Abstract
Flammulina velutipes is a widely cultivated edible mushroom in East Asia, recognized for its nutritional benefits and distinct morphology characterized by a long stipe and a compact, hemispherical pileus. The pileus not only plays a critical biological role in reproduction through spore formation but also serves as a key commercial trait influencing consumer preference and market value. Despite its economic importance, pileus development in F. velutipes is highly sensitive to environmental factors, among which carbon dioxide (CO2) concentration is particularly influential under indoor cultivation conditions. While previous studies have reported that elevated CO2 levels can inhibit pileus expansion in other mushroom species, the molecular mechanisms by which CO2 affects pileus growth in F. velutipes remain poorly understood. In this study, we investigated the impact of CO2 concentration on pileus morphology and gene expression in F. velutipes by cultivating fruiting bodies under two controlled atmospheric conditions: low (1000 ppm) and high (10,000 ppm) CO2. Morphometric analysis revealed that elevated CO2 levels significantly suppressed pileus expansion, reducing the average diameter by more than 50% compared to the low CO2 condition. To elucidate the underlying genetic response, we conducted RNA sequencing and identified 102 differentially expressed genes (DEGs), with 78 being downregulated under elevated CO2. Functional enrichment analysis highlighted the involvement of cyclin-dependent protein kinase regulatory pathways in this response. Two cyclin genes were found to be significantly downregulated under elevated CO2 conditions, and their suppression was validated through quantitative real-time PCR. These genes, possessing conserved cyclin_N domains, are implicated in the regulation of the eukaryotic cell cycle, particularly in mitotic growth. These results indicate that CO2-induced downregulation of cyclin genes may underlie cell cycle arrest, contributing to inhibited pileus development. This study is the first to provide transcriptomic evidence that elevated CO2 concentrations specifically repress PHO80-like cyclin genes in F. velutipes, revealing a molecular mechanism by which CO2 stress inhibits pileus development. These findings suggest that elevated CO2 triggers a morphogenetic checkpoint by repressing PHO80-like cyclins, thereby modulating cell cycle progression during fruiting body development. This study provides the first evidence of such a transcriptional response in edible mushrooms and offers promising molecular targets for breeding CO2-resilient strains and optimizing commercial cultivation conditions.
1. Introduction
Flammulina velutipes, commonly known as the winter mushroom or enokitake, is a commercially important edible basidiomycete belonging to the family Physalacriaceae. It is extensively cultivated in East Asian countries, particularly in Republic of Korea, Japan, and China, due to its high nutritional value, subtle flavor, and desirable texture. The species is characterized by a long, slender stipe and a small, dome-shaped pileus, which is a reproductive structure responsible for spore production and dispersal [1,2,3]. Among the morphological traits, the size and shape of the pileus are considered critical determinants of both reproductive fitness and commercial value [1,2,3].
The global demand for F. velutipes has grown steadily over the past few decades due to increasing consumer interest in functional foods and natural health products. Rich in dietary fiber, antioxidants, and immunomodulatory compounds such as polysaccharides and lectins, F. velutipes has garnered attention not only in gastronomy but also in nutraceutical and pharmaceutical industries [4,5]. Consequently, optimizing cultivation practices to improve yield and morphological uniformity has become a focal point in mushroom biotechnology.
Among the numerous environmental variables influencing mushroom cultivation—such as temperature, humidity, photoperiod, and substrate composition—gas composition, particularly the concentration of CO2, has emerged as a key factor that modulates the morphogenesis of fungal fruiting bodies [6,7,8,9,10,11]. In intensive cultivation systems, especially those employing sealed or semi-sealed containers, CO2 accumulation is common and can significantly affect the developmental trajectory of mushrooms. Elevated CO2 levels have been reported to suppress pileus expansion and alter the differentiation of various mushroom species, leading to reduced aesthetic quality and lower marketability [6,7,8,9,10,11].
Although the inhibitory effects of elevated CO2 on pileus development have been previously observed in species such as Pleurotus ostreatus and F. filiformis, the underlying molecular mechanisms remain incompletely understood [7,11]. In these species, transcriptomic and proteomic studies suggest that elevated CO2 may interfere with genes and proteins involved in the cell cycle, cell wall biosynthesis, and energy metabolism [7,11]. However, despite the agricultural and economic importance of F. velutipes, detailed investigations into its transcriptional response to CO2 stress have been limited. No studies to date have conclusively identified the key regulatory genes mediating pileus suppression under elevated CO2 conditions in F. velutipes.
Given the growing need for precision agriculture in fungal biotechnology, there is a pressing demand to understand how environmental cues like CO2 concentration affect gene expression and developmental pathways in commercially valuable mushrooms. Such knowledge could facilitate the development of CO2-resilient strains through targeted breeding or genetic engineering, thereby enhancing productivity and quality under suboptimal cultivation conditions.
In this study, we investigated the effects of CO2 concentration on the morphological development of F. velutipes, with a particular focus on pileus size. We cultivated F. velutipes under two controlled CO2 conditions—1000 ppm (ambient) and 10,000 ppm (elevated)—and performed a comparative transcriptomic analysis to identify differentially expressed genes (DEGs). Functional enrichment analyses revealed that two cyclin genes, known for their pivotal role in cell cycle regulation, were significantly downregulated under elevated CO2 conditions. Quantitative PCR confirmed the downregulation of cyclin genes under elevated CO2, consistent with RNA-Seq results. Accordingly, this study was designed to explore the transcriptomic mechanisms of CO2-induced morphological changes in F. velutipes, aiming to identify key genes involved in developmental regulation and provide a foundation for future strain improvement efforts.
2. Materials and Methods
2.1. Fungal Strain and Cultivation Conditions
The F. velutipes strain CBMFV-72 was obtained from the Agricultural Research and Extension Service, Cheongju-si, Republic of Korea. This strain was selected based on its stable morphological traits and widespread use in commercial cultivation studies. The mycelia were initially cultured on potato dextrose agar (PDA; Difco, Seoul, Republic of Korea) at 25 °C for 20 days to promote vegetative growth. Following the pre-culture stage, actively growing mycelial plugs were transferred to a solid fruiting substrate composed of 80% sawdust and 20% rice bran, which was sterilized and incubated under controlled environmental conditions. The incubation for mycelial colonization was carried out at 18 °C with 62–65% relative humidity for 25 days in darkness. Subsequently, to induce fruiting body development, the temperature was sequentially reduced to 14–16 °C under 80% humidity and exposed to light (approximately 300 lux) for 8 h per day.
Two different CO2 concentrations were maintained during the fruiting phase using an automated gas regulation system incorporating a non-dispersive infrared (NDIR) sensor (SH-VT260; SOHA Tech, Seoul, Republic of Korea): a low CO2 condition (1000 ppm, simulating ambient atmospheric levels) and an elevated CO2 condition (10,000 ppm), which reflects accumulation levels frequently encountered in enclosed cultivation facilities. Gas levels were continuously monitored using infrared CO2 sensors to ensure stability. Fruiting bodies were collected at the stage of morphological maturity, characterized by full elongation of the stipe and initial pileus expansion.
