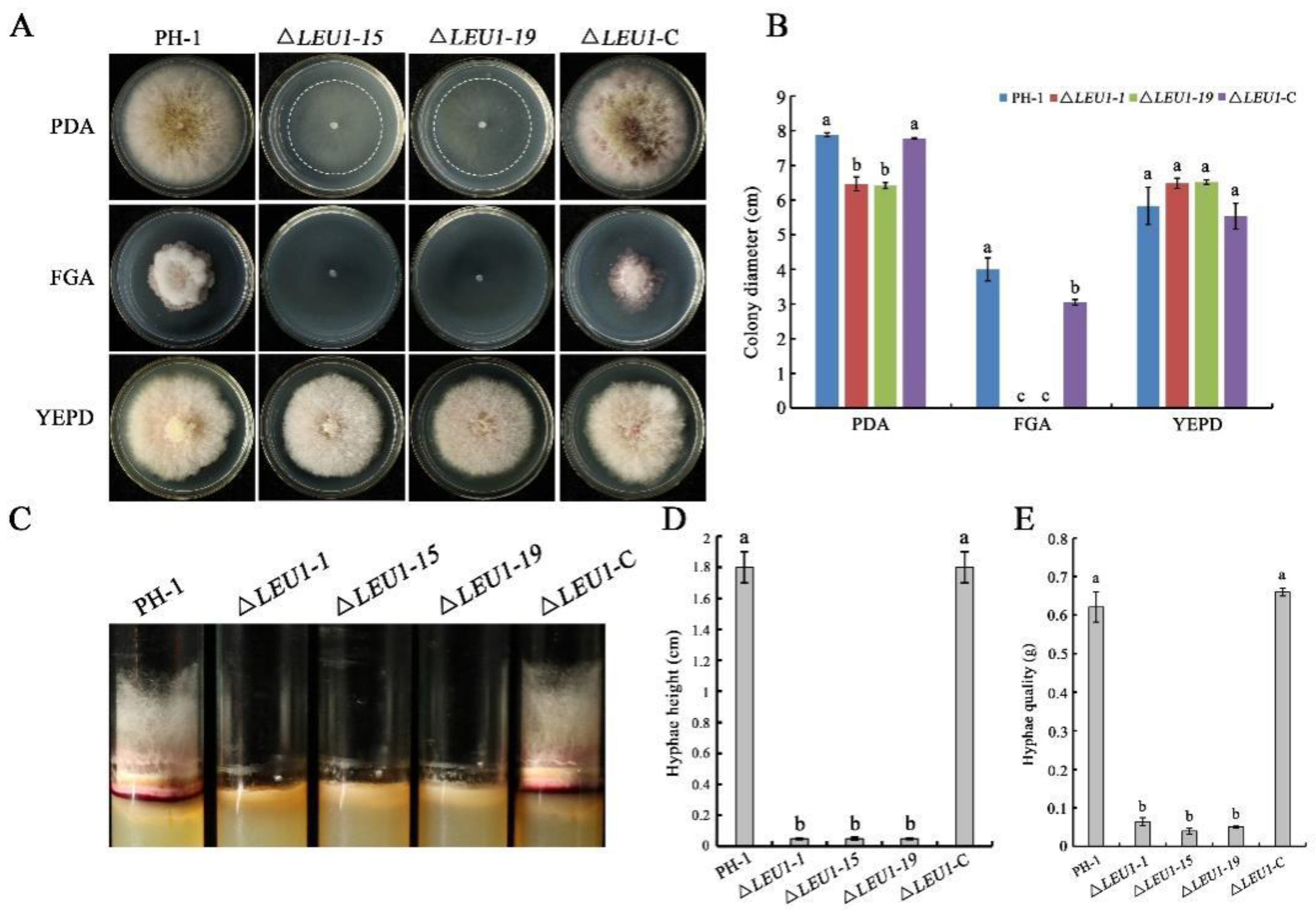
3.2. FgLEU1 Is Responsible for Hyphal Growth, Conidiation, and Conidial Germination
3.3. FgLEU1 Is Necessary for Sexual Reproduction
3.4. FgLEU1 Is Essential for Pathogenicity and DON Production
3.5. Deletion of FgLEU1 Results in Reduced Red Pigmentation
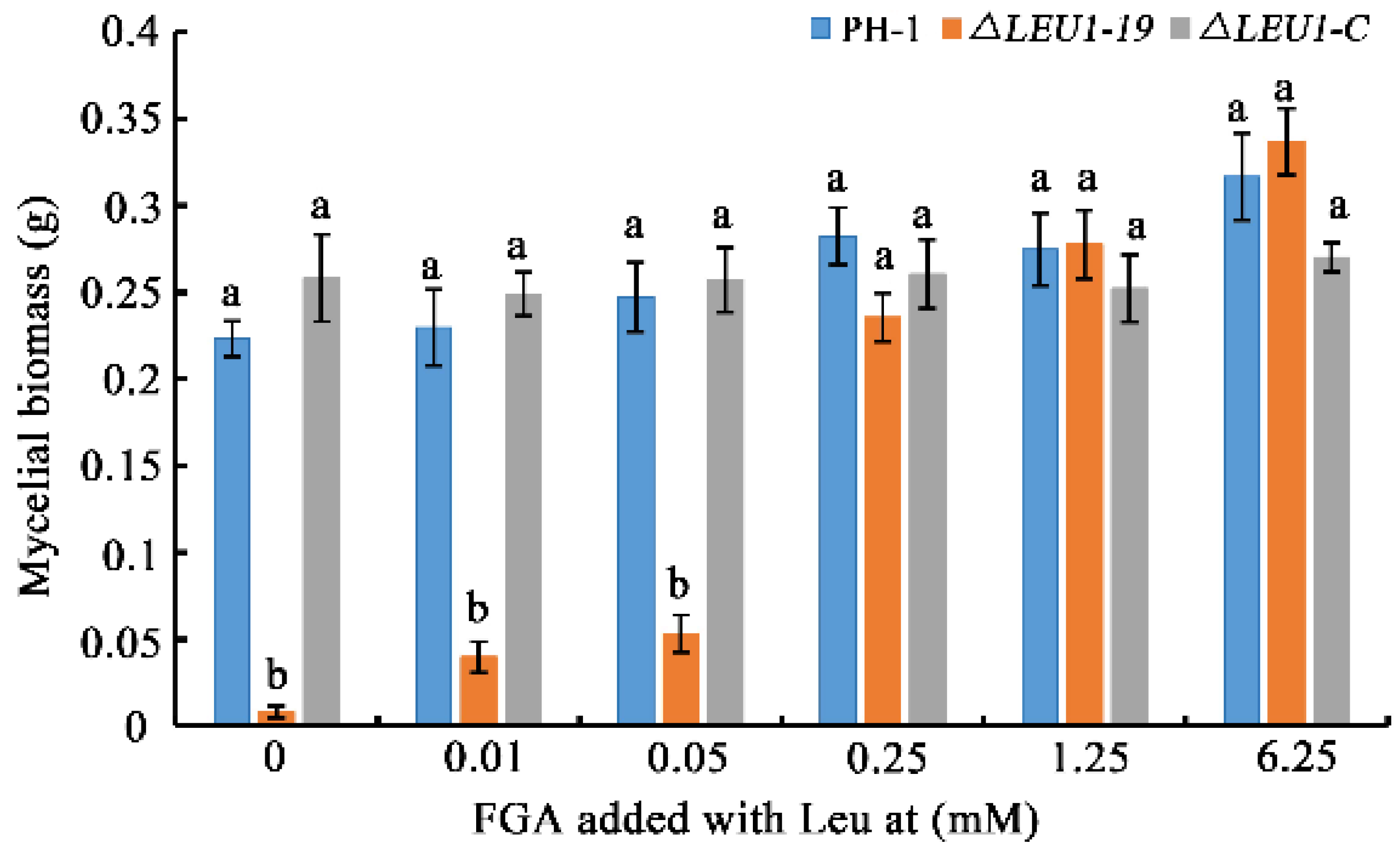
3.6. FgLEU1 Plays a Crucial Role in Leu Biosynthesis in F. graminearum
3.7. Effects of FgLEU1 Deletion on the Maintenance of Cell Wall Integrity
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Goswami, R.S.; Kistler, H.C. Heading for disaster: Fusarium graminearum on cereal crops. Mol. Plant Pathol. 2004, 5, 515–525. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Nicholson, P. Community ecology of fungal pathogens causing wheat head blight. Ann. Rev. Phytopathol. 2009, 47, 83–103. [Google Scholar] [CrossRef]
- McMullen, M.; Jones, R.; Gallenberg, D. Scab of Wheat and Barley: A Re-emerging Disease of Devastating Impact. Plant Dis. 1997, 81, 1340–1348. [Google Scholar] [CrossRef]
- Pestka, J.J. Deoxynivalenol: Mechanisms of action, human exposure, and toxicological relevance. Arch. Toxicol. 2010, 84, 663–679. [Google Scholar] [CrossRef] [PubMed]
- Pestka, J.J.; Zhou, H.R.; Moon, Y.; Chung, Y.J. Cellular and molecular mechanisms for immune modulation by deoxynivalenol and other trichothecenes: Unraveling a paradox. Toxicol. Lett. 2004, 153, 61–73. [Google Scholar] [CrossRef]
- Cuomo, C.A.; Güldener, U.; Xu, J.R.; Trail, F.; Turgeon, B.G.; Di Pietro, A.; Walton, J.D.; Ma, L.J.; Baker, S.E.; Rep, M.; et al. The Fusarium graminearum genome reveals a link between localized polymorphism and pathogen specialization. Science 2007, 317, 1400–1402. [Google Scholar] [CrossRef]
- Fan, X.; He, F.; Ding, M.; Geng, C.; Chen, L.; Zou, S.; Liang, Y.; Yu, J.; Dong, H. Thioredoxin Reductase Is Involved in Development and Pathogenicity in Fusarium graminearum. Front. Microbiol. 2019, 10, 393. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, G.; Dai, Y.; Wang, Y.; Lee, Y.W.; Shi, J.; Xu, J. Biodegradation of Deoxynivalenol by a Novel Microbial Consortium. Front. Microbiol. 2019, 10, 2964. [Google Scholar] [CrossRef]
