
Genome-Wide Identification of bZIP Transcription Factor Genes and Functional Analyses of Two Members in Cytospora chrysosperma


Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification of bZIP Transcription Factors in C. chrysosperma
2.2. Gene Structural Characterization and Phylogenetic Analysis
2.3. Conditions for Fungal Strains’ Growth and Treatment
2.4. Generation of CcbZIP Gene Deletion Mutants
2.5. Pathogenicity Tests
2.6. RNA Extraction and RT-qPCR Analysis
3. Results
3.1. Identification of bZIP Genes in C. chrysosperma
3.2. Analysis of Conserved Domains and Motif in CcbZIP Proteins
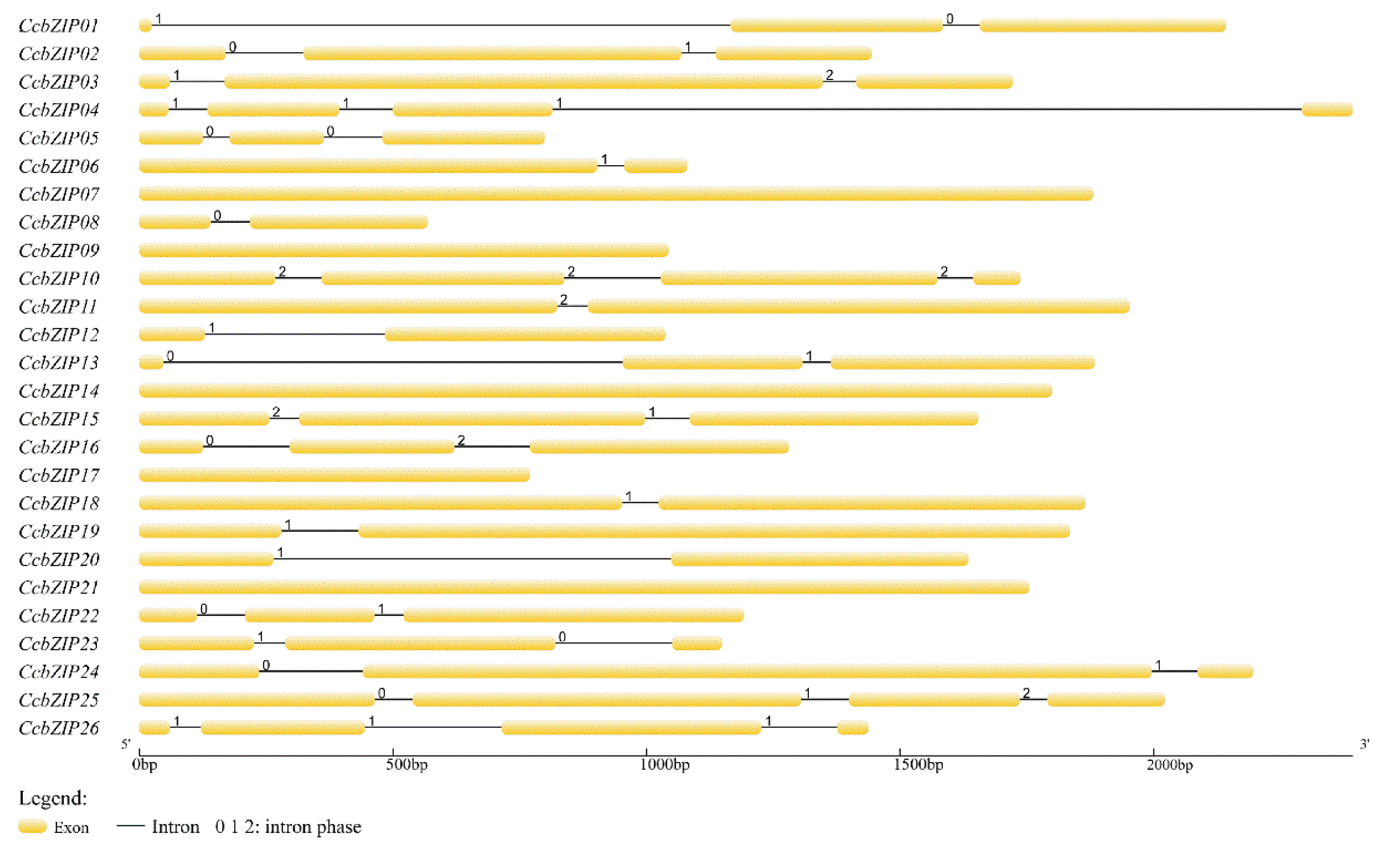
3.3. Intron Numbers and Their Distribution Pattern in CcbZIPs
3.4. Phylogenetic Analysis of bZIP Gene Family
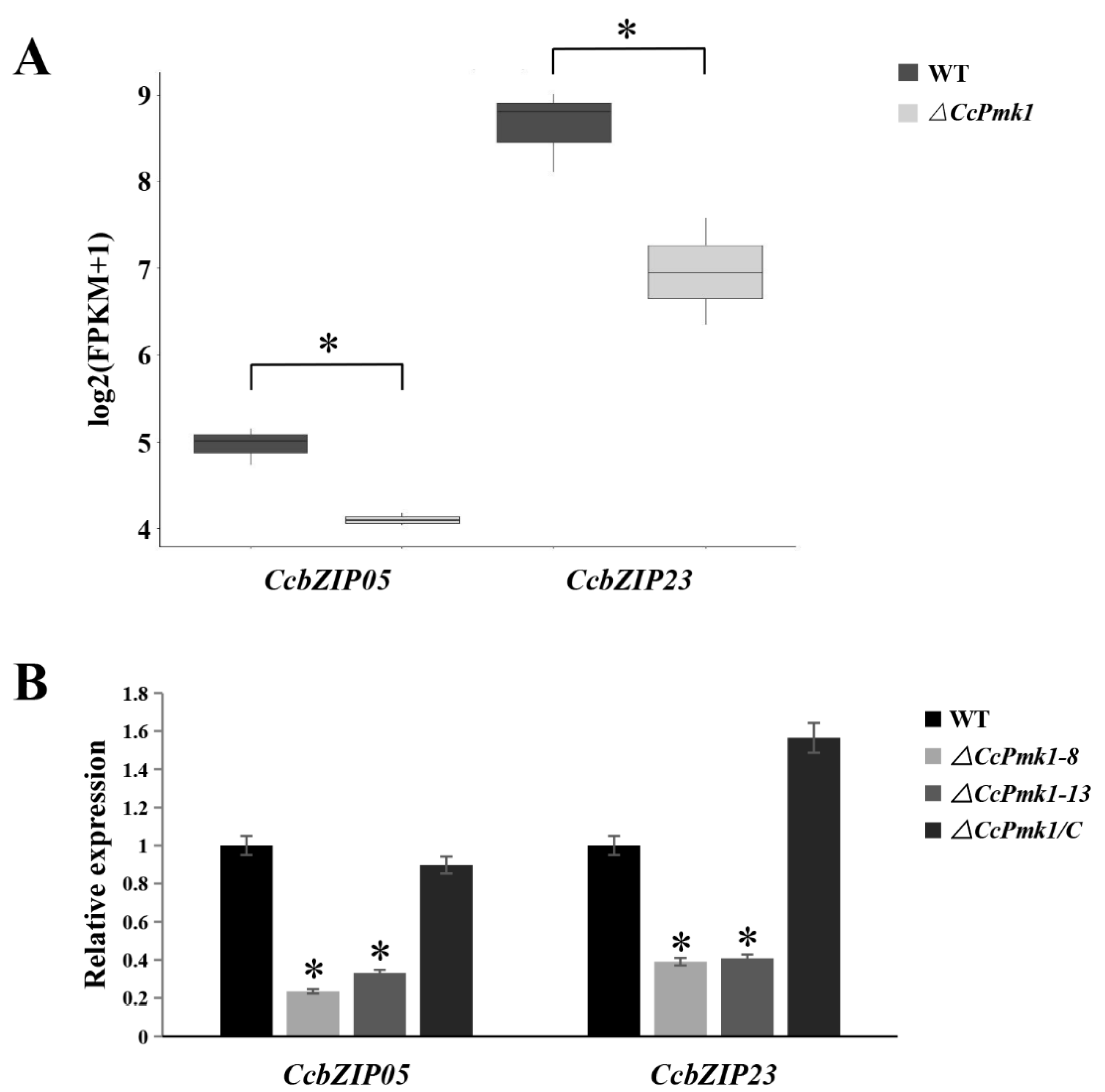
3.5. Expression Patterns of CcbZIPs during the Initial Infection Stages
3.6. Construction of CcbZIP Deletion Mutants
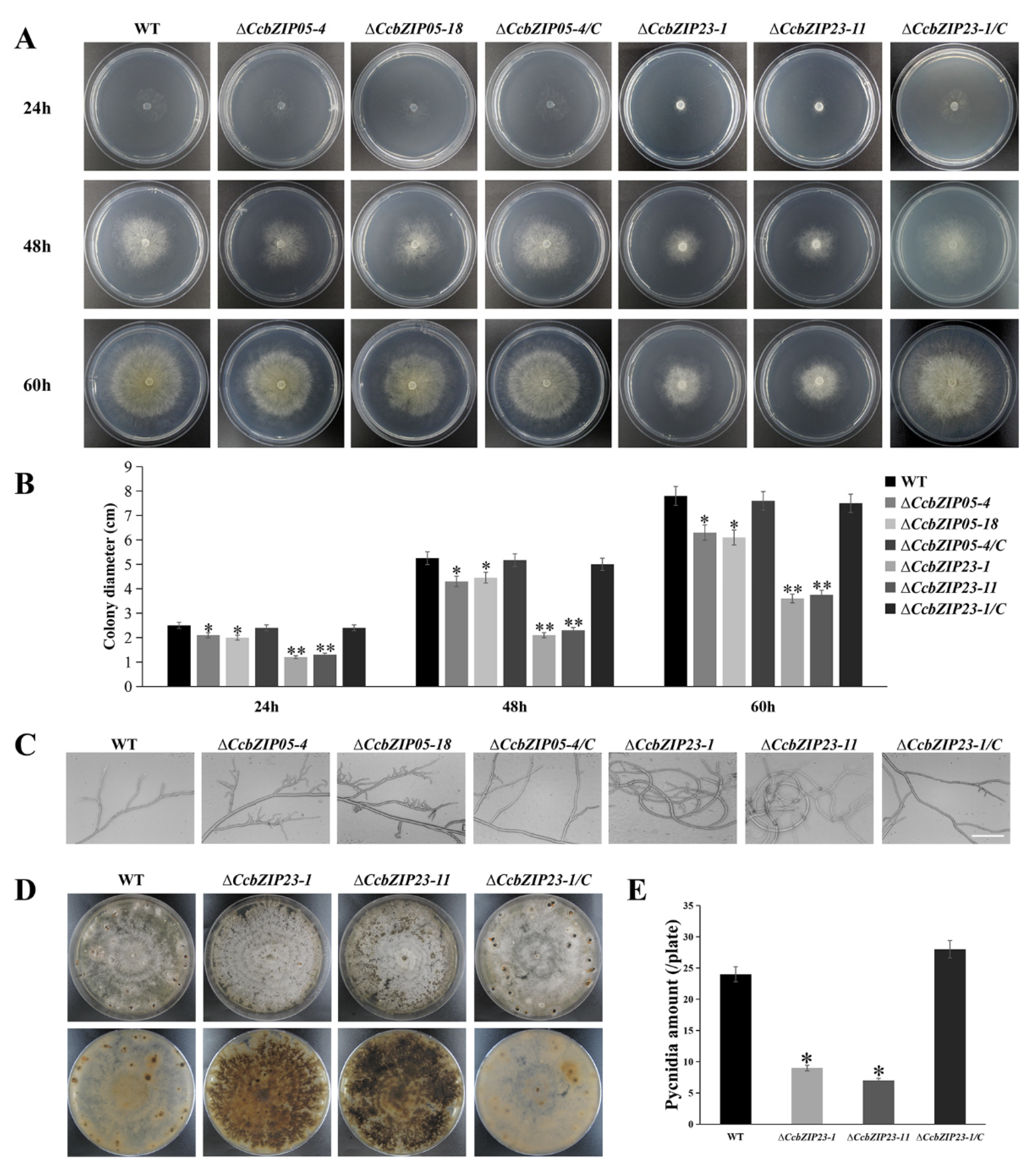
3.7. CcbZIP05 and CcbZIP23 Are Important for the Development of C. chrysosperma
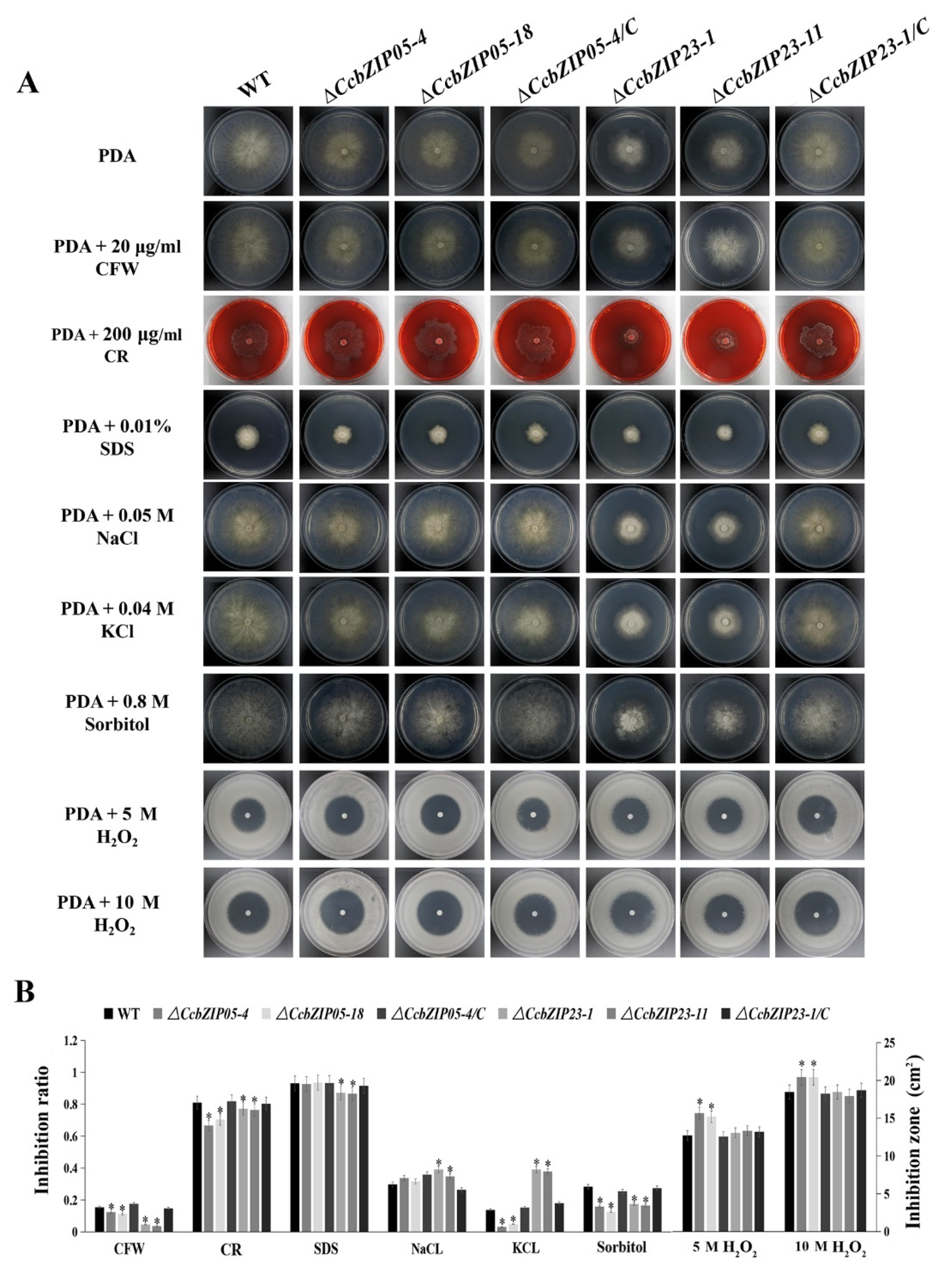
3.8. CcbZIP05 and CcbZIP23 Are Required for the Stress Responses
3.9. CcbZIP05 and CcbZIP23 Are Involved in Pathogenicity
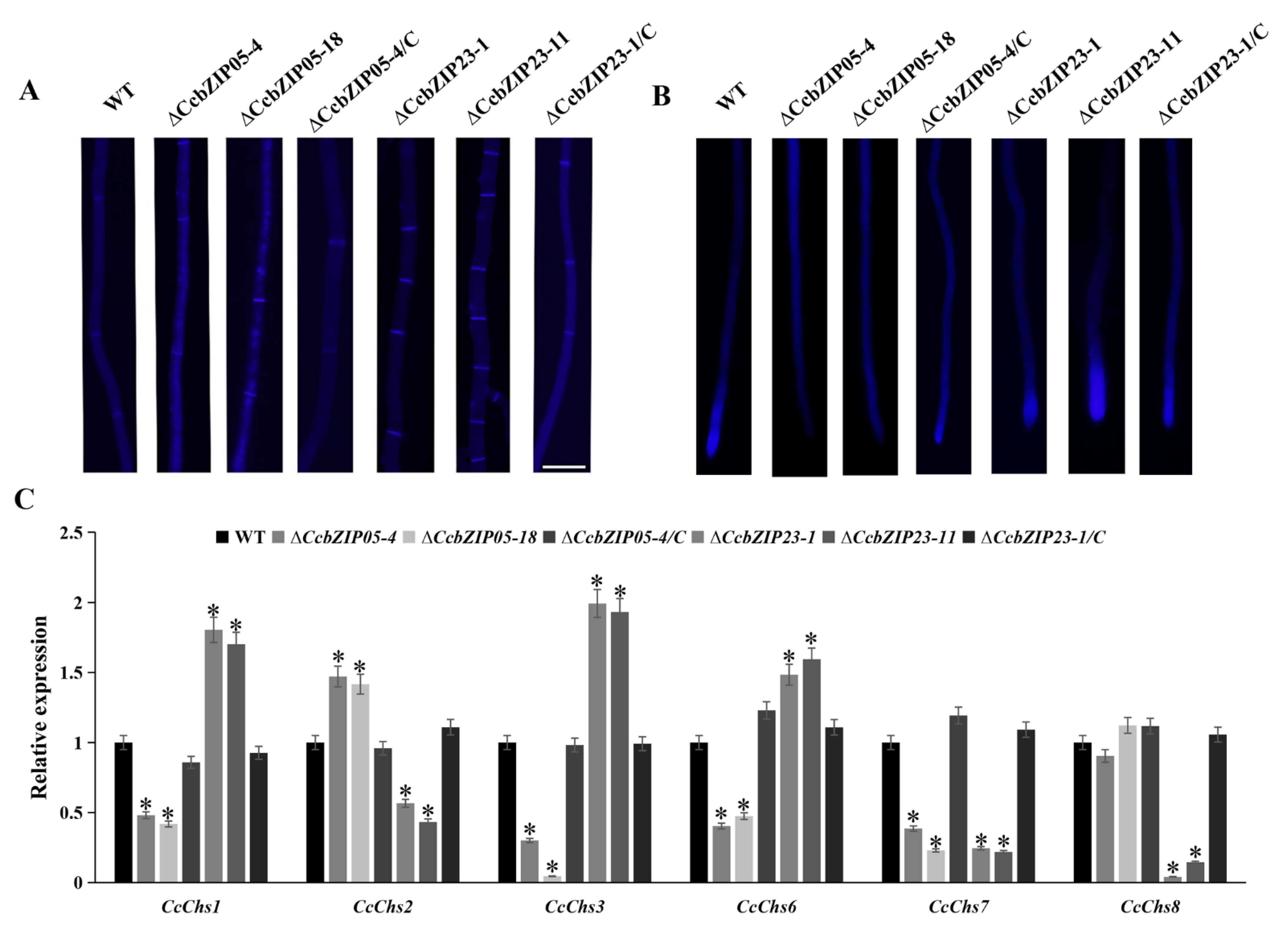
3.10. CcbZIP05 and CcbZIP23 Differentially Regulate the Expression of Putative Effector Genes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Choi, J.; Kim, Y.; Kim, S.; Park, J.; Lee, Y.H. MoCRZ1, a gene encoding a calcineurin-responsive transcription factor, regulates fungal growth and pathogenicity of Magnaporthe oryzae. Fungal Genet. Biol. 2009, 46, 243–254. [Google Scholar] [CrossRef]
- Kim, S.; Park, S.Y.; Kim, K.S.; Rho, H.S.; Chi, M.H.; Choi, J.; Park, J.; Kong, S.; Park, J.; Goh, J.; et al. Homeobox transcription factors are required for conidiation and appressorium development in the rice blast fungus Magnaporthe oryzae. PLoS Genet. 2009, 5, e1000757. [Google Scholar] [CrossRef] [Green Version]
- Mehrabi, R.; Ding, S.; Xu, J.R. MADS-box transcription factor mig1 is required for infectious growth in Magnaporthe grisea. Eukaryot. Cell 2008, 7, 791–799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwechheimer, C.; Bevan, M. The regulation of transcription factor activity in plants. Trends Plant Sci. 1998, 3, 378–383. [Google Scholar] [CrossRef]
- Meshi, T.; Iwabuchi, M. Plant transcription factors. Plant Cell Physiol. 1995, 36, 1405–1420. [Google Scholar]
- Pabo, C.O.; Sauer, R.T. Transcription factors: Structural families and principles of DNA recognition. Annu. Rev. Biochem. 1992, 61, 1053–1095. [Google Scholar] [CrossRef]
- Park, J.; Park, J.; Jang, S.; Kim, S.; Kong, S.; Choi, J.; Ahn, K.; Kim, J.; Lee, S.; Kim, S.; et al. FTFD: An informatics pipeline supporting phylogenomic analysis of fungal transcription factors. Bioinformatics 2008, 24, 1024–1025. [Google Scholar] [CrossRef] [Green Version]
- Gerin, D.; Garrapa, F.; Ballester, A.R.; González-Candelas, L.; De Miccolis Angelini, R.M.; Faretra, F.; Pollastro, S. Functional role of Aspergillus carbonarius AcOTAbZIP gene, a bZIP transcription factor within the OTA gene cluster. Toxins 2021, 13, 111. [Google Scholar] [CrossRef]
- Leiter, É.; Emri, T.; Pákozdi, K.; Hornok, L.; Pócsi, I. The impact of bZIP Atf1ortholog global regulators in fungi. Appl. Microbiol. Biotechnol. 2021, 105, 5769–5783. [Google Scholar] [CrossRef] [PubMed]