2.2. Morphological Analysis
To assess the impact of CO2 concentration on pileus development, pileus diameter was measured from at least 30 fruiting bodies per treatment group using digital calipers. Data were statistically analyzed using Student’s t-test, with a significance threshold of p < 0.001.
2.3. RNA Extraction and Sequencing
Total RNA was extracted from freshly harvested pileus tissues, which were immediately cryopreserved in liquid nitrogen and ground to a fine powder using a pre-cooled mortar and pestle. RNA was isolated using TRIzol reagent (Thermo Fisher Scientific, Seoul, Republic of Korea) following the manufacturer’s instructions, and further purified using the RNeasy Mini Kit (Qiagen, Seoul, Republic of Korea) with on-column DNase I treatment to remove genomic DNA contamination. RNA quality and concentration were assessed using a NanoDrop spectrophotometer (Thermo Fisher Scientific, Seoul, Republic of Korea) and agarose gel electrophoresis. High-quality RNA samples (RIN > 7.0) were selected for sequencing.
We constructed RNA-Seq libraries from 1 μg of total RNA and sequenced them using the Illumina HiSeq 2000 platform (Illumina Korea, Seoul, Republic of Korea), generating paired-end reads. Raw reads were trimmed for quality using a Phred score cutoff of 30 and processed using the Trinity de novo assembly pipeline (v2.15.0) [12]. TransDecoder (v5.5.0) [13] was used to predict coding regions, and CD-HIT (v4.8.1) [14] was employed to reduce redundancy through clustering and deduplication of transcripts (Tables S1 and S2).
2.4. Differential Gene Expression and Enrichment Analysis
Differentially expressed genes (DEGs) between the low and elevated CO2 groups were identified using DESeq2 (v1.48.1) integrated within the Trinity pipeline [15]. The thresholds for significance were set at |log2 fold change| > 1 and p < 0.001. Functional annotation and enrichment analysis of DEGs were performed using the STRING database (v12.0) [16] and the topGO R package (v2.54.0) [17]. GO terms were assigned across three categories: biological process, molecular function, and cellular component. Genes were annotated with domain information using Pfam (24 February 2024) [18] and NCBI Conserved Domain Database (CDD) searches [19].
2.5. Quantitative Real-Time PCR Validation
To validate the RNA-Seq results, quantitative real-time PCR (qRT-PCR) was performed on a subset of differentially expressed genes identified under elevated CO2 conditions. Total RNA was extracted separately from (i) whole fruiting bodies and (ii) dissected pileus and stipe tissues to assess possible tissue-specific expression patterns. In mushrooms, the pileus is the cap-like upper structure that expands and bears the spore-producing hymenium, whereas the stipe is the stem-like supporting structure that elevates the pileus. First-strand cDNA synthesis was carried out using 1 μg of total RNA, oligo(dT) primers, 5× RT buffer, and M-MLV Reverse Transcriptase (Bioneer, Daejeon, Republic of Korea). The reaction mixture was incubated at 25 °C for 5 min, followed by 42 °C for 60 min.
qPCR was conducted using the Bioline SensiFAST SYBR No-ROX kit (Joagene Bioscience, Seoul, Republic of Korea) on a Rotor-Gene 6000 instrument (Qiagen). Each reaction contained 100 ng cDNA, 10 pM gene-specific primers, and SYBR master mix in a final volume of 20 µL. The primer sequences used for amplification are listed in Table S3. Gene expression levels were normalized to an internal reference gene, and relative expression was calculated using the ΔΔCt method. qRT-PCR was performed with three biological replicates, each with three technical replicates. Statistical analysis was performed using t-tests to evaluate significant differences between CO2 treatments.
3. Results
3.1. Elevated CO2 Concentration Significantly Reduces Pileus Size in F. velutipes
The morphological assessment of F. velutipes cultivated under two CO2 concentrations—1000 ppm (low) and 10,000 ppm (high)—revealed a pronounced effect of elevated CO2 on pileus development. As shown in Figure 1a, fruiting bodies grown under elevated CO2 conditions displayed a visibly smaller and less expanded pileus compared to those grown under low CO2. However, the stipe length did not show a statistically significant difference between CO2 treatments. Quantitative measurements confirmed that the mean pileus diameter under elevated CO2 was 4.82 ± 1.11 mm, significantly smaller than the 10.23 ± 2.55 mm observed under low CO2 conditions (p < 0.001, t-test) (Figure 1b). These results are consistent with previous studies in F. filiformis and Pleurotus ostreatus, which reported inhibition of pileus expansion under elevated CO2 environments due to altered cellular proliferation and expansion mechanisms [7,11].

Figure 1.
Effect of CO2 concentration on the pileus of F. velutipes. (a) Pileus of the fruiting bodies of F. velutipes cultivated under CO2 concentrations of 1000 ppm (left) and 10,000 ppm (right). (b) Pileus sizes of F. velutipes under different CO2 concentrations. Data were analyzed using t-tests. *** p < 0.001.
In F. velutipes, this reduction in pileus size further suggests that elevated CO2 may suppress normal fruiting body morphogenesis through changes in physiological or transcriptional regulation. Considering the pileus is the site of spore production and a key trait for commercial evaluation, this CO2-induced morphological alteration presents both biological and industrial concerns for mushroom growers [6,7,11].
3.2. CO2-Induced Transcriptomic Changes in F. velutipes
To explore the molecular mechanisms associated with CO2-induced morphological changes, RNA-Seq analysis was performed, resulting in the detection of 17,038 transcripts across all samples. A comparative transcriptomic analysis was then conducted to identify CO2-responsive gene expression patterns and assess their potential functional relevance (Table S4). Differential gene expression analysis using DESeq2 identified significant transcriptomic alterations, with 20 top-ranked differentially expressed genes (DEGs) selected based on p-value < 0.01 and |log2 fold change| ≥ 1 (Figure 2, Table 1 and Table S5). Among the upregulated genes under elevated CO2, transcripts such as DN3185_c0_g1_i20, DN95_c0_g1_i7, and DN985_c0_g1_i3 exhibited strong induction, with log2 fold changes exceeding seven. These genes are predicted to encode proteins involved in methyltransferase activity, heat shock protein function, and redox metabolism, respectively, suggesting enhanced cellular remodeling and stress adaptation [20,21,22]. Conversely, transcripts including DN1803_c0_g1_i17 and DN4990_c0_g1_i119 were among the most downregulated genes under elevated CO2. These transcripts are associated with major facilitator superfamily transporters and SPX (SYG1, PHO81, and XPR1) domain-containing proteins, potentially linked to nutrient signaling or membrane transport, both of which may be suppressed in carbon-rich environments [23,24]. Notably, genes predicted to encode enzymes such as cytochrome P450s (e.g., DN28_c0_g1_i39, DN5300_c0_g1_i5) were differentially expressed, implicating a role for oxidative metabolism in CO2 response. Several DEGs belonged to protein families involved in structural regulation, such as ankyrin repeats and WD40 domains, which are often associated with cytoskeletal organization and developmental signaling [25,26]. These transcriptional changes mirror phenotypic responses observed in F. velutipes under elevated CO2, including enhanced stipe elongation and delayed cap development. Such morphological alterations are commonly leveraged in commercial cultivation to optimize fruiting body appearance [6]. The upregulation of genes potentially involved in cell wall modification (e.g., chitin biosynthesis, β-glucan remodeling) and stress response indicates a shift toward promoting vegetative growth and suppressing reproductive development [27,28,29]. Furthermore, downregulated genes were enriched in pathways potentially related to aerobic respiration, consistent with metabolic adjustments under limited oxygen availability and excess CO2 [22,30,31,32]. Together, these findings suggest that F. velutipes reprograms its gene expression landscape in response to CO2 levels, modulating energy metabolism, stress tolerance, and developmental timing.
