
Development of a Simple and Robust Whole Blood Assay with Dual Co-Stimulation to Quantify the Release of T-Cellular Signature Cytokines in Response to Aspergillus fumigatus Antigens


 , , ,
, , , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Blood Collection
2.2. Generation of Aspergillus fumigatus mycelial lysate (AfuLy)
2.3. Generation of Recombinant Aspf4
2.4. Quality Control of Lysates and Aspf4 Antigen
2.5. Preparation of WB Stimulation Tubes
2.6. WB Stimulation and Quantification of Cytokine Concentrations in Plasma Supernatants
2.7. T-Cell Depletion of WB Samples
2.8. Flow Cytometry
2.9. Statistics
3. Results
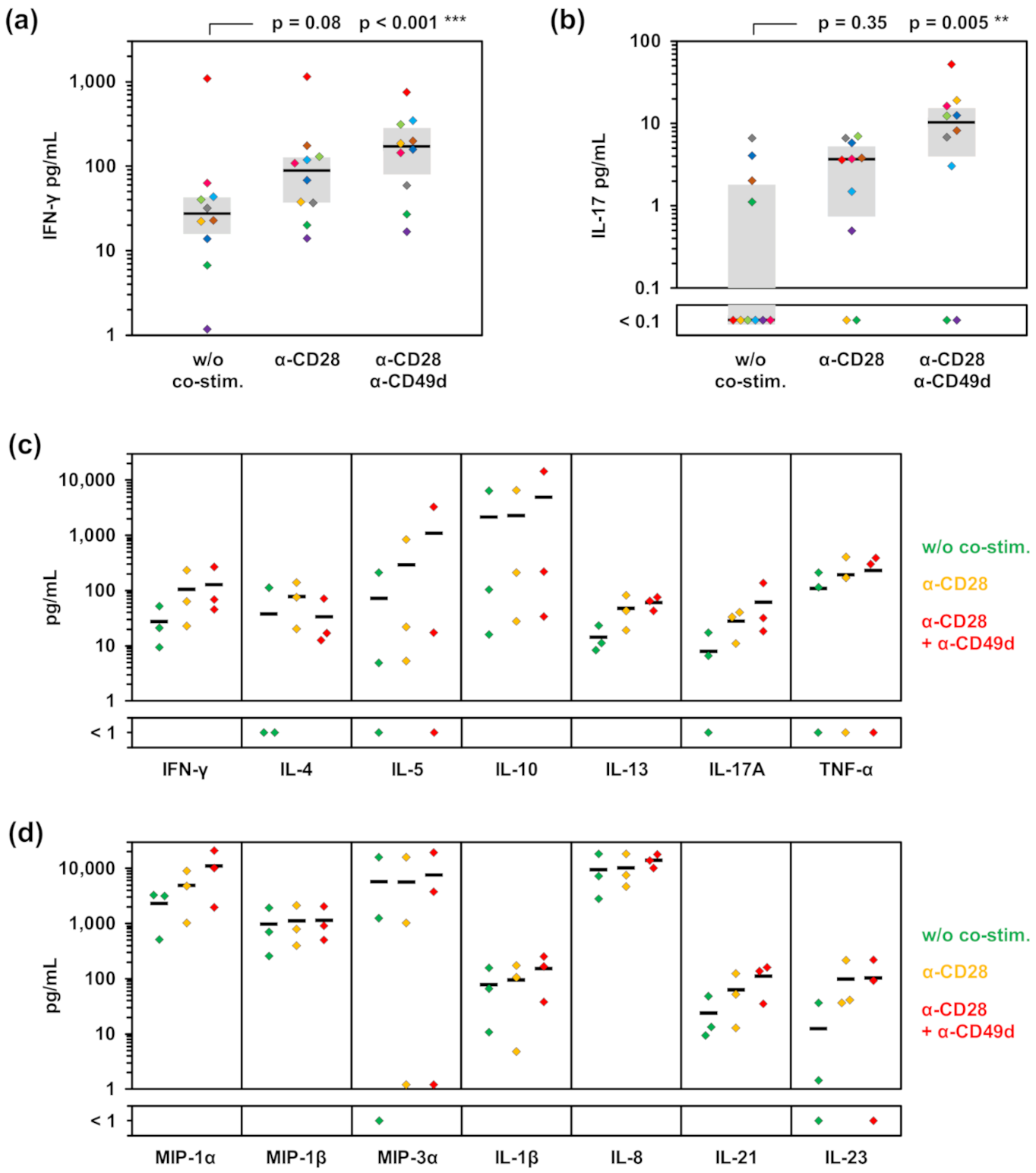
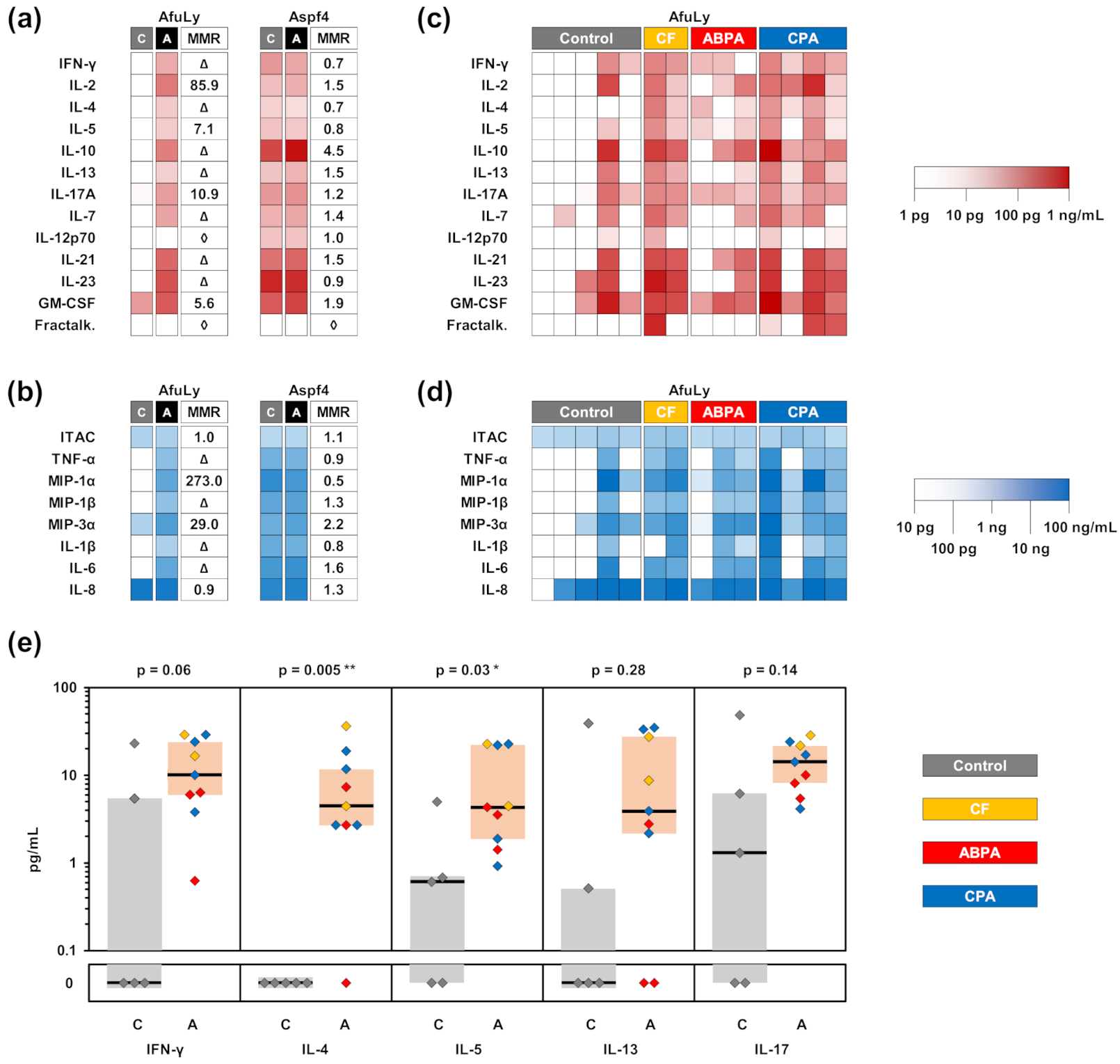
3.1. Impact of Co-Stimulatory Factors on Aspergillus-Induced Cytokine Release
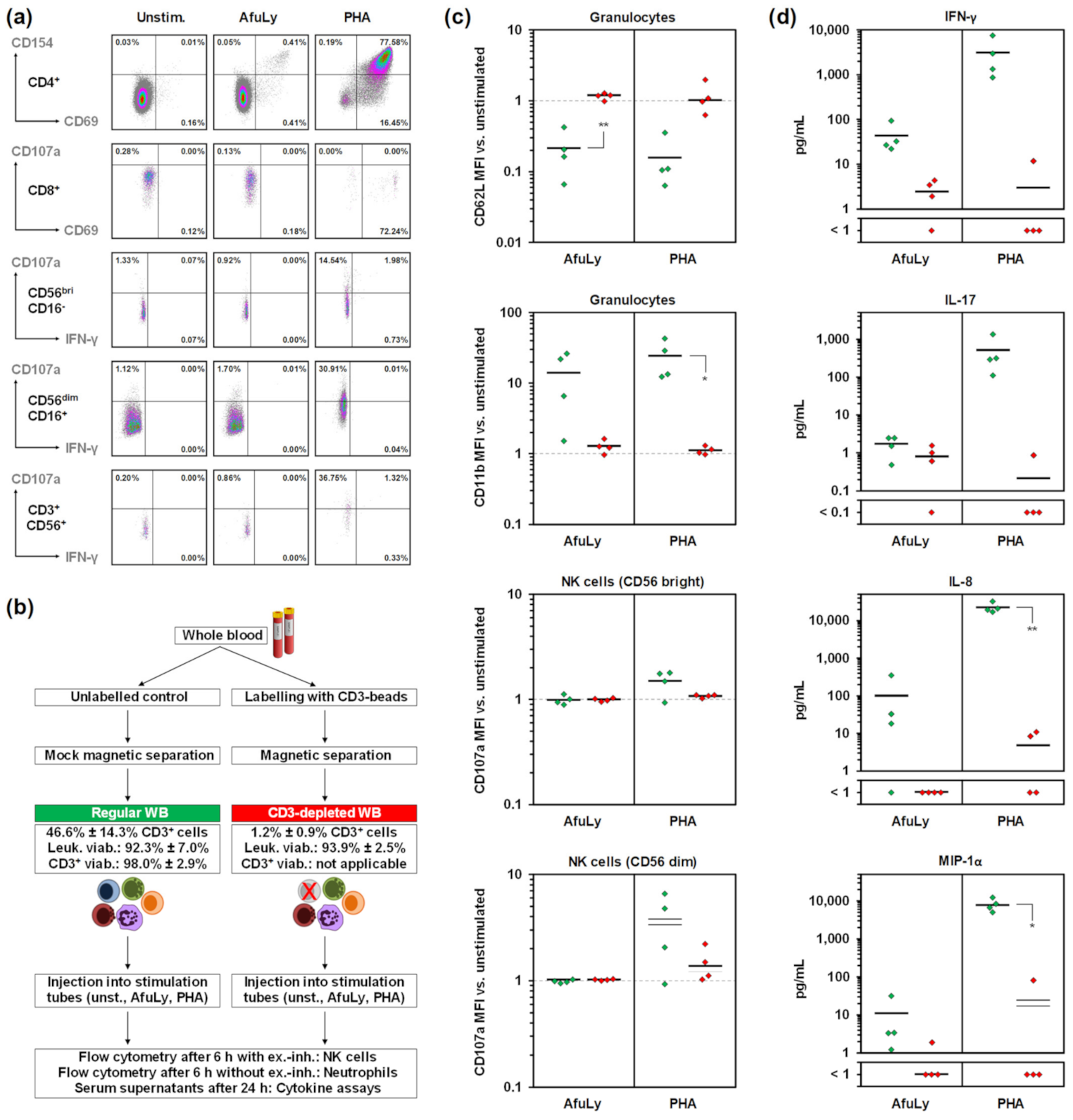
3.2. T-Cell Dependency of Immune Cell Stimulation
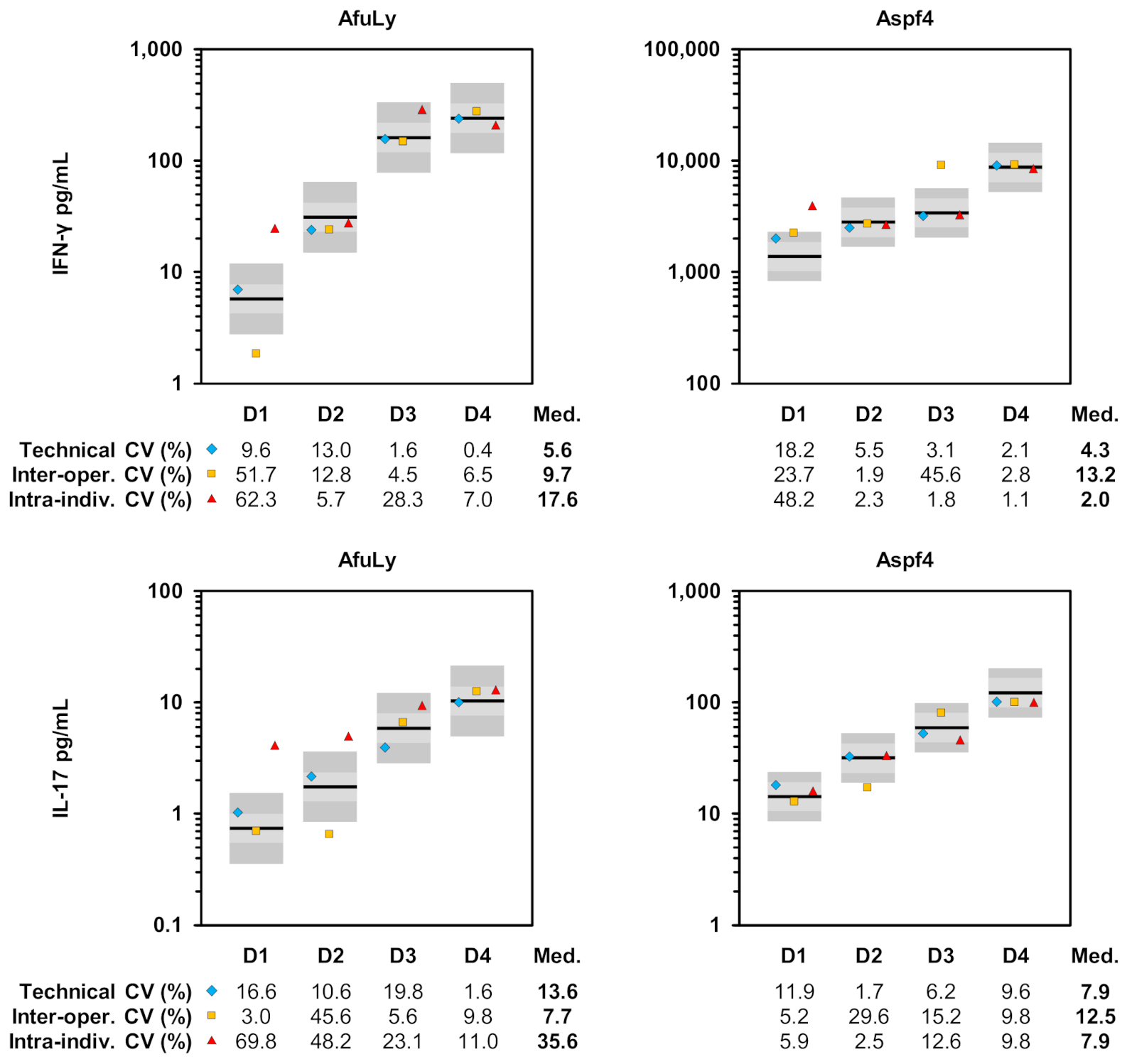
3.3. Evaluation of Assay Reproducibility
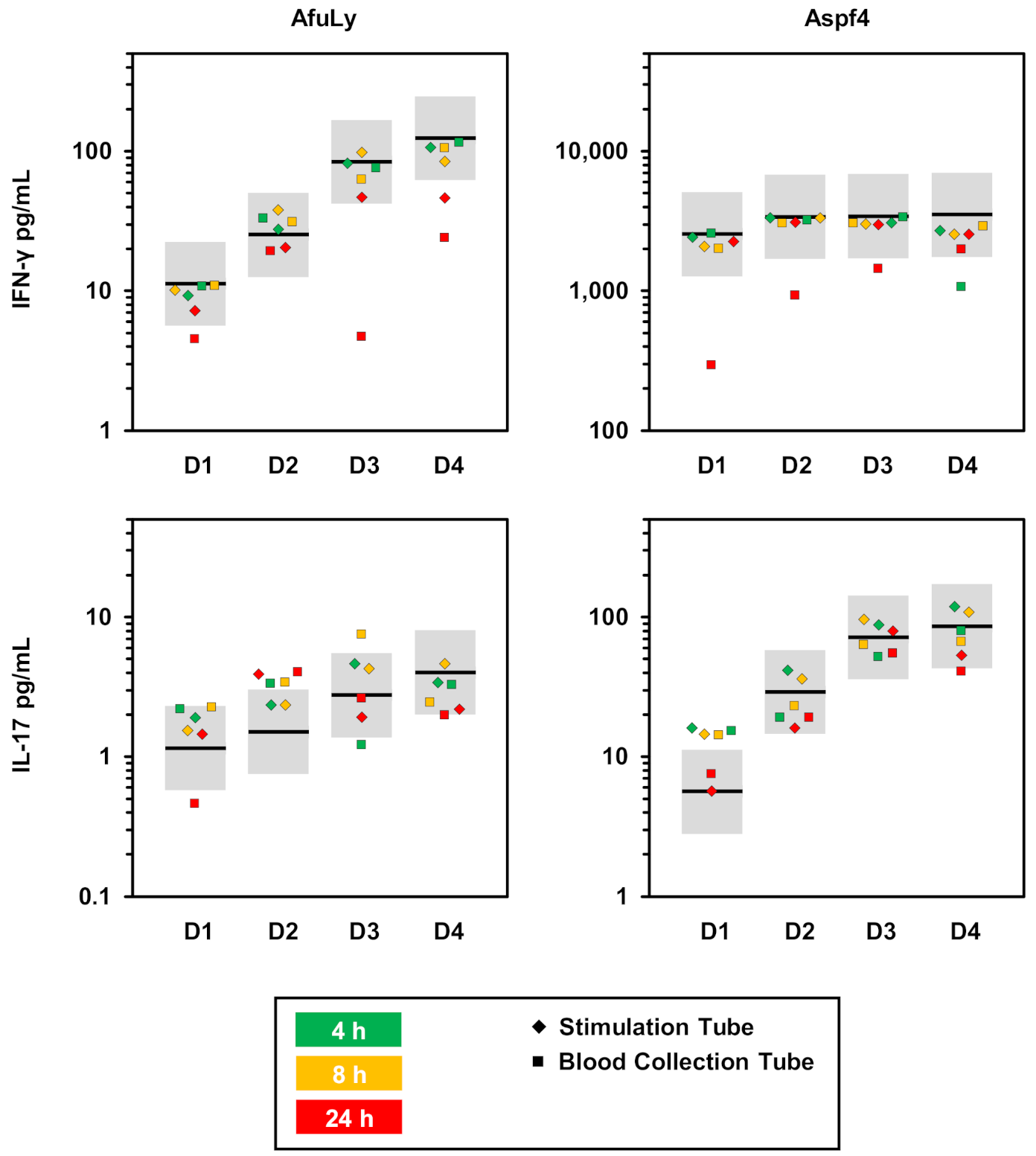
3.4. Susceptibility to Pre-Analytic Delays
3.5. Application of WB-ELISA in Patients with Aspergillus Sensitization
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dewi, I.; van de Veerdonk, F.; Gresnigt, M. The Multifaceted Role of T-Helper Responses in Host Defense against Aspergillus fumigatus. J. Fungi 2017, 3, 55. [Google Scholar] [CrossRef]
- Park, S.J.; Hughes, M.A.; Burdick, M.; Strieter, R.M.; Mehrad, B. Early NK cell-derived IFN-{gamma} is essential to host defense in neutropenic invasive aspergillosis. J. Immunol. 2009, 182, 4306–4312. [Google Scholar] [CrossRef] [PubMed]
- Wurster, S.; Weis, P.; Page, L.; Helm, J.; Lazariotou, M.; Einsele, H.; Ullmann, A.J. Intra- and inter-individual variability of Aspergillus fumigatus reactive T-cell frequencies in healthy volunteers in dependency of mould exposure in residential and working environment. Mycoses 2017, 60, 668–675. [Google Scholar] [CrossRef]
- Jolink, H.; Hagedoorn, R.S.; Lagendijk, E.L.; Drijfhout, J.W.; van Dissel, J.T.; Falkenburg, J.H.; Heemskerk, M.H. Induction of A. fumigatus-specific CD4-positive T cells in patients recovering from invasive aspergillosis. Haematologica 2014, 99, 1255–1263. [Google Scholar] [CrossRef] [PubMed]