- López-Berges, M.S.; Scheven, M.T.; Hortschansky, P.; Misslinger, M.; Baldin, C.; Gsaller, F.; Werner, E.R.; Krüger, T.; Kniemeyer, O.; Weber, J.; et al. The bZIP transcription factor HapX Is post-translationally regulated to control iron homeostasis in Aspergillus fumigatus. Int. J. Mol. Sci. 2021, 22, 7739. [Google Scholar] [CrossRef] [PubMed]
- Hurst, H.C. Transcription factors 1: bZIP proteins. Protein Profile 1995, 2, 101–168. [Google Scholar]
- Vinson, C.; Acharya, A.; Taparowsky, E.J. Deciphering B-ZIP transcription factor interactions in vitro and in vivo. Biochim. Biophys. Acta 2006, 1759, 4–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vinson, C.R.; Sigler, P.B.; McKnight, S.L. Scissors-grip model for DNA recognition by a family of leucine zipper proteins. Science 1989, 246, 911–916. [Google Scholar] [CrossRef] [PubMed]
- Jakoby, M.; Weisshaar, B.; Dröge-Laser, W.; Vicente-Carbajosa, J.; Tiedemann, J.; Kroj, T.; Parcy, F. bZIP transcription factors in Arabidopsis. Trends Plant Sci. 2002, 7, 106–111. [Google Scholar] [CrossRef]
- Fassler, J.; Landsman, D.; Acharya, A.; Moll, J.R.; Bonovich, M.; Vinson, C. B-ZIP proteins encoded by the Drosophila genome: Evaluation of potential dimerization partners. Genome Res. 2002, 12, 1190–1200. [Google Scholar] [CrossRef] [Green Version]
- Landschulz, W.H.; Johnson, P.F.; McKnight, S.L. The leucine zipper: A hypothetical structure common to a new class of DNA binding proteins. Science 1988, 240, 1759–1764. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Xiong, L.; Gao, J.; Zhang, H.Y. Molecular mechanisms of the protein-protein interaction-regulated binding specificity of basic-region leucine zipper transcription factors. J. Mol. Model. 2019, 25, 246. [Google Scholar] [CrossRef]
- Vinson, C.R.; Hai, T.; Boyd, S.M. Dimerization specificity of the leucine zipper-containing bZIP motif on DNA binding: Prediction and rational design. Genes Dev. 1993, 7, 1047–1058. [Google Scholar] [CrossRef] [Green Version]
- Liao, Y.; Zou, H.F.; Wei, W.; Hao, Y.J.; Tian, A.G.; Huang, J.; Liu, Y.F.; Zhang, J.S.; Chen, S.Y. Soybean GmbZIP44, GmbZIP62 and GmbZIP78 genes function as negative regulator of ABA signaling and confer salt and freezing tolerance in transgenic Arabidopsis. Planta 2008, 228, 225–240. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Chen, N.; Chen, F.; Cai, B.; Dal Santo, S.; Tornielli, G.B.; Pezzotti, M.; Cheng, Z.M. Genome-wide analysis and expression profile of the bZIP transcription factor gene family in grapevine (Vitis vinifera). BMC Genom. 2014, 15, 281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niu, X.; Guiltinan, M.J. DNA binding specificity of the wheat bZIP protein EmBP-1. Nucleic Acids Res. 1994, 22, 4969–4978. [Google Scholar] [CrossRef] [Green Version]
- Kong, S.; Park, S.Y.; Lee, Y.H. Systematic characterization of the bZIP transcription factor gene family in the rice blast fungus, Magnaporthe oryzae. Environ. Microbiol. 2015, 17, 1425–1443. [Google Scholar] [CrossRef]
- Xu, Y.; Wang, Y.; Zhao, H.; Wu, M.; Zhang, J.; Chen, W.; Li, G.; Yang, L. Genome-wide identification and expression analysis of the bzip transcription factors in the mycoparasite Coniothyrium minitans. Microorganisms 2020, 8, 1045. [Google Scholar] [CrossRef] [PubMed]
