Effect of Juvenile Hormone on Resistance against Entomopathogenic Fungus Metarhizium robertsii Differs between Sexes


 , and
, and 
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Animals
2.2. Experimental Treatments
2.3. Statistical Analyses
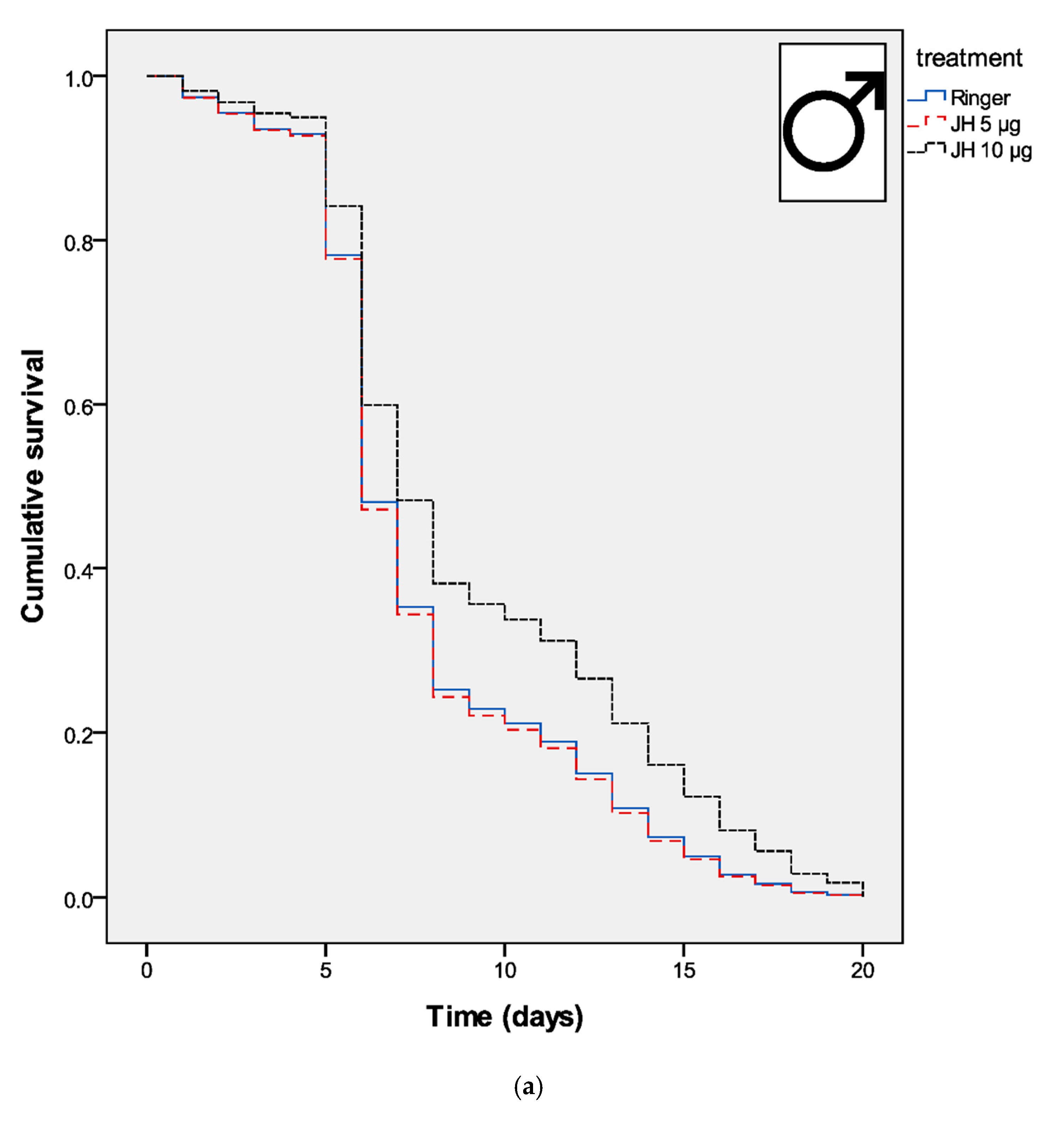
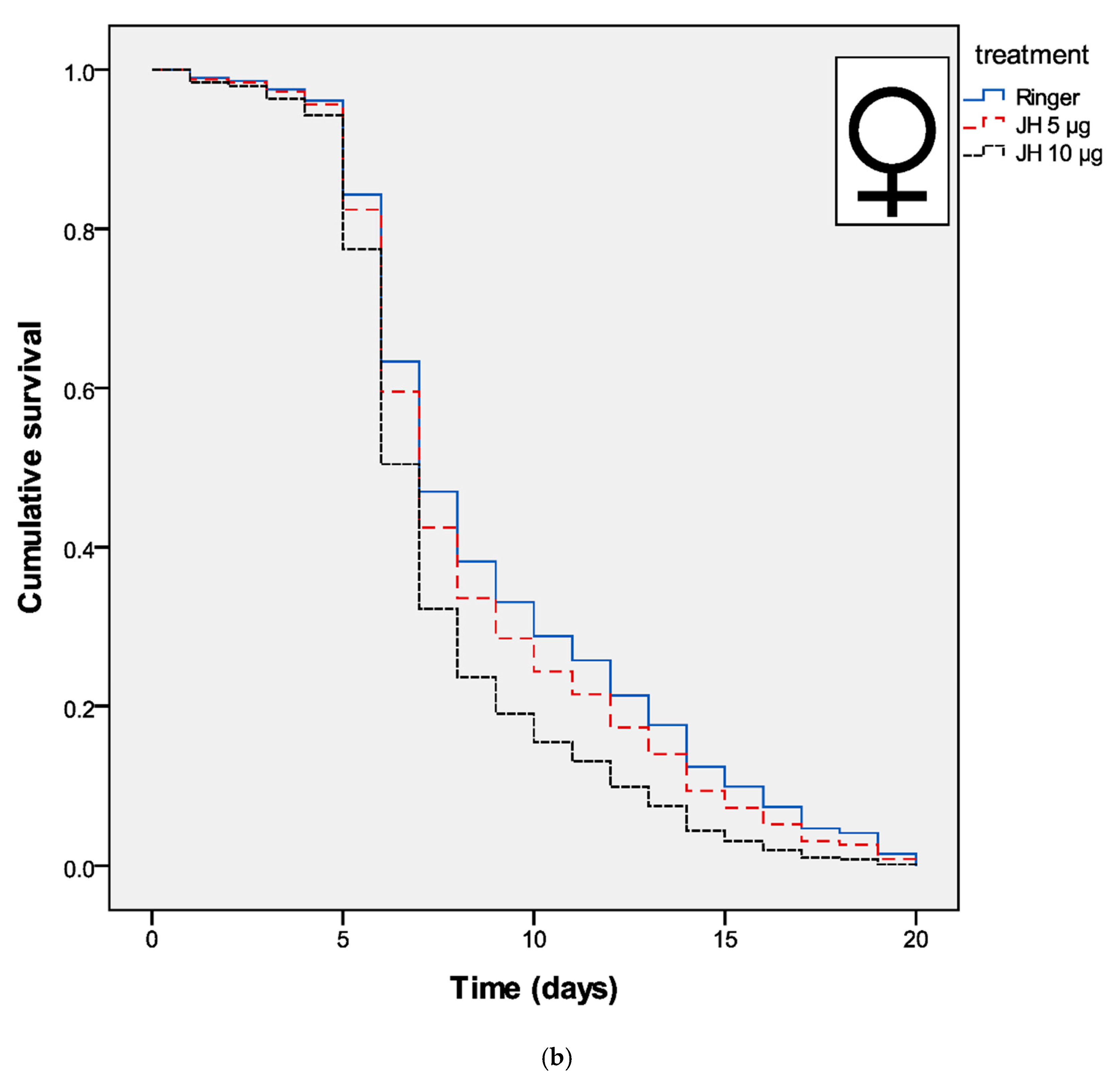
3. Results
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Folstad, I.; Karter, A.J. Parasites, bright males, and the immunocompetence handicap. Am. Nat. 1992, 139, 603–622. [Google Scholar] [CrossRef]
- Roberts, M.L.; Buchanan, K.L.; Evans, M.R. Testing the immunocompetence handicap hypothesis: A review of the evidence. Anim. Behav. 2004, 68, 227–239. [Google Scholar] [CrossRef]
- Rantala, M.; Koskimaki, J.; Taskinen, J.; Tynkkynen, K.; Suhonen, J. Immunocompetence, developmental stability and wingspot size in the damselfly Calopteryx splendens L. Proc. R. Soc. B Biol. Sci. 2000, 267, 2453–2457. [Google Scholar] [CrossRef]
- Rantala, M.; Jokinen, I.; Kortet, R.; Vainikka, A.; Suhonen, J. Do pheromones reveal male immunocompetence? Proc. R. Soc. B Biol. Sci. 2002, 269, 1681–1685. [Google Scholar] [CrossRef] [PubMed]
- Siva-Jothy, M. A mechanistic link between parasite resistance and expression of a sexually selected trait in a damselfly. Proc. R. Soc. B Biol. Sci. 2000, 267, 2523–2527. [Google Scholar] [CrossRef] [PubMed]
- Ahtiainen, J.; Alatalo, R.; Kortet, R.; Rantala, M. Sexual advertisement and immune function in an arachnid species (Lycosidae). Behav. Ecol. 2004, 15, 602–606. [Google Scholar] [CrossRef]
- Krams, I.; Daukšte, J.; Kivleniece, I.; Krama, T.; Rantala, M.J. Previous encapsulation response enhances within individual protection against fungal parasite in the mealworm beetle Tenebrio molitor. Insect Sci. 2013, 20, 771–777. [Google Scholar] [CrossRef]