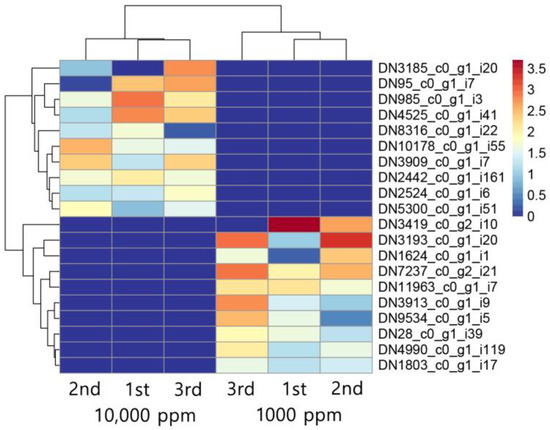
Figure 2.
Top 10 upregulated and top 10 downregulated transcripts in F. velutipes in response to CO2 treatment (1000 ppm and 10,000 ppm). Heatmap shows hierarchical clustering of normalized expression levels across three biological replicates per condition.

Table 1.
Differentially expressed genes in F. velutipes under CO2 treatment (1000 ppm and 10,000 ppm). The table lists the top 10 downregulated and top 10 upregulated genes based on log2 fold change values. Gene expression levels are shown under two CO2 concentrations, and corresponding protein domain annotations are retrieved from InterPro (24 February 2024) and Pfam (24 February 2024) databases. An en dash (“–”) indicates no significant domain hit.
3.3. CO2-Responsive Transcriptomic Profiling of CAZyme Genes
Genome-wide analysis of F. velutipes revealed a diverse array of carbohydrate-active enzymes (CAZymes), including glycoside hydrolases (GHs), auxiliary activities (AAs), carbohydrate esterases (CEs), polysaccharide lyases (PLs), and carbohydrate-binding modules (CBMs), underscoring its capacity for efficient lignocellulosic biomass degradation [1]. CAZyme-related genes were predicted based on annotation using the CAZy database (14 March 2024) (Table S6) [33]. Prominent GH families such as GH5, GH16, and GH43 were identified, which are known to participate in the breakdown of cellulose and hemicellulose, while multiple AA9 genes encoding lytic polysaccharide monooxygenases (LPMOs) suggest a strong oxidative degradation capability that synergizes with hydrolytic enzymes (Table S6) [34]. The presence of CE and PL genes further implies the ability to remove complex side chains and target diverse polysaccharide components, whereas CBM-containing enzymes—particularly those with CBM1 and CBM13—likely enhance substrate binding and catalytic efficiency [34]. Signal peptide predictions in many CAZyme genes suggest their secretion into the extracellular environment, aligning with the saprotrophic lifestyle of F. velutipes [34].
In addition to genomic potential, transcriptomic profiling under varying CO2 concentrations revealed dynamic expression changes in response to environmental carbon levels. Notably, six CAZyme-related genes exhibited significant differential expression between high and low CO2 conditions, indicating a transcriptional regulatory mechanism responsive to atmospheric cues (Figure 3a and Table 2) [35]. These included genes encoding GH114 and GH71 enzymes, which are involved in the degradation of α-glucans and β-glucans, respectively, as well as AA9-type LPMOs that mediate oxidative cleavage of cellulose [33]. Two additional genes encoding CE5 and PL38 family proteins also showed significant expression changes, suggesting CO2 sensitivity not only in catalytic enzymes but also in polysaccharide lyases. Notably, a gene containing both CBM49 and GH13_S domains was also differentially expressed, indicating possible involvement in starch-related substrate recognition. Interestingly, the highly upregulated genes under low CO2 concentrations included those related to hemicellulose and pectin degradation, implying that F. velutipes may prioritize the breakdown of accessible carbohydrates under limited CO2 conditions as a strategy to balance energy demand and metabolic flexibility [35]. This CO2-responsive expression pattern highlights the plasticity of the fungal degradative system and suggests that environmental CO2 not only acts as a substrate-related cue but may also serve as a signaling molecule influencing carbon acquisition strategies. While definitive evidence is still lacking, the observed upregulation of CAZyme genes under low CO2 conditions may be indicative of enhanced structural plasticity and potential expansibility of the fruiting body. This expression pattern could reflect an adaptive developmental strategy whereby F. velutipes modulates its cell wall remodeling capacity in response to environmental signals, thereby optimizing morphogenesis under fluctuating atmospheric conditions.
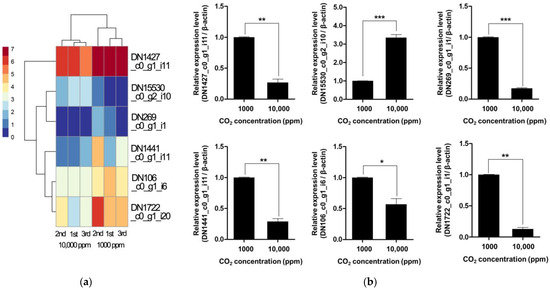
Figure 3.
Heatmap (a) and qRT-PCR validation (b) of differentially expressed CAZyme genes in F. velutipes under low (1000 ppm) and high (10,000 ppm) CO2. Asterisks indicate statistically significant differences between treatments (* p < 0.05, ** p < 0.01, *** p < 0.001; Student’s t-test).
Table 2.
CAZyme-related genes in F. velutipes differentially expressed under CO2 treatment, selected based on log2 fold change thresholds (≥1 or ≤−1).