- Son, H.; Seo, Y.S.; Min, K.; Park, A.R.; Lee, J.; Jin, J.M.; Lin, Y.; Cao, P.; Hong, S.Y.; Kim, E.K.; et al. A phenome-based functional analysis of transcription factors in the cereal head blight fungus, Fusarium graminearum. PLoS Pathog. 2011, 7, e1002310. [Google Scholar] [CrossRef] [PubMed]
- Yin, W.; Cui, P.; Wei, W.; Lin, Y.; Luo, C. Genome-wide identification and analysis of the basic leucine zipper (bZIP) transcription factor gene family in Ustilaginoidea virens. Genome 2017, 60, 1051–1059. [Google Scholar] [CrossRef] [Green Version]
- Guo, M.; Guo, W.; Chen, Y.; Dong, S.; Zhang, X.; Zhang, H.; Song, W.; Wang, W.; Wang, Q.; Lv, R.; et al. The basic leucine zipper transcription factor Moatf1 mediates oxidative stress responses and is necessary for full virulence of the rice blast fungus Magnaporthe oryzae. Mol. Plant-Microbe Interact. 2010, 23, 1053–1068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, C.; Zhang, S.; Zhang, Q.; Tao, Y.; Wang, C.; Xu, J.R. FgSKN7 and FgATF1 have overlapping functions in ascosporogenesis, pathogenesis and stress responses in Fusarium graminearum. Environ. Microbiol. 2015, 17, 1245–1260. [Google Scholar] [CrossRef]
- Nathues, E.; Joshi, S.; Tenberge, K.B.; von den Driesch, M.; Oeser, B.; Bäumer, N.; Mihlan, M.; Tudzynski, P. CPTF1, a CREB-like transcription factor, is involved in the oxidative stress response in the phytopathogen Claviceps purpurea and modulates ROS level in its host Secale cereale. Mol. Plant-Microbe Interact. 2004, 17, 383–393. [Google Scholar] [CrossRef] [Green Version]
- Qi, X.; Guo, L.; Yang, L.; Huang, J. Foatf1, a bZIP transcription factor of Fusarium oxysporum f. sp. cubense, is involved in pathogenesis by regulating the oxidative stress responses of Cavendish banana (Musa spp.). Physiol. Mol. Plant Pathol. 2013, 84, 76–85. [Google Scholar] [CrossRef]
- Temme, N.; Oeser, B.; Massaroli, M.; Heller, J.; Simon, A.; Collado, I.G.; Viaud, M.; Tudzynski, P. BcAtf1, a global regulator, controls various differentiation processes and phytotoxin production in Botrytis cinerea. Mol. Plant Pathol. 2012, 13, 704–718. [Google Scholar] [CrossRef]
- Van Nguyen, T.; Kröger, C.; Bönnighausen, J.; Schäfer, W.; Bormann, J. The ATF/CREB transcription factor Atf1 is essential for full virulence, deoxynivalenol production, and stress tolerance in the cereal pathogen Fusarium graminearum. Mol. Plant-Microbe Interact. 2013, 26, 1378–1394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.; Wang, Y.; Tian, C. bZIP transcription factor CgAP1 is essential for oxidative stress tolerance and full virulence of the poplar anthracnose fungus Colletotrichum gloeosporioides. Fungal Genet. Biol. 2016, 95, 58–66. [Google Scholar] [CrossRef]
- López-Berges, M.S.; Capilla, J.; Turrà, D.; Schafferer, L.; Matthijs, S.; Jöchl, C.; Cornelis, P.; Guarro, J.; Haas, H.; Di Pietro, A. HapX-mediated iron homeostasis is essential for rhizosphere competence and virulence of the soilborne pathogen Fusarium oxysporum. Plant Cell 2012, 24, 3805–3822. [Google Scholar] [CrossRef] [Green Version]
- Szabó, Z.; Pákozdi, K.; Murvai, K.; Pusztahelyi, T.; Kecskeméti, Á.; Gáspár, A.; Logrieco, A.F.; Emri, T.; Ádám, A.L.; Leiter, É.; et al. FvatfA regulates growth, stress tolerance as well as mycotoxin and pigment productions in Fusarium verticillioides. Appl. Microbiol. Biotechnol. 2020, 104, 7879–7899. [Google Scholar] [CrossRef] [PubMed]