- Wang, G.; Wang, Y.; Ji, F.; Xu, L.; Yu, M.; Shi, J.; Xu, J. Biodegradation of deoxynivalenol and its derivatives by Devosia insulae A16. Food Chem. 2019, 276, 436–442. [Google Scholar] [CrossRef]
- Binder, S. Branched-Chain Amino Acid Metabolism in Arabidopsis thaliana. Arab. Book. 2010, 8, e0137. [Google Scholar] [CrossRef]
- Boigegrain, R.A.; Liautard, J.P.; Köhler, S. Targeting of the virulence factor acetohydroxyacid synthase by sulfonylureas results in inhibition of intramacrophagic multiplication of Brucella suis. Antimicrob Agents Chemother. 2005, 49, 3922–3925. [Google Scholar] [CrossRef]
- Liu, Y.; Li, Y.; Wang, X. Acetohydroxyacid synthases: Evolution, structure, and function. Appl. Microbiol. Biotechnol. 2016, 100, 8633–8649. [Google Scholar] [CrossRef]
- Wei, W.; Zhou, S.; Cheng, D.; Li, Y.; Liu, J.; Xie, Y.; Li, Y.; Li, Z. Design, synthesis and herbicidal activity study of aryl 2,6-disubstituted sulfonylureas as potent acetohydroxyacid synthase inhibitors. Bioorg. Med. Chem. Lett. 2017, 27, 3365–3369. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Jiang, Y.; Zhang, Y.; Yu, M.; Jiang, H.; Xu, J.; Shi, J. FgIlv3a is crucial in branched-chain amino acid biosynthesis, vegetative differentiation, and virulence in Fusarium graminearum. J. Microbiol. 2019, 57, 694–703. [Google Scholar] [CrossRef]
- Kohlhaw, G.B. Leucine biosynthesis in fungi: Entering metabolism through the back door. Microbiol. Mol. Biol. Rev. 2003, 67, 1–15, table of contents. [Google Scholar] [CrossRef] [PubMed]
- Gruer, M.J.; Artymiuk, P.J.; Guest, J.R. The aconitase family: Three structural variations on a common theme. Trends Biochem. Sci. 1997, 22, 3–6. [Google Scholar] [CrossRef]
- Bedekovics, T.; Li, H.; Gajdos, G.B.; Isaya, G. Leucine biosynthesis regulates cytoplasmic iron-sulfur enzyme biogenesis in an Atm1p-independent manner. J. Biol. Chem. 2011, 286, 40878–40888. [Google Scholar] [CrossRef]
