

Food Safety Aspects of Breeding Maize to Multi-Resistance against the Major (Fusarium graminearum, F. verticillioides, Aspergillus flavus) and Minor Toxigenic Fungi (Fusarium spp.) as Well as to Toxin Accumulation, Trends, and Solutions—A Review

Abstract
1. Introduction
2. Ear Rot, Mycotoxins, and Losses in Maize Production
3. Ear Rot-Causing Agents and Toxin Relations, General View
4. Which Mycotoxins Are Important?
- The first is that the mycotoxins produced by the toxigenic fungi that occur most frequently in a given region should be listed first. Examples include DON and ZEN from F. graminearum, FB1 from F. verticillioides, and aflatoxin B1 from Aspergillus flavus. These mycotoxins have global significance [3]. To these, we should add the different masked variants, isomers, and so on. Then, locally important toxigenic fungi should be considered; however, breeding against these is normally not carried out, and resistance to the most important species remains unknown, so they are problematic and uncontrolled. It is not an accident that the brochures for hybrids normally do not address this problem, with the yielding ability being the most decisive element. As an example, members of the Gibberella fujikuroi clade, such as F. verticillioides, are known to produce MON (without exception) and BEA (rather rarely), causing additional food and feed safety problems [89].
- The second source of knowledge is multi-toxin analyses of infected food and feed samples. There are two modes of analysis: targeted and non-targeted. To obtain a full picture, a non-targeted analysis should be conducted, through which even 50–100 toxigenic compounds can be detected—mostly for research purposes—with the objective of determining those that should be the subject of targeted analyses. The authors of [89,90,91] identified 12 mycotoxins in maize and its products, with Fusarium toxins found mostly in whole maize and maize gluten samples. Aflatoxins, ochratoxin A, T-2 toxin, and HT-2 were concentrated in maize gluten feed. Meanwhile, the starch part was much less contaminated. Guan et al. [2] checked 11 mycotoxins, and, beyond the regular AFs, DON, FBs, and ZEN, OTA was also identified. Eckhard et al. [92] observed DON in every silo maize sample, but (in decreasing order) nivalenol, T-2, HT-2, acetylated DON, and FBs were also detected. From these studies, we can conclude that, beyond the standard four toxins, we should also consider nivalenol, T-2, HT-2, and (maybe) OTA; the necessary list may be much longer. Non-regulated mycotoxins represent an important analysis object, and, in one study, 38 different combinations were detected, with combinations in one sample containing up to 12 mycotoxins [93]. The extent to which this is a breeding problem remains unknown, but the problem is considerable. Among these mycotoxins, nivalenol (NIV), enniatin B (ENB), and enniatin B1 (ENB1) were the most frequent. As nivalenol can be produced by strains of F. graminearum sensu stricto and some F. culmorum isolates—as well as by F. asiaticum and other members of the F. graminearum clade—we should pay attention to such species.
- The third source of information is urine and blood mycotoxin analyses in humans and animals. Lemming et al. [94] tested 3000 school students for urine and blood sera. In urine, DON, DON-15-β-D-O-glucuronide (DON-15GlcA, 9.1%), dihydro-citrinone (DH-CIT, 0.5%), HT-2-glucuronide (HT-2-3-GlcA, 0.1%), and ochratoxin A (OTA, 0.1%) were identified, while OTA was found in all sera samples. Furthermore, 2′R-OTA occurred in 8.3%, and enniatin B in 99.2% of all samples. The cereal background was found for DON, ENB, and OTA. Izzo et al. [95] compared targeted (for six toxins) and non-targeted analyses on the same samples. They concluded that the targeted test was better when the toxin background had been determined prior. Abia et al. [96] identified aflatoxin M1, fumonisin B1, ochratoxin A, and DON from urine. De Santis et al. [97] screened eight mycotoxins from urine and found DON, ZEN, FBs, T2/HT2, and NIV to be at critical levels. Schmidt et al. [97] reported the occurrence of aflatoxin M1 (AFM1), altenuene (ALT), alternariol monomethyl ether (AME), alternariol (AOH), citrinin (CIT) and its metabolite dihydrocitrinone (DH-CIT), fumonisin B1 (FB1), and ochratoxin A (OTA). Additionally, ZEN, as well as α- and β-zearalenol, FB1, and OTA, was found in 100% and 38% of human urine samples, while AFM1, ZEN, citrinin, and DH-CIT were observed in 10–20% of samples. Novel Spanish data [98] reported on the presence of all aflatoxins, fumonisin B1 + B2, ochratoxin A, DON (with 3 and 15 ADON), and T-2-HT2. The overall list is longer, but at least 4–6 mycotoxins appear to occur at levels higher than the limits and, so, can be considered responsible (alone or in combination) for toxicoses in humans and animals.