- Rantala, M.; Vainikka, A.; Kortet, R. The role of juvenile hormone in immune function and pheromone production trade-offs: A test of the immunocompetence handicap principle. Proc. R. Soc. B Biol. Sci. 2003, 270, 2257–2261. [Google Scholar] [CrossRef]
- Sreng, L.; Leoncini, I.; Clement, J.L. Regulation of sex pheromone production in the male Nauphoeta cinerea cockroach: Role of brain extracts, corpora allata (CA), and juvenile hormone (JH). Arch. Insect Biochem. Physiol. 1999, 40, 165–172. [Google Scholar] [CrossRef]
- Marquez-Garcia, A.; Canales-Lazcano, J.; Rantala, M.J.; Contreras-Garduno, J. Is juvenile hormone a potential mechanism that underlay the “branched Y-model”? Gen. Comp. Endocrinol. 2016, 230–231, 170–176. [Google Scholar] [CrossRef]
- Contreras-Garduno, J.; Cordoba-Aguilar, A.; Lanz-Mendoza, H.; Rivera, A.C. Territorial behaviour and immunity are mediated by juvenile hormone: The physiological basis of honest signalling? Funct. Ecol. 2009, 23, 157–163. [Google Scholar] [CrossRef]
- Flatt, T.; Tu, M.P.; Tatar, M. Hormonal pleiotropy and the juvenile hormone regulation of Drosophila development and life history. BioEssays 2005, 27, 999–1010. [Google Scholar] [CrossRef] [PubMed]
- Kwon, S.; Kim, Y. Immunosuppressive action of pyriproxyfen, a juvenile hormone analog, enhances pathogenicity of Bacillus thuringiensis subsp kurstaki against diamondback moth, Plutella xylostella (Lepidoptera: Yponoineutidae). Biol. Control 2007, 42, 72–76. [Google Scholar] [CrossRef]
- Lavine, M.D.; Strand, M.R. Insect hemocytes and their role in immunity. Insect Biochem. Mol. Biol. 2002, 32, 1295–1309. [Google Scholar] [CrossRef]
- Rolff, J.; Siva-Jothy, M. Copulation corrupts immunity: A mechanism for a cost of mating in insects. Proc. Natl. Acad. Sci. USA 2002, 99, 9916–9918. [Google Scholar] [CrossRef]