- Lordan, J.L.; Bucchieri, F.; Richter, A.; Konstantinidis, A.; Holloway, J.W.; Thornber, M.; Puddicombe, S.M.; Buchanan, D.; Wilson, S.J.; Djukanovic, R.; et al. Cooperative effects of Th2 cytokines and allergen on normal and asthmatic bronchial epithelial cells. J. Immunol. 2002, 169, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Zelante, T.; De Luca, A.; Bonifazi, P.; Montagnoli, C.; Bozza, S.; Moretti, S.; Belladonna, M.L.; Vacca, C.; Conte, C.; Mosci, P.; et al. IL-23 and the Th17 pathway promote inflammation and impair antifungal immune resistance. Eur. J. Immunol. 2007, 37, 2695–2706. [Google Scholar] [CrossRef]
- Bacher, P.; Steinbach, A.; Kniemeyer, O.; Hamprecht, A.; Assenmacher, M.; Vehreschild, M.J.G.T.; Vehreschild, J.J.; Brakhage, A.A.; Cornely, O.A.; Scheffold, A. Fungus-Specific CD4+ T Cells for Rapid Identification of Invasive Pulmonary Mold Infection. Am. J. Respir. Crit. Care Med. 2015, 191, 348–352. [Google Scholar] [CrossRef]
- Koehler, F.C.; Cornely, O.A.; Wisplinghoff, H.; Schauss, A.C.; Salmanton-Garcia, J.; Ostermann, H.; Ziegler, M.; Bacher, P.; Scheffold, A.; Alex, R.; et al. Candida-Reactive T Cells for the Diagnosis of Invasive Candida Infection-A Prospective Pilot Study. Front. Microbiol. 2018, 9, 1381. [Google Scholar] [CrossRef]
- Potenza, L.; Vallerini, D.; Barozzi, P.; Riva, G.; Forghieri, F.; Beauvais, A.; Beau, R.; Candoni, A.; Maertens, J.; Rossi, G.; et al. Characterization of specific immune responses to different Aspergillus antigens during the course of invasive Aspergillosis in hematologic patients. PLoS ONE 2013, 8, e74326. [Google Scholar] [CrossRef]
- Potenza, L.; Vallerini, D.; Barozzi, P.; Riva, G.; Gilioli, A.; Forghieri, F.; Candoni, A.; Cesaro, S.; Quadrelli, C.; Maertens, J.; et al. Mucorales-Specific T Cells in Patients with Hematologic Malignancies. PLoS ONE 2016, 11, e0149108. [Google Scholar] [CrossRef]
- Steinbach, A.; Cornely, O.A.; Wisplinghoff, H.; Schauss, A.C.; Vehreschild, J.J.; Rybniker, J.; Hamprecht, A.; Richter, A.; Bacher, P.; Scheffold, A.; et al. Mould-reactive T cells for the diagnosis of invasive mould infection-A prospective study. Mycoses 2019, 62, 562–569. [Google Scholar] [CrossRef]
- Lauruschkat, C.D.; Wurster, S.; Page, L.; Lazariotou, M.; Dragan, M.; Weis, P.; Ullmann, A.J.; Einsele, H.; Loffler, J. Susceptibility of A. fumigatus-specific T-cell assays to pre-analytic blood storage and PBMC cryopreservation greatly depends on readout platform and analytes. Mycoses 2018, 61, 549–560. [Google Scholar] [CrossRef]
- Wurster, S.; Weis, P.; Page, L.; Lazariotou, M.; Einsele, H.; Ullmann, A.J. Quantification of A. fumigatus-specific CD154+ T-cells-preanalytic considerations. Med. Mycol. 2017, 55, 223–227. [Google Scholar] [CrossRef][Green Version]
- McKenna, K.C.; Beatty, K.M.; Vicetti Miguel, R.; Bilonick, R.A. Delayed processing of blood increases the frequency of activated CD11b+ CD15+ granulocytes which inhibit T cell function. J. Immunol. Methods 2009, 341, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Rubio-Carrasquilla, M.; Santa, C.D.; Rendon, J.P.; Botero-Garces, J.; Guimaraes, A.J.; Moreno, E.; Cano, L.E. An interferon gamma release assay specific for Histoplasma capsulatum to detect asymptomatic infected individuals: A proof of concept study. Med. Mycol. 2019, 57, 724–732. [Google Scholar] [CrossRef]
- Weis, P.; Helm, J.; Page, L.; Lauruschkat, C.D.; Lazariotou, M.; Einsele, H.; Loeffler, J.; Ullmann, A.J.; Wurster, S. Development and evaluation of a whole blood-based approach for flow cytometric quantification of CD154+ mould-reactive T cells. Med. Mycol. 2020, 58, 187–196. [Google Scholar] [CrossRef]
- Douglas, A.P.; Yu, L.; Sundararajan, V.; Szer, J.; Ritchie, D.; Slavin, M.A.; Sasadeusz, J.; Visvanathan, K. The QuantiFERON Monitor((R)) assay is predictive of infection post allogeneic hematopoietic cell transplantation. Transpl. Infect. Dis. 2020, 22, e13260. [Google Scholar] [CrossRef] [PubMed]
- Silveira, M.B.V.; Ferrarini, M.A.G.; Viana, P.O.; Succi, R.C.; Terreri, M.T.; Costa-Carvalho, B.; Carlesse, F.; de Moraes-Pinto, M.I. Contribution of the interferon-gamma release assay to tuberculosis diagnosis in children and adolescents. Int. J. Tuberc. Lung. Dis. 2018, 22, 1172–1178. [Google Scholar] [CrossRef] [PubMed]
- Theel, E.S.; Hilgart, H.; Breen-Lyles, M.; McCoy, K.; Flury, R.; Breeher, L.E.; Wilson, J.; Sia, I.G.; Whitaker, J.A.; Clain, J.; et al. Comparison of the QuantiFERON-TB Gold Plus and QuantiFERON-TB Gold In-Tube Interferon Gamma Release Assays in Patients at Risk for Tuberculosis and in Health Care Workers. J. Clin. Microbiol. 2018, 56. [Google Scholar] [CrossRef]
- Doberne, D.; Gaur, R.L.; Banaei, N. Preanalytical delay reduces sensitivity of QuantiFERON-TB gold in-tube assay for detection of latent tuberculosis infection. J. Clin. Microbiol. 2011, 49, 3061–3064. [Google Scholar] [CrossRef]
- Herrera, V.; Yeh, E.; Murphy, K.; Parsonnet, J.; Banaei, N. Immediate incubation reduces indeterminate results for QuantiFERON-TB Gold in-tube assay. J. Clin. Microbiol. 2010, 48, 2672–2676. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kim, E.Y.; Lim, J.E.; Jung, J.Y.; Son, J.Y.; Lee, K.J.; Yoon, Y.W.; Park, B.H.; Moon, J.W.; Park, M.S.; Kim, Y.S.; et al. Performance of the tuberculin skin test and interferon-gamma release assay for detection of tuberculosis infection in immunocompromised patients in a BCG-vaccinated population. BMC Infect. Dis. 2009, 9, 207. [Google Scholar] [CrossRef] [PubMed]
- Page, L.; Lauruschkat, C.D.; Helm, J.; Weis, P.; Lazariotou, M.; Einsele, H.; Ullmann, A.J.; Loeffler, J.; Wurster, S. Impact of immunosuppressive and antifungal drugs on PBMC- and whole blood-based flow cytometric CD154(+) Aspergillus fumigatus specific T-cell quantification. Med. Microbiol. Immunol. 2020, 209, 579–592. [Google Scholar] [CrossRef] [PubMed]
- Deborska-Materkowska, D.; Perkowska-Ptasinska, A.; Sadowska, A.; Gozdowska, J.; Ciszek, M.; Serwanska-Swietek, M.; Domagala, P.; Miszewska-Szyszkowska, D.; Sitarek, E.; Jozwik, A.; et al. Diagnostic utility of monitoring cytomegalovirus-specific immunity by QuantiFERON-cytomegalovirus assay in kidney transplant recipients. BMC Infect. Dis. 2018, 18, 179. [Google Scholar] [CrossRef]
- El Haddad, L.; Ariza-Heredia, E.; Shah, D.P.; Jiang, Y.; Blanchard, T.; Ghantoji, S.S.; El Chaer, F.; El-Haddad, D.; Prayag, A.; Nesher, L.; et al. The Ability of a Cytomegalovirus ELISPOT Assay to Predict Outcome of Low-Level CMV Reactivation in Hematopoietic Cell Transplant Recipients. J. Infect. Dis. 2019, 219, 898–907. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Li, R.; Shen, J.; He, L.; Li, Y.; Zhang, N.; Wu, Q.; Zhang, J.; Zheng, J.; Wang, X. Clinical effect of T-SPOT.TB test for the diagnosis of tuberculosis. BMC Infect. Dis. 2019, 19, 993. [Google Scholar] [CrossRef] [PubMed]
- Pourakbari, B.; Mamishi, S.; Benvari, S.; Mahmoudi, S. Comparison of the QuantiFERON-TB Gold Plus and QuantiFERON-TB Gold In-Tube interferon-gamma release assays: A systematic review and meta-analysis. Adv. Med. Sci. 2019, 64, 437–443. [Google Scholar] [CrossRef]
- Lemieux, J.; Jobin, C.; Simard, C.; Neron, S. A global look into human T cell subsets before and after cryopreservation using multiparametric flow cytometry and two-dimensional visualization analysis. J. Immunol. Methods 2016, 434, 73–82. [Google Scholar] [CrossRef]
- Thakur, A.; Riber, U.; Davis, W.C.; Jungersen, G. Increasing the ex vivo antigen-specific IFN-gamma production in subpopulations of T cells and NKp46+ cells by anti-CD28, anti-CD49d and recombinant IL-12 costimulation in cattle vaccinated with recombinant proteins from Mycobacterium avium subspecies paratuberculosis. Vet. Immunol. Immunopathol. 2013, 155, 276–283. [Google Scholar] [CrossRef][Green Version]
- Udagawa, T.; Woodside, D.G.; McIntyre, B.W. Alpha 4 beta 1 (CD49d/CD29) integrin costimulation of human T cells enhances transcription factor and cytokine induction in the absence of altered sensitivity to anti-CD3 stimulation. J. Immunol. 1996, 157, 1965–1972. [Google Scholar]
- Brakhage, A.A.; Van den Brulle, J. Use of reporter genes to identify recessive trans-acting mutations specifically involved in the regulation of Aspergillus nidulans penicillin biosynthesis genes. J. Bacteriol. 1995, 177, 2781–2788. [Google Scholar] [CrossRef] [PubMed]