- Do, E.; Hu, G.; Caza, M.; Oliveira, D.; Jung, W.H. Leu1 plays a role in iron metabolism and is required for virulence in Cryptococcus neoformans. Fungal Genet. Biol. 2015, 75, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Wang, X.; Gao, T.; Gu, C.; Sun, F.; Yu, L.; Hu, J. Characterization of the complex involved in regulating V-ATPase activity of the vacuolar and endosomal membrane. J. Bioenerg. Biomembr. 2017, 49, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.; Jiang, H.; Zheng, Q.; Chen, X.; Wang, R.; Yang, S.; Zhao, G.; Liu, J.; Norvienyeku, J.; Wang, Z. Isopropylmalate isomerase MoLeu1 orchestrates leucine biosynthesis, fungal development, and pathogenicity in Magnaporthe oryzae. Appl. Microbiol. Biotechnol. 2019, 103, 327–337. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.Y.; Yu, Q.; Dong, B.; Zhang, Y.; Liu, X.H.; Lin, F.C.; Liang, S. MoLEU1, MoLEU2, and MoLEU4 regulated by MoLEU3 are involved in leucine biosynthesis, fungal development, and pathogenicity in Magnaporthe oryzae. Environ. Microbiol. Rep. 2019, 11, 784–796. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Han, Q.; Wang, J.; Wang, X.; Xu, J.; Shi, J. Two FgLEU2 Genes with Different Roles in Leucine Biosynthesis and Infection-Related Morphogenesis in Fusarium graminearum. PLoS ONE 2016, 11, e0165927. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Zhou, B.; Gao, Z.; Liang, Y. Effects of Phospholipase C on Fusarium graminearum Growth and Development. Curr. Microbiol. 2015, 71, 632–637. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhang, L.; Liu, C.; Sun, S.; Liu, A.; Liang, Y.; Yu, J. The roles of FgPEX2 and FgPEX12 in virulence and lipid metabolism in Fusarium graminearum. Fungal Genet. Biol. 2020, 135, 103288. [Google Scholar] [CrossRef] [PubMed]
- Ding, M.; Zhu, Q.; Liang, Y.; Li, J.; Fan, X.; Yu, X.; He, F.; Xu, H.; Liang, Y.; Yu, J. Differential roles of three FgPLD genes in regulating development and pathogenicity in Fusarium graminearum. Fungal Genet. Biol. 2017, 109, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Mirocha, C.J.; Kolaczkowski, E.; Xie, W.; Yu, H.; Jelen, H. Analysis of deoxynivalenol and its derivatives (batch and single kernel) using gas chromatography/M\mass spectrometry. J. Agric. Food Chem. 1998, 46, 1414–1418. [Google Scholar] [CrossRef]
- Zheng, Q.; Hou, R.; Zhang, J.; Ma, J.; Wu, Z.; Wang, G.; Wang, C.; Xu, J.R. The MAT locus genes play different roles in sexual reproduction and pathogenesis in Fusarium graminearum. PLoS ONE 2013, 8, e66980. [Google Scholar] [CrossRef]
- Ram, A.F.; Klis, F.M. Identification of fungal cell wall mutants using susceptibility assays based on Calcofluor white and Congo red. Nat. Protoc. 2006, 1, 2253–2256. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, J.C.; Omer, S.; Sheldon, J.R.; Welch, I.; Heinrichs, D.E. Role of BrnQ1 and BrnQ2 in branched-chain amino acid transport and virulence in Staphylococcus aureus. Infect. Immun. 2015, 83, 1019–1029. [Google Scholar] [CrossRef]
- Divon, H.H.; Fluhr, R. Nutrition acquisition strategies during fungal infection of plants. FEMS Microbiol. Lett. 2007, 266, 65–74. [Google Scholar] [CrossRef]
- Hoffmeister, D.; Keller, N.P. Natural products of filamentous fungi: Enzymes, genes, and their regulation. Nat. Prod. Rep. 2007, 24, 393–416. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Wang, C.; Hou, R.; Zhou, X.; Li, G.; Zhang, S.; Xu, J.R. The AMT1 arginine methyltransferase gene is important for plant infection and normal hyphal growth in Fusarium graminearum. PLoS ONE 2012, 7, e38324. [Google Scholar] [CrossRef]
- Liu, X.; Xu, J.; Wang, J.; Ji, F.; Yin, X.; Shi, J. Involvement of threonine deaminase FgIlv1 in isoleucine biosynthesis and full virulence in Fusarium graminearum. Curr. Genet. 2015, 61, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Han, Q.; Xu, J.; Wang, J.; Shi, J. Acetohydroxyacid synthase FgIlv2 and FgIlv6 are involved in BCAA biosynthesis, mycelial and conidial morphogenesis, and full virulence in Fusarium graminearum. Sci. Rep. 2015, 5, 16315. [Google Scholar] [CrossRef]
- Liu, X.; Wang, J.; Xu, J.; Shi, J. FgIlv5 is required for branched-chain amino acid biosynthesis and full virulence in Fusarium graminearum. Microbiology 2014, 160, 692–702. [Google Scholar] [CrossRef]
- Yang, P.; Chen, Y.; Wu, H.; Fang, W.; Liang, Q.; Zheng, Y.; Olsson, S.; Zhang, D.; Zhou, J.; Wang, Z.; et al. The 5-oxoprolinase is required for conidiation, sexual reproduction, virulence and deoxynivalenol production of Fusarium graminearum. Curr. Genet. 2018, 64, 285–301. [Google Scholar] [CrossRef]
- Chen, X.; Nielsen, K.F.; Borodina, I.; Kielland-Brandt, M.C.; Karhumaa, K. Increased isobutanol production in Saccharomyces cerevisiae by overexpression of genes in valine metabolism. Biotechnol. Biofuels 2011, 4, 21. [Google Scholar] [CrossRef]
- Trail, F. For blighted waves of grain: Fusarium graminearum in the postgenomics era. Plant Physiol. 2009, 149, 103–110. [Google Scholar] [CrossRef]
- Seong, K.Y.; Pasquali, M.; Zhou, X.; Song, J.; Hilburn, K.; McCormick, S.; Dong, Y.; Xu, J.R.; Kistler, H.C. Global gene regulation by Fusarium transcription factors Tri6 and Tri10 reveals adaptations for toxin biosynthesis. Mol. Microbiol. 2009, 72, 354–367. [Google Scholar] [CrossRef]
- Desjardins, A.E.; Bai, G.H.; Plattner, R.D.; Proctor, R.H. Analysis of aberrant virulence of Gibberella zeae following transformation-mediated complementation of a trichothecene-deficient (Tri5) mutant. Microbiology 2000, 146 Pt 8, 2059–2068. [Google Scholar] [CrossRef]
- Cambaza, E. Comprehensive Description of Fusarium graminearum Pigments and Related Compounds. Foods 2018, 7, 165. [Google Scholar] [CrossRef] [PubMed]
- Que, Y.; Yue, X.; Yang, N.; Xu, Z.; Tang, S.; Wang, C.; Lv, W.; Xu, L.; Talbot, N.J.; Wang, Z. Leucine biosynthesis is required for infection-related morphogenesis and pathogenicity in the rice blast fungus Magnaporthe Oryzae. Curr. Genet. 2020, 66, 155–171. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Conidiation (105 conidia/mL) α | Germination (%) β | Perithecia Counts γ | DON Production (ppm) δ |
|---|---|---|---|---|
| PH-1 | 7.33 ± 0.74 a | 60.34 ± 0.02 b | 38.44 ± 2.2305 a | 14.3733 ± 0.6 b |
| ΔLEU1-15 | 1.67 ± 0.4 b | 42.32 ± 0.03 c | 0 b | 0.0133 ± 0.01 c |
| ΔLEU1-19 | 1.42 ± 0.48 b | 41.44 ± 0.03 c | 0 b | 0.0367 ± 0.03 c |
| ΔLEU1-C | 7.12 ± 0.98 a | 71.91 ± 0.00 a | 38.55 ± 1.0943 a | 17.0533 ± 0.34 a |
| Strain | Relative Expression Level | ||
|---|---|---|---|
| Tri5 | Tri6 | Tri10 | |
| PH-1 | 1 ± 0.02 a | 1 ± 0.01 a | 1 ± 0.02 a |
| ΔLEU1-19 | 0.115 ± 0.02 b | 0.1691 ± 0.01 d | 0.0936 ± 0.00 b |
| ΔLEU1-19 + Leu | 0.944 ± 0.02 a | 0.5274 ± 0.03 c | 1.0341 ± 0.03 a |
| ΔLEU1-C | 1.0257 ± 0.14 a | 0.8332 ± 0.02 b | 0.9622 ± 0.04 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, S.; Wang, M.; Liu, C.; Tao, Y.; Wang, T.; Liang, Y.; Zhang, L.; Yu, J. FgLEU1 Is Involved in Leucine Biosynthesis, Sexual Reproduction, and Full Virulence in Fusarium graminearum. J. Fungi 2022, 8, 1090. https://doi.org/10.3390/jof8101090
Sun S, Wang M, Liu C, Tao Y, Wang T, Liang Y, Zhang L, Yu J. FgLEU1 Is Involved in Leucine Biosynthesis, Sexual Reproduction, and Full Virulence in Fusarium graminearum. Journal of Fungi. 2022; 8(10):1090. https://doi.org/10.3390/jof8101090
Chicago/Turabian StyleSun, Shaohua, Mingyu Wang, Chunjie Liu, Yilin Tao, Tian Wang, Yuancun Liang, Li Zhang, and Jinfeng Yu. 2022. "FgLEU1 Is Involved in Leucine Biosynthesis, Sexual Reproduction, and Full Virulence in Fusarium graminearum" Journal of Fungi 8, no. 10: 1090. https://doi.org/10.3390/jof8101090
APA StyleSun, S., Wang, M., Liu, C., Tao, Y., Wang, T., Liang, Y., Zhang, L., & Yu, J. (2022). FgLEU1 Is Involved in Leucine Biosynthesis, Sexual Reproduction, and Full Virulence in Fusarium graminearum. Journal of Fungi, 8(10), 1090. https://doi.org/10.3390/jof8101090