
Transmission and Colonization of Pneumocystis jirovecii


Abstract
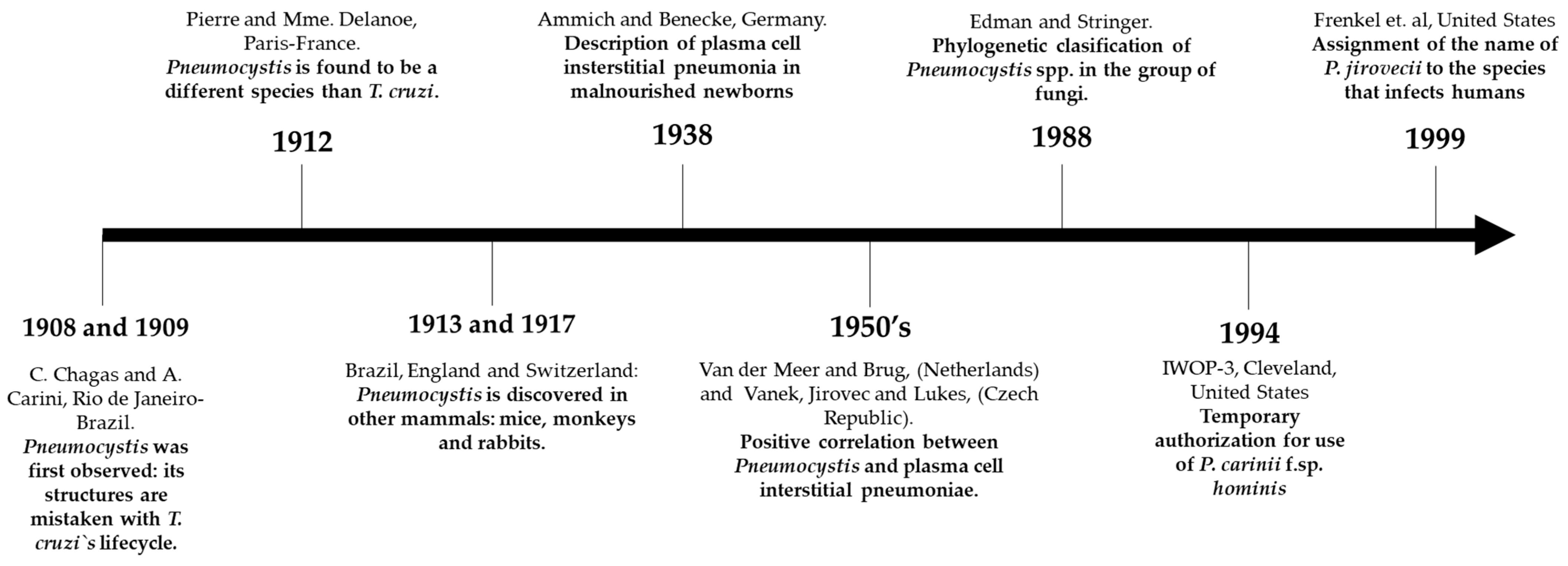
1. Pneumocystis spp.: Brief Description of Its Discovery
2. Nomenclature and Genetic Characteristics
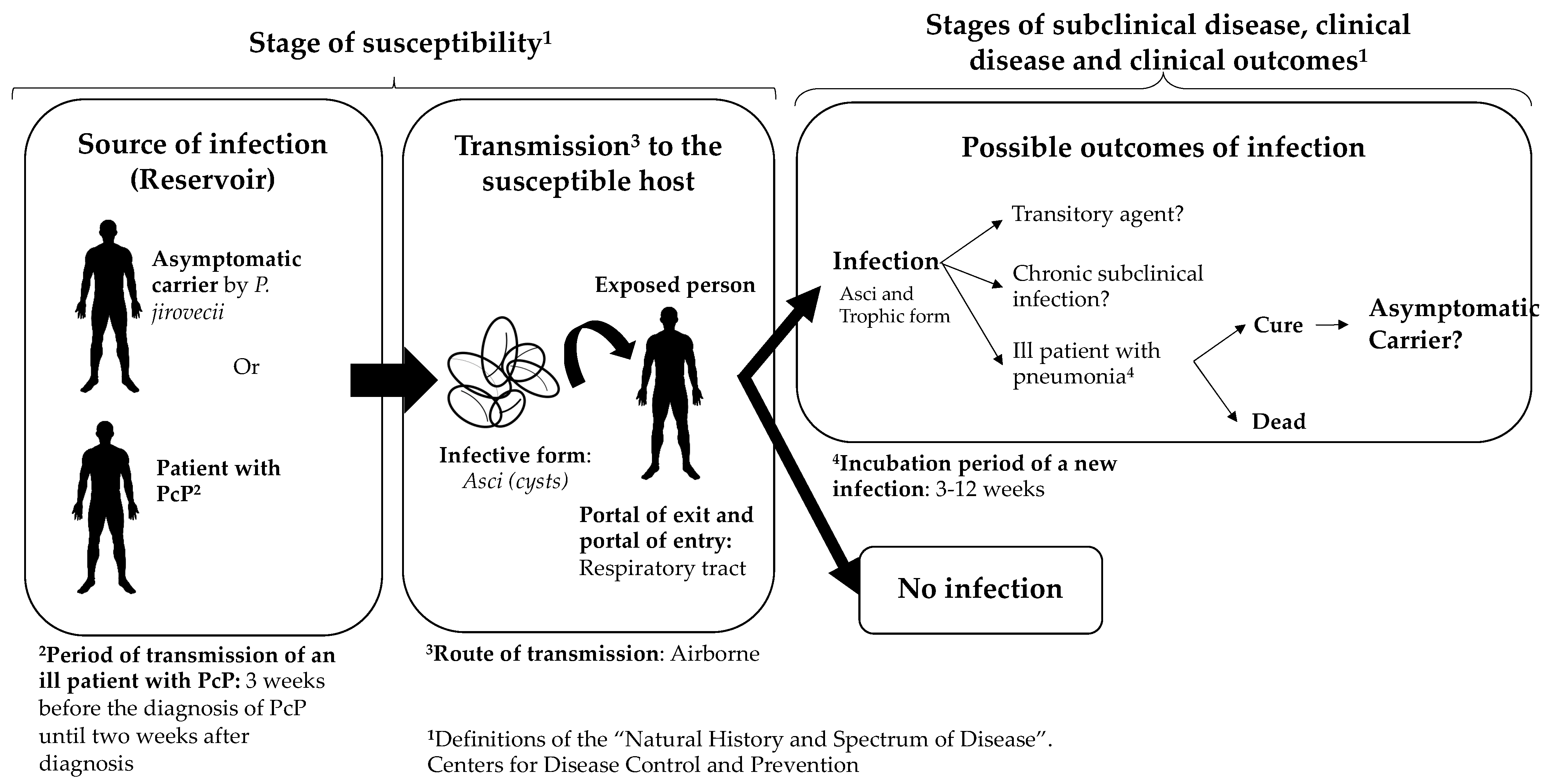
3. Transmission of Pneumocystis jirovecii
4. Weather and P. jirovecii
5. Colonization of P. jirovecii in Different Population Groups
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chagas, C. Nova tripanozomiaze humana. Estudios sobre a morfolojia e o ciclo evolutivo do Schizotrypanum cruzi n. gen., n. sp., ajente etiolojico de nova entidade morbida do homen. Mem. Inst. Oswaldo Cruz. 1909, 1, 159–218. [Google Scholar] [CrossRef]
- Delanöe, P.; Delanöe, M. Sur les rapports des kystes de carinii du poumon des rats avec le Trypanosoma lewisi. CR Acad. Sci. 1912, 155, 658–660. [Google Scholar]
- Keely, S.P.; Stringer, J.R. Part One. THE ORGANISM. 1. Historical Overview. In Pneumocystis Pneumonia, 3rd ed.; rev.expanded; Walzer, P.D., Cushion, M.T., Eds.; Marcel Dekker: New York, NY, USA, 2005; pp. 39–55, (Lung biology in health and disease). [Google Scholar]
- Calderón-Sandubete, E.J.; Varela-Aguilar, J.M.; Medrano-Ortega, F.J.; Nieto-Guerrer, V.; Respaldiza-Salas, N.; de la Horra-Padilla, C.; Dei-Cas, E. Historical perspective on Pneumocystis carinii infection. Protist 2002, 153, 303–310. [Google Scholar] [CrossRef]
- Gajdusek, D.C. Pneumocystis Carinii—Etiologic Agent of Interstitial Plasma Cell Pneumonia of Premature and Young Infants. Pediatrics 1957, 19, 543–565. [Google Scholar] [PubMed]
- Vavra, J.; Kucera, K. Pneumocystis carinii Delanoë, its Ultrastructure and Ultrastructural Affinities*. J. Protozool. 1970, 17, 463–483. [Google Scholar] [CrossRef]
- Edman, J.R.; Kovacs, J.A.; Masur, H.; Santi, D.; Elwood, H.J.; Sogin, M.L. Ribosomal RNA sequence shows Pneumocystis carinii to be a member of the Fungi. Nature 1988, 334, 519–522. [Google Scholar] [CrossRef]
- Stringer, S.L.; Stringer, J.R.; Blase, M.A.; Walzer, P.D.; Cushion, M.T. Pneumocystis carinii: Sequence from ribosomal RNA implies a close relationship with fungi. Exp. Parasitol. 1989, 68, 450–461. [Google Scholar] [CrossRef]
- Wakefield, A.E.; Stringer, J.R.; Tamburrini, E.; Dei-Cas, E. Genetics, metabolism and host specificity of Pneumocystis carinii. Med. Mycol. 1998, 36 (Suppl. 1), 183–193. [Google Scholar] [PubMed]
- Bartlett, M.; Cushion, M.T.; Fishman, J.A.; Kaneshiro, E.S.; Lee, C.H.; Leibowitz, M.J.; Lu, J.J.; Lundgren, B.; Peters, S.E.; Smith, J.W.; et al. Revised nomenclature for Pneumocystis carinii. The Pneumocystis Workshop. J. Eukaryot. Microbiol. 1994, 41, 121S–122S. [Google Scholar]
- Frenkel, J.K. Pneumocystis jiroveci n. sp. from man: Morphology, physiology, and immunology in relation to pathology. Natl. Cancer Inst. Monogr. 1976, 43, 13–30. [Google Scholar]
- Redhead, S.A.; Cushion, M.T.; Frenkel, J.K.; Stringer, J.R. Pneumocystis and Trypanosoma cruzi: Nomenclature and typifications. J. Eukaryot. Microbiol. 2006, 53, 2–11. [Google Scholar] [CrossRef]
- Cushion, M.T.; Kaselis, M.; Stringer, S.L.; Stringer, J.R. Genetic stability and diversity of Pneumocystis carinii infecting rat colonies. Infect. Immun. 1993, 61, 4801–4813. [Google Scholar] [CrossRef]
- Cushion, M.T.; Smulian, A.G. The pneumocystis genome project: Update and issues. J. Eukaryot. Microbiol. 2001, 48, 182S–183S. [Google Scholar] [CrossRef]
- Sesterhenn, T.M.; Slaven, B.E.; Smulian, A.G.; Cushion, M.T. Generation of sequencing libraries for the Pneumocystis Genome project. J. Eukaryot. Microbiol. 2003, 50, 663–665. [Google Scholar] [CrossRef]
- Slaven, B.E.; Meller, J.; Porollo, A.; Sesterhenn, T.; Smulian, A.G.; Cushion, M.T. Draft Assembly and Annotation of the Pneumocystis carinii Genome. J. Eukaryot. Microbiol. 2006, 53, S89–S91. [Google Scholar] [CrossRef] [PubMed]