To validate these transcriptomic findings, we performed qRT-PCR for six representative CAZyme genes, confirming significant expression differences between 1000 ppm and 10,000 ppm CO2 conditions (Figure 3b). These results support the conclusion that CAZyme expression is transcriptionally regulated by atmospheric CO2 levels. These findings have implications for fungal adaptation in changing atmospheric environments and could be leveraged for optimizing biomass degradation in industrial applications under controlled CO2 conditions.
3.4. Transcriptome Profiling Reveals Downregulation of Cell Cycle-Related Genes Under Elevated CO2 Conditions
Out of the 17,038 transcripts detected by RNA-Seq, a total of 102 genes were identified as differentially expressed between high and low CO2 conditions, based on a p-value < 0.01 and |log2 fold change| ≥ 1 (Table S5). Among these, 78 genes were downregulated and 24 were upregulated under elevated CO2 conditions. Gene Ontology (GO) enrichment analysis of the downregulated genes identified significant terms related to cell cycle regulation, particularly cyclin-dependent kinase (CDK) activity and protein kinase binding (Figure 4). Notably, two genes (DN1560_c0_g1_i9 and DN6954_c0_g1_i2) were commonly detected as members of the cyclin family in the enrichment analysis based on the translated protein sequences (Table 3). Pfam domain analysis further confirmed the presence of conserved cyclin_N box domains at the N-termini of these proteins (residues 62–157). In addition, comparative sequence analysis using the NCBI Conserved Domain Database (CDD) analysis revealed that both proteins share high sequence similarity with members of the cyclin_SF superfamily in Saccharomyces cerevisiae, particularly with G1/S-specific cyclins such as PCL1 and PCL2, which are known to interact with the Pho85 cyclin-dependent kinase and regulate the G1/S phase transition in the eukaryotic cell cycle (Figure S1 and Table S5) [36,37]. In contrast, GO enrichment analysis of the upregulated genes did not yield any significantly enriched terms, suggesting that these genes may not be involved in coherent functional pathways or may participate in more diverse or poorly characterized processes.
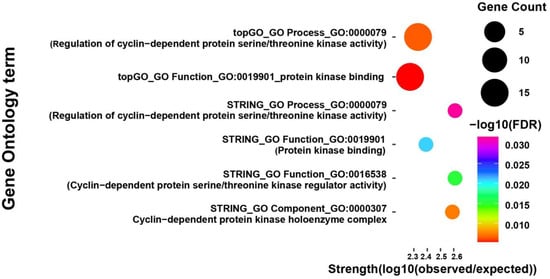
Figure 4.
Gene Ontology (GO) enrichment analysis of downregulated genes in F. velutipes under elevated CO2 conditions (10,000 ppm). GO terms were identified using STRING and topGO analysis tools and are grouped into three categories: biological process, molecular function, and cellular component. Each bubble represents an enriched GO term. The size of the bubble corresponds to the number of genes associated with that term, the color scale indicates statistical significance as −log10 of the false discovery rate (FDR), and the x-axis represents enrichment strength.
Table 3.
List of enriched GO terms among significantly downregulated DEGs in F. velutipes cultivated under 10,000 ppm CO2, with commonly identified genes shown in bold.
The downregulation of cyclin genes under elevated CO2 conditions strongly suggests that elevated CO2 disrupts normal mitotic processes required for cell proliferation during pileus development. Cyclins are essential components of the cell cycle machinery, forming complexes with CDKs to regulate progression through various checkpoints [38,39]. In fungi, especially in basidiomycetes like F. velutipes, cell division and tissue differentiation are tightly coordinated during the formation of reproductive structures [40,41,42]. Suppression of these regulatory genes could impair cellular growth in the pileus region, leading to reduced expansion and final size.
3.5. Evolutionary Conservation of Cyclin Sequences Among Basidiomycota
To support the transcriptomic findings indicating CO2-responsive suppression of cyclin genes, we conducted a comprehensive investigation into the evolutionary conservation and domain architecture of the identified cyclin candidates using PSI-BLAST (26 March 2024) (threshold 0.005) and InterPro protein family classification (e-value < 0.001). Homology searches against fungal protein databases (NCBI non-redundant database) revealed that the majority of top cyclin homologs were derived from members of the phylum Basidiomycota, including the genera Lentinula, Armillaria, Mycena, and Pleurotus (Figure S2), suggesting that cyclin-mediated cell cycle regulation is evolutionarily conserved among mushroom-forming fungi.
Multiple sequence alignment of these homologs revealed strong conservation within the canonical cyclin box, which is thought to mediate interactions with cyclin-dependent kinases (CDKs) [43]. Although detailed structural data are limited for fungal cyclins, residues such as leucine (L), glutamic acid (E), and phenylalanine (F) are commonly involved in hydrophobic interactions in cyclin–CDK complexes in other eukaryotes (Figure 5) [44]. Their conservation across Basidiomycota homologs suggests a potential functional role in CDK association and maintenance of cyclin structure.
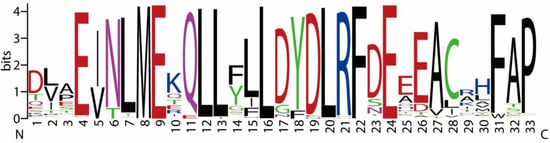
Figure 5.
Conserved amino acid regions identified from PSI-BLAST alignment of F. velutipes PHO80-like cyclin homologs (DN1560_c0_g1_i9 and DN6954_c0_g1_i2) with cyclin sequences from other fungal species.
InterPro classification of the F. velutipes transcriptome identified seven transcripts belonging to the PHO80-like cyclin protein family (IPR013922), which includes a group of eukaryotic cyclins involved in regulating the G1/S phase transition of the cell cycle (Table 4) [45,46,47]. Among these, only two transcripts (DN1560_c0_g1_i9 and DN6954_c0_g1_i2) were significantly downregulated under elevated CO2 conditions and were concurrently enriched in Gene Ontology (GO) terms related to “regulation of CDK activity” and “protein kinase binding”. These two transcripts were the only PHO80-like cyclins to satisfy both differential expression and functional enrichment thresholds, indicating their unique regulatory roles in the CO2-responsive gene network. Moreover, comparative sequence analysis among PHO80-like cyclin genes within the F. velutipes genome revealed that only these two genes shared specific conserved sequence regions (outside the canonical cyclin_N domain), which are also commonly found across other Basidiomycota species (Figures S2 and S3), highlighting their evolutionary conservation and suggesting they act as key regulators of cell cycle control and environmental adaptation in response to CO2. Notably, DN1560_c0_g1_i9 was identified as an isoform from a single gene locus, and only this isoform was differentially expressed under elevated CO2. This suggests that structural differences between isoforms may affect their transcriptional responsiveness, highlighting the regulatory importance of transcript diversity in environmental adaptation.
Table 4.
Predicted cyclin-related genes in F. velutipes identified by transcriptome analysis.