- Selliah, N.; Eck, S.; Green, C.; Oldaker, T.; Stewart, J.; Vitaliti, A.; Litwin, V. Flow Cytometry Method Validation Protocols. Curr. Protoc. Cytom. 2019, 87, e53. [Google Scholar] [CrossRef]
- Page, L.; Weis, P.; Muller, T.; Dittrich, M.; Lazariotou, M.; Dragan, M.; Waaga-Gasser, A.M.; Helm, J.; Dandekar, T.; Einsele, H.; et al. Evaluation of Aspergillus and Mucorales specific T-cells and peripheral blood mononuclear cell cytokine signatures as biomarkers of environmental mold exposure. Int. J. Med. Microbiol. 2018, 308, 1018–1026. [Google Scholar] [CrossRef] [PubMed]
- David, J.M.; Dominguez, C.; Hamilton, D.H.; Palena, C. The IL-8/IL-8R Axis: A Double Agent in Tumor Immune Resistance. Vaccines 2016, 4, 22. [Google Scholar] [CrossRef] [PubMed]
- Lindell, D.M.; Standiford, T.J.; Mancuso, P.; Leshen, Z.J.; Huffnagle, G.B. Macrophage inflammatory protein 1alpha/CCL3 is required for clearance of an acute Klebsiella pneumoniae pulmonary infection. Infect. Immun. 2001, 69, 6364–6369. [Google Scholar] [CrossRef]
- Meniailo, M.E.; Malashchenko, V.V.; Shmarov, V.A.; Gazatova, N.D.; Melashchenko, O.B.; Goncharov, A.G.; Seledtsova, G.V.; Seledtsov, V.I. Interleukin-8 favors pro-inflammatory activity of human monocytes/macrophages. Int. Immunopharmacol. 2018, 56, 217–221. [Google Scholar] [CrossRef]
- Narni-Mancinelli, E.; Campisi, L.; Bassand, D.; Cazareth, J.; Gounon, P.; Glaichenhaus, N.; Lauvau, G. Memory CD8+ T cells mediate antibacterial immunity via CCL3 activation of TNF/ROI+ phagocytes. J. Exp. Med. 2007, 204, 2075–2087. [Google Scholar] [CrossRef] [PubMed]
- Yu, B.; Yang, H. Evaluation of Different Estimation Methods for Accuracy and Precision in Biological Assay Validation. PDA J. Pharm. Sci. Technol. 2017, 71, 297–305. [Google Scholar] [CrossRef]
- U.S. Department of Health and Human Services, Food and Drug Administration. Bioanalytical Method Validation Guidance for Industry. 2018. Available online: https://www.fda.gov/files/drugs/published/Bioanalytical-Method-Validation-Guidance-for-Industry.pdf (accessed on 8 June 2021).