- Cissé, O.H.; Pagni, M.; Hauser, P.M. De novo assembly of the Pneumocystis jirovecii genome from a single bronchoalveolar lavage fluid specimen from a patient. mBio 2012, 4, e00412–e00428. [Google Scholar] [CrossRef]
- Ma, L.; Huang, D.-W.; Cuomo, C.A.; Sykes, S.; Fantoni, G.; Das, B.; Sherman, B.T.; Yang, J.; Huber, C.; Xia, Y.; et al. Sequencing and characterization of the complete mitochondrial genomes of three Pneumocystis species provide new insights into divergence between human and rodent Pneumocystis. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2013, 27, 1962–1972. [Google Scholar] [CrossRef] [PubMed]
- Cushion, M.T.; Keely, S.P. Assembly and annotation of Pneumocystis jirovecii from the human lung microbiome. mBio 2013, 4, e00224. [Google Scholar] [CrossRef]
- Ma, L.; Chen, Z.; Huang, D.W.; Kutty, G.; Ishihara, M.; Wang, H.; Abouelleil, A.; Bishop, L.; Davey, E.; Deng, R.; et al. Genome analysis of three Pneumocystis species reveals adaptation mechanisms to life exclusively in mammalian hosts. Nat. Commun. 2016, 7, 10740. [Google Scholar] [CrossRef] [PubMed]
- Cissé, O.H.; Hauser, P.M. Genomics and evolution of Pneumocystis species. Infect. Genet. Evol. 2018, 65, 308–320. [Google Scholar] [CrossRef]
- Cissé, O.H.; Ma, L.; Dekker, J.P.; Khil, P.P.; Youn, J.-H.; Brenchley, J.M.; Blair, R.; Pahar, P.; Chabé, M.; Van Rompay, K.; et al. Genomic insights into the host specific adaptation of the Pneumocystis genus. Commun. Biol. 2021, 4, 305. [Google Scholar] [CrossRef]
- Cissé, O.H.; Pagni, M.; Hauser, P.M. Comparative genomics suggests that the human pathogenic fungus Pneumocystis jirovecii acquired obligate biotrophy through gene loss. Genome Biol. Evol. 2014, 6, 1938–1948. [Google Scholar] [CrossRef] [PubMed]
- Hauser, P.M. Genomic insights into the fungal pathogens of the genus pneumocystis: Obligate biotrophs of humans and other mammals. PLoS Pathog. 2014, 10, e1004425. [Google Scholar] [CrossRef]
- Palluault, F.; Slomianny, C.; Soulez, B.; Dei-Cas, E.; Camus, D. High osmotic pressure enables fine ultrastructural and cytochemical studies onPneumocystic carinii. Parasitol. Res. 1992, 78, 437–444. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, H.; Yoshida, Y. Localization of silver deposits on Pneumocystis carinii treated with Gomori’s methenamine silver nitrate stain. Zentralblatt. Für Bakteriol. Mikrobiol. Hyg. Ser. Med. Microbiol. Infect. Dis. Virol. Parasitol. 1987, 264, 363–372. [Google Scholar] [CrossRef]
- Cushion, M.T.; Ashbaugh, A.; Hendrix, K.; Linke, M.J.; Tisdale, N.; Sayson, S.G.; Porollo, A. Gene Expression of Pneumocystis murina after Treatment with Anidulafungin Results in Strong Signals for Sexual Reproduction, Cell Wall Integrity, and Cell Cycle Arrest, Indicating a Requirement for Ascus Formation for Proliferation. Antimicrob. Agents Chemother. 2018, 62, e02513–e02517. [Google Scholar] [CrossRef] [PubMed]
- Opata, M.M.; Ye, Z.; Hollifield, M.; Garvy, B.A. B cell production of tumor necrosis factor in response to Pneumocystis murina infection in mice. Infect. Immun. 2013, 81, 4252–4260. [Google Scholar] [CrossRef][Green Version]
- Linke, M.J.; Ashbaugh, A.A.; Koch, J.V.; Levin, L.; Tanaka, R.; Walzer, P.D. Effects of surfactant protein-A on the interaction of Pneumocystis murina with its host at different stages of the infection in mice. J. Eukaryot. Microbiol. 2009, 56, 58–65. [Google Scholar] [CrossRef]
- Dei-Cas, E.; Chabé, M.; Moukhlis, R.; Durand-Joly, I.; Aliouat, E.M.; Stringer, J.R.; Cushion, M.; Noël, C.; de Hoog, G.S.; Guillot, J.; et al. Pneumocystis oryctolagi sp. nov., an uncultured fungus causing pneumonia in rabbits at weaning: Review of current knowledge, and description of a new taxon on genotypic, phylogenetic and phenotypic bases. FEMS Microbiol. Rev. 2006, 30, 853–871. [Google Scholar] [CrossRef]
- Ma, L.; Cissé, O.H.; Kovacs, J.A. A Molecular Window into the Biology and Epidemiology of Pneumocystis spp. Clin. Microbiol. Rev. 2018, 31, e00009-18. [Google Scholar] [CrossRef]