- Iracane, E.; Donovan, P.D.; Ola, M.; Butler, G.; Holland, L.M. Identification of an exceptionally long intron in the HAC1 gene of Candida parapsilosis. mSphere 2018, 3, e00532-18. [Google Scholar] [CrossRef] [Green Version]
- Wong, K.H.; Hynes, M.J.; Todd, R.B.; Davis, M.A. Transcriptional control of nmrA by the bZIP transcription factor MeaB reveals a new level of nitrogen regulation in Aspergillus nidulans. Mol. Microbiol. 2007, 66, 534–551. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.S.; Kim, Y.H.; Shin, K.S. The developmental regulators, FlbB and FlbE, are involved in the virulence of Aspergillus fumigatus. J. Microbiol. Biotechnol. 2013, 23, 766–770. [Google Scholar] [CrossRef] [Green Version]
- Tang, W.; Ru, Y.; Hong, L.; Zhu, Q.; Zuo, R.; Guo, X.; Wang, J.; Zhang, H.; Zheng, X.; Wang, P.; et al. System-wide characterization of bZIP transcription factor proteins involved in infection-related morphogenesis of Magnaporthe oryzae. Environ. Microbiol. 2015, 17, 1377–1396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, G.C.; Roux, J.; Wingfield, M.J. Cytospora species (Ascomycota, Diaporthales, Valsaceae): Introduced and native pathogens of trees in South Africa. Australas. Plant Pathol. 2006, 35, 521–548. [Google Scholar] [CrossRef]
- Bagherabadi, S.; Zafari, D.; Soleimani, M.J. Morphological and molecular identification of Cytospora chrysosperma causing canker disease on Prunus persica. Australas. Plant Dis. Notes 2017, 12, 26. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Zang, R.; Yin, Z.; Kang, Z.; Huang, L. Delimiting cryptic pathogen species causing apple Valsa canker with multilocus data. Ecol. Evol. 2014, 4, 1369–1380. [Google Scholar] [CrossRef]
- Fan, X.; Hyde, K.; Yang, Q.; Liang, Y.-M.; Ma, R.; Tian, C. Cytospora species associated with canker disease of three anti-desertification plants in northwestern China. Phytotaxa 2015, 197, 227–244. [Google Scholar] [CrossRef] [Green Version]
- Biggs, A.R.; Davis, D.D.; Merrill, W. Histopathology of cankers on Populus caused by Cytospora chrysosperma. Can. J. Bot. 1983, 61, 563–574. [Google Scholar] [CrossRef]
- Tao, D.; Li, P.H.; Carter, J.V.; Ostry, M.E. Relationship of environmental stress and Cytospora chrysosperma infection to spring dieback of poplar shoots. Forest Sci. 1984, 30, 645–651. [Google Scholar] [CrossRef]
- Zhang, J.E.; Liang, Y.M.; Tian, C.M. Development process of pycnidia in Cytospora chrysosperma. Mycosystema 2017, 36, 573–581. [Google Scholar] [CrossRef]
- Wang, X.; Zha, W.; Liang, L.; Fasoyin, O.E.; Wu, L.; Wang, S. The bZIP transcription factor AflRsmA regulates Aflatoxin B(1) biosynthesis, oxidative stress response and sclerotium formation in Aspergillus flavus. Toxins 2020, 12, 271. [Google Scholar] [CrossRef] [Green Version]
- Xiong, D.; Yu, L.; Shan, H.; Tian, C. CcPmk1 is a regulator of pathogenicity in Cytospora chrysosperma and can be used as a potential target for disease control. Mol. Plant Pathol. 2021, 22, 710–726. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Xiong, D.; Han, Z.; Liang, Y.; Tian, C. The mitogen-activated protein kinase gene CcPmk1 is required for fungal growth, cell wall integrity and pathogenicity in Cytospora chrysosperma. Fungal Genet. Biol. 2019, 128, 1–13. [Google Scholar] [CrossRef]
