
Beauveria bassiana Endophytic Strain as Plant Growth Promoter: The Case of the Grape Vine Vitis vinifera

 ,
, 
 and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material, Fungus, Inst and Experimental Design
2.2. Endophytic re Isolation from Plant Tissues
2.3. Effect of Endophytic B. bassiana on V. vinifera Plants
2.4. Fungi Re-Isolation from the Soil
2.5. Fungi Identification Methods
2.6. Statistical Analysis
3. Results
3.1. Molecular and Microscopic Identification of the Fungi
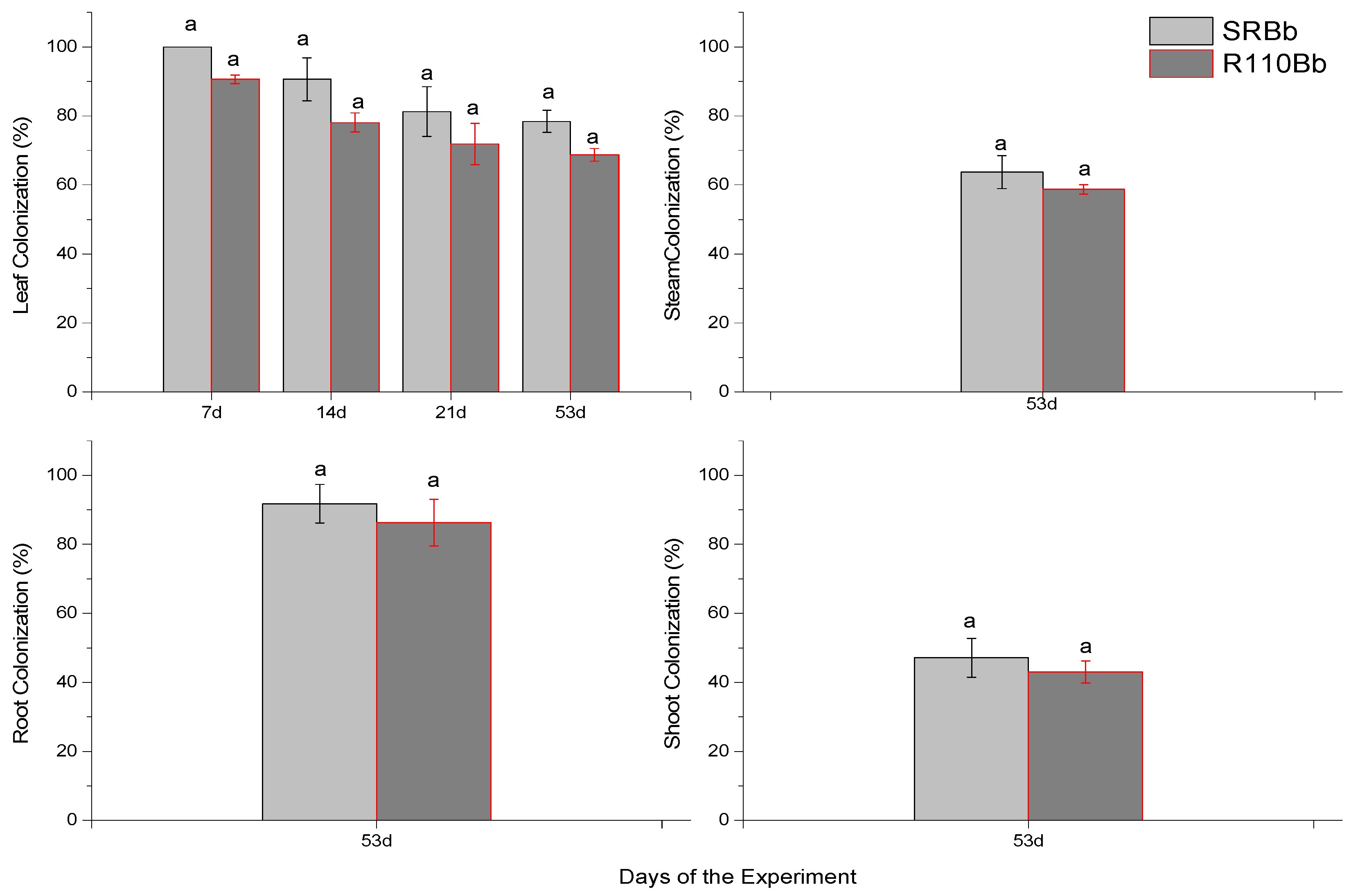
3.2. Re-Isolation of Entomopathogenic Fungi from Leaves, Shoots, Stem and Roots on SDA Substrate
3.3. Growth and Mass Parameters
3.4. Fungus Soil Re-Isolation with T. confusum Bait
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nassiri-Asl, M.; Hosseinzadeh, H.I. Review of the Pharmacological Effects of Vitis vinifera (Grape) and its Bioactive Compounds. Phyther. Res. 2009, 23, 1197–1204. [Google Scholar] [CrossRef]
- Tabeshpour, J.; Mehri, S.; Shaebani Behbahani, F.; Hosseinzadeh, H. Protective effects of Vitis vinifera (grapes) and one of its biologically active constituents, resveratrol, against natural and chemical toxicities: A comprehensive review. Phyther. Res. 2018, 32, 2164–2190. [Google Scholar] [CrossRef] [PubMed]
- Fleet, G.H. Yeast interactions and wine flavour. Int. J. Food. Microbiol. 2003, 86, 11–22. [Google Scholar] [CrossRef]
- Strobel, G.; Daisy, B. Bioprospecting for Microbial Endophytes and Their Natural Products. Microbiol. Mol. Biol. Rev. 2003, 67, 491–502. [Google Scholar] [CrossRef]
- Strobel, G.A. Symposium contribution Microbial gifts from rain forests. Can. J. Plant Pathol. 2002, 24, 14–20. [Google Scholar] [CrossRef]
- Polishook, J.D.; Dombrowski, A.W.; Tsou, N.N.; Salituro, G.M.; Curotto, J.E. Preussomerin D from the Endophyte Hormonema Dematioides. Mycologia 1993, 85, 62–64. [Google Scholar] [CrossRef]
- Pelaez, F.; Cabello, A.; Platas, G.; Díez, M.T.; González del Val, A.; Basilio, A.; Martán, I.; Vicente, F.; Bills, G.E.; Giacobbe, R.A.; et al. The discovery of enfumafungin, a novel antifungal compound produced by an endophytic Hormonema species biological activity and taxonomy of the producing organisms. Syst. Appl. Microbiol. 2000, 23, 333–343. [Google Scholar] [CrossRef]
- Aly, A.H.; Debbab, A.; Kjer, J.; Proksch, P. Fungal endophytes from higher plants: A prolific source of phytochemicals and other bioactive natural products. Fungal Divers. 2010, 41, 1–16. [Google Scholar] [CrossRef]
- Lin, X.; Huang, Y.J.; Zheng, Z.H.; Su, W.J.; Qian, X.M.; Shen, Y.M. Endophytes from the pharmaceutical plant, Annona squamosa: Isolation, bioactivity, identification and diversity of its polyketide synthase gene. Fungal Divers. 2010, 41, 41–51. [Google Scholar] [CrossRef]
- Bamisile, B.S.; Dash, C.K.; Akutse, K.S.; Qasim, M.; Ramos Aguila, L.C.; Wang, F.; Keppanan, R.; Wang, L. Endophytic Beauveria bassiana in foliar-treated citrus limon plants acting as a growth suppressor to three successive generations of diaphorina citri kuwayama (Hemiptera: Liviidae). Insects 2019, 10, 176. [Google Scholar] [CrossRef]
