In Vitro and In Vivo Inhibitory Activity of Limonene against Different Isolates of Candida spp.

 ,
,  , ,
, ,  , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Material and Methods
2.1. Animals
2.2. Candida albicans Strains
2.3. Antimicrobial Compounds
2.4. Conditions of Limonene Exposure for Growth Inhibition Assays
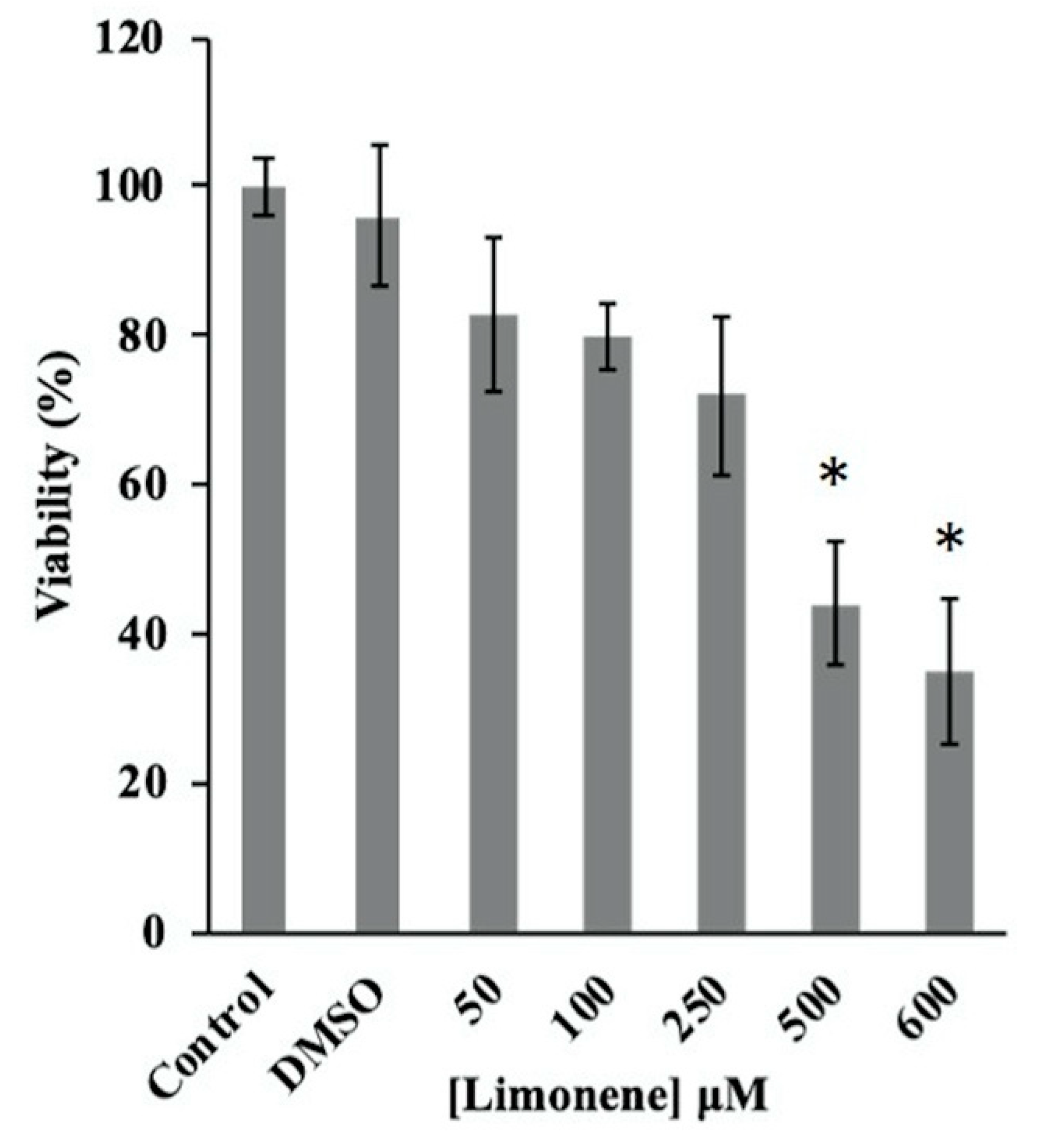
2.5. MTT Assay
2.6. Intravaginal Infection of BALB/c Mice
2.7. Assay for Colony-Forming Units (CFUs)
2.8. Histopathology
2.9. Transmission Electron Microscopy (TEM)
2.10. Scanning Electron Microscopy (SEM)
2.11. Statistical Analysis
3. Results
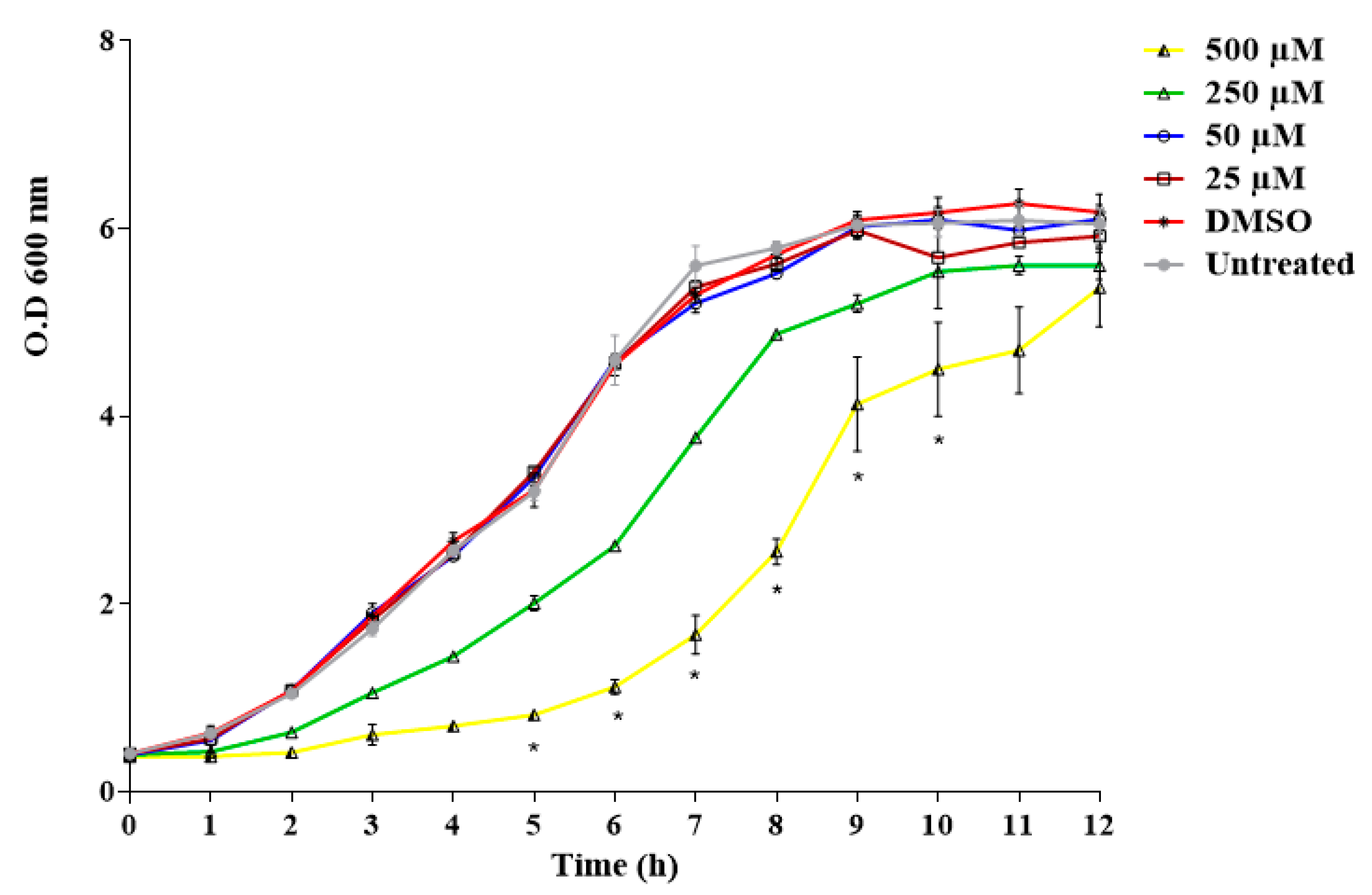
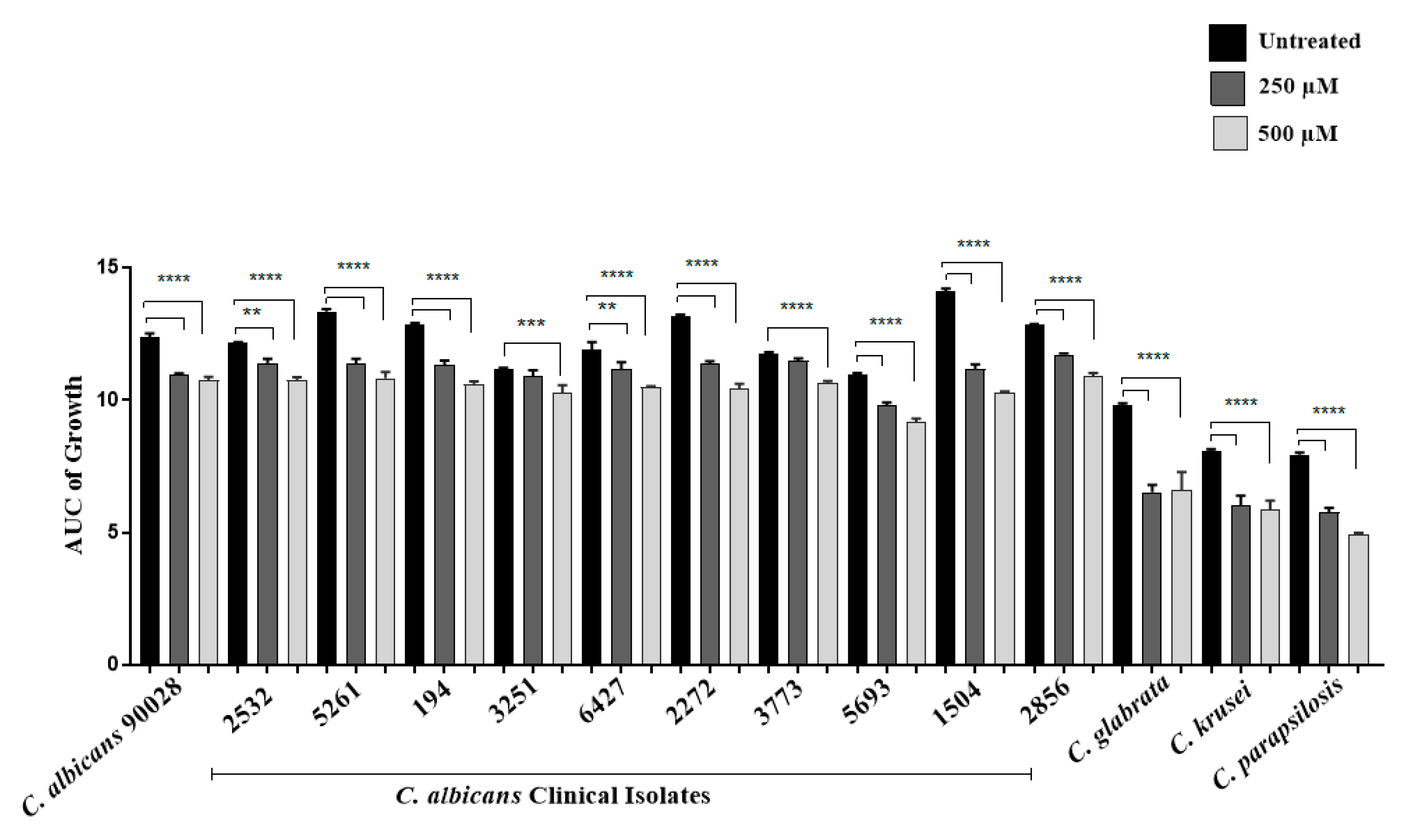
3.1. Evaluation of the Antifungal Activity of Limonene In Vitro
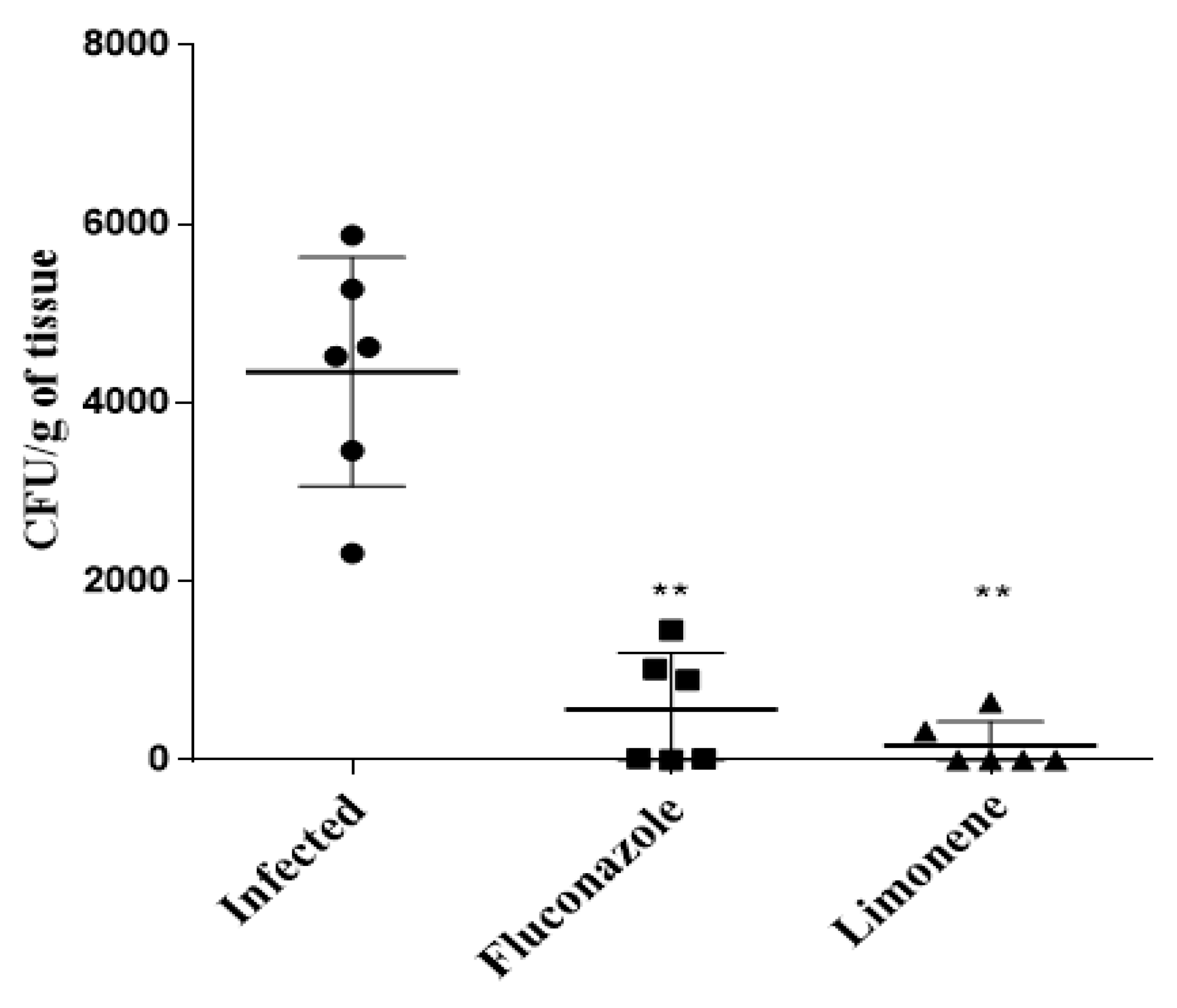
3.2. Treatment with Limonene Reduces the Fungal Burden in Mice with Vaginal Candidiasis
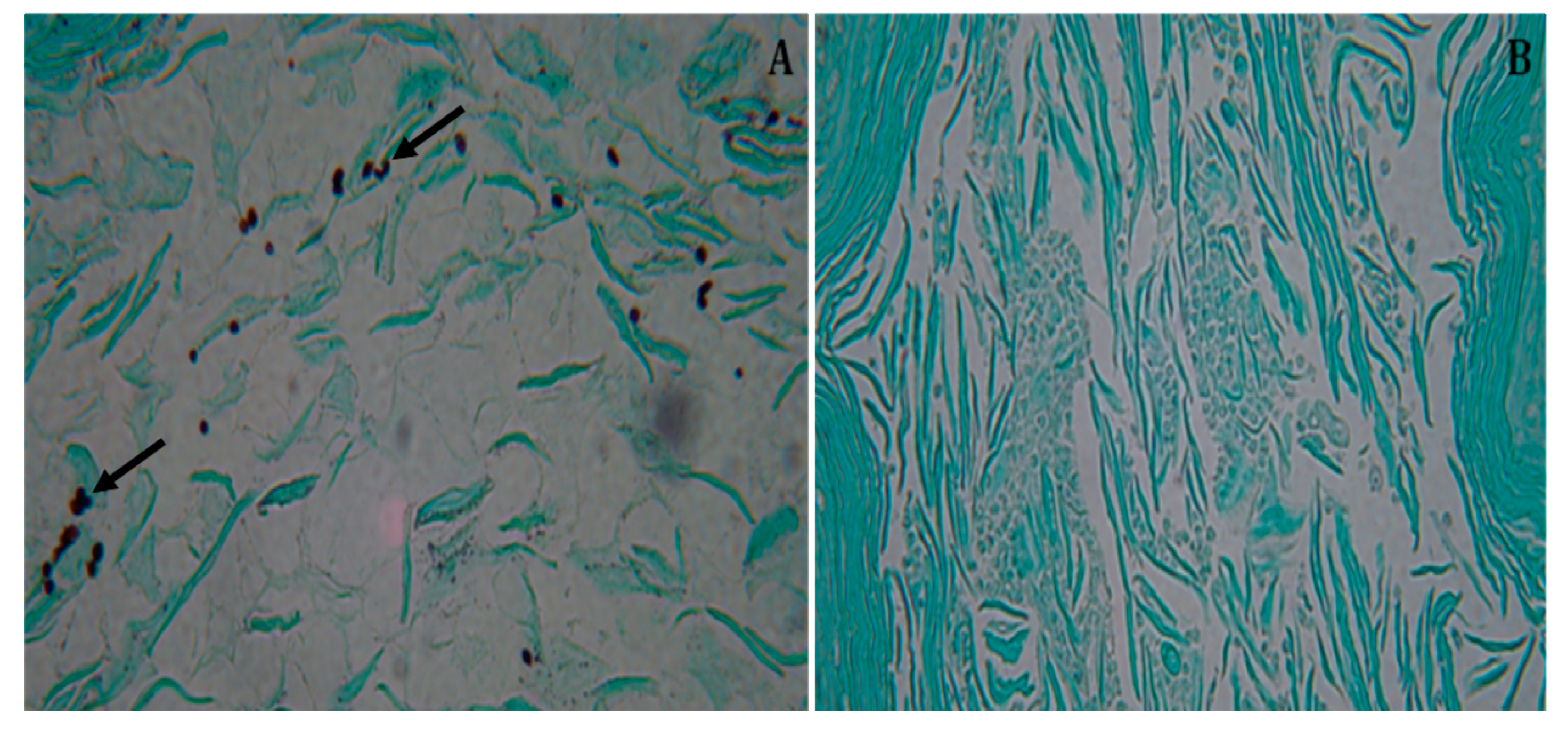
3.3. Histopathology of the Vaginal canal in Treated, Intravaginally Infected, BALB/c Mice
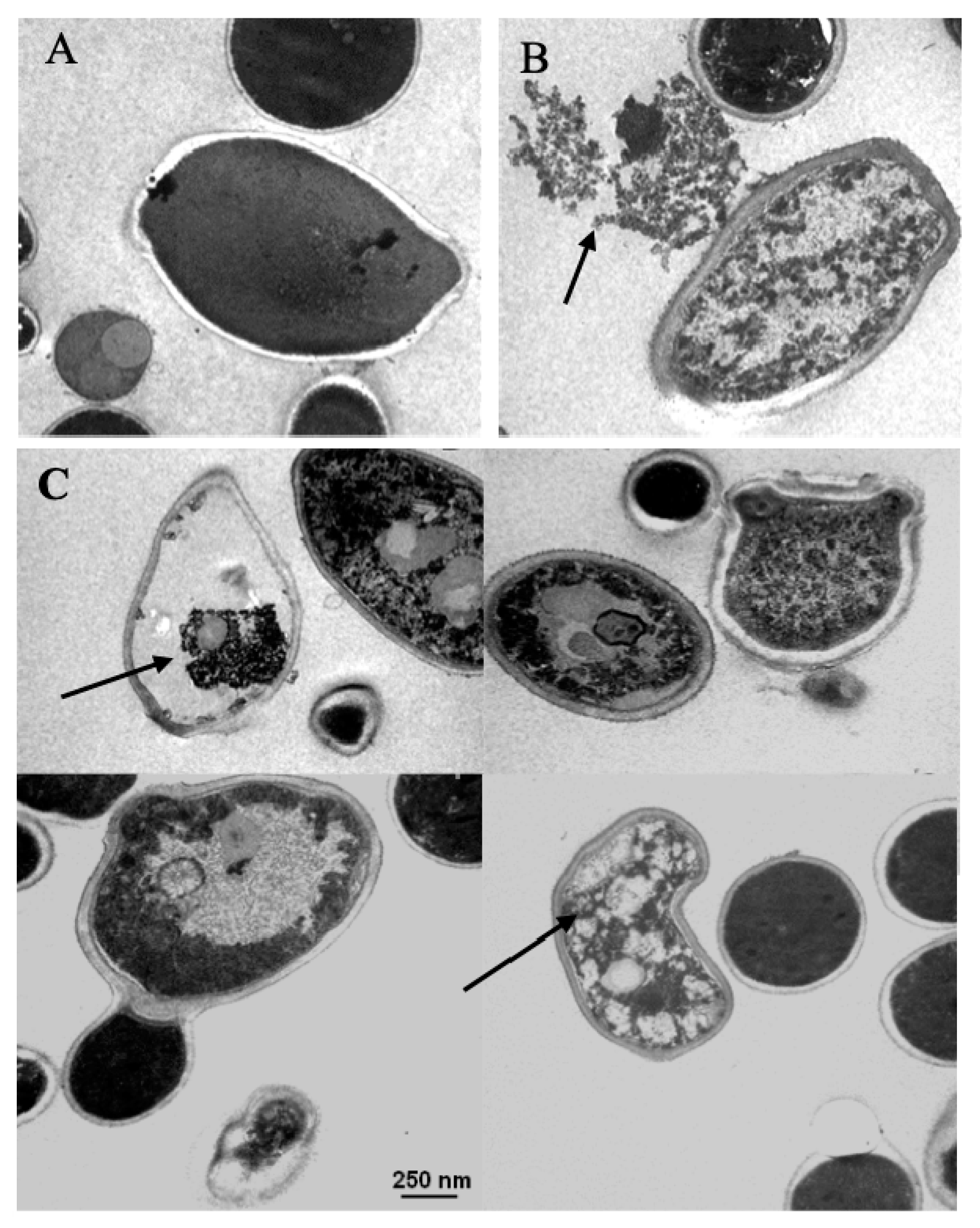
3.4. Transmission Electron Microscopy (TEM)
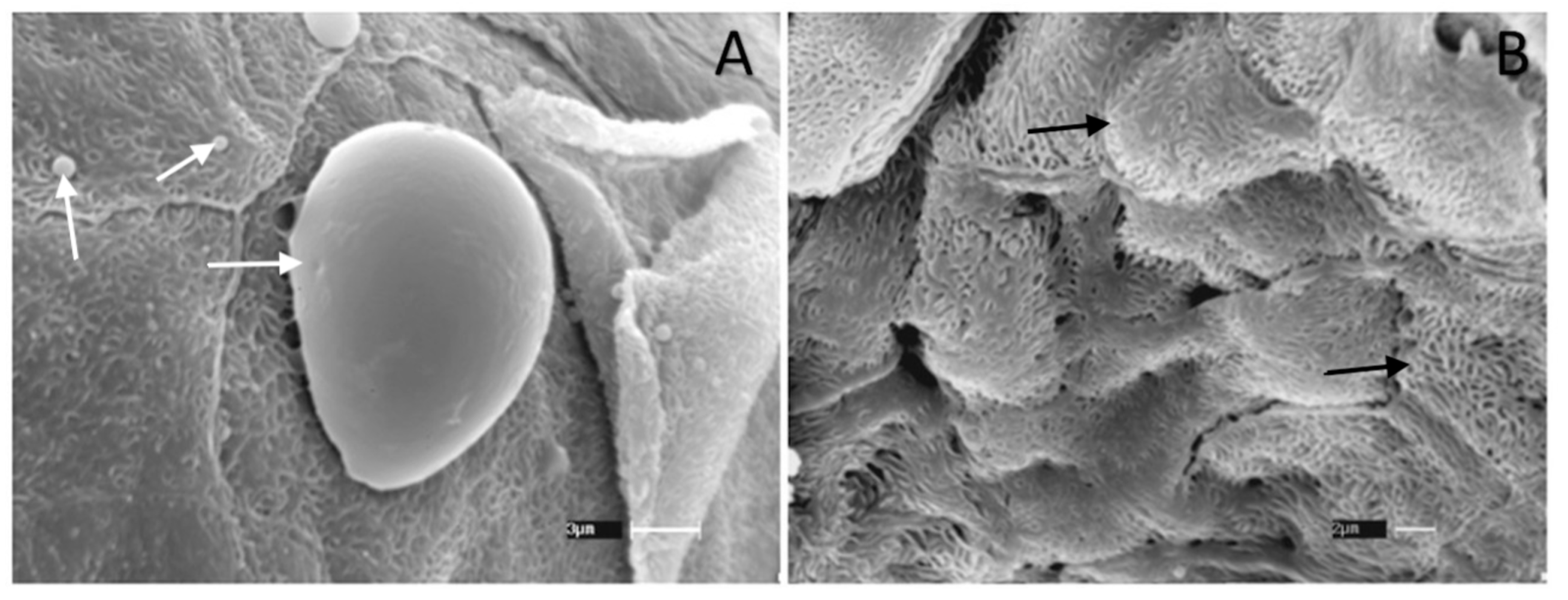
3.5. Scanning Electron Microscopy (SEM)
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Miceli, M.H.; Díaz, J.A.; Lee, S.A. Emerging opportunistic yeast infections. Lancet Infect. Dis. 2011, 11, 142–151. [Google Scholar] [CrossRef]
- Alvarez-Moreno, C.; Cortes, J.; Denning, D. Burden of Fungal Infections in Colombia. J. Fungi 2018, 4, 41. [Google Scholar] [CrossRef]
- Vila, T.; Sultan, A.S.; Montelongo-Jauregui, D.; Jabra-Rizk, M.A. Oral candidiasis: A disease of opportunity. J. Fungi 2020, 6, 15. [Google Scholar] [CrossRef]
- Shapiro, R.S.; Robbins, N.; Cowen, L.E. Regulatory circuitry governing fungal development, drug resistance, and disease. Microbiol. Mol. Biol. Rev. 2011, 75, 213–267. [Google Scholar] [CrossRef]
- Arruda, D.C.; Alexandri, F.L.; Katzin, A.M.; Uliana, S.R.B. Antileishmanial Activity of the Terpene Nerolidol. Antimicrob. Agents Chemother. 2005, 49, 1679–1687. [Google Scholar] [CrossRef]
- Slade, D.; Galal, A.M.; Gul, W.; Radwan, M.M.; Ahmed, S.A.; Khan, S.I.; Tekwani, B.L.; Jacob, M.R.; Ross, S.A.; ElSohly, M.A. Antiprotozoal, anticancer and antimicrobial activities of dihydroartemisinin acetal dimers and monomers. Bioorganic Med. Chem. 2009, 17, 7949–7957. [Google Scholar] [CrossRef]
- Matsuo, A.L.; Figueiredo, C.R.; Arruda, D.C.; Pereira, F.V.; Borin Scutti, J.A.; Massaoka, M.H.; Travassos, L.R.; Sartorelli, P.; Lago, J.H.G. α-Pinene isolated from Schinus terebinthifolius Raddi (Anacardiaceae) induces apoptosis and confers antimetastatic protection in a melanoma model. Biochem. Biophys. Res. Commun. 2011, 411, 449–454. [Google Scholar] [CrossRef]
- Langford, M.L.; Atkin, A.L.; Nickerson, K.W. Cellular interactions of farnesol, a quorum-sensing molecule produced by Candida albicans. Future Microbiol. 2009, 4, 1353–1362. [Google Scholar] [CrossRef]
- Ricci, D.; Braga, P.C. Atomic Force Microscopy in Biomedical Research. Methods Mol. Biol. 2011, 736, 355–377. [Google Scholar]
- Elson, C.E.; Maltzman, T.H.; Boston, J.L.; Tanner, M.A.; Gould, M.N. Anti-carcinogenic activity of d-limonene during the initiation and promotion/progression stages of dmba-induced rat mammary carcinogenesis. Carcinogenesis 1988, 9, 331–332. [Google Scholar] [CrossRef]
- Yu, X.; Lin, H.; Wang, Y.; Lv, W.; Zhang, S.; Qian, Y.; Deng, X.; Feng, N.; Yu, H.; Qian, B. D-limonene exhibits antitumor activity by inducing autophagy and apoptosis in lung cancer. Onco. Targets. Ther. 2018, 11, 1833–1847. [Google Scholar] [CrossRef]
- Yang, C.; Chen, H.; Chen, H.; Zhong, B.; Luo, X.; Chun, J. Antioxidant and anticancer activities of essential oil from gannan navel orange peel. Molecules 2017, 22, 1391. [Google Scholar] [CrossRef]
- Haag, J.D.; Lindstrom, M.J.; Gould, M.N. Limonene-induced Regression of Mammary Carcinomas. Cancer Res. 1992, 52, 4021–4026. [Google Scholar]
- Del Toro-Arreola, S.; Flores-Torales, E.; Torres-Lozano, C.; Del Toro-Arreola, A.; Tostado-Pelayo, K.; Ramirez-Dueñas, M.G.; Daneri-Navarro, A. Effect of D-limonene on immune response in BALB/c mice with lymphoma. Int. Immunopharmacol. 2005, 5, 829–838. [Google Scholar] [CrossRef]
- Lu, X.G.; Zhan, L.B.; Feng, B.A.; Qu, M.Y.; Yu, L.H.; Xie, J.H. Inhibition of growth and metastasis of human gastric cancer implanted in nude mice by d-limonene. World J. Gastroenterol. 2004, 10, 2140–2144. [Google Scholar] [CrossRef]
- Murali, R.; Karthikeyan, A.; Saravanan, R. Protective Effects of d-Limonene on Lipid Peroxidation and Antioxidant Enzymes in Streptozotocin-Induced Diabetic Rats. Basic Clin. Pharmacol. Toxicol. 2013, 112, 175–181. [Google Scholar] [CrossRef]
- Sakeena, M.H.F.; Elrashid, S.M.; Muthanna, F.A.; Ghassan, Z.A.; Kanakal, M.M.; Laila, L.; Munavvar, A.S.; Azmin, M.N. Effect of limonene on permeation enhancement of ketoprofen in palm oil esters nanoemulsion. J. Oleo Sci. 2010, 59, 395–400. [Google Scholar]
- Krishnaiah, Y.; Al-Saidan, S. Limonene Enhances the In Vitro and In Vivo Permeation of Trimetazidine Across a Membrane-Controlled Transdermal Therapeutic System. Curr. Drug Deliv. 2008, 5, 70–76. [Google Scholar] [CrossRef]
- Goulart, H.R.; Kimura, E.A.; Peres, V.J.; Couto, A.S.; Duarte, F.A.A.; Katzin, A.M. Terpenes arrest parasite development and inhibit biosynthesis of isoprenoids in Plasmodium falciparum. Antimicrob. Agents Chemother. 2004, 48, 2502–2509. [Google Scholar] [CrossRef]
- Arruda, D.C.; Miguel, D.C.; Yokoyama-Yasunaka, J.K.U.; Katzin, A.M.; Uliana, S.R.B. Inhibitory activity of limonene against Leishmania parasites in vitro and in vivo. Biomed. Pharmacother. 2009, 63, 643–649. [Google Scholar] [CrossRef]
- Moreno, É.M.; Leal, S.M.; Stashenko, E.E.; García, L.T. Induction of programmed cell death in Trypanosoma cruzi by Lippia alba essential oils and their major and synergistic terpenes (citral, limonene and caryophyllene oxide). BMC Complement. Altern. Med. 2018, 18, 1–16. [Google Scholar] [CrossRef]
- Hamada, M.; Uezu, K.; Matsushita, J.; Yamamoto, S.; Kishino, Y. Distribution and immune responses resulting from oral administration of D-limonene in rats. J. Nutr. Sci. Vitaminol. (Tokyo) 2002, 48, 155–160. [Google Scholar] [CrossRef]
- Evans, D.L.; Miller, D.M.; Jacobsen, K.L.; Bush, P.B. Modulation of immune responses in mice by d-limonene. J. Toxicol. Environ. Health 1987, 20, 51–66. [Google Scholar] [CrossRef]
- Hamad, M.; Abu-Elteen, K.H.; Ghaleb, M. Estrogen-dependent induction of persistent vaginal candidosis in naive mice. Ostrogen-abhangige Induktion der persistierenden Vaginalcandidose in naiven Mausen. Mycoses 2004, 47, 304–309. [Google Scholar] [CrossRef]
- Muñoz, J.E.; Rossi, D.C.P.; Ishida, K.; Spadari, C.C.; Melhem, M.S.; Garcia, D.M.; Caires, A.C.F.; Taborda, C.P.; Rodrigues, E.G. Antifungal activity of the biphosphinic cyclopalladate C7a against Candida albicans yeast forms in vitro and in vivo. Front. Microbiol. 2017, 8, 771. [Google Scholar] [CrossRef]
- Polonelli, L.; Ciociola, T.; Elviri, L.; Zanello, P.P.; Giovati, L.; Arruda, D.C.; Muñoz, J.E.; Mortara, R.A.; Morace, G.; Borghi, E.; et al. A Naturally Occurring Antibody Fragment Neutralizes Infectivity of Diverse Infectious Agents. Sci. Rep. 2016, 6, 35018. [Google Scholar] [CrossRef]
- Ravichandran, C.; Badgujar, P.C.; Gundev, P.; Upadhyay, A. Review of toxicological assessment of d-limonene, a food and cosmetics additive. Food Chem. Toxicol. 2018, 120, 668–680. [Google Scholar] [CrossRef]
- Crowell, P.L.; Elson, C.E.; Bailey, H.H.; Elegbede, A.; Haag, J.D.; Gould, M.N. Human metabolism of the experimental cancer therapeutic agent d-limonene. Cancer Chemother. Pharmacol. 1994, 35, 31–37. [Google Scholar] [CrossRef]
- Freire, J.C.P.; de Oliveira Júnior, J.K.; de Figueredo Silva, D.; de Sousa, J.P.; Guerra, F.Q.S.; de Oliveira Lima, E. Antifungal Activity of Essential Oils against Candida albicans Strains Isolated from Users of Dental Prostheses. Evid.-Based Complement. Altern. Med. 2017, 2017, 1–9. [Google Scholar] [CrossRef]
- Leite, M.C.A.; De Brito Bezerra, A.P.; De Sousa, J.P.; Guerra, F.Q.S.; De Oliveira Lima, E. Evaluation of antifungal activity and mechanism of action of citral against Candida albicans. Evid.-Based Complement. Altern. Med. 2014. [Google Scholar] [CrossRef]
- Thakre, A.; Zore, G.; Kodgire, S.; Kazi, R.; Mulange, S.; Patil, R.; Shelar, A.; Santhakumari, B.; Kulkarni, M.; Kharat, K.; et al. Limonene inhibits Candida albicans growth by inducing apoptosis. Med. Mycol. 2018, 56, 565–578. [Google Scholar]
- Mondello, F.; De Bernardis, F.; Girolamo, A.; Salvatore, G.; Cassone, A. In vitro and in vivo activity of tea tree oil against azole-susceptible and -resistant human pathogenic yeasts. J. Antimicrob. Chemother. 2003, 51, 1223–1229. [Google Scholar] [CrossRef]
- Yano, J.; Sobel, J.D.; Nyirjesy, P.; Sobel, R.; Williams, V.L.; Yu, Q.; Noverr, M.C.; Fidel, P.L. Current patient perspectives of vulvovaginal candidiasis: Incidence, symptoms, management and post-treatment outcomes. BMC Womens. Health 2019, 19, 1–9. [Google Scholar] [CrossRef]
- Cassone, A. Vulvovaginal Candida albicans infections: Pathogenesis, immunity and vaccine prospects. BJOG Int. J. Obstet. Gynaecol. 2015, 122, 785–794. [Google Scholar] [CrossRef]
- Babula, O.; Lazdane, G.; Kroica, J.; Ledger, W.J.; Witkin, S.S. Relation between Recurrent Vulvovaginal Candidiasis, Vaginal Concentrations of Mannose-Binding Lectin, and a Mannose-Binding Lectin Gene Polymorphism in Latvian Women. Clin. Infect. Dis. 2003, 37, 733–737. [Google Scholar] [CrossRef]
- Bae, Y.S.; Rhee, M.S. Short-Term Antifungal Treatments of Caprylic Acid with Carvacrol or Thymol Induce Synergistic 6-Log Reduction of Pathogenic Candida albicans by Cell Membrane Disruption and Efflux Pump Inhibition. Cell. Physiol. Biochem. 2019, 53, 285–300. [Google Scholar]
- Sun, J. D-limonene: Safety and clinical applications. Altern. Med. Rev. 2007, 12, 259–264. [Google Scholar]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muñoz, J.E.; Rossi, D.C.P.; Jabes, D.L.; Barbosa, D.A.; Cunha, F.F.M.; Nunes, L.R.; Arruda, D.C.; Pelleschi Taborda, C. In Vitro and In Vivo Inhibitory Activity of Limonene against Different Isolates of Candida spp. J. Fungi 2020, 6, 183. https://doi.org/10.3390/jof6030183
Muñoz JE, Rossi DCP, Jabes DL, Barbosa DA, Cunha FFM, Nunes LR, Arruda DC, Pelleschi Taborda C. In Vitro and In Vivo Inhibitory Activity of Limonene against Different Isolates of Candida spp. Journal of Fungi. 2020; 6(3):183. https://doi.org/10.3390/jof6030183
Chicago/Turabian StyleMuñoz, Julián E., Diego C. P. Rossi, Daniela L. Jabes, David Aciole Barbosa, Fernanda F. M. Cunha, Luiz R. Nunes, Denise C. Arruda, and Carlos Pelleschi Taborda. 2020. "In Vitro and In Vivo Inhibitory Activity of Limonene against Different Isolates of Candida spp." Journal of Fungi 6, no. 3: 183. https://doi.org/10.3390/jof6030183
APA StyleMuñoz, J. E., Rossi, D. C. P., Jabes, D. L., Barbosa, D. A., Cunha, F. F. M., Nunes, L. R., Arruda, D. C., & Pelleschi Taborda, C. (2020). In Vitro and In Vivo Inhibitory Activity of Limonene against Different Isolates of Candida spp. Journal of Fungi, 6(3), 183. https://doi.org/10.3390/jof6030183