- Vossen, M.E.; Beckers, P.J.; Meuwissen, J.H.; Stadhouders, A.M. Developmental biology of Pneumocystis carinii, and alternative view on the life cycle of the parasite. Z. Parasitenkd Berl. Ger. 1978, 55, 101–118. [Google Scholar] [CrossRef]
- Yoshida, Y. Ultrastructural studies of Pneumocystis carinii. J. Protozool. 1989, 36, 53–60. [Google Scholar] [CrossRef]
- Hauser, P.M.; Cushion, M.T. Is sex necessary for the proliferation and transmission of Pneumocystis? PLoS Pathog. 2018, 14, e1007409. [Google Scholar] [CrossRef]
- Matsumoto, Y.; Yoshida, Y. Sporogony in Pneumocystis carinii: Synaptonemal complexes and meiotic nuclear divisions observed in precysts. J. Protozool. 1984, 31, 420–428. [Google Scholar] [CrossRef] [PubMed]
- Richard, S.; Almeida, J.M.G.C.F.; Cissé, O.H.; Luraschi, A.; Nielsen, O.; Pagni, M.; Houser, P.M. Functional and Expression Analyses of the Pneumocystis MAT Genes Suggest Obligate Sexuality through Primary Homothallism within Host Lungs. mBio 2018, 9, e02201-17. [Google Scholar] [CrossRef]
- Hauser, P.M. Pneumocystis Mating-Type Locus and Sexual Cycle during Infection. Microbiol. Mol. Biol. Rev. MMBR 2021, 85, e0000921. [Google Scholar] [CrossRef]
- Cissé, O.H.; Ma, L.; Jiang, C.; Snyder, M.; Kovacs, J.A. Humans Are Selectively Exposed to Pneumocystis jirovecii. mBio 2020, 11, e03138-19. [Google Scholar] [CrossRef] [PubMed]
- Dumoulin, A.; Mazars, E.; Seguy, N.; Gargallo-Viola, D.; Vargas, S.; Cailliez, J.C.; Aliouat, E.M.; Wakefield, A.E.; Dei-Cas, E. Transmission of Pneumocystis carinii Disease from Immunocompetent Contacts of Infected Hosts to Susceptible Hosts. Eur. J. Clin. Microbiol. Infect. Dis. 2000, 19, 671–678. [Google Scholar] [CrossRef]
- Le Gal, S.; Pougnet, L.; Damiani, C.; Fréalle, E.; Guéguen, P.; Virmaux, M.; Ansart, S.; Jaffuel, S.; Couturaud, F.; Delluc, A.; et al. Pneumocystis jirovecii in the air surrounding patients with Pneumocystis pulmonary colonization. Diagn. Microbiol. Infect. Dis. 2015, 82, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Hughes, W.T.; Bartley, D.L.; Smith, B.M. A natural source of infection due to pneumocystis carinii. J. Infect. Dis. 1983, 147, 595. [Google Scholar] [CrossRef]
- Bartlett, M.S.; Vermund, S.H.; Jacobs, R.; Durant, P.J.; Shaw, M.M.; Smith, J.W.; Tang, X.; Lu, J.J.; Li, B.; Jin, S.; et al. Detection of Pneumocystis carinii DNA in air samples: Likely environmental risk to susceptible persons. J. Clin. Microbiol. 1997, 35, 2511–2513. [Google Scholar] [CrossRef] [PubMed]
- Casanova-Cardiel, L.; Leibowitz, M.J. Presence of Pneumocystis carinii DNA in pond water. J. Eukaryot. Microbiol. 1997, 44, 28S. [Google Scholar] [CrossRef]
- Choukri, F.; Menotti, J.; Sarfati, C.; Lucet, J.-C.; Nevez, G.; Garin, Y.J.F.; Derouin, F.; Totet, A. Quantification and spread of Pneumocystis jirovecii in the surrounding air of patients with Pneumocystis pneumonia. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2010, 51, 259–265. [Google Scholar] [CrossRef] [PubMed]
- Cushion, M.T.; Linke, M.J.; Ashbaugh, A.; Sesterhenn, T.; Collins, M.S.; Lynch, K.; Brubaker, R.; Walzer, P.D. Echinocandin treatment of pneumocystis pneumonia in rodent models depletes cysts leaving trophic burdens that cannot transmit the infection. PLoS ONE 2010, 5, e8524. [Google Scholar] [CrossRef]
- Sing, A.; Wonhas, C.; Bader, L.; Luther, M.; Heesemann, J. Detection of Pneumocystis carinii DNA in the air filter of a ventilated patient with AIDS. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 1999, 29, 952–953. [Google Scholar] [CrossRef][Green Version]
- Olsson, M.; Lidman, C.; Latouche, S.; Björkman, A.; Roux, P.; Linder, E.; Wahlgren, M. Identification of Pneumocystis carinii f. sp. hominis gene sequences in filtered air in hospital environments. J. Clin. Microbiol. 1998, 36, 1737–1740. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi-Ghalehbin, B.; Habibzadeh, S.; Arzanlou, M.; Teimourpour, R.; Amani Ghayum, S. Colonization of Pneumocystis jirovecii in Patients Who Received and not Received Corticosteroids Admitted to the Intensive Care Unit: Airborne Transmission Approach. Iran J. Pathol. 2018, 13, 136–143. [Google Scholar] [CrossRef]