- Liu, J.; Liu, G. Analysis of sondary metabolites from plant endophytic fungi. Methods Mol. Biol. 2018, 1848, 25–38. [Google Scholar]
- Gómez, O.C.; Luiz, J.H.H. Endophytic fungi isolated from medicinal plants: Future prospects of bioactive natural products from Tabebuia/Handroanthus endophytes. Appl. Microbiol. Biotechnol. 2018, 102, 9105–9119. [Google Scholar] [CrossRef]
- Vidal, S.; Jaber, L.R. Entomopathogenic fungi as endophytes: Plant-endophyte-herbivore interactions and prospects for use in biological control. Curr. Sci. 2015, 109, 46–54. [Google Scholar]
- Jaber, L.R.; Ownley, B.H. Can we use entomopathogenic fungi as endophytes for dual biological control of inst pests and plant pathogens? Biol. Control 2018, 116, 36–45. [Google Scholar] [CrossRef]
- Moonjely, S.; Barelli, L.; Bidochka, M.J. Inst Pathogenic Fungi as Endophytes. Adv. Genet. 2016, 94, 107–135. [Google Scholar]
- Tan, R.X.; Zou, W.X. Endophytes: A rich source of functional metabolites. Nat. Prod. Rep. 2001, 18, 448–459. [Google Scholar] [CrossRef]
- Huang, W.; Cai, Y.; Surveswaran, S.; Hyde, K.D.; Corke, H.; Sun, M. Molecular phylogenetic identification of endophytic fungi isolated from three Artemisia species. Fungal Divers. 2009, 36, 69–88. [Google Scholar]
- Arnold, A.E.; Mejía, L.C.; Kyllo, D.; Rojas, E.I.; Maynard, Z.; Robbins, N.; Herre, E.A. Fungal endophytes limit pathogen damage in a tropical tree. Proc. Natl. Acad. Sci. USA 2003, 100, 15649–15654. [Google Scholar] [CrossRef]
- Hallman, J.; Quadt-Hallman, A.; Mahafee, W.F.; Kloepper, J.W. Bacterial endophytes in agricultural crops. Can. J. Microbiol. 1997, 43, 895–914. [Google Scholar] [CrossRef]
- Sturz, A.V.; Nowak, J. Endophytic communities of rhizobacteria and the strategies required to create yield enhancing associations with crops. Appl. Soil Ecol. 2000, 15, 183–190. [Google Scholar] [CrossRef]
- Conrath, U.; Pieterse, C.M.J.; Mauch-Mani, B. Priming in plantpathogen interactions. Trends Plant. Sci. 2002, 7, 210–216. [Google Scholar] [CrossRef]
- Welbaum, G.E.; Sturz, A.V.; Dong, Z.; Nowak, J. Managing soil microorganisms to improve productivity of agro-ecosystems. Crit. Rev. Plant. Sci. 2004, 23, 175–193. [Google Scholar] [CrossRef]
- Mishra, A.; Singh, S.P.; Mahfooz, S.; Bhattacharya, A.; Mishra, N.; Shirke, P.A.; Nautiyal, C.S. Bacterial endophytes modulates the withanolide biosynthetic pathway and physiological performance in Withania somnifera under biotic stress. Microbiol. Res. 2018, 212–213, 17–28. [Google Scholar] [CrossRef]
- Ciccillo, F.; Fiore, A.; Bevivino, A.; Dalmastri, C.; Tabacchioni, S.; Chiarini, L. Effects of two different application methods of Burkholderia ambifaria MCI 7 on plant growth and rhizospheric bacterial diversity. Environ. Microbiol. 2002, 4, 238–245. [Google Scholar] [CrossRef]
- Lodewyckx, C.; Vangronsveld, J.; Porteous, F.; Moore, E.R.B.; Taghavi, S.; Mezgeay, M.; van der Lelie, D. Endophytic bacteria and their potential applications. Crit. Rev. Plant. Sci. 2002, 21, 583–606. [Google Scholar] [CrossRef]
- van Loon, L.C.; Bakker, P.A.H.M. Signalling in rhizobacteria plant interactions. In Ecological Studies, Root Ecology; De Kroon, J., Visser, E.J.W., Eds.; Springer: Berlin, Germany, 2004; Volume 168, pp. 287–330. [Google Scholar]
- Dash, C.K.; Bamisile, B.S.; Keppanan, R.; Qasim, M.; Lin, Y.; Islam, S.U.; Hussain, M.; Wang, L. Endophytic entomopathogenic fungi enhance the growth of Phaseolus vulgaris L. (Fabaceae) and negatively affect the development and reproduction of Tetranychus urticae Koch (Acari: Tetranychidae). Microb. Pathog. 2018, 125, 385–392. [Google Scholar] [CrossRef]
- Jaber, L.R. Seed inoculation with endophytic fungal entomopathogens promotes plant growth and reduces crown and root rot (CRR) caused by Fusarium culmorum in wheat. Planta 2018, 248, 1525–1535. [Google Scholar] [CrossRef]
- Yuan, Z.S.; Liu, F.; Xie, B.G.; Zhang, G.F. The growth-promoting effects of endophytic bacteria on Phyllostachys edulis. Arch. Microbiol. 2018, 200, 921–927. [Google Scholar] [CrossRef]
- Tall, S.; Meyling, N.V. Probiotics for Plants? Growth Promotion by the Entomopathogenic Fungus Beauveria bassiana Depends on Nutrient Availability. Microb. Ecol. 2018, 76, 1002–1008. [Google Scholar] [CrossRef] [PubMed]
- Elena, G.J.; Beatriz, P.J.; Alejandro, P.; Lecuona, R. Metarhizium anisopliae (Metschnikoff) Sorokin promotes growth and has endophytic activity in tomato plants. Adv. Biol. Res. 2011, 5, 22–27. [Google Scholar]
- Kuzhuppillymyal-Prabhakarankutty, L.; Tamez-Guerra, P.; Gomez-Flores, R.; Rodriguez-Padilla, M.C.; Ek-Ramos, M.J. Endophytic Beauveria bassiana promotes drought tolerance and early flowering in corn. World J. Microbiol. Biotechnol. 2020, 36, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; He, X.; Hou, L.; Ren, Y.; Wang, S.; Su, F. Dark septate endophytes isolated from a xerophyte plant promote the growth of Ammopiptanthus mongolicus under drought condition. Sci. Rep. 2018, 8, 26–28. [Google Scholar] [CrossRef]
- Mascarin, G.M.; Jaronski, S.T. The production and uses of Beauveria bassiana as a microbial insticide. World J. Microbiol. Biotechnol. 2016, 32, 177. [Google Scholar] [CrossRef] [PubMed]
- Renuka, S.; Ramanujam, B.; Poornesha, B. Endophytic Ability of Different Isolates of Entomopathogenic Fungi Beauveria bassiana (Balsamo) Vuillemin in Stem and Leaf Tissues of Maize (Zea mays L.). Indian J. Microbiol. 2016, 56, 126–133. [Google Scholar] [CrossRef]
- Ownley, B.H.; Griffin, M.R.; Klingeman, W.E.; Gwinn, K.D.; Moulton, J.K.; Pereira, R.M. Beauveria bassiana: Endophytic colonization and plant disease control. J. Invertebr. Pathol. 2008, 98, 267–270. [Google Scholar] [CrossRef]
- Russo, M.L.; Scorsetti, A.C.; Vianna, M.F.; Cabello, M.N. Endophytic Effects of Beauveria bassiana on Corn (Zea mays) and Its Herbivore, Rachiplusia nu (Lepidoptera: Noctuidae). Insects 2019, 10, 110. [Google Scholar] [CrossRef]
- Akutse, K.S.; Fiaboe, K.K.M.; Van Den Berg, J.; Ekesi, S.; Maniania, N.K. Effects of endophyte colonization of vicia faba (fabaceae) plants on the life-history of leafminer parasitoids phaedrotoma scabriventris (hymenoptera: Braconidae) and diglyphus isaea (hymenoptera: Eulophidae). PLoS ONE 2014, 9, e109965. [Google Scholar] [CrossRef]
- Ownley, B.H.; Gwinn, K.D.; Vega, F.E. Endophytic fungal entomopathogens with activity against plant pathogens: Ecology and evolution. BioControl 2010, 55, 113–128. [Google Scholar] [CrossRef]
- Parsa, S.; Ortiz, V.; Vega, F.E. Establishing fungal entomopathogens as endophytes: Towards endophytic biological control. J. Vis. Exp. 2013, 74, 1–5. [Google Scholar] [CrossRef]
- Vega, F.E. Inst pathology and fungal endophytes. J. Invertebr. Pathol. 2008, 98, 277–279. [Google Scholar] [CrossRef]
- Vega, F.E. The use of fungal entomopathogens as endophytes in biological control: A review. Mycologia 2018, 110, 4–30. [Google Scholar] [CrossRef] [PubMed]
- Ramos, Y.; Portal, O.; Lysøe, E.; Meyling, N.V.; Klingen, I. Diversity and abundance of Beauveria bassiana in soils, stink bugs and plant tissues of common bean from organic and conventional fields. J. Invertebr. Pathol. 2017, 150, 114–120. [Google Scholar] [CrossRef] [PubMed]
- Lopez, D.C.; Sword, G.A. The endophytic fungal entomopathogens Beauveria bassiana and Purpureocillium lilacinum enhance the growth of cultivated cotton (Gossypium hirsutum) and negatively affect survival of the cotton bollworm (Helicoverpa zea). Biol. Control. 2015, 89, 53–60. [Google Scholar] [CrossRef]
- McKinnon, A.C.; Glare, T.R.; Ridgway, H.J.; Mendoza-Mendoza, A.; Holyoake, A.; Godsoe, W.K.; Bufford, J.L. Detection of the entomopathogenic Fungus Beauveria bassiana in the rhizosphere of wound-stressed Zea mays plants. Front. Microbiol. 2018, 9, 1–16. [Google Scholar] [CrossRef]
- Akello, J.; Dubois, T.; Gold, C.S.; Coyne, D.; Nakavuma, J.; Paparu, P. Beauveria bassiana (Balsamo) Vuillemin as an endophyte in tissue culture banana (Musa spp.). J. Invertebr. Pathol. 2007, 96, 34–42. [Google Scholar] [CrossRef]
- Posada, F.; Aime, M.C.; Peterson, S.W.; Rehner, S.A.; Vega, F.E. Inoculation of coffee plants with the fungal entomopathogen Beauveria bassiana (Ascomycota: Hypocreales). Mycol. Res. 2007, 111, 748–757. [Google Scholar] [CrossRef]
- Tefera, T.; Vidal, S. Effect of inoculation method and plant growth medium on endophytic colonization of sorghum by the entomopathogenic fungus Beauveria bassiana. BioControl 2009, 54, 663–669. [Google Scholar] [CrossRef]
- Behie, S.W.; Jones, S.J.; Bidochka, M.J. Plant tissue localization of the endophytic inst pathogenic fungi Metarhizium and Beauveria. Fungal Ecol. 2015, 13, 112–119. [Google Scholar] [CrossRef]
- Jaber, L.R.; Enkerli, J. Fungal entomopathogens as endophytes: Can they promote plant growth? Biocontrol Sci. Technol. 2017, 27, 28–41. [Google Scholar] [CrossRef]
- Jaber, L.R.; Araj, S.-E. Interactions among endophytic fungal entomopathogens (Ascomycota: Hypocreales), the green peach aphid Myzus persicae Sulzer (Homoptera:Aphididae), and the aphid endoparasitoid Aphidius colemani Viereck (Hymenoptera:Braconidae). Biol. Control. 2017, 116, 53–61. [Google Scholar] [CrossRef]
- Jaber, L.R.; Enkerli, J. Effect of seed treatment duration on growth and colonizationof Vicia faba by endophytic Beauveria bassiana and Metarhizium brunneum. Biol. Control. 2016, 103, 187–195. [Google Scholar] [CrossRef]
- Mantzoukas, S.; Lagogiannis, I.; Karmakolia, K.; Rodi, A.; Gazepi, M.; Eliopoulos, P. The effect of grain type on virulence of entomopathogenic fungi against stored product pests. SI: Entomopathogenic Microorganisms: A Viable Tool for Controlling Serious Pests. Appl. Sci. 2020, 10, 2970. [Google Scholar] [CrossRef]
- Mantzoukas, S.; Lagogiannis, I.; Ntoukas, A.; Eliopoulos, P.A.; Kouretas, D.; Karpouzas, D.G.; Poulas, K. Trapping entomopathogenic fungi from vine terroir soil samples with inst baits for controlling serious pests. Appl. Sci. 2020, 10, 3539. [Google Scholar] [CrossRef]
- Quesada-Moraga, E.; Navas-Cortes, J.A.; Maranhao, A.A.; Ortiz-Urquiza, A.; Santiago-Alvarez, C. Factors affecting the occurence and distribution of entomopathogenic fungi in natural and cultivated soils. Mycol. Res. 2007, 111, 947–966. [Google Scholar] [CrossRef] [PubMed]
- Wagner, B.L.; Lewis, L.C. Colonization of corn, Zea mays, by the entomopathogenic fungus Beauveria bassiana. Appl. Environ. Microbiol. 2000, 66, 3468–3473. [Google Scholar] [CrossRef]
- Mantzoukas, S.; Pettas, I.; Lagogiannis, I. Stored product pests as models for trapping entomopathogenic fungi from olive tree orchards in Western Greece. J. Stored Prod. Res. 2020, 87, 101584. [Google Scholar] [CrossRef]
- Mantzoukas, S.; Chondrogiannis, C.; Grammatikopoulos, G. Effects of three endophytic entomopathogens on sweet sorghum and on the larvae of the stalk borer Sesamia nonagrioides. Entomol. Exp. Appl. 2015, 154, 78–87. [Google Scholar] [CrossRef]
- Tkaczuk, C. Występowanie i potencjał infek-cyjny grzybów owadobójczych w glebach agroce-noza I środowisk seminaturalnych w krajobrazie rolniczym. (Occurrence and infective potential of entomopathogenic fungi in the soils of agrocenoses and semi-natural habitats in agricultural landscape). Rozpr. Nauk. 2008, 94, 160. [Google Scholar]
- Meyling, N.V.; Eilenberg, J. Occurrence and distribution of soil borne entomopathogenic fungi within a single organic agroecosystem. Agric. Ecosyst. Environ. 2006, 113, 336–341. [Google Scholar] [CrossRef]
- Mantzoukas, S.; Lagogiannis, I. Endophytic Colonization of Pepper (Capsicum anuum) controls Aphids (Myzus persicae Sulzer). Appl. Sci. 2019, 9, 2239. [Google Scholar] [CrossRef]
- Rogers, S.O.; Bendich, A.J. Extraction of DNA from milligram amounts of fresh, herbarium and mummified plant tissues. Plant. Mol. Biol. 1985, 5, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Destefano, R.H.R.; Destefano, S.A.L.; Messias, C.L. Detection of Metarhizium anisopliae var. anisopliae within infected sugarcane borer Diatraea saccharalis (Lepidoptera, Pyralidae) using specific primers. Genet. Mol. Biol. 2004, 27, 245–252. [Google Scholar] [CrossRef][Green Version]
- Mantzoukas, S.; Grammatikopoulos, G. The effect of three entomopathogenic endophytes of the sweet sorghum on the growth and feeding performance of its pest, Sesamia nonagrioides larvae, and their efficacy under field conditions. Crop. Prot. 2020, 127, 104952. [Google Scholar] [CrossRef]
- Carroll, F.E.; Mu¨ller, E.; Sutton, B.C. Preliminary studies on the incidence of needle endophytes in some European conifers. Sydowia 1977, 29, 89–103. [Google Scholar]
- Bertoni, M.D.; Cabral, D. Phyllosphere of Eucalyptus viminalis. II:Distribution of endophytes. Nova Hedwig. 1988, 46, 491–502. [Google Scholar]
- Fisher, P.J.; Petrini, O.; Webstar, J. Aquatic hyphomycetes and other fungi in living aquatic and terrestrial roots of Alnus glutinosa. Mycol. Res. 1991, 95, 543–547. [Google Scholar] [CrossRef]
- Petrini, O.; Fisher, P.J. Fungal endophytes in Salicornia perennis. Trans. Br. Mycol. Soc. 1987, 87, 647–651. [Google Scholar] [CrossRef]
- Liang-Dong, G.; Guo-Rui, H.; Yu, W. Seasonal and tissue age influences on endophytic fungi of Pinus tabulaeformis (Pinaceae) in the Dongling Mountains, Beijing. J. Integr. Biol. 2008, 50, 997–1003. [Google Scholar]
- Allegrucci, N.; Velazquez, M.S.; Russo, M.L.; Perez, E.; Scorsetti, A.C. Endophytic colonisation of tomato by the entomopathogenic fungus Beauveria bassiana: The use of different inoculation techniques and their effects on the tomato leafminer Tuta absoluta (Lepidoptera: Gelechiidae). J. Plant. Prot. Res. 2017, 57, 205–211. [Google Scholar] [CrossRef]
- Greenfield, M.; Gómez-Jiménez, M.; Ortiz, V.; Vega, F.E.; Kramer, M.; Parsa, S. Beauveria bassiana and Metarhizium anisopliae endophytically colonize cassava roots following soil drench inoculation. Biol. Control. 2016, 95, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Marasco, R.; Rolli, E.; Fusi, M.; Michoud, G.; Daffonchio, D. Grapevine rootstocks shape underground bacterial microbiome and networking but not potential functionality. Microbiome 2018, 6, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Company, S.; Reiter, B.; Sessitsch, A.; Nowak, J.; Clément, C.; Ait Barka, E. Endophytic colonization of Vitis vinifera L. by plant growth-promoting bacterium Burkholderia sp. strain PsJN. Appl. Environ. Microbiol. 2005, 71, 1685–1693. [Google Scholar] [CrossRef]
- Quesada-Moraga, E.; Munoz-Ledesma, F.J.; Santiago-Alvarez, C. Systemic protection of papaver somniferum L. against Iraella luteipes (Hymenoptera: Cynipidae) by an endophytic strain of beauveria bassiana (Ascomycota: Hypocreales). Environ. Entomol. 2009, 38, 723–730. [Google Scholar] [CrossRef]
- Ownley, B.H.; Dee, M.M.; Gwinn, K. Effect of conidial seed treatment rate of entomopathogenic Beauveria bassiana11-98 on endophytic colonization of tomato seedlings and control of Rhizoctonia disease. Phytopathology 2008, 98, S118. [Google Scholar]
- Akutse, K.S.; Maniania, N.K.; Fiaboe, K.K.M.; van den Berg, J.; Ekesi, S. Endophytic colonization of Vicia faba and Phaseolus vulgaris (Fabaceae) by fungal pathogens and their effects on the life-history parameters of Liriomyza huidobrensis (Diptera: Agromyzidae). Fungal Ecol. 2013, 6, 293–301. [Google Scholar] [CrossRef]
- Lingg, A.J.; Donaldson, M.D. Biotic and abiotic factors affecting stability of Beauveria bassiana conidia in soil. J. Invertebr. Pathol. 1981, 38, 191–200. [Google Scholar] [CrossRef]
- Meyling, N.V.; Hajek, A.E. Principles from community and metapopulation ecology: Application to fungal entomopathogens. BioControl 2010, 55, 39–54. [Google Scholar] [CrossRef]
- Zimmermann, G. The Galleria bait method for detection of entomopathogenic fungi in soil. J. Appl. Entomol. 1986, 102, 213–215. [Google Scholar] [CrossRef]
- Rodriguez, R.J.; White, J.F.; Arnold, A.E.; Redman, R.S. Fungal endophytes: Diversity and functional roles. New Phytol. 2009, 182, 314–330. [Google Scholar] [CrossRef]
- Yan, Z.I.; Zhang, C.; Lin, F.C. Role of diverse non-systemic fungal endophytes in plant performance and response of stress: Progress and approaches. Plant. Growth Regul. 2010, 29, 116–126. [Google Scholar] [CrossRef]
- Kannadan, S.; Rudgers, J.A. Endophyte symbiosis benefits a rare grass under low water availability. Funct. Ecol. 2008, 22, 706–713. [Google Scholar] [CrossRef]
- Rondot, Y.; Reineke, A. Endophytic Beauveria bassiana in grapevine Vitis vinifera (L.) reduces infestation with piercing-sucking insts. Biol. Control. 2018, 116, 82–89. [Google Scholar] [CrossRef]
- Porras-Alfaro, A.; Bayman, P. Hidden fungi, emergent properties: Endophytes and microbiomes. Annu. Rev. Phytopathol. 2011, 49, 291–315. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Vidal, S.; Salinas, J.; Tena, M.; Lopez-Llorca, L.V. Proteomic analysis of date palm (Phoenix dactylifera L.) responses to endophytic colonization by entomopathogenic fungi. Electrophoresis 2009, 30, 2996–3005. [Google Scholar]
- Jackson, M.; Dunlap, C.; Jaronski, S. Ecological considerations in producing and formulating fungal entomopathogens for use in inst biocontrol. Biocontrol 2010, 55, 129–145. [Google Scholar] [CrossRef]
- Vega, F.E.; Posada, F.; Aime, M.C.; Pava-Ripoll, M.; Infante, F.; Rehner, S.A. Entomopathogenic fungal endophytes. Biol. Control. 2008, 46, 72–82. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Fungal Species | DNA | Sequence Length | Blast ID Number | Conidia Microscope Evaluation | Match Percent (%) |
|---|---|---|---|---|---|---|
| Plant Tissues | B. bassiana | CCGGTAAACCACAGTATCTACCTGATTCGAGGTCACGTTCAGAAGTTGGGTTTTTACGGCGTGGCCGCGTCGGGGTTCCGGTGCGAGCTGTATTACTGCGCAGAGGTCGCCGCGGACGGGCCGCCACTCCATTTCAGGGCCGGCGGTGTGCTGCCGGTCCCCAACGCCGACCTCCCCAAGGGGAGGTCGAGGGTTGAAATGACGCTCGA ACAGGCATGCCCGCCAGAATGCTGGCGGGCGCAATGTGCGTTCAAAGATTCGATGATTCACTGGATTCTGCAATTCACATTACTTATCGCGTTTCGCTGCGTTCTTCATCGATGCCAGAGCCAAGAGATCCGTTGTTGAAAGTTTTGATTCATTTGTTTTGCCTTGCGGCGTATTCAGAAGATGCTGGAATACAAGAGTTTGAGGTCCCCGGCGGGCCGCTGGTCCAGTCCGCGTCCGGGCTGGGGCGAGTCCGCCGAAGCAACGATAGGTAGGTTCACAGAAGGGTTAGGGAGTTGAAAACTCGGTAATGATCCCTCCGCTGGTTCACCAACGGAGACCTTGTTACGACTTTTACTTCCA | 555 | 20200105GS1P2_E01_20 | Yes | 100 |