The exclusive identification of DN1560_c0_g1_i9 and DN6954_c0_g1_i2 as core members of the PHO80-like family with environmental responsiveness suggests a high degree of functional specialization. This specificity may reflect regulatory divergence at the transcriptional level, differential promoter sensitivity to environmental stimuli, or spatial–temporal expression patterns within developing tissues. While the PHO80-like family encompasses multiple cyclins with structural similarity, only these two genes appear to be functionally integrated into the CO2-regulated developmental framework of F. velutipes.
Taken together, the convergence of phylogenetic conservation, family-level classification, and transcriptional responsiveness under CO2 stress highlights DN1560_c0_g1_i9 and DN6954_c0_g1_i2 as pivotal regulators of cell cycle progression. These genes represent promising molecular targets for developing CO2-tolerant mushroom strains and provide valuable insights into how environmental factors modulate morphogenetic pathways in basidiomycetous fungi.
3.6. Validation and Tissue-Specific Regulation of CO2-Induced Cyclin Gene Repression
To validate the transcriptomic observation of cyclin gene repression under elevated CO2 (10,000 ppm), we conducted qRT-PCR targeting two PHO80-like cyclin genes, DN6954_c0_g1_i2 and DN1560_c0_g1_i9, using RNA isolated from three distinct tissues of F. velutipes: the pileus, stipe, and whole fruiting body.
As shown in Figure 6, DN1560_c0_g1_i9 was significantly downregulated across all tissue types, with reductions of approximately 72% in the pileus, 64% in the stipe, and 68% in the whole fruiting body under high CO2 conditions (p < 0.01). These findings suggest a broad, CO2-responsive role of this gene in cell cycle regulation during fruiting body development.
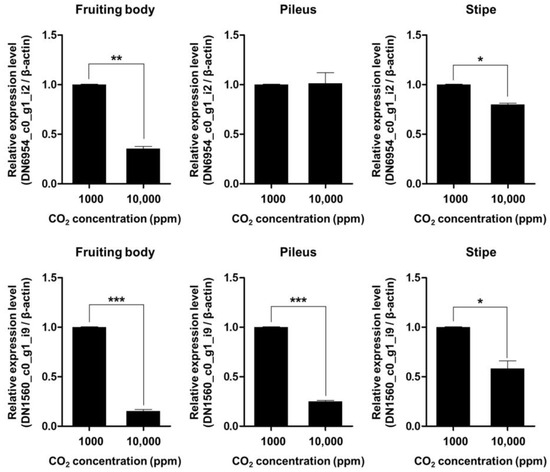
Figure 6.
Quantitative real-time PCR validation of the relative expression levels of two PHO80-like cyclin genes (DN6954_c0_g1_i2 and DN1560_c0_g1_i9) in F. velutipes under different CO2 concentrations (1000 ppm and 10,000 ppm). Expression values were measured in the whole fruiting body, pileus, and stipe tissues. Bars represent mean ± standard error (n = 3 biological replicates). Asterisks indicate statistically significant differences between treatments (* p < 0.05, ** p < 0.01, *** p < 0.001; Student’s t-test).
In contrast, DN6954_c0_g1_i2 exhibited tissue-specific repression. While its expression was significantly decreased in the stipe (~81%) and whole fruiting body (~88%) (p < 0.001), no significant change was observed in the pileus (p > 0.05). This suggests that DN6954_c0_g1_i2 may play a more specialized role in regulating mitotic activity in the stipe rather than in the pileus.
Interestingly, despite the lack of differential expression in the pileus, DN6954_c0_g1_i2 still exhibited strong downregulation at the whole-fruiting-body level. This discrepancy likely reflects the disproportionately large biomass contribution of the stipe relative to the pileus in F. velutipes [1]. Given that the stipe is elongated and dominates the total volume of the fruiting body, expression changes in this tissue can strongly influence overall transcript levels. Moreover, the stipe undergoes vigorous cell division and elongation during development, further supporting the idea that DN6954_c0_g1_i2 functions primarily in stipe-specific cell cycle control [6].
These tissue-dependent patterns of gene expression repression imply that F. velutipes differentially regulates cyclin genes in response to CO2 stress depending on anatomical location. While DN1560_c0_g1_i9 appears to be a global regulator affecting multiple tissues, DN6954_c0_g1_i2 may serve as a stipe-specific CO2-responsive gene involved in controlling longitudinal growth. This is consistent with the observed CO2-induced morphological changes, in which stipe elongation is promoted while pileus expansion is inhibited under elevated CO2 conditions [7].
Overall, our qRT-PCR validation confirms the RNA-Seq results and reveals a spatially resolved regulatory mechanism, where CO2 influences the transcriptional repression of key cyclin genes in a tissue-dependent manner. These findings suggest that DN6954_c0_g1_i2 and DN1560_c0_g1_i9 represent valuable molecular markers for breeding CO2-resilient mushroom strains, and they offer important insights into how atmospheric conditions modulate organ-specific growth processes in basidiomycetes. However, further functional characterization will be necessary to validate the tissue-specific roles of these genes and to elucidate the underlying regulatory mechanisms in response to CO2 stress.
4. Discussion
In general, high concentrations of CO2 inhibit the growth of many aerobic microorganisms, but wood-decaying basidiomycetes such as F. velutipes are known as capnophilic or facultative anaerobes that thrive in low-oxygen, high-carbon environments [22,48]. However, our results suggest that CO2 may have inhibitory effects when it exceeds a certain threshold, especially during sensitive developmental stages such as fruiting body formation [6,36].
Transcriptome analysis showed that high concentrations of CO2 downregulated the expression of key regulators of cell cycle progression, particularly PHO80-like cyclins, suggesting that they act as regulators of pileus size under abnormal atmospheric conditions. This may be an adaptive strategy to conserve energy in environments with limited ventilation such as decaying logs [9,22,49,50,51,52].
Interestingly, under high CO2 conditions, the growth of the pileus was suppressed, but the number of fruiting bodies significantly increased (Figure S4). Analysis of the number of fruiting bodies showed that F. velutipes cultured under 10,000 ppm CO2 produced significantly more fruiting bodies than those under 1000 ppm CO2, suggesting a shift to a compensatory developmental strategy that prioritizes increased reproductive output over pileus expansion. This morphological change may be a fruiting body development strategy for efficient spore dispersal and environmental adaptation in an oxygen-limited environment.
These results suggest that CO2 may act as a signal that affects not only pileus size but also the number of fruiting bodies in F. velutipes. Therefore, the suppression of cyclin expression and the associated morphological changes in F. velutipes may reflect an adaptive strategy to gas changes in the atmosphere. Although this study did not specifically clarify the ecological implications of environmental changes in F. velutipes, it provides an interesting hypothesis that should be examined in future studies.