- Arichi, N.; Kishikawa, H.; Mitsui, Y.; Kato, T.; Nishimura, K.; Tachikawa, R.; Tomii, K.; Shiina, H.; Igawa, M.; Ichikawa, Y. Cluster outbreak of Pneumocystis pneumonia among kidney transplant patients within a single center. Transplant. Proc. 2009, 41, 170–172. [Google Scholar] [CrossRef]
- Rostved, A.A.; Sassi, M.; Kurtzhals, J.A.L.; Sørensen, S.S.; Rasmussen, A.; Ross, C.; Gogineni, E.; Huber, C.; Kutty, G.; Kovacs, J.A.; et al. Outbreak of pneumocystis pneumonia in renal and liver transplant patients caused by genotypically distinct strains of Pneumocystis jirovecii. Transplantation 2013, 96, 834–842. [Google Scholar] [CrossRef]
- Chandola, P.; Lall, M.; Sen, S.; Bharadwaj, R. Outbreak of Pneumocystis jirovecii pneumonia in renal transplant recipients on prophylaxis: Our observation and experience. Indian J. Med. Microbiol. 2014, 32, 333–336. [Google Scholar] [CrossRef]
- Chave, J.P.; David, S.; Wauters, J.P.; Van Melle, G.; Francioli, P. Transmission of Pneumocystis carinii from AIDS patients to other immunosuppressed patients: A cluster of Pneumocystis carinii pneumonia in renal transplant recipients. AIDS Lond. Engl. 1991, 5, 927–932. [Google Scholar] [CrossRef]
- Manoloff, E.S.; Francioli, P.; Taffé, P.; van Melle, G.; Bille, J.; Hauser, P.M. Risk for Pneumocystis carinii Transmission among Patients with Pneumonia: A Molecular Epidemiology Study. Emerg. Infect. Dis. 2003, 9, 132–134. [Google Scholar] [CrossRef]
- De Boer, M.G.J.; van Bruijnesteijn Coppenraet, L.E.S.; Gaasbeek, A.; Berger, S.P.; Gelinck, L.B.S.; van Houwelingen, H.C.; van den Broek, P.; Kuijper, E.D.J.; Kroon, F.P.; Vandenbroucke, J.P.; et al. An Outbreak of Pneumocystis jiroveci Pneumonia with 1 Predominant Genotype among Renal Transplant Recipients: Interhuman Transmission or a Common Environmental Source? Clin. Infect. Dis. 2007, 44, 1143–1149. [Google Scholar] [CrossRef] [PubMed]
- Rabodonirina, M.; Vanhems, P.; Couray-Targe, S.; Gillibert, R.-P.; Ganne, C.; Nizard, N.; Colin, C.; Fabry, J.; Touraine, J.L.; van Melle, G.; et al. Molecular evidence of interhuman transmission of Pneumocystis pneumonia among renal transplant recipients hospitalized with HIV-infected patients. Emerg. Infect. Dis. 2004, 10, 1766–1773. [Google Scholar] [CrossRef]
- Jiang, C.; Wang, X.; Li, X.; Inlora, J.; Wang, T.; Liu, Q.; Snyder, M. Dynamic Human Environmental Exposome Revealed by Longitudinal Personal Monitoring. Cell 2018, 175, 277–291.e31. [Google Scholar] [CrossRef]
- Gigliotti, F.; Wright, T.W. Pneumocystis: Where does it live? PLoS Pathog. 2012, 8, e1003025. [Google Scholar] [CrossRef] [PubMed]
- Le Gal, S.; Damiani, C.; Rouillé, A.; Grall, A.; Tréguer, L.; Virmaux, M.; Moalic, E.; Quinio, D.; Moal, M.C.; Berthou, C.; et al. A cluster of Pneumocystis infections among renal transplant recipients: Molecular evidence of colonized patients as potential infectious sources of Pneumocystis jirovecii. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2012, 54, e62–e71. [Google Scholar] [CrossRef]
- Vanhems, P.; Hirschel, B.; Morabia, A. Seasonal incidence of Pneumocystis carinii pneumonia. Lancet Lond. Engl. 1992, 339, 1182. [Google Scholar] [CrossRef]
- De la López Osa, A.; Gutiérrez, F.; Sánchez, C.; Mediavilla, J.D.; Aliaga, L. Do seasonal changes have any influence on Pneumocystis carinii pneumonia? Enferm. Infecc. Microbiol. Clin. 1998, 16, 435. [Google Scholar]
- Miller, R.F.; Evans, H.E.R.; Copas, A.J.; Huggett, J.F.; Edwards, S.G.; Walzer, P.D. Seasonal variation in mortality of Pneumocystis jirovecii pneumonia in HIV-infected patients. Int. J. STD Aids 2010, 21, 497–503. [Google Scholar] [CrossRef] [PubMed]