- Han, Z.; Yu, R.; Xiong, D.; Tian, C. A Sge1 homolog in Cytospora chrysosperma governs conidiation, virulence and the expression of putative effectors. Gene 2021, 778, 145474. [Google Scholar] [CrossRef] [PubMed]
- Han, Z.; Xiong, D.; Xu, Z.; Liu, T.; Tian, C. The Cytospora chrysosperma virulence effector CcCAP1 mainly localizes to the plant nucleus to suppress plant immune responses. mSphere 2021, 6, e00883-20. [Google Scholar] [CrossRef]
- Prakash, A.; Jeffryes, M.; Bateman, A.; Finn, R.D. The HMMER web server for protein sequence similarity search. Curr. Protoc. Bioinform. 2017, 60, 3–15. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicholas, K.; Nicholas, H.; Deerfield, D. GeneDoc: Analysis and visualization of genetic variation. Embnew. News 1997, 4, 14. [Google Scholar]
- Silvestro, D.; Michalak, I. raxmlGUI: A graphical front-end for RAxML. Org. Divers. Evol. 2012, 12, 335–337. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Fan, X.L.; Bezerra, J.D.P.; Tian, C.M.; Crous, P.W. Cytospora (Diaporthales) in China. Persoonia 2020, 45, 1–45. [Google Scholar] [CrossRef]
- Liu, L.L.; Wang, Y.L.; Xiong, D.G.; Xu, X.; Tian, C.M.; Liang, Y.M. Genetic transformation system of Cytospora chrysosperma, the causal agent of poplar canker. Microbiol. China 2017, 44, 2487–2497. [Google Scholar] [CrossRef]
- Boudet, N.; Aubourg, S.; Toffano-Nioche, C.; Kreis, M.; Lecharny, A. Evolution of intron/exon structure of DEAD helicase family genes in Arabidopsis, Caenorhabditis, and Drosophila. Genome Res. 2001, 11, 2101–2114. [Google Scholar] [CrossRef] [Green Version]
- Callis, J.; Fromm, M.; Walbot, V. Introns increase gene expression in cultured maize cells. Genes Dev. 1987, 1, 1183–1200. [Google Scholar] [CrossRef] [Green Version]
- Jindrich, K.; Degnan, B.M. The diversification of the basic leucine zipper family in eukaryotes correlates with the evolution of multicellularity. BMC Evol. Biol. 2016, 16, 28. [Google Scholar] [CrossRef] [Green Version]
- Li, X.Y.; Xiong, D.G.; Tian, C.M. Genome-wide identification, phylogeny and transcriptional profiling of SNARE genes in Cytospora chrysosperma. J. Phytopathol. 2021, 169, 471–485. [Google Scholar] [CrossRef]
- Turrà, D.; Segorbe, D.; Di Pietro, A. Protein kinases in plant-pathogenic fungi: Conserved regulators of infection. Annu. Rev. Phytopathol. 2014, 52, 267–288. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.; Bui, D.C.; Kim, S.; Jung, S.Y.; Nam, H.J.; Lim, J.Y.; Choi, G.J.; Lee, Y.W.; Kim, J.E.; Son, H. The novel bZIP transcription factor Fpo1 negatively regulates perithecial development by modulating carbon metabolism in the ascomycete fungus Fusarium graminearum. Environ. Microbiol. 2020, 22, 2596–2612. [Google Scholar] [CrossRef] [PubMed]
- Gladieux, P.; Ropars, J.; Badouin, H.; Branca, A.; Aguileta, G.; de Vienne, D.M.; Rodríguez de la Vega, R.C.; Branco, S.; Giraud, T. Fungal evolutionary genomics provides insight into the mechanisms of adaptive divergence in eukaryotes. Mol. Ecol. 2014, 23, 753–773. [Google Scholar] [CrossRef]
- Nuruzzaman, M.; Manimekalai, R.; Sharoni, A.M.; Satoh, K.; Kondoh, H.; Ooka, H.; Kikuchi, S. Genome-wide analysis of NAC transcription factor family in rice. Gene 2010, 465, 30–44. [Google Scholar] [CrossRef]