In terms of application, this study also provides important implications for mushroom cultivation. In particular, in modern bottle or bag cultivation systems, CO2 accumulation due to insufficient ventilation causes pileus size variation and inconsistent product quality [6,9,53]. The CO2-responsive cyclin gene of F. velutipes identified in this study can be utilized as a candidate molecular marker for breeding high CO2-tolerant strains. In addition, it will be possible to improve cultivation conditions that maintain both yield and morphological quality through real-time environmental monitoring and air flow optimization.
In this study, transcriptome analysis of F. veutipes revealed a significant association between cyclin genes and changes in pileus size and number of fruiting bodies. However, further studies involving mutation or overexpression are needed to clarify the functional roles of these cyclin genes, and in parallel, future research should explore how elevated CO2 levels influence quality-related traits such as taste, texture, and nutritional composition, in addition to morphological changes.
5. Conclusions
In this study, we demonstrated that elevated atmospheric CO2 concentration (10,000 ppm) significantly inhibits pileus development in F. velutipes by downregulating two PHO80-like cyclin genes, DN1560_c0_g1_i9 and DN6954_c0_g1_i2, which are implicated in mitotic regulation during fruiting body formation. This repression was validated through RNA-Seq and qRT-PCR across multiple tissue types and shows strong homology to the PCL1/PCL2 family of S. cerevisiae, suggesting conserved regulatory mechanisms across fungi.
Our results represent the first clear indication that repression of specific cyclin genes contributes to reduced pileus expansion under elevated CO2 in a cultivated mushroom species. These observations imply that CO2 may play a dual role—not only limiting development physically but also acting as a molecular signal that activates checkpoint pathways. Altogether, our results shed light on how fungi adjust their developmental processes at the transcriptomic level, and they may offer practical clues for enhancing traits in cultivated strains.
Furthermore, increased CO2 concentration led to a reduction in pileus size, accompanied by a compensatory rise in fruiting body number of F. velutips. This finding raises the possibility of a trade-off between pileus expansion and fruiting body proliferation under changing environmental conditions. This observation indicates the complexity of interpreting growth changes caused by the environment. It also emphasizes the need for an integrated understanding through physiological and molecular analyses in future studies.
Overall, these findings help us understand how environmental changes affect the development of mushroom-forming fungi and provide a foundation for developing CO2-tolerant mushroom cultivation strategies based on transcriptome-level insights.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/jof11080551/s1, Figure S1: Conserved domain analysis of two cyclin genes identified under elevated CO2 conditions in F. velutipes. NCBI Conserved Domain Database (CDD) search results showing the presence of conserved cyclin_N box domains at the N-termini of DN1560_c0_g1_i9 and DN6954_c0_g1_i2. The alignment highlights regions of high sequence similarity to the cyclin_SF superfamily, particularly PHO85-type cyclins from Saccharomyces cerevisiae; Figure S2: Multiple sequence alignment of conserved amino acid regions outside the canonical cyclin_N domain in cyclin homologs identified by PSI-BLAST. Sequences were aligned using PSI-BLAST hits from F. velutipes cyclin genes and representative basidiomycete species; Figure S3: Comparative sequence analysis of PHO80-like cyclin genes within the F. velutipes genome. Boxes indicate the specific conserved sequence regions that are commonly shared with Basidiomycota homologs; Figure S4: Comparison of fruiting body numbers in F. velutipes under 1000 ppm and 10,000 ppm CO2. Representative images of culture bottles grown under 1000 ppm CO2 (left) and 10,000 ppm CO2 (right) (a). Average number of fruiting bodies per bottle under each condition (b). Asterisks indicate statistically significant differences between treatments (p < 0.05; Student’s t-test); Table S1: Predicted amino acid sequences of non-redundant coding transcripts identified from the F. velutipes CBMFV-72 strain; Table S2: Predicted coding DNA sequences (CDSs) of assembled transcripts from the F. velutipes CBMFV-72 strain; Table S3: List of primers used in this study; Table S4: Full list of differentially expressed genes in F. velutipes under CO2 treatment (1000 ppm and 10,000 ppm); Table S5: List of differentially expressed genes identified through RNA-Seq analysis of F. velutipes exposed to high and low CO2 concentrations (p < 0.01, |log2FC| > 1); Table S6: Prediction of carbohydrate-active enzyme (CAZyme) genes in F. velutipes CBMFV-72 using the CAZy database.
Author Contributions
Conceptualization, Y.-J.P. and K.-W.L.; methodology, Y.-J.P.; validation, K.-W.L. and C.-H.P.; formal analysis, C.-H.P. and J.-H.S.; investigation, K.-W.L.; resources, K.-W.L.; data curation, Y.-J.P. and S.-C.L.; writing—original draft preparation, Y.-J.P., K.-W.L. and C.-H.P.; writing—review and editing, Y.-J.P., K.-W.L. and C.-H.P.; visualization, Y.-J.P.; supervision, Y.-J.P.; project administration, Y.-J.P.; funding acquisition, Y.-J.P. All authors have read and agreed to the published version of the manuscript.
Funding
This paper was supported by Konkuk University in 2025.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Raw sequencing reads were deposited in the NCBI Sequence Read Archive (SRA) under the accession numbers SRR28157287–SRR28157292.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Park, Y.J.; Baek, J.H.; Lee, S.; Kim, C.; Rhee, H.; Kim, H.; Seo, J.S.; Park, H.R.; Yoon, D.E.; Nam, J.Y.; et al. Whole genome and global gene expression analyses of the model mushroom Flammulina velutipes reveal a high capacity for lignocellulose degradation. PLoS ONE 2014, 9, e93560. [Google Scholar] [CrossRef]
- Kim, E.S.; Woo, S.I.; Oh, M.J.; Oh, Y.L.; Shin, P.G.; Jang, K.Y.; Kong, W.S.; Lee, C.S. Characteristics of ‘Baekjung’, a variety adaptable to high temperature in Flammulina velutipes. J. Mushroom 2015, 13, 203–206. [Google Scholar] [CrossRef]
- Kong, W.S.; Jang, K.Y.; Lee, C.Y.; Gu, J.S.; Shin, P.G.; Jhune, C.S.; Oh, Y.L.; Yoo, Y.B.; Suh, J.S. Breeding progress and characterization of a Korean white variety ‘Baek-A’ in Flammulina velutipes. J. Mushroom 2013, 11, 159–163. [Google Scholar] [CrossRef]
- Ye, S.; Gao, Y.; Hu, X.; Cai, J.; Sun, S.; Jiang, J. Research progress and future development potential of Flammulina velutipes polysaccharides in the preparation process, structure analysis, biology, and pharmacology: A review. Int. J. Biol. Macromol. 2024, 267, 131467. [Google Scholar] [CrossRef]
- Lu, Y.P.; Chen, R.L.; Long, Y.; Li, X.; Jiang, Y.J.; Xie, B.G. A Jacalin-related lectin regulated the formation of aerial mycelium and fruiting body in Flammulina velutipes. Int. J. Mol. Sci. 2016, 17, 1884. [Google Scholar] [CrossRef]
- Sakamoto, Y. Influences of environmental factors on fruiting body induction, development and maturation in mushroom-forming fungi. Fungal Biol. Rev. 2018, 32, 236–248. [Google Scholar] [CrossRef]
- Yan, J.J.; Tong, Z.J.; Liu, Y.Y.; Li, Y.N.; Zhao, C.; Mukhtar, I.; Tao, Y.X.; Chen, B.Z.; Deng, Y.J.; Xie, B.G. Comparative transcriptomics of Flammulina filiformis suggests a elevated CO2 concentration inhibits early pileus expansion by decreasing cell division control pathways. Int. J. Mol. Sci. 2019, 20, 5923. [Google Scholar] [CrossRef]
- Kinugawa, K.; Tanesaka, E. Changes in the rate of CO2 release from cultures of three basidiomycetes during cultivation. Trans. Mycol. Soc. Jpn. 1990, 31, 489–500. [Google Scholar]
- Kinugawa, K.; Suzuki, A.; Takamatsu, Y.; Kato, M.; Tanaka, K. Effects of concentrated carbon dioxide on the fruiting of several cultivated basidiomycetes (II). Mycoscience 1994, 35, 345–352. [Google Scholar] [CrossRef]
- Jang, K.Y.; Jhune, C.S.; Park, J.S.; Cho, S.M.; Weon, H.Y.; Cheong, J.C.; Choi, S.G.; Sung, J.M. Characterization of fruitbody morphology on various environmental conditions in Pleurotus ostreatus. Mycobiology 2003, 31, 145–150. [Google Scholar] [CrossRef]
- Lin, R.; Zhang, L.; Yang, X.; Li, Q.; Zhang, C.; Guo, L.; Yu, H.; Yu, H. Responses of the mushroom Pleurotus ostreatus under different CO2 concentration by comparative proteomic analyses. J. Fungi 2022, 8, 652. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. A Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef]
- Haas, B.J.; Papanicolaou, A.; Yassour, M.; Grabherr, M.; Blood, P.D.; Bowden, J.; Couger, M.B.; Eccles, D.; Li, B.; Lieber, M.; et al. De novo transcript sequence reconstruction from RNA-seq using the Trinity platform for reference generation and analysis. Nat. Protoc. 2013, 8, 1494–1512. [Google Scholar] [CrossRef]
- Fu, L.; Niu, B.; Zhu, Z.; Wu, S.; Li, W. CD-HIT: Accelerated for clustering the next-generation sequencing data. Bioinformatics 2012, 28, 3150–3152. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, e550. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Nastou, K.C.; Lyon, D.; Kirsch, R.; Pyysalo, S.; Doncheva, N.T.; Legeay, M.; Fang, T.; Bork, P.; et al. The STRING database in 2021: Customizable protein-protein networks, and functional characterization of user-uploaded gene/measurement sets. Nucleic Acids Res. 2021, 49, D605–D612. [Google Scholar] [CrossRef]
- Alexa, A.; Rahnenfuhrer, J. topGO: Enrichment Analysis for Gene Ontology. 2024. Available online: https://bioconductor.org/packages/release/bioc/html/topGO.html (accessed on 3 May 2025).
- Paysan-Lafosse, T.; Andreeva, A.; Blum, M.; Chuguransky, S.R.; Grego, T.; Pinto, B.L.; Salazar, G.A.; Bileschi, M.L.; Llinares-López, F.; Meng-Papaxanthos, L.; et al. The Pfam protein families database: Embracing AI/ML. Nucleic Acids Res. 2025, 53, D523–D534. [Google Scholar] [CrossRef]
- Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Gonzales, N.R.; Gwadz, M.; Lu, S.; Marchler, G.H.; Song, J.S.; Thanki, N.; Yamashita, R.A.; et al. The conserved domain database in 2023. Nucleic Acids Res. 2023, 51, D384–D388. [Google Scholar] [CrossRef]
- Huang, R.; Ding, Q.; Xiang, Y.; Gu, T.; Li, Y.; Li, Y. Comparative analysis of DNA methyltransferase gene family in fungi: A focus on Basidiomycota. Front. Plant Sci. 2016, 7, 1556. [Google Scholar] [CrossRef]
- Kim, M.S.; Ko, Y.J.; Maeng, S.; Floyd, A.; Heitman, J.; Bahn, Y.S. Comparative transcriptome analysis of the CO2 sensing pathway via differential expression of carbonic anhydrase in Cryptococcus neoformans. Genetics 2010, 185, 1207–1219. [Google Scholar] [CrossRef]
- Chadwick, B.J.; Lin, X. Effects of CO2 in fungi. Curr. Opin. Microbiol. 2024, 79, 102488. [Google Scholar] [CrossRef]
- Perlin, M.H.; Andrews, J.M.; Toh, S.S. Essential letters in the fungal alphabet: ABC and MFS transporters and their roles in survival and pathogenicity. Adv. Genet. 2014, 85, 201–253. [Google Scholar] [CrossRef]
- Chabert, V.; Kim, G.D.; Qiu, D.; Liu, G.; Michaillat Mayer, L.; Jamsheer, K.M.; Jessen, H.J.; Mayer, A. Inositol pyrophosphate dynamics reveals control of the yeast phosphate starvation program through 1,5-IP8 and the SPX domain of Pho81. eLife 2023, 12, RP87956. [Google Scholar] [CrossRef] [PubMed]