| T. confusum baits | B. bassiana | 20200105GS1P2_E02_20 | Yes | 100 |
| Plant Growth Parameters | Self-Rooted Control | Self-Rooted Endophtyic B. bassiana | R110 Control | R110 Endophtyic B. bassiana |
|---|---|---|---|---|
| Number Leaves | 33.18 ± 3.71a | 40.84 ± 2.9b | 29.25 ± 5.77a | 40.73 ± 3.15b |
| Height of plant | 40.09 ± 1.10b | 45.08 ± 2.69c | 34 ± 1.96a | 40.53 ± 1.80b |
| Thickness of the stem | 10.63 ± 1.15a | 11.58 ± 1.67a | 11.08 ± 2.63a | 11.84 ± 2.23a |
| Length of clematis | 47.36 ± 8.62a | 58.77 ± 19.15a | 51.20 ± 10.83a | 56.12 ± 17.66a |
| Length of Root | 29.18 ± 3.83a | 39 ± 2.56b | 29.58 ± 4.55a | 37.75 ± 1.58b |
| Leaf dry weight | 19.04 ± 1.98a | 22.43 ± 1.81a | 20.69 ± 3.69a | 25.18 ± 2.53a |
| Shoot dry weight | 14 ± 6.14a | 19.56 ± 6.52a | 17.78 ± 10.31a | 20.99 ± 8.46a |
| Stem dry weight | 18.75 ± 4.59a | 20.63 ± 6.16a | 19.03 ± 8.72a | 22.59 ± 12.99a |
| Root dry weight | 35.02 ± 1.34a | 57.63 ± 3.02b | 49.48 ± 2.42a | 81.15 ± 5.74c |
| Aboveground dry weight, A (g) | 51.79 | 60.46 | 57.50 | 68.76 |
| Belowground dry weight, B (g) | 35.02 | 57.63 | 49.48 | 81.15 |
| Ratio A/B | 1.47 | 1.04 | 1.16 | 0.84 |
| Total biomass (g) | 86.81 ± 2.54a | 118.09 ± 1.98c | 106.98 ± 1.85b | 149.91 ± 3.17d |
| Soil Mortality of T. confusum | ||||
|---|---|---|---|---|
| Treatment | 2d | 4d | 6d | Mycelium Present on Dead Adults |
| Self-rooted Control | 3.67 ± 2.75a | 6.13 ± 1.96a | 7.89 ± 2.11a | 0 ± 0a |
| Self-rooted Endophytic B. bassiana | 44.06 ± 3.21b | 63.54 ± 5.40b | 91.09 ± 3.56b | 87.97 ± 5.08b |
| R110 Control | 4.54 ± 2.67a | 6.75 ± 4.13a | 8.32 ± 2.51a | 0 ± 0a |
| R110 Endophytic B. bassiana | 52.79 ± 4.67c | 71.11 ± 0.42c | 87.47 ± 1.96b | 73.45 ± 3.51c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mantzoukas, S.; Lagogiannis, I.; Mpousia, D.; Ntoukas, A.; Karmakolia, K.; Eliopoulos, P.A.; Poulas, K. Beauveria bassiana Endophytic Strain as Plant Growth Promoter: The Case of the Grape Vine Vitis vinifera. J. Fungi 2021, 7, 142. https://doi.org/10.3390/jof7020142
Mantzoukas S, Lagogiannis I, Mpousia D, Ntoukas A, Karmakolia K, Eliopoulos PA, Poulas K. Beauveria bassiana Endophytic Strain as Plant Growth Promoter: The Case of the Grape Vine Vitis vinifera. Journal of Fungi. 2021; 7(2):142. https://doi.org/10.3390/jof7020142
Chicago/Turabian StyleMantzoukas, Spiridon, Ioannis Lagogiannis, Dionusia Mpousia, Aristeidis Ntoukas, Katerina Karmakolia, Panagiotis A. Eliopoulos, and Konstantinos Poulas. 2021. "Beauveria bassiana Endophytic Strain as Plant Growth Promoter: The Case of the Grape Vine Vitis vinifera" Journal of Fungi 7, no. 2: 142. https://doi.org/10.3390/jof7020142
APA StyleMantzoukas, S., Lagogiannis, I., Mpousia, D., Ntoukas, A., Karmakolia, K., Eliopoulos, P. A., & Poulas, K. (2021). Beauveria bassiana Endophytic Strain as Plant Growth Promoter: The Case of the Grape Vine Vitis vinifera. Journal of Fungi, 7(2), 142. https://doi.org/10.3390/jof7020142