- Valtonen, T.M.; Viitaniemi, H.; Rantala, M.J. Copulation enhances resistance against an entomopathogenic fungus in the mealworm beetle Tenebrio molitor. Parasitology 2010, 137, 985–989. [Google Scholar] [CrossRef]
- Butt, T.M.; Coates, C.J.; Dubovskiy, I.M.; Ratcliffe, N.A. Chapter nine—Entomopathogenic fungi: New insights into host–pathogen interactions. Adv. Gen. 2016, 94, 307–364. [Google Scholar]
- Krams, I.; Daukšte, J.; Kivleniece, I.; Krama, T.; Rantala, M.J.; Ramey, G.; Šauša, L. Female choice reveals terminal investment in male mealworm beetles, Tenebrio molitor, after a repeated activation of the immune system. J. Insect Sci. 2011, 11, 56. [Google Scholar] [CrossRef]
- Mukherjee, K.; Dubovskiy, I.; Grizanova, E.; Lehmann, R.; Vilcinskas, A. Epigenetic mechanisms mediate the experimental evolution of resistance against parasitic fungi in the greater wax moth Galleria mellonella. Sci. Rep. 2019, 9, 1626. [Google Scholar] [CrossRef]
- Braude, S.; Tang-Martinez, Z.; Taylor, G.T. Stress, testosterone, and the immunoredistribution hypothesis. Behav. Ecol. 1999, 10, 345–350. [Google Scholar] [CrossRef]
- Zuk, M.; Stoehr, A.M. Immune defence and host life history. Am. Nat. 2002, 160, S9–S22. [Google Scholar] [CrossRef] [PubMed]
- Kecko, S.; Mihailova, A.; Kangassalo, K.; Elferts, D.; Krama, T.; Krams, R.; Luoto, S.; Rantala, M.J.; Krams, I.A. Sex-specific compensatory growth in the larvae of the greater wax moth Galleria mellonella. J. Evol. Biol. 2017, 30, 1910–1918. [Google Scholar] [CrossRef] [PubMed]
- Sheridan, L.A.D.; Poulin, R.; Ward, D.F.; Zuk, M. Sex differences in parasitic infections among arthropod hosts: Is there a male bias? Oikos 2000, 88, 327–334. [Google Scholar] [CrossRef]
- Rantala, M.J.; Roff, D.A.; Rantala, M.J. Forceps size and immune function in the earwig Forficula auricularia L. Biol. J. Linn. Soc. 2007, 90, 509–516. [Google Scholar] [CrossRef]
- McKean, K.A.; Nunney, L. Bateman’s principle and immunity: Phenotypically plastic reproductive strategies predict changes in immunological sex differences. Evolution 2005, 59, 1510–1517. [Google Scholar] [CrossRef]
- Rantala, M.; Roff, D. Inbreeding and extreme outbreeding cause sex differences in immune defence and life history traits in Epirrita autumnata. Heredity 2007, 98, 329–336. [Google Scholar] [CrossRef]
- Vilcinskas, A.; Matha, V. Effect of the entomopathogenic fungus Beauveria bassiana on the humoral immune response of Galleria mellonella larvae (Lepidoptera: Pyralidae). Eur. J. Entomol. 1997, 94, 461–472. [Google Scholar]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rantala, M.J.; Dubovskiy, I.M.; Pölkki, M.; Krama, T.; Contreras-Garduño, J.; Krams, I.A. Effect of Juvenile Hormone on Resistance against Entomopathogenic Fungus Metarhizium robertsii Differs between Sexes. J. Fungi 2020, 6, 298. https://doi.org/10.3390/jof6040298
Rantala MJ, Dubovskiy IM, Pölkki M, Krama T, Contreras-Garduño J, Krams IA. Effect of Juvenile Hormone on Resistance against Entomopathogenic Fungus Metarhizium robertsii Differs between Sexes. Journal of Fungi. 2020; 6(4):298. https://doi.org/10.3390/jof6040298
Chicago/Turabian StyleRantala, Markus J., Ivan M. Dubovskiy, Mari Pölkki, Tatjana Krama, Jorge Contreras-Garduño, and Indrikis A. Krams. 2020. "Effect of Juvenile Hormone on Resistance against Entomopathogenic Fungus Metarhizium robertsii Differs between Sexes" Journal of Fungi 6, no. 4: 298. https://doi.org/10.3390/jof6040298
APA StyleRantala, M. J., Dubovskiy, I. M., Pölkki, M., Krama, T., Contreras-Garduño, J., & Krams, I. A. (2020). Effect of Juvenile Hormone on Resistance against Entomopathogenic Fungus Metarhizium robertsii Differs between Sexes. Journal of Fungi, 6(4), 298. https://doi.org/10.3390/jof6040298