- Chaudhary, N.; Staab, J.F.; Marr, K.A. Healthy human T-Cell Responses to Aspergillus fumigatus antigens. PLoS ONE 2010, 5, e9036. [Google Scholar] [CrossRef]
- Agarwal, R.; Chakrabarti, A.; Shah, A.; Gupta, D.; Meis, J.F.; Guleria, R.; Moss, R.; Denning, D.W.; ABPA complicating asthma ISHAM working group. Allergic bronchopulmonary aspergillosis: Review of literature and proposal of new diagnostic and classification criteria. Clin. Exp. Allergy 2013, 43, 850–873. [Google Scholar] [CrossRef]
- Wang, F.; Zhang, C.; Jiang, Y.; Kou, C.; Kong, Q.; Long, N.; Lu, L.; Sang, H. Innate and adaptive immune response to chronic pulmonary infection of hyphae of Aspergillus fumigatus in a new murine model. J. Med. Microbiol. 2017, 66, 1400–1408. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Biagini Myers, J.M.; Brandt, E.B.; Ryan, P.H.; Lindsey, M.; Mintz-Cole, R.A.; Reponen, T.; Vesper, S.J.; Forde, F.; Ruff, B.; et al. β-Glucan exacerbates allergic asthma independent of fungal sensitization and promotes steroid-resistant TH2/TH17 responses. J. Allergy Clin. Immunol. 2017, 139, 54–65.e8. [Google Scholar] [CrossRef]
- Gour, N.; Wills-Karp, M. IL-4 and IL-13 signaling in allergic airway disease. Cytokine 2015, 75, 68–78. [Google Scholar] [CrossRef]
- Emson, C.; Pham, T.H.; Manetz, S.; Newbold, P. Periostin and Dipeptidyl Peptidase-4: Potential Biomarkers of Interleukin 13 Pathway Activation in Asthma and Allergy. Immunol. Allergy Clin. North. Am. 2018, 38, 611–628. [Google Scholar] [CrossRef]
- Duffy, D.; Rouilly, V.; Libri, V.; Hasan, M.; Beitz, B.; David, M.; Urrutia, A.; Bisiaux, A.; Labrie, S.T.; Dubois, A.; et al. Functional analysis via standardized whole-blood stimulation systems defines the boundaries of a healthy immune response to complex stimuli. Immunity 2014, 40, 436–450. [Google Scholar] [CrossRef] [PubMed]
- Deenadayalan, A.; Maddineni, P.; Raja, A. Comparison of whole blood and PBMC assays for T-cell functional analysis. BMC Res. Notes 2013, 6, 120. [Google Scholar] [CrossRef] [PubMed]
- Lagrelius, M.; Jones, P.; Franck, K.; Gaines, H. Cytokine detection by multiplex technology useful for assessing antigen specific cytokine profiles and kinetics in whole blood cultured up to seven days. Cytokine 2006, 33, 156–165. [Google Scholar] [CrossRef]
- Silva, D.; Ponte, C.G.; Hacker, M.A.; Antas, P.R. A whole blood assay as a simple, broad assessment of cytokines and chemokines to evaluate human immune responses to Mycobacterium tuberculosis antigens. Acta Trop. 2013, 127, 75–81. [Google Scholar] [CrossRef]
- Minns, D.; Smith, K.J.; Findlay, E.G. Orchestration of Adaptive T Cell Responses by Neutrophil Granule Contents. Mediat. Inflamm. 2019, 2019, 8968943. [Google Scholar] [CrossRef]
- Duechting, A.; Przybyla, A.; Kuerten, S.; Lehmann, P.V. Delayed Activation Kinetics of Th2- and Th17 Cells Compared to Th1 Cells. Cells 2017, 6, 29. [Google Scholar] [CrossRef]
- Esensten, J.H.; Helou, Y.A.; Chopra, G.; Weiss, A.; Bluestone, J.A. CD28 Costimulation: From Mechanism to Therapy. Immunity 2016, 44, 973–988. [Google Scholar] [CrossRef] [PubMed]
- Gauduin, M.C. Intracellular cytokine staining for the characterization and quantitation of antigen-specific T lymphocyte responses. Methods 2006, 38, 263–273. [Google Scholar] [CrossRef] [PubMed]
- Waldrop, S.L.; Davis, K.A.; Maino, V.C.; Picker, L.J. Normal human CD4+ memory T cells display broad heterogeneity in their activation threshold for cytokine synthesis. J. Immunol. 1998, 161, 5284–5295. [Google Scholar] [PubMed]
- BD. BD FastImmune™CD28/CD49d. Available online: https://www.bdbiosciences.com/ds/pm/tds/347690.pdf (accessed on 8 June 2021).
- Suni, M.A.; Picker, L.J.; Maino, V.C. Detection of antigen-specific T cell cytokine expression in whole blood by flow cytometry. J. Immunol. Methods 1998, 212, 89–98. [Google Scholar] [CrossRef]
- Lauruschkat, C.D.; Page, L.; Etter, S.; Weis, P.; Gamon, F.; Kraus, S.; Einsele, H.; Wurster, S.; Loeffler, J. T-Cell Immune Surveillance in Allogenic Stem Cell Transplant Recipients: Are Whole Blood-Based Assays Ready to Challenge ELISPOT? Open Forum Infect. Dis. 2021, 8, ofaa547. [Google Scholar] [CrossRef] [PubMed]
- Bacher, P.; Hohnstein, T.; Beerbaum, E.; Rocker, M.; Blango, M.G.; Kaufmann, S.; Rohmel, J.; Eschenhagen, P.; Grehn, C.; Seidel, K.; et al. Human Anti-fungal Th17 Immunity and Pathology Rely on Cross-Reactivity against Candida albicans. Cell 2019, 176, 1340–1355.e15. [Google Scholar] [CrossRef]
- Iwanaga, N.; Kolls, J.K. Updates on T helper type 17 immunity in respiratory disease. Immunology 2019, 156, 3–8. [Google Scholar] [CrossRef]
- Zelante, T.; Bozza, S.; De Luca, A.; D’Angelo, C.; Bonifazi, P.; Moretti, S.; Giovannini, G.; Bistoni, F.; Romani, L. Th17 cells in the setting of Aspergillus infection and pathology. Med. Mycol. 2009, 47 (Suppl. 1), S162–S169. [Google Scholar] [CrossRef]
- Vogel, K.; Pierau, M.; Arra, A.; Lampe, K.; Schlueter, D.; Arens, C.; Brunner-Weinzierl, M.C. Developmental induction of human T-cell responses against Candida albicans and Aspergillus fumigatus. Sci. Rep. 2018, 8, 16904. [Google Scholar] [CrossRef]
- Murdock, B.J.; Shreiner, A.B.; McDonald, R.A.; Osterholzer, J.J.; White, E.S.; Toews, G.B.; Huffnagle, G.B. Coevolution of TH1, TH2, and TH17 responses during repeated pulmonary exposure to Aspergillus fumigatus conidia. Infect. Immun. 2011, 79, 125–135. [Google Scholar] [CrossRef]
- Becker, K.L.; Gresnigt, M.S.; Smeekens, S.P.; Jacobs, C.W.; Magis-Escurra, C.; Jaeger, M.; Wang, X.; Lubbers, R.; Oosting, M.; Joosten, L.A.; et al. Pattern recognition pathways leading to a Th2 cytokine bias in allergic bronchopulmonary aspergillosis patients. Clin. Exp. Allergy 2015, 45, 423–437. [Google Scholar] [CrossRef] [PubMed]
- Knutsen, A.P.; Slavin, R.G. Allergic bronchopulmonary aspergillosis in asthma and cystic fibrosis. Clin. Dev. Immunol. 2011, 2011, 843763. [Google Scholar] [CrossRef] [PubMed]
- Bacher, P.; Kniemeyer, O.; Schonbrunn, A.; Sawitzki, B.; Assenmacher, M.; Rietschel, E.; Steinbach, A.; Cornely, O.A.; Brakhage, A.A.; Thiel, A.; et al. Antigen-specific expansion of human regulatory T cells as a major tolerance mechanism against mucosal fungi. Mucosal Immunol. 2014, 7, 916–928. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| α-CD28 | α-CD49d | AfuLy | Aspf4 | PHA | RPMI | |
|---|---|---|---|---|---|---|
| Concentration in the ready-to-use stimulation tube | 2 µg/mL | 2 µg/mL | 100 µg/mL | 60 µg/mL | 10 µg/mL | ad 500 µL |
| Final concentration afterinjection of 500 µL WB | 1 µg/mL | 1 µg/mL | 50 µg/mL | 30 µg/mL | 5 µg/mL | n/a |
| Unstimulated control | X | X | X | |||
| AfuLy stimulation | X | X | X | X | ||
| Aspf4 protein stimulation | X | X | X | X | ||
| Positive control | X | X |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lauruschkat, C.D.; Page, L.; White, P.L.; Etter, S.; Davies, H.E.; Duckers, J.; Ebel, F.; Schnack, E.; Backx, M.; Dragan, M.; et al. Development of a Simple and Robust Whole Blood Assay with Dual Co-Stimulation to Quantify the Release of T-Cellular Signature Cytokines in Response to Aspergillus fumigatus Antigens. J. Fungi 2021, 7, 462. https://doi.org/10.3390/jof7060462
Lauruschkat CD, Page L, White PL, Etter S, Davies HE, Duckers J, Ebel F, Schnack E, Backx M, Dragan M, et al. Development of a Simple and Robust Whole Blood Assay with Dual Co-Stimulation to Quantify the Release of T-Cellular Signature Cytokines in Response to Aspergillus fumigatus Antigens. Journal of Fungi. 2021; 7(6):462. https://doi.org/10.3390/jof7060462
Chicago/Turabian StyleLauruschkat, Chris D., Lukas Page, P. Lewis White, Sonja Etter, Helen E. Davies, Jamie Duckers, Frank Ebel, Elisabeth Schnack, Matthijs Backx, Mariola Dragan, and et al. 2021. "Development of a Simple and Robust Whole Blood Assay with Dual Co-Stimulation to Quantify the Release of T-Cellular Signature Cytokines in Response to Aspergillus fumigatus Antigens" Journal of Fungi 7, no. 6: 462. https://doi.org/10.3390/jof7060462
APA StyleLauruschkat, C. D., Page, L., White, P. L., Etter, S., Davies, H. E., Duckers, J., Ebel, F., Schnack, E., Backx, M., Dragan, M., Schlegel, N., Kniemeyer, O., Brakhage, A. A., Einsele, H., Loeffler, J., & Wurster, S. (2021). Development of a Simple and Robust Whole Blood Assay with Dual Co-Stimulation to Quantify the Release of T-Cellular Signature Cytokines in Response to Aspergillus fumigatus Antigens. Journal of Fungi, 7(6), 462. https://doi.org/10.3390/jof7060462