- Demanche, C.; Wanert, F.; Herrenschmidt, N.; Moussu, C.; Durand-Joly, I.; Dei-Cas, E.; Chermette, R.; Guillot, J. Influence of climatic factors on Pneumocystis carriage within a socially organized group of immunocompetent macaques (Macaca fascicularis). J. Eukaryot. Microbiol. 2003, 50, 611–613. [Google Scholar] [CrossRef]
- Laakkonen, J.; Henttonen, H.; Niemimaa, J.; Soveri, T. Seasonal dynamics of Pneumocystis carinii in the field vole, Microtus agrestis, and in the common shrew, Sorex araneus, in Finland. Parasitology 1999, 118 Pt 1, 1–5. [Google Scholar] [CrossRef]
- Djawe, K.; Levin, L.; Swartzman, A.; Fong, S.; Roth, B.; Subramanian, A.; Grieco, K.; Jarlsberg, L.; Miller, R.F.; Huang, L.; et al. Environmental risk factors for Pneumocystis pneumonia hospitalizations in HIV patients. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2013, 56, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Lubis, N.; Baylis, D.; Short, A.; Stebbing, J.; Teague, A.; Portsmouth, S.; Bower, M.; Nelson, M.; Gazzard, B. Prospective cohort study showing changes in the monthly incidence of Pneumocystis carinii pneumonia. Postgrad. Med. J. 2003, 79, 164–166. [Google Scholar] [CrossRef]
- Varela, J.M.; Regordán, C.; Medrano, F.J.; Respaldiza, N.; de La Horra, C.; Montes-Cano, M.A.; Calderón, E.J. Climatic factors and Pneumocystis jiroveci infection in southern Spain. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2004, 10, 770–772. [Google Scholar] [CrossRef] [PubMed]
- Calderón, E.J.; Varela, J.M.; Medrano, F.J.; Nieto, V.; González-Becerra, C.; Respaldiza, N.; De La Horra, C.; Montes-Cano, M.A.; Vigil, E.; González de la Puente, M.A.; et al. Epidemiology of Pneumocystis carinii pneumonia in southern Spain. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2004, 10, 673–676. [Google Scholar] [CrossRef][Green Version]
- Sing, A.; Schmoldt, S.; Laubender, R.P.; Heesemann, J.; Sing, D.; Wildner, M. Seasonal variation of Pneumocystis jirovecii infection: Analysis of underlying climatic factors. Clin. Microbiol. Infect. 2009, 15, 957–960. [Google Scholar] [CrossRef]
- Cushion, M.T. Are members of the fungal genus pneumocystis (a) commensals, (b) opportunists, (c) pathogens, or (d) all of the above? PLoS Pathog. 2010, 6, e1001009. [Google Scholar] [CrossRef] [PubMed]
- Morris, A.; Norris, K.A. Colonization by Pneumocystis jirovecii and its role in disease. Clin. Microbiol. Rev. 2012, 25, 297–317. [Google Scholar] [CrossRef]
- Cushion, M.T.; Stringer, J.R. Stealth and opportunism: Alternative lifestyles of species in the fungal genus Pneumocystis. Annu. Rev. Microbiol. 2010, 64, 431–452. [Google Scholar] [CrossRef] [PubMed]
- Medrano, F.J.; Montes-Cano, M.; Conde, M.; de la Horra, C.; Respaldiza, N.; Gasch, A.; Perez-Lozano, M.J.; Varela, J.M.; Calderon, E.J. Pneumocystis jirovecii in general population. Emerg. Infect. Dis. 2005, 11, 245–250. [Google Scholar] [CrossRef]
- Vargas, S.L.; Ponce, C.A.; Sanchez, C.A.; Ulloa, A.V.; Bustamante, R.; Juarez, G. Pregnancy and asymptomatic carriage of Pneumocystis jiroveci. Emerg. Infect. Dis. 2003, 9, 605–606. [Google Scholar] [CrossRef] [PubMed]
- Le Gal, S.; Bonnet, P.; Huguenin, A.; Chapelle, C.; Boulic, P.; Tonnelier, J.-M.; Moal, M.-C.; Gut-Gobert, C.; Barnier, A.; Nevez, G. The shift from pulmonary colonization to Pneumocystis pneumonia. Med. Mycol. 2021, 59, 510–513. [Google Scholar] [CrossRef] [PubMed]
- Maskell, N.A.; Waine, D.J.; Lindley, A.; Pepperell, J.C.T.; Wakefield, A.E.; Miller, R.F.; Davies, R.J.O. Asymptomatic carriage of Pneumocystis jiroveci in subjects undergoing bronchoscopy: A prospective study. Thorax 2003, 58, 594–597. [Google Scholar] [CrossRef] [PubMed]
- Nevez, G.; Magois, E.; Duwat, H.; Gouilleux, V.; Jounieaux, V.; Totet, A. Apparent absence of Pneumocystis jirovecii in healthy subjects. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2006, 42, e99–e101. [Google Scholar] [CrossRef]
- Ponce, C.A.; Gallo, M.; Bustamante, R.; Vargas, S.L. Pneumocystis colonization is highly prevalent in the autopsied lungs of the general population. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2010, 50, 347–353. [Google Scholar] [CrossRef]
- Keely, S.P.; Stringer, J.R.; Baughman, R.P.; Linke, M.J.; Walzer, P.D.; Smulian, A.G. Genetic variation among Pneumocystis carinii hominis isolates in recurrent pneumocystosis. J. Infect. Dis. 1995, 172, 595–598. [Google Scholar] [CrossRef]
- Tsolaki, A.G.; Miller, R.F.; Underwood, A.P.; Banerji, S.; Wakefield, A.E. Genetic diversity at the internal transcribed spacer regions of the rRNA operon among isolates of Pneumocystis carinii from AIDS patients with recurrent pneumonia. J. Infect. Dis. 1996, 174, 141–156. [Google Scholar] [CrossRef]
- Vargas, S.L.; Ponce, C.A.; Gallo, M.; Pérez, F.; Astorga, J.-F.; Bustamante, R.; Chabé, M.; Durand-Joly, I.; Iturra, P.; Miller, R.F.; et al. Near-universal prevalence of Pneumocystis and associated increase in mucus in the lungs of infants with sudden unexpected death. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2013, 56, 171–179. [Google Scholar] [CrossRef]
- Miller, R.F.; Ambrose, H.E.; Novelli, V.; Wakefield, A.E. Probable mother-to-infant transmission of Pneumocystis carinii f. sp. hominis infection. J. Clin. Microbiol. 2002, 40, 1555–1557. [Google Scholar] [CrossRef]
- Rodríguez Álvarez, M.; Sosa González, L.C.; Marcos Rodríguez, A.; de Araña, M.J.; AlermGonzález, A.; León Pimentel, A. Evaluación de niveles séricos de citocinas proinflamatorias y marcadores de estrés oxidativo en mujeres embarazadas a término. Rev. Cuba Endocrinol. 2005, 16, 1–12. [Google Scholar]
- Djawe, K.; Daly, K.R.; Vargas, S.L.; Santolaya, M.E.; Ponce, C.A.; Bustamante, R.; Koch, J.; Levin, L.; Walzer, P.D. Seroepidemiological study of Pneumocystis jirovecii infection in healthy infants in Chile using recombinant fragments of the P. jirovecii major surface glycoprotein. Int. J. Infect. Dis. IJID Off. Publ. Int. Soc. Infect. Dis. 2010, 14, e1060–e1066. [Google Scholar] [CrossRef] [PubMed]
- Monroy-Vaca, E.X.; de Armas, Y.; Illnait-Zaragozí, M.T.; Toraño, G.; Diaz, R.; Vega, D.; Alvarez-Lam, I.; Calderón, E.J.; Stensvold, C.R. Prevalence and genotype distribution of Pneumocystis jirovecii in Cuban infants and toddlers with whooping cough. J. Clin. Microbiol. 2014, 52, 45–51. [Google Scholar] [CrossRef][Green Version]
- Respaldiza, N.; Medrano, F.J.; Medrano, A.C.; Varela, J.M.; de la Horra, C.; Montes-Cano, M.; Ferrer, S.; Wichmann, I.; Gargallo-Viola, D.; Calderon, E.J. High seroprevalence of Pneumocystis infection in Spanish children. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2004, 10, 1029–1031. [Google Scholar] [CrossRef] [PubMed]
- Vargas, S.L.; Hughes, W.T.; Santolaya, M.E.; Ulloa, A.V.; Ponce, C.A.; Cabrera, C.E.; Cumsille, F.; Gigliotti, F. Search for primary infection by Pneumocystis carinii in a cohort of normal, healthy infants. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2001, 32, 855–861. [Google Scholar] [CrossRef] [PubMed]
- Vera, C.; Aguilar, Y.A.; Vélez, L.A.; Rueda, Z.V. High transient colonization by Pneumocystis jirovecii between mothers and newborn. Eur. J. Pediatr. 2017, 176, 1619–1627. [Google Scholar] [CrossRef]
- Garcia, C.; Ochoa, T.; Neyra, E.; Bustamante, B.; Ponce, C.; Calderon, E.J.; Vargas, S.L. Pneumocystis jirovecii colonisation in pregnant women and newborns in Lima, Peru. Rev. Iberoam Micol. 2020, 37, 24–27. [Google Scholar] [CrossRef] [PubMed]
- Nevez, G.; Totet, A.; Pautard, J.C.; Raccurt, C. Pneumocystis carinii detection using nested-PCR in nasopharyngeal aspirates of immunocompetent infants with bronchiolitis. J. Eukaryot. Microbiol. 2001, 122S–123S. [Google Scholar] [CrossRef]
- Totet, A.; Latouche, S.; Duwat, H.; Magois, E.; Lacube, P.; Pautard, J.-C.; Schmit, J.-L.; Jounieaux, V.; Roux, P.; Raccurt, C.; et al. Multilocus genotyping of Pneumocystis jirovecii in patients developing diverse forms of parasitism: Implication for a wide human reservoir for the fungus. J. Eukaryot. Microbiol. 2003, 50, 670–671. [Google Scholar] [CrossRef]