- Mosavi, L.K.; Cammett, T.J.; Desrosiers, D.C.; Peng, Z.Y. The ankyrin repeat as molecular architecture for protein recognition. Protein Sci. 2004, 13, 1435–1448. [Google Scholar] [CrossRef] [PubMed]
- Jain, B.P.; Pandey, S. WD40 repeat proteins: Signaling scaffold with diverse functions. Protein J. 2018, 37, 391–406. [Google Scholar] [CrossRef]
- Kappel, L.; Münsterkötter, M.; Sipos, G.; Escobar Rodriguez, C.; Gruber, S. Chitin and chitosan remodeling defines vegetative development and Trichoderma biocontrol. PLoS Pathog. 2020, 16, e1008320. [Google Scholar] [CrossRef]
- Gow, N.A.R.; Latge, J.P.; Munro, C.A. The fungal cell wall: Structure, biosynthesis, and function. Microbiol. Spectr. 2017, 5, 28513415. [Google Scholar] [CrossRef]
- Robledo-Briones, M.; Ruiz-Herrera, J. Regulation of genes involved in cell wall synthesis and structure during Ustilago maydis dimorphism. FEMS Yeast Res. 2013, 13, 74–84. [Google Scholar] [CrossRef]
- Masuo, S.; Terabayashi, Y.; Shimizu, M.; Fujii, T.; Kitazume, T.; Takaya, N. Global gene expression analysis of Aspergillus nidulans reveals metabolic shift and transcription suppression under hypoxia. Mol. Genet. Genom. 2010, 284, 415–424. [Google Scholar] [CrossRef]
- Chung, H.; Lee, Y.H. Hypoxia: A double-edged sword during fungal pathogenesis? Front. Microbiol. 2020, 11, 1920. [Google Scholar] [CrossRef]
- Hillmann, F.; Shekhova, E.; Shekhova, E.; Kniemeyer, O.; Kniemeyer, O. Insights into the cellular responses to hypoxia in filamentous fungi. Curr. Genet. 2015, 61, 441–455. [Google Scholar] [CrossRef]
- Zheng, J.; Ge, Q.; Yan, Y.; Zhang, X.; Huang, L.; Yin, Y. dbCAN3: Automated carbohydrate-active enzyme and substrate annotation. Nucleic Acids Res. 2023, 51, W115–W121. [Google Scholar] [CrossRef]
- Yu, H.W.; Im, J.H.; Kong, W.S.; Park, Y.J. Comparative analysis of carbohydrate active enzymes in the Flammulina velutipes var. lupinicola Genome. Microorganisms 2020, 9, 20. [Google Scholar] [CrossRef]
- Mattila, H.K.; Mäkinen, M.; Mäkinen, M.; Lundell, T. Hypoxia is regulating enzymatic wood decomposition and intracellular carbohydrate metabolism in filamentous white rot fungus. Biotechnol. Biofuels 2020, 13, 26. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Friesen, H.; Andrews, B.J. Pho85, a multifunctional cyclin-dependent protein kinase in budding yeast. Mol. Microbiol. 2007, 66, 303–314. [Google Scholar] [CrossRef] [PubMed]
- Hadwiger, J.A.; Wittenberg, C.; Richardson, H.E.; de Barros Lopes, M.; Reed, S.I. A family of cyclin homologs that control the G1 phase in yeast. Proc. Natl. Acad. Sci. USA. 1989, 86, 6255–6259. [Google Scholar] [CrossRef]
- Lents, N.H.; Piszczatowski, R.T. Cyclins, Cyclin-Dependent Kinases, and Cyclin-Dependent Kinase Inhibitors. In Encyclopedia of Cell Biology, 2nd ed.; Bradshaw, R.A., Hart, G.W., Stahl, P.D., Eds.; Academic Press: Oxford, UK, 2023; Volume 5, pp. 224–234. [Google Scholar] [CrossRef]
- Barnum, K.J.; O’Connell, M.J. Cell cycle regulation by checkpoints. Methods Mol. Biol. 2014, 1170, 29–40. [Google Scholar] [CrossRef]
- Pelkmans, J.F.; Lugones, L.G.; Wösten, H.A.B. Fruiting Body Formation in Basidiomycetes. In The mycota, 3rd ed.; Wendland, J., Ed.; Springer: Cham, Switzerland, 2016; Volume 1, pp. 387–405. [Google Scholar] [CrossRef]
- Coelho, M.A.; Bakkeren, G.; Sun, S.; Hood, M.E.; Giraud, T. Fungal sex: The Basidiomycota. Microbiol. Spectr. 2017, 5, 147–175. [Google Scholar] [CrossRef]
- Nagy, L. Evolutionary Morphogenesis of Sexual Fruiting Bodies in Basidiomycota: Toward a New Evo-Devo Synthesis. Microbiol. Mol. Biol. Rev. 2022, 86, e0001921. [Google Scholar] [CrossRef]
- Malumbres, M.; Harlow, E.; Hunt, T.; Hunter, T.; Lahti, J.M.; Manning, G.; Morgan, D.O.; Tsai, L.H.; Wolgemuth, D.J. Cyclin-dependent kinases: A family portrait. Nat. Cell Biol. 2009, 11, 1275–1276. [Google Scholar] [CrossRef]
- Heitz, F.; Morris, M.C.; Fesquet, D.; Cavadore, J.C.; Dorée, M.; Divita, G. Interactions of cyclins with cyclin-dependent kinases: A common interactive mechanism. Biochemistry 1997, 36, 4995–5003. [Google Scholar] [CrossRef]
- Neganova, I.; Lako, M. G1 to S phase cell cycle transition in somatic and embryonic stem cells. J. Anat. 2008, 213, 30–44. [Google Scholar] [CrossRef]
- Turnaev, I.I.; Gunbin, K.V.; Kolchanov, N.A. The evolution of key cell cycle proteins correlates with an increase in the complexity of eukaryotic organisms. Dokl. Biochem. Biophys. 2009, 426, 147–151. [Google Scholar] [CrossRef] [PubMed]
- Measday, V.; Moore, L.; Retnakaran, R.; Lee, J.; Donoviel, M.; Neiman, A.M.; Andrews, B. A family of cyclin-like proteins that interact with the Pho85 cyclin-dependent kinase. Mol. Cell. Biol. 1997, 17, 1212–1223. [Google Scholar] [CrossRef] [PubMed]
- Mukhin, V.A.; Diyarova, D.K. Eco-Physiological Adaptations of the Xylotrophic Basidiomycetes Fungi to CO2 and O2 Mode in the Woody Habitat. J. Fungi 2022, 8, 1296. [Google Scholar] [CrossRef] [PubMed]
- Camilo, C.M.; Gomes, S.L. Transcriptional response to hypoxia in the aquatic fungus Blastocladiella emersonii. Eukaryot. Cell 2010, 9, 915–925. [Google Scholar] [CrossRef]
- Bonaccorsi, E.D.; Ferreira, A.J.; Chambergo, F.S.; Ramos, A.S.; Mantovani, M.C.; Farah, J.P.; Sorio, C.S.; Gombert, A.K.; Tonso, A.; El-Dorry, H. Transcriptional response of the obligatory aerobe Trichoderma reesei to hypoxia and transient anoxia: Implications for energy production and survival in the absence of oxygen. Biochemistry 2006, 45, 3912–3924. [Google Scholar] [CrossRef]
- Chun, C.D.; Liu, O.W.; Madhani, H.D. A link between virulence and homeostatic responses to hypoxia during infection by the human fungal pathogen Cryptococcus neoformans. PLoS Pathog. 2007, 3, e22. [Google Scholar] [CrossRef]
- Maček, I. Fungi in Hypoxic Soils and Aquatic Sediments. In Extremophilic Fungi, 1st ed.; Sahay, S., Ed.; Springer: Singapore, 2022; pp. 219–243. [Google Scholar] [CrossRef]
- Chi, J.H.; Kim, J.H.; Ju, Y.; Seo, G.S.; Kang, H.W. Effects of Elevated Carbon Dioxide on the Fruiting Initiation and Development of Grifola frondosa. Korean J. Mycol. 2009, 37, 60–64. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).