- Amaike, S.; Affeldt, K.J.; Yin, W.B.; Franke, S.; Choithani, A.; Keller, N.P. The bZIP protein MeaB mediates virulence attributes in Aspergillus flavus. PLoS ONE 2013, 8, e74030. [Google Scholar] [CrossRef] [Green Version]
- Tang, C.; Li, T.; Klosterman, S.J.; Tian, C.; Wang, Y. The bZIP transcription factor VdAtf1 regulates virulence by mediating nitrogen metabolism in Verticillium dahliae. New Phytol. 2020, 226, 1461–1479. [Google Scholar] [CrossRef]
- Wang, Y.; Deng, C.; Tian, L.; Xiong, D.; Tian, C.; Klosterman, S.J. The transcription factor VdHapX controls iron homeostasis and is crucial for virulence in the vascular pathogen Verticillium dahliae. mSphere 2018, 3, e00400-18. [Google Scholar] [CrossRef] [Green Version]
- Gai, Y.; Liu, B.; Ma, H.; Li, L.; Chen, X.; Moenga, S.; Riely, B.; Fayyaz, A.; Wang, M.; Li, H. The methionine biosynthesis regulator AaMetR contributes to oxidative stress tolerance and virulence in Alternaria alternata. Microbiol. Res. 2019, 219, 94–109. [Google Scholar] [CrossRef]
- Guo, X.Y.; Li, Y.; Fan, J.; Xiong, H.; Xu, F.X.; Shi, J.; Shi, Y.; Zhao, J.Q.; Wang, Y.F.; Cao, X.L.; et al. Host-induced gene silencing of MoAP1 confers broad-spectrum resistance to Magnaporthe oryzae. Front. Plant Sci. 2019, 10, 433. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wu, Y.; Liu, Z.; Zhang, C. The function and transcriptome analysis of a bZIP transcription factor CgAP1 in Colletotrichum gloeosporioides. Microbiol. Res. 2017, 197, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.L.; Xiong, D.G.; Tian, L.Y.; Tang, C.; Wang, Y.L.; Tian, C.M. Functional characterization of two bZIP transcription factors in Verticillium dahliae. Gene 2017, 626, 386–394. [Google Scholar] [CrossRef] [PubMed]
- Takada, H.; Nishida, A.; Domae, M.; Kita, A.; Yamano, Y.; Uchida, A.; Ishiwata, S.; Fang, Y.; Zhou, X.; Masuko, T.; et al. The cell surface protein gene ecm33+ is a target of the two transcription factors Atf1 and Mbx1 and negatively regulates Pmk1 MAPK cell integrity signaling in fission yeast. Mol. Biol. Cell 2010, 21, 674–685. [Google Scholar] [CrossRef] [Green Version]
- Soanes, D.M.; Chakrabarti, A.; Paszkiewicz, K.H.; Dawe, A.L.; Talbot, N.J. Genome-wide transcriptional profiling of appressorium development by the rice blast fungus Magnaporthe oryzae. PLoS Pathog. 2012, 8, e1002514. [Google Scholar] [CrossRef] [Green Version]
- Keller, N.P. Fungal secondary metabolism: Regulation, function and drug discovery. Nat. Rev. Microbiol. 2019, 17, 167–180. [Google Scholar] [CrossRef]
- Xu, D.; Xue, M.; Shen, Z.; Jia, X.; Hou, X.; Lai, D.; Zhou, L. Phytotoxic secondary metabolites from fungi. Toxins 2021, 13, 261. [Google Scholar] [CrossRef]
- Macheleidt, J.; Mattern, D.J.; Fischer, J.; Netzker, T.; Weber, J.; Schroeckh, V.; Valiante, V.; Brakhage, A.A. Regulation and role of fungal secondary metabolites. Annu. Rev. Genet. 2016, 50, 371–392. [Google Scholar] [CrossRef] [PubMed]
- Guan, X.; Zhao, Y.; Liu, X.; Shang, B.; Xing, F.; Zhou, L.; Wang, Y.; Zhang, C.; Bhatnagar, D.; Liu, Y. The bZIP transcription factor Afap1 mediates the oxidative stress response and aflatoxin biosynthesis in Aspergillus flavus. Rev. Argent. Microbiol. 2019, 51, 292–301. [Google Scholar] [CrossRef]
- Yin, W.B.; Amaike, S.; Wohlbach, D.J.; Gasch, A.P.; Chiang, Y.M.; Wang, C.C.; Bok, J.W.; Rohlfs, M.; Keller, N.P. An Aspergillus nidulans bZIP response pathway hardwired for defensive secondary metabolism operates through aflR. Mol. Microbiol. 2012, 83, 1024–1034. [Google Scholar] [CrossRef] [Green Version]
- Kong, L.A.; Yang, J.; Li, G.T.; Qi, L.L.; Zhang, Y.J.; Wang, C.F.; Zhao, W.S.; Xu, J.R.; Peng, Y.L. Different chitin synthase genes are required for various developmental and plant infection processes in the rice blast fungus Magnaporthe oryzae. PLoS Pathog. 2012, 8, e1002526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Zhang, X.; Liu, X.; Fu, C.; Han, X.; Yin, Y.; Ma, Z. The chitin synthase FgChs2 and other FgChss co-regulate vegetative development and virulence in F. graminearum. Sci. Rep. 2016, 6, 34975. [Google Scholar] [CrossRef] [Green Version]
- Riquelme, M.; Bartnicki-García, S.; González-Prieto, J.M.; Sánchez-León, E.; Verdín-Ramos, J.A.; Beltrán-Aguilar, A.; Freitag, M. Spitzenkorper localization and intracellular traffic of green fluorescent protein-labeled CHS-3 and CHS-6 chitin synthases in living hyphae of Neurospora crassa. Eukaryot. Cell 2007, 6, 1853–1864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, M.; Ngou, B.P.M.; Ding, P.; Xin, X.F. PTI-ETI crosstalk: An integrative view of plant immunity. Curr. Opin. Plant Biol. 2021, 62, 102030. [Google Scholar] [CrossRef] [PubMed]
- Michielse, C.B.; van Wijk, R.; Reijnen, L.; Manders, E.M.; Boas, S.; Olivain, C.; Alabouvette, C.; Rep, M. The nuclear protein Sge1 of Fusarium oxysporum is required for parasitic growth. PLoS Pathog. 2009, 5, e1000637. [Google Scholar] [CrossRef] [Green Version]
- Santhanam, P.; Thomma, B.P. Verticillium dahliae Sge1 differentially regulates expression of candidate effector genes. Mol. Plant-Microbe Interact. 2013, 26, 249–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gui, Y.J.; Chen, J.Y.; Zhang, D.D.; Li, N.Y.; Li, T.G.; Zhang, W.Q.; Wang, X.Y.; Short, D.P.G.; Li, L.; Guo, W.; et al. Verticillium dahliae manipulates plant immunity by glycoside hydrolase 12 proteins in conjunction with carbohydrate-binding module 1. Environ. Microbiol. 2017, 19, 1914–1932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanneganti, T.D.; Huitema, E.; Cakir, C.; Kamoun, S. Synergistic interactions of the plant cell death pathways induced by Phytophthora infestans Nepl-like protein PiNPP1.1 and INF1 elicitin. Mol. Plant-Microbe Interact. 2006, 19, 854–863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nazar Pour, F.; Cobos, R.; Rubio Coque, J.J.; Serôdio, J.; Alves, A.; Félix, C.; Ferreira, V.; Esteves, A.C.; Duarte, A.S. Toxicity of Recombinant Necrosis and Ethylene-Inducing Proteins (NLPs) from Neofusicoccum parvum. Toxins 2020, 12, 235. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CcbZIP Number | Locus Number | gDNA(bp) | cDNA(bp) | Intron | Protein Length (aa) | bZIP Domain Position | MW (kDa) | pI | Location on Supercontig |
|---|---|---|---|---|---|---|---|---|---|
| Ccbzip01 | GME965_g | 2143 | 930 | 2 | 309 | 132–199 | 32.57 | 6.097 | Scaffold1:3424294:3426436: − |
| Ccbzip02 | GME1427_g | 1444 | 1224 | 2 | 407 | 65–127 | 43.65 | 5.919 | Scaffold1:5014645:5016088: − |
| Ccbzip03 | GME1822_g | 1723 | 1551 | 2 | 516 | 401–466 | 54.55 | 6.532 | Scaffold1:6461072:6462794: + |
| Ccbzip04 | GME2112_g | 2393 | 684 | 2 | 244 | 30–91 | 26.88 | 8.302 | Scaffold1:7520918:7523310: + |
| Ccbzip05 | GME2174_g | 800 | 632 | 2 | 210 | 101–165 | 23.16 | 5.537 | Scaffold1:7806590:7807389: + |
| Ccbzip06 | GME4009_g | 1081 | 1029 | 1 | 342 | 162–233 | 37.89 | 5.715 | Scaffold4:2286665:2287745: − |
| Ccbzip07 | GME4152_g | 1881 | 1881 | 0 | 626 | 40–108 | 66.21 | 7.38 | Scaffold4:2835713:2837593: − |
| Ccbzip08 | GME4174_g | 569 | 492 | 1 | 163 | 41–105 | 18.24 | 5.664 | Scaffold4:2902921:2903489: + |
| Ccbzip09 | GME4540_g | 1044 | 1044 | 0 | 347 | 232–296 | 37.37 | 6.81 | Scaffold7:356347:357390: + |
| Ccbzip10 | GME4781_g | 1738 | 1389 | 3 | 462 | 76–137 | 49.77 | 7.161 | Scaffold3:232131:233868: + |
| Ccbzip11 | GME4894_g | 1953 | 1893 | 1 | 630 | 259–323 | 69.87 | 4.991 | Scaffold3:653712:655664: + |
| Ccbzip12 | GME5553_g | 1038 | 735 | 1 | 227 | 54–113 | 24.96 | 4.635 | Scaffold3:2936931:2937968: − |
| Ccbzip13 | GME6051_g | 1884 | 924 | 2 | 307 | 234–299 | 33.9 | 4.825 | Scaffold5:131906:133789: + |
| Ccbzip14 | GME6598_g | 1800 | 1800 | 0 | 599 | 349–412 | 63.93 | 11.082 | Scaffold5:2160568:2162367: − |
| Ccbzip15 | GME7181_g | 1654 | 1509 | 2 | 502 | 94–158 | 54.69 | 8.404 | Scaffold6:723065:724718: − |
| Ccbzip16 | GME7284_g | 1281 | 963 | 2 | 320 | 133–201 | 35.05 | 5.258 | Scaffold6:1084879:1086159: + |
| Ccbzip17 | GME8123_g | 771 | 771 | 0 | 256 | 160–221 | 28.21 | 5.995 | Scaffold2:1195066:1195836: + |
| Ccbzip18 | GME8194_g | 1866 | 1794 | 1 | 597 | 293–358 | 65.51 | 4.698 | Scaffold2:1441851:1443716: + |
| Ccbzip19 | GME8362_g | 1835 | 1683 | 1 | 560 | 85–151 | 61.53 | 6.693 | Scaffold2:2031527:2033361: + |
| Ccbzip20 | GME8516_g | 1635 | 852 | 1 | 283 | 174–235 | 31.53 | 5.486 | Scaffold2:2636438:2638072: − |
| Ccbzip21 | GME9378_g | 1755 | 1755 | 0 | 584 | 448–514 | 64.68 | 6.148 | Scaffold2:5864980:5866734: − |
| Ccbzip22 | GME9737_g | 1192 | 1041 | 2 | 346 | 294–341 | 37.37 | 5.741 | Scaffold2:7266318:7267509: − |
| Ccbzip23 | GME9746_g | 1149 | 858 | 2 | 285 | 63–108 | 31.3 | 7.702 | Scaffold2:7310294:7311442: − |
| Ccbzip24 | GME9819_g | 2194 | 1902 | 2 | 633 | 79–138 | 69.52 | 5.308 | Scaffold2:7606002:7608197: + |
| Ccbzip25 | GME9912_g | 2022 | 1800 | 3 | 599 | 149–214 | 64.89 | 4.863 | Scaffold2:7925885:7927906: + |
| Ccbzip26 | GME10373_g | 1438 | 960 | 3 | 319 | 8–61 | 35.7 | 6.338 | Scaffold2:9540692:9542129: − |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wen, D.; Yu, L.; Xiong, D.; Tian, C. Genome-Wide Identification of bZIP Transcription Factor Genes and Functional Analyses of Two Members in Cytospora chrysosperma. J. Fungi 2022, 8, 34. https://doi.org/10.3390/jof8010034
Wen D, Yu L, Xiong D, Tian C. Genome-Wide Identification of bZIP Transcription Factor Genes and Functional Analyses of Two Members in Cytospora chrysosperma. Journal of Fungi. 2022; 8(1):34. https://doi.org/10.3390/jof8010034
Chicago/Turabian StyleWen, Dasen, Lu Yu, Dianguang Xiong, and Chengming Tian. 2022. "Genome-Wide Identification of bZIP Transcription Factor Genes and Functional Analyses of Two Members in Cytospora chrysosperma" Journal of Fungi 8, no. 1: 34. https://doi.org/10.3390/jof8010034
APA StyleWen, D., Yu, L., Xiong, D., & Tian, C. (2022). Genome-Wide Identification of bZIP Transcription Factor Genes and Functional Analyses of Two Members in Cytospora chrysosperma. Journal of Fungi, 8(1), 34. https://doi.org/10.3390/jof8010034