- Leigh, T.R.; Kangro, H.O.; Gazzard, B.G.; Jeffries, D.J.; Collins, J.V. DNA amplification by the polymerase chain reaction to detect sub-clinical Pneumocystis carinii colonization in HIV-positive and HIV-negative male homosexuals with and without respiratory symptoms. Respir. Med. 1993, 87, 525–529. [Google Scholar] [CrossRef]
- Matos, O.; Costa, M.C.; Lundgren, B.; Caldeira, L.; Aguiar, P.; Antunes, F. Effect of oral washes on the diagnosis of Pneumocystis carinii pneumonia with a low parasite burden and on detection of organisms in subclinical infections. Eur. J. Clin. Microbiol. Infect Dis. Off. Publ. Eur. Soc. Clin. Microbiol. 2001, 20, 573–575. [Google Scholar] [CrossRef] [PubMed]
- Helweg-Larsen, J.; Jensen, J.S.; Dohn, B.; Benfield, T.L.; Lundgren, B. Detection of Pneumocystis DNA in samples from patients suspected of bacterial pneumonia—A case-control study. BMC Infect. Dis. 2002, 2, 28. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Crothers, K.; Morris, A.; Groner, G.; Fox, M.; Turner, J.R.; Merrifield, C.; Eiser, S.; Zucchi, P.; Beard, C.B. Pneumocystis colonization in HIV-infected patients. J. Eukaryot. Microbiol. 2003, 50, 616–617. [Google Scholar] [CrossRef] [PubMed]
- Vidal, S.; de la Horra, C.; Martín, J.; Montes-Cano, M.A.; Rodríguez, E.; Respaldiza, N.; Rodríguez, F.; Varela, J.M.; Medrano, F.J.; Calderón, E.J. Pneumocystis jirovecii colonisation in patients with interstitial lung disease. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2006, 12, 231–235. [Google Scholar] [CrossRef]
- Matos, O.; Costa, M.; Correia, I.; Monteiro, P.; Vieira, J.R.; Soares, J.; Bonnet, M.; Esteves, F.; Antunes, F. Infecção por Pneumocystis jirovecii em imunocompetentes com patologia pulmonar, em Portugal. Acta Med. Port. 2006, 19, 121–126. [Google Scholar]
- Morris, A.; Sciurba, F.C.; Lebedeva, I.P.; Githaiga, A.; Elliott, W.M.; Hogg, J.C.; Huang, L.; Norris, K.A. Association of chronic obstructive pulmonary disease severity and Pneumocystis colonization. Am. J. Respir. Crit. Care Med. 2004, 170, 408–413. [Google Scholar] [CrossRef]
- Calderón, E.J.; Rivero, L.; Respaldiza, N.; Morilla, R.; Montes-Cano, M.A.; Friaza, V.; Muñoz-Lobato, F.; Varela, J.M.; Medrano, F.J.; de la Horra, C. Systemic Inflammation in Patients with Chronic Obstructive Pulmonary Disease Who Are Colonized with Pneumocystis jiroveci. Clin. Infect. Dis. 2007, 45, e17–e19. [Google Scholar] [CrossRef]
- Gal, S.L.; Héry-Arnaud, G.; Ramel, S.; Virmaux, M.; Damiani, C.; Totet, A.; Nevez, G. Pneumocystis jirovecii and cystic fibrosis in France. Scand. J. Infect. Dis. 2010, 42, 225–227. [Google Scholar] [CrossRef]
- Mekinian, A.; Durand-Joly, I.; Hatron, P.-Y.; Moranne, O.; Denis, G.; Dei-Cas, E.; Morell-Dubois, S.; Lambert, M.; Launay, D.; Delhaes, L.; et al. Pneumocystis jirovecii colonization in patients with systemic autoimmune diseases: Prevalence, risk factors of colonization and outcome. Rheumatol. Oxf. Engl. 2011, 50, 569–577. [Google Scholar] [CrossRef]
- Pereira, R.M.; Müller, A.L.; Zimerman, R.A.; Antunes, D.B.; Zinn, V.F.; Friaza, V.; de la Horra, C.; Calderón, E.J.; Wissmann, G. High prevalence of Pneumocystis jirovecii colonization among HIV-positive patients in southern Brazil. Med. Mycol. 2014, 52, 804–809. [Google Scholar] [CrossRef]
- Wang, D.-D.; Zheng, M.-Q.; Zhang, N.; An, C.-L. Investigation of Pneumocystis jirovecii colonization in patients with chronic pulmonary diseases in the People’s Republic of China. Int. J. Chron Obstruct. Pulmon Dis. 2015, 10, 2079–2085. [Google Scholar] [PubMed]
- Morris, A.; Wei, K.; Afshar, K.; Huang, L. Epidemiology and clinical significance of pneumocystis colonization. J. Infect. Dis. 2008, 197, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Respaldiza, N.; Montes-Cano, M.A.; Dapena, F.J.; de la Horra, C.; Mateos, I.; Medrano, F.J.; Calderon, E.; Varela, J.M. Prevalence of colonisation and genotypic characterisation of Pneumocystis jirovecii among cystic fibrosis patients in Spain. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2005, 11, 1012–1015. [Google Scholar] [CrossRef] [PubMed]
- Sivam, S.; Sciurba, F.C.; Lucht, L.A.; Zhang, Y.; Duncan, S.R.; Norris, K.A.; Morris, A. Distribution of Pneumocystis jirovecii in lungs from colonized COPD patients. Diagn. Microbiol. Infect. Dis. 2011, 71, 24–28. [Google Scholar] [CrossRef]
- Fitzpatrick, M.E.; Tedrow, J.R.; Hillenbrand, M.E.; Lucht, L.; Richards, T.; Norris, K.A.; Zhang, Y.; Sciurba, F.C.; Kaminski, N.; Morris, A. Pneumocystis jirovecii colonization is associated with enhanced Th1 inflammatory gene expression in lungs of humans with chronic obstructive pulmonary disease. Microbiol. Immunol. 2014, 58, 202–211. [Google Scholar] [CrossRef] [PubMed]
- Morris, A.; Sciurba, F.C.; Norris, K.A. Pneumocystis: A novel pathogen in chronic obstructive pulmonary disease? COPD 2008, 5, 43–51. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Author | Year | Country | Patients | n | Colonization Percentage | Biological Sample 2 | Diagnostic Technique 3 |
|---|---|---|---|---|---|---|---|
| Leigh TR [91]. | 1993 | UK | HET 1, HOM 1 and HIV-seropositive | 90 | Between 10 and 40%. | IS | PCR |
| Matos O [92]. | 2001 | Portugal | Adults HIV-infected | 104 | 14.5% | IS and OW | PCR and conventional stain |
| Helweg-Larsen J [93]. | 2002 | Denmark | Adults with suspected bacterial pneumoniae | 367 | 4.0% | BAL, tracheal aspirates and sputum | PCR |
| Vargas SL [73] | 2003 | Chile | Immunocompetent adult, pregnant and non-pregnant women | 33 and 28, respectively | 15.0% and 0.0%, respectively | Nasopharyngeal aspirates | PCR |
| Huang L. [94]. | 2003 | EEUU | HIV-infected Adults | 32 | 69% | IS or BAL | PCR |
| Maskell NA [75]. | 2003 | EEUU | Immunosuppressed HIV-negative adults | 93 | 18.0% | BAL | PCR |
| Vidal S [95]. | 2006 | Spain | Adults with Interstitial lung disease | 80 | 33.8% | BAL | PCR |
| Matos O [96]. | 2006 | Portugal | Immunocompetent adults with pulmonary disease and | 45 | 24.4 | BAL | PCR |
| Nevez G [76]. | 2006 | France | Adults with COPD and healthy adults | 50 and 30, respectively | 16% and 0%, respectively | Sputum | PCR |
| Morris A [97]. | 2006 | EEUU | Adults with COPD | 68 | 19.1% | Lung tissue Pendiente | PCR |
| Calderón EJ [98]. | 2007 | Spain | Adults with COPD | 51 | 55.0% | Sputum | PCR |
| Gal SL [99]. | 2010 | France | Adults and children with cystic fibrosis | 76 patients and 146 samples | 1.3% | Sputum | PCR |
| Mekinian A [100]. | 2011 | France | Adults with systemic autoimmune diseases | 67 | 16.0% | IS | PCR |
| Pereira RM [101]. | 2014 | Brazil | HIV-seropositive Adults | 58 | 44.8% | Oropharyngeal samples | PCR |
| Wang D-D [102] | 2015 | China | Adults with chronic pulmonary diseases | 98 | 63.3 | Sputum | LAMP and PCR |
| Vera C [87] | 2017 | Colombia | Immunocompetent and pregnant women and newborns | 43 and 43, respectively | 46.5% and 74.4%, respectively | NPS | PCR |
| García C. [88] | 2020 | Perú | HIV-negative women and newborns | 92 and 87 | 5.43% and 0.0%, respectively | OW, NPS, and placenta samples | PCR |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vera, C.; Rueda, Z.V. Transmission and Colonization of Pneumocystis jirovecii. J. Fungi 2021, 7, 979. https://doi.org/10.3390/jof7110979
Vera C, Rueda ZV. Transmission and Colonization of Pneumocystis jirovecii. Journal of Fungi. 2021; 7(11):979. https://doi.org/10.3390/jof7110979
Chicago/Turabian StyleVera, Cristian, and Zulma Vanessa Rueda. 2021. "Transmission and Colonization of Pneumocystis jirovecii" Journal of Fungi 7, no. 11: 979. https://doi.org/10.3390/jof7110979
APA StyleVera, C., & Rueda, Z. V. (2021). Transmission and Colonization of Pneumocystis jirovecii. Journal of Fungi, 7(11), 979. https://doi.org/10.3390/jof7110979