- The fourth information source is the toxin regulations that set binding or suggested limits. The limits are binding for human consumption, while, for animal husbandry, suggested limits are provided for all except aflatoxins, as animals typically cannot be supplied with human-quality feed in agriculture. This is changing: as new research results on the critical toxicity of mycotoxins are being published and confirmed, the EFSA in Europe has suggested that the EU modify the relevant regulations [99,100,101]. Although the regulations do not have the same limits in all countries, human health and feed export/import considerations are controlled. For example, when an individual wishes to export maize from Europe, they must further consider the European regulations in the case when their home country regulatory limit is less strict. A recent review has mentioned mycotoxins such as trichothecenes, fumonisins, zearalenone, aflatoxins, and ochratoxin A as the most-researched items [102]; however, useful data for breeding are available only for DON, fumonisins, and (to a lesser extent) aflatoxins, and they are far from perfect. Other mostly emerging mycotoxins, such as fusarenon-X, beauvericin, enniatins (A and B), and moniliformin, have been obtained from both raw and processed cereals and possible resistances are yet unknown.
5. Resistance to Ear Rots and Toxin Contamination
5.1. General Considerations
5.2. Gibberella Ear Rot
5.3. Fusarium Ear Rot
5.4. Aspergillus Ear Rot
5.5. Common or Diverse Resistance Mechanisms against Ear Rot Fungi
5.6. Resistance to Other Toxigenic Fungi and Their Toxin Accumulation
6. Influence of Climate Change on the Resistance Breeding Activity
7. Methodology Aspects
8. Resistance and Agronomic Practices
9. Breeding Aspects
10. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AER | Aspergillus ear rot |
| AFB1 | aflatoxin B1 |
| AFM1 | aflatoxin M1 |
| Afs | aflatoxins |
| ALT | altenuene |
| AME | alternariol monomethyl ether |
| AOH | alternariol |
| BEA | beauvericin |
| CIT | citrinin |
| DEG | differently expressing gene |
| DH-CIT | dihydrocitrinone |
| DON | deoxynivalenol |
| DON-15GlcA | DON-15-β-D-O-glucuronide A |
| ENB | enniatin B |
| FAO | Food and Agricultural Organization of United Nations |
| FB1 | fumonisin B1 |
| FBs | fumonisins |
| FDK | Fusarium-damaged kernel |
| FER | Fusarium ear rot |
| FHB | Fusarium head blight |
| FUM | fumonisin(s) |
| GCA | general combining ability |
| GCA | general combining ability |
| GCA | general combining ability |
| GCA | general combining ability |
| GER | Gibberella ear rot |
| GM | genetically modified |
| GOSZ-VSZT | National Association of Hungarian Cereal Producers |
| GUS | beta glucuronidase |
| GWAM | genome-wide association mapping |
| GWAS | genome-wide association study |
| HIGS | host-induced gene silencing |
| HT-2-3-GlcA | HT-2-glucuronide |
| MON | moniliformin |
| MQTL | meta-QTL |
| NIV | nivalenol |
| OTA | ochratoxin A |
| PCR | polymerase chain reaction |
| QDR | quantitatively inherited resistance |
| QTL | quantitative trait locus |
| RIL | recombinant inbred line |
| SCA | specific combining ability |
| SNP | single-nucleotide polymorphism |
| ZEN | zearalenone |
References
- Magan, N.; Medina, A.; Aldred, D. Possible climate-change effects on mycotoxin contamination of food crops pre- and postharvest. Plant Pathol. 2011, 60, 150–163. [Google Scholar] [CrossRef]
- Guan, X.; Feng, Y.; Suo, D.; Xiao, Z.; Wang, S.; Liang, Y.; Fan, X. Simultaneous Determination of 11 Mycotoxins in maize via multiple-impurity adsorption combined with liquid chromatography–tandem mass spectrometry. Foods 2022, 11, 3624. [Google Scholar] [CrossRef] [PubMed]
- Munkvold, G.P.; White, D.G. (Eds.) Compendium of Corn Diseases; The American Phytopathological Society (APS) Press: Saint Paul, MN, USA, 2016; 165p. [Google Scholar]
- Cary, J.W.; Rajasekaran, K.; Brown, R.L.; Luo, M.; Chen, Z.-Y.; Bhatnagar, D. Developing resistance to aflatoxin in maize and cottonseed. Toxins 2011, 3, 678–696. [Google Scholar] [CrossRef] [PubMed]
- Lanubile, A.; Machietto, V.; Marocco, A. Breeding maize for resistance to mycotoxins. In Mycotoxin Reduction in Grain Chains; Leslie, J.F., Logrieco, A.F., Eds.; Wiley Blackwell: Ames, IA, USA; Chichester, UK, 2014; pp. 37–58. ISBN 978-0-8138-2083-5. [Google Scholar]
- Campbell, K.W.; White, D.G. Evaluation of corn genotypes for resistance to Aspergillus ear rot, kernel infection, and aflatoxin production. Plant Dis. 1995, 79, 1039–1045. [Google Scholar] [CrossRef]
- Bhatnagar, D.; Rajasekaran, K.; Cary, J.W.; Brown, R.L.; Yu, J.; Cleveland, T.E. Molecular approaches to development of resistance to preharvest aflatoxin contamination. In Mycotoxins: Detection Methods, Management, Public Health, and Agricultural Trade; CABI (CAB International): Cambridge, MA, USA, 2008; pp. 257–276. [Google Scholar]
- Mesterházy, A.; Lemmens, M.; Reid, L.M. Breeding for resistance to ear rots caused by Fusarium spp. in maize—A review. Plant Breed. 2012, 131, 1–19. [Google Scholar] [CrossRef]
- Battilani, P.; Toscano, P.; van der Fels-Klerx, H.J.; Moretti, A.; Leggieri, M.C.; Brera, C.; Rortais, A.; Goumperis, T.; Robinson, T. Aflatoxin B1 contamination in maize in Europe increases due to climate change. Sci. Rep. 2016, 6, 24328. [Google Scholar] [CrossRef] [PubMed]
- Mesterhazy, A.; Szieberth, D.; Toldine, E.T.; Nagy, Z.; Szabó, B.; Herczig, B.; Bors, I.; Tóth, B. Updating the Methodology of Identifying Maize Hybrids Resistant to Ear Rot Pathogens and Their Toxins—Artificial Inoculation Tests for Kernel Resistance to Fusarium graminearum, F. verticillioides, and Aspergillus flavus. J. Fungi 2022, 8, 293. [Google Scholar] [CrossRef]
- Bandyopadhyay, R.; Atehnkeng, J.; Ortega-Beltran, A.; Akande, A.; Falade, T.D.O.; Cotty, P.J. “ground-truthing” efficacy of biological control for aflatoxin mitigation in farmers’ fields in Nigeria: From field trials to commercial usage, a 10-year study. Front. Microbiol. 2019, 10, 25281. [Google Scholar] [CrossRef]
- Kaale, L.D.; Kimanya, M.E.; Macha, L.J.; Mlalila, N. Aflatoxin contamination and recommendations to improve its control: A review. World Mycotoxin J. 2021, 14, 27–40. [Google Scholar] [CrossRef]
- Senatore, M.T.; Prodi, A.; Tini, F.; Balmas, V.; Infantino, A.; Onofri, A.; Eleonora Cappelletti, E.; Oufensou, S.; Sulyok, M.; Covarelli, L.; et al. Different diagnostic approaches for the characterization of the fungal community and Fusarium species complex composition of Italian durum wheat grain and correlation with secondary metabolite accumulation. J. Sci. Food Agric. 2023, 103, 4503–4521. [Google Scholar] [CrossRef]
- Afolabi, C.G.; Ojiambo, P.S.; Ekpo, E.J.A.; Menkir, A.; Bandyopadhyay, R. Evaluation of maize inbred lines for resistance to Fusarium ear rot and fumonisin accumulation in grain in tropical Africa. Plant Dis. 2007, 91, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Mesterházy, Á. Relationship between resistance to stalk rot and ear rot of corn influenced by rind resistance, premature death, and the rate of drying of the ear. Maydica 1983, 28, 425–437. [Google Scholar]
- Bernardi, J.; Stagnati, L.; Lucini, L.; Rocchetti, G.; Lanubile, A.; Cortellini, C.; De Poli, G.; Busconi, M.; Marocco, A. Phenolic profile and susceptibility to Fusarium infection of pigmented maize cultivars. Front. Plant Sci. 2018, 9, 1189. [Google Scholar] [CrossRef] [PubMed]
- Santiago, R.; Cao, A.; Butrón, A. Genetic factors involved in fumonisin accumulation in maize kernels and their implications in maize agronomic management and breeding. Toxins 2015, 7, 3267–3296. [Google Scholar] [CrossRef] [PubMed]
- Butrón, A.; Santiago, R.; Cao, A.; Samayoa, L.F.; Malvar, R.A. QTLs for resistance to Fusarium ear rot in a multiparent advanced generation intercross (MAGIC) maize population. Plant Dis. 2019, 103, 397–404. [Google Scholar] [CrossRef] [PubMed]
- Cao, A.; Santiago, R.; Ramos, A.J.; Souto, X.C.; Aguín, O.; Malvar, R.A.; Butrón, A. Critical environmental and genotypic factors for Fusarium verticillioides infection, fungal growth and fumonisin contamination in maize grown in northwestern Spain. Int. J. Food Microbiol. 2014, 177, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Cao, A.; Butron, A.; Ramos, A.J.; Marin, S.; Souto, C.; Santiago, R. Assessing white maize resistance to fumonisin contamination. Eur. J. Plant Pathol. 2014, 138, 283–292. [Google Scholar] [CrossRef]
- Cao, A.; de la Fuente, M.; Gesteiro, N.; Santiago, R.; Malvar, R.A.; Butrón, A. Genomics and pathways involved in maize resistance to Fusarium ear rot and kernel contamination with fumonisins. Front. Plant Sci. 2022, 13, 866478. [Google Scholar] [CrossRef]
- Chen, J.; Shrestha, R.; Ding, J.; Zheng, H.; Mu, C.; Wu, J.; Mahuku, G. Genome-wide association study and QTL mapping reveal genomic loci associated with Fusarium ear rot resistance in tropical maize germplasm. G3 Genes Genomes Genet. 2016, 6, 3803–3815. [Google Scholar] [CrossRef]
- Chiuraise, N.; Derera, J.; Yobo, K.S.; Magorokosho, C.; Nunkumar, A.; Qwabe, N.F.P. Progress in stacking aflatoxin and fumonisin contamination resistance genes in maize hybrids. Euphytica 2016, 207, 49–67. [Google Scholar] [CrossRef]
- Gesteiro, N.; Cao, A.; Santiago, R.; Malvar, R.A.; Butron, A. Genomics of maize resistance to kernel contamination with fumonisins using a multiparental advanced generation InterCross maize population (MAGIC). BMC Plant Biol. 2021, 21, 596. [Google Scholar] [CrossRef] [PubMed]
- Guche, M.D.; Pilati, S.; Trenti, F.; Dalla Costa, L.; Giorni, P.; Guella, G.; Marocco, A.; Lanubile, A. Functional study of lipoxygenase- mediated resistance against Fusarium verticillioides and Aspergillus flavus infection in maize. Int. J. Mol. Sci. 2022, 23, 10894. [Google Scholar] [CrossRef] [PubMed]
- Guo, B.; Ji, X.-G.; Ni, X.; Fountain, J.C.; Hong Li, H.; Abbas, H.K.; Lee, R.D.; Brian, T.; Scully, B.T. Evaluation of maize inbred lines for resistance to pre-harvest aflatoxin and fumonisin contamination. in the field. Crop J. 2017, 5, 259–264. [Google Scholar] [CrossRef]
- Henry, W.B.; Williams, W.P.; Gary, L.; Windham, G.L.; Hawkins, L.K. Evaluation of maize inbred lines for resistance to Aspergillus and Fusarium ear rot and mycotoxin accumulation. Agron. J. 2009, 101, 1219–1226. [Google Scholar] [CrossRef]
- Ivic, D.; Cabric, M.; Palaveršic, B.; Cvjetkovic, B. No correlation between pericarp thickness and Fusarium ear rot (Fusarium verticillioides) in Croatian maize hybrids and lines. Maydica 2008, 53, 297–301. [Google Scholar]
- Lanubile, A.; Ferrarini, A.; Maschietto, V.; Delledonne, M.; Marocco, A.; Bellin, D. Functional genomic analysis of constitutive and inducible defense responses to Fusarium verticillioides infection in maize genotypes with contrasting ear rot resistance. BMC Genom. 2014, 15, 710. Available online: http://www.biomedcentral.com/1471-2164/15/710 (accessed on 14 February 2023). [CrossRef]
- Lanubile, A.; Maschietto, V.; Borrelli, V.M.; Stagnati, L.; Logrieco, A.F.; Marocco, A. Molecular Basis of Resistance to Fusarium Ear Rot in Maize. Front. Plant Sci. 2017, 8, 1774. [Google Scholar] [CrossRef]
- Lanzanova, C.; Arrigoni, A.; Valoti, P.; Alfieri, M.; Locatelli, S. Agronomic performance, chemical composition, and Fusarium verticillioides resistance of Italian white maize varieties. Qual. Assur. Saf. Crops Foods 2019, 11, 549–559. [Google Scholar] [CrossRef]
- Bolduan, C.; Miedaner, T.; Schipprack, W.; Dhillon, B.S.; Melchinger, A.E. Genetic variation for resistance to ear rots and mycotoxins contamination in early European maize inbred lines. Crop Sci. 2009, 49, 2019–2028. [Google Scholar] [CrossRef]
- Loeffler, M.; Kessel, B.; Ouzunova, M.; Miedaner, T. Population parameters for resistance to Fusarium graminearum and Fusarium verticillioides ear rot among large sets of early, mid-late and late maturing European maize (Zea mays L.) inbred lines. Theor. Appl. Genet. 2010, 120, 1053–1062. [Google Scholar] [CrossRef]
- Schjøth, J.E.; Visconti, A.; Sundheim, L. Fumonisins in maize in relation to climate, planting time and hybrids in two agroecological zones in Zambia. Mycopathologia 2009, 167, 209–219. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Zhou, Z.; Dong, C.; Chen, J.; Ding, J.; Zhang, X.; Mu, C.; Chen, Y.; Li, X.P.; Li, H.; et al. Linkage mapping and genome-wide association study reveals conservative QTL and candidate genes for Fusarium rot resistance in maize. BMC Genom. 2020, 21, 357. [Google Scholar] [CrossRef] [PubMed]
- Zila, C.T.; Ogut, F.; Romay, M.C.; Gardner, C.A.; Buckler, E.S.; Holland, J.B. Genome-wide association study of fusarium ear rot disease in the USA maize inbred line collection. BMC Plant Biol. 2014, 14, 372. [Google Scholar] [CrossRef] [PubMed]
- Abbas, H.K.; Mascagni, H.J., Jr.; Bruns, H.A.; Shier, W.T.; Damann, K.E. Effect of planting density, irrigation regimes, and maize hybrids with varying ear size on yield, and aflatoxin and fumonisin contamination levels. Am. J. Plant Sci. 2012, 3, 1341–1354. [Google Scholar] [CrossRef]
- Henry, W.B.; Windham, G.L.; Blanco, M.H. Evaluation of maize germplasm for resistance to aflatoxin accumulation. Agronomy 2012, 2, 28–39. [Google Scholar] [CrossRef]
- Warburton, M.L.; Williams, W.P. Aflatoxin Resistance in Maize: What Have We Learned Lately? Adv. Bot. 2014, 2014, 352831. [Google Scholar] [CrossRef]
- Ali, L.K.; Jeff, H.; Taylor, L.J.; Sun, G.; William, M.; Kasha, K.J.; Reid, L.M.; Pauls, K.P. Molecular mapping of QTLs for resistance to Gibberella ear rot, in corn, caused by Fusarium graminearum. Genome 2005, 48, 521–533. [Google Scholar] [CrossRef] [PubMed]
- Park, D.L.; Njapau, H.; Boutrif, E. Minimizing risks posed by mycotoxins utilizing the HACCP concept. FAO Food. Nutr. Agric. J. 1999, 23, 49–56. [Google Scholar]
- Galiano-Carneiro, A.L.; Kessel, B.; Presterl, T.; Gaikpa, D.S.; Kistner, M.B.; Miedaner, T. Multi-parent QTL mapping reveals stable QTL conferring resistance to Gibberella ear rot in maize. Euphytica 2021, 217, 2. [Google Scholar] [CrossRef]
- Kebede, A.Z.; Woldemariam, T.; Reid, L.M.; Harris, L.J. Quantitative trait loci mapping for Gibberella ear rot resistance and associated agronomic traits using genotyping-by-sequencing in maize. Theor. Appl. Genet. 2016, 129, 17–29. [Google Scholar] [CrossRef]
- Reid, L.M.; Hamilton, R.E.; Mather, D.E. Screening Maize for Resistance to Gibberella Ear Rot; Publication 1996–5E; Agriculture and Agri-Food Canada; Technical Bulletin: Ottawa, ON, Canada, 1996; 62p. [Google Scholar]
- Reid, L.M.; Hamilton, R.I.; Mather, D.E. Effect of macroconidial suspension volume and concentration on expression of resistance to 1749 Fusarium graminearum in maize. Plant Dis. 1995, 79, 461–466. [Google Scholar] [CrossRef]
- Zhou, G.; Li, S.; Ma, L.; Wang, F.; Jiang, F.; Sun, Y.; Ruan, X.; Cao, Y.; Wang, Q.; Zhang, Y.; et al. Mapping and validation of a stable quantitative trait locus conferring maize resistance to Gibberella Ear Rot. Plant Dis. 2021, 105, 1984–1991. [Google Scholar] [CrossRef] [PubMed]
- Schaafsma, A.W.; Nicol, R.W.; Reid, L.M. Evaluating commercial maize hybrids for resistance to Gibberella ear rot. Eur. J. Plant Path. 1997, 103, 737–746. [Google Scholar] [CrossRef]
- Schaafsma, A.W.; Miller, J.D.; Savard, M.E.; Ewing, R.J. Ear rot development and mycotoxin production in corn in relation to inoculation method, corn hybrid, and species of Fusarium. Can. J. Plant Pathol. 1993, 15, 185–192. [Google Scholar] [CrossRef]
- Stagnati, I.; Martino, M.; Battilani, P.; Busconi, M.; Lanubile, A.; Marocco, A. Development of early maturity maize hybrids for resistance to Fusarium and Aspergillus ear rots and their associated mycotoxins. World Mycotoxin J. 2020, 13, 459–471. [Google Scholar] [CrossRef]
- Szabo, B.; Toth, B.; Toth Toldine, E.; Varga, M.; Kovacs, N.; Varga, J.; Kocsube, S.; Palagyi, A.; Bagi, F.; Budakov, D.; et al. a new concept to secure food safety standards against Fusarium species and Aspergillus flavus and their toxins in maize. Toxins 2018, 10, 372. [Google Scholar] [CrossRef] [PubMed]
- Mesterhazy, A.; Szieberth, D.; Tóth, E.T.; Nagy, Z.; Szabó, B.; Herczig, B.; Bors, I.; Tóth, B. The role of preharvest natural infection and toxin contamination in food and feed safety in maize, South-East Hungary, 2014–2021. J. Fungi 2022, 8, 1104. [Google Scholar] [CrossRef]
- Mesterhazy, A.; Toldine Toth, E.; Szel, S.; Varga, M.; Toth, B. Resistance of maize hybrids to Fusarium graminearum, F. culmorum, and F. verticillioides ear rots with toothpick and silk channel inoculation, as well as their toxin production. Agronomy 2020, 10, 1283. [Google Scholar] [CrossRef]
- Mesterházy, Á.; Bartók, T.; Lamper, C. Influence of cultivar resistance, epidemic severity, and Fusarium species on the efficacy of fungicide control of Fusarium head blight in wheat and deoxynivalenol (DON) contamination of grain. Plant Dis. 2003, 87, 1107–1115. [Google Scholar] [CrossRef]
- Mesterházy, Á.; Varga, M.; Tóth, B.; Kótai, C.; Bartók, T.; Véha, A.; Ács, K.; Vágvölgyi, C.; Lehoczki-Krsjak, S. Reduction of deoxynivalenol (DON) contamination by improved fungicid e use in wheat. Part 1. Dependence on epidemic severity and resistance level in small plot. Tests with artificial inoculation. Eur. J. Plant Pathol. 2018, 151, 39–55. [Google Scholar] [CrossRef]
- Eskola, M.; Kos, G.; Elliott, C.T.; Hajslova, J.; Mayar, S.; Krska, R. Worldwide contamination of food-crops with mycotoxins: Validity of the widely cited ‘FAO estimate’ of 25%. Crit. Rev. Food Sci. Nutr. 2019, 60, 2773–2789. [Google Scholar] [CrossRef]
- Available online: https://www.atlasbig.com/en-ae/countries-by-corn-production (accessed on 9 December 2023).
- Mesterházy, Á.; Oláh, J.; Popp, J. Losses in the grain supply chain: Causes and solutions. Sustainability 2020, 12, 2342. [Google Scholar] [CrossRef]
- GOSZ-VSZT-NAK. Kukorica Posztregisztrációs Fajtakísérletek (Maize Post Registration Variety Tests for Grain Hybrids, 2021. Innovation Council of Variety Testing. 44p. Available online: https://vszt.hu/wp-content/uploads/2023/09/gosz_vszt_nak_kukorica_2021.pdf (accessed on 11 October 2023). (In Hungarian).
- GOSZ-VSZT-NAK. Kukorica Posztregisztrációs Fajtakísérletek (Maize Post Registration Variety Tests for Grain Hybrids, 2020. Innovation Council of Variety Testing. 44p. Available online: https://vszt.hu/wp-content/uploads/2023/09/gosz_vszt_nak_kukorica_2020.pdf (accessed on 11 October 2023). (In Hungarian).
- Gaikpa, D.S.; Kessel, B.; Presterl, T.; Ouzunova, M.; Galiano-Carneiro, A.L.; Mayer, M.; Melchinger, A.M.; Schön, C.-C.; Miedaner, T. Exploiting genetic diversity in two European maize landraces for improving Gibberella ear rot resistance using genomic tools. Theo. Appl. Genet. 2021, 134, 793–805. [Google Scholar] [CrossRef] [PubMed]
- Foley, D.C. Systemic infection of corn by Fusarium moniliforme. Phytopathology 1962, 52, 870–872. [Google Scholar]
- Miedaner, T.; Juroszek, P. Global warming and increasing maize cultivation demand comprehensive efforts in disease and insect resistance breeding in north-western Europe. Plant Pathol. 2021, 70, 1032–1046. [Google Scholar] [CrossRef]
- McMillan, W.W. Maize plant resistance to insect damage and associated aflatoxin development. In Aflatoxin in Maize: Proceedings of the Workshop; Zuber, M.S., Lillehoj, E.B., Renfro, B.L., Eds.; CIMMYT: México-Veracruz, Mexico, 1987; pp. 250–253. [Google Scholar]
- Zhou, D.; Wang, X.; Chen, G.; Sun, S.; Yang, Y.; Zhu, Z.; Duan, C. The major Fusarium species causing maize ear and kernel rot and their toxigenicity in Chongqing, China. Toxins 2018, 10, 90. [Google Scholar] [CrossRef] [PubMed]
- Bartok, T.; Tolgyesi, L.; Mesterházy, A.; Bartok, M.; Szecsi, A. Identification of the first fumonisin mycotoxins with three acyl groups by ESI-ITMS and ESI-TOFMS following RP-HPLC separation: Palmitoyl, linoleoyl and oleoyl EFB1 fumonisin isomers from a solid culture of Fusarium verticillioides. Food Addit. Contam. A 2010, 27, 1714–1723. [Google Scholar] [CrossRef]
- Bartók, T.; Tölgyesi, L.; Szekeres, A.; Varga, M.; Bartha, R.; Szécsi, Á.; Bartók, M.; Mesterházy, Á. Detection and characterization of twenty-eight isomers of fumonisin B1 (FB1) mycotoxin in a solid rice culture infected with Fusarium verticillioides by reserved phase high-performance liquid chromatography/electro spray ionization time-of-flight and ion trap mass spectrometry. Rapid Commun. Mass. Spectrom. 2010, 24, 35–42. [Google Scholar]
- Bartok, T.; Szecsi, A.; Juhasz, K.; Bartok, M.; Mesterházy, A. ESI-MS and MS/MS identification of the first ceramide analogues of fumonisin B-1 mycotoxin from a Fusarium verticillioides culture following RP-HPLC separation. Food Addit. Contam. Part A 2013, 30, 1651–1659. [Google Scholar] [CrossRef]
- Bartok, T.; Tölgyesi, L.; Szecsi, A.; Varga, J.; Bartok, M.; Mesterházy, A.; Gyimes, E.; Veha, A. Identification of unknown isomers of fumonisin b-5 mycotoxin in a Fusarium verticillioides culture by high-performance liquid chromatography/electrospray ionization time-of-flight and ion trap mass spectrometry. J. Liq. Chrom. Relat. Technol. 2013, 36, 1549–1561. [Google Scholar] [CrossRef]
- Ghiasian, S.A.; Kord-Bacheh, P.; Rezayat, S.M.; Maghsood, H.A.; Taherkhani, H. Mycoflora of Iranian maize harvested in the main production areas in 2000. Mycopathologia 2004, 158, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Castañares, E.; Martínez, M.; Cristos, D.; Rojas, D.; Lara, B.; Stenglein, S.; Dinolfo, M.I. Fusarium species and mycotoxin contamination in maize in Buenos Aires province, Argentina. Eur. J. Plant Pathol. 2019, 155, 1265–1275. [Google Scholar] [CrossRef]
- Dorn, B.; Forrer, H.-F.; Schürch, S.; Vogelgsang, S. Fusarium species complex on maize in Switzerland: Occurrence, prevalence, impact, and mycotoxins in commercial hybrids under natural infection. Eur. J. Plant Pathol. 2009, 125, 51–61. [Google Scholar] [CrossRef]
- Logrieco, A.F.; Battilani, P.; Camardo Leggieri, M.; Haesaert, G.; Jiang, Y.; Lanubile, A.; Mahuku, G.; Mesterhazy, A.; Ortega-Beltran, A.; Pasti, M.A.; et al. Perspectives on global mycotoxin issues and management from the MycoKey Maize Working Group. Plant Dis. 2021, 105, 525–537. [Google Scholar] [CrossRef] [PubMed]
- Logrieco, A.; Mule, G.; Moretti, A.; Bottalico, A. Toxigenic Fusarium species and mycotoxins associated with maize ear rot in Europe. Eur. J. Plant Pathol. 2002, 108, 597–609. [Google Scholar] [CrossRef]
- Almeida, A.P.; Corrêa, B.; Marisa, A.B.; Mallozzi, M.A.B.; Sawazaki, E.; Valente Soares, L.M. Mycoflora and aflatoxin/fumonisin production by fungal isolates from freshly harvested corn hybrids. Braz. J. Microbiol. 2000, 31, 321–326. [Google Scholar] [CrossRef]
- Mesterházy, Á.; Vojtovics, M. A kukorica Fusarium okozta fertõzöttségének vizsgálata 1972–1975-ben. (Investigation of Fusarium species in corn kernels 1972–1975). Növénytermelés 1977, 26, 367–378. [Google Scholar]
- Alexander, N.J.; Proctor, R.H.; McCormick, S.P. Genes, gene clusters, and biosynthesis of trichothecenes and fumonisins in Fusarium. Toxin Rev. 2009, 28, 198–215. [Google Scholar] [CrossRef]
- Ding, L.; Xu, H.; Yi, H.; Yang, L.; Kong, Z.; Zhang, L.; Xue, S.; Jia, H.; Ma, Z. Resistance to hemi-biotrophic F. graminearum infection is associated with coordinated and ordered expression of diverse defense signaling pathways. PLoS ONE 2011, 6, e19008. [Google Scholar] [CrossRef]
- Hao, G.; McCormick, S.; Usgaard, T.; Tiley, H.; Vaughan, M.M. Characterization of three Fusarium graminearum effectors and their roles during Fusarium head blight. Front. Plant Sci. 2020, 11, 579553. [Google Scholar] [CrossRef]
- Christensen, C.M.; Kaufmann, H.H. Grain Storage; The University of Minnesota Press: Minneapolis, MN, USA, 1969; 153p. [Google Scholar]
- Varga, J.; Tóth, B.; Mesterhazy, A.; Teren, J.; Fazekas, B. Mycotoxigenic fungi and mycotoxins in foods and feeds in Hungary. In An Overview on Toxigenic Fungi and Mycotoxins in Europe; Logrieco, A., Visconti, A., Eds.; Kluwer Acedemic Publishers: Dordecht, The Netherlands, 2004; pp. 123–139. 252p, ISBN 1-4020-2645-5. [Google Scholar]
- Shanon, G.M.; Rodricks, J.V.; Hesseltine, C.W.; Shotwell, O.L.; Mehlman, M.A. (Eds.) Minicolumn Detection methods for aflatoxin in yellow corn. J. Assoc. Off. Anal. Chem. 1979, 62, 1070–1075. [Google Scholar]
- Anderson, H.W.; Nehring, E.W.; Wichser, W.R. Aflatoxin contamination of com in the field. J. Agric. Food Chem. 1975, 23, 775–782. [Google Scholar] [CrossRef] [PubMed]
- Lillehoj, E.B.; Kwolek, W.F.; Fennell, D.I.; Milburn, M.S. Aflatoxin incidence and association with bright greenish yellow fluorescence and insect damage in a limited survey of freshly harvested high-moisture com. Cereal Chem. 1975, 52, 403–412. [Google Scholar]
- Lillehoj, E.B.; Kwolek, W.F.; Vandegraft, E.E.; Zuber, M.S.; Calvert, O.H.; Widstrom, N.; Futrell, M.C.; Bockholt, A.J. Aflatoxin production in Aspergillus flavus inoculated ears of corn grown at diverse locations. Crop Sci. 1975, 15, 267–270. [Google Scholar] [CrossRef]
- Widstrom, N.W. Breeding strategies to control aflatoxin contamination of maize through host plant resistance. In Aflatoxin in Maize: Proceedings of the Workshop; Zuber, M.S., Lillehoj, E.B., Renfro, B.L., Eds.; CIMMYT: México-Veracruz, El Batan, Mexico, 1987; pp. 212–220. [Google Scholar]
- Mesterhazy, A. Updating the breeding philosophy of wheat to Fusarium head blight (FHB): Resistance components, QTL identification and phenotyping- a review. Plants 2020, 9, 1702. [Google Scholar] [CrossRef] [PubMed]
- Mesterházy, Á.; Bartók, T.; Kászonyi, G.; Varga, M.; Tóth, B.; Varga, J. Common resistance to different Fusarium spp. causing Fusarium head blight in wheat. Eur. J. Plant Path. 2005, 112, 267–281. [Google Scholar] [CrossRef]
- Mesterházy, Á. Role of deoxynivalenol in aggressiveness of Fusarium graminearum and F. culmorum and in resistance to Fusarium head blight. Eur. J. Plant Pathol. 2002, 108, 675–684. [Google Scholar] [CrossRef]
- Darnetty, T.; Salleh, B. Toxigenicity of Fusarium species in Gibberella fujikuroi species complex (GFSC) associated with stalk and ear rot disease of corn. ESci. J. Plant Pathol. 2013, 2, 147–154. [Google Scholar] [CrossRef]
- Adejumo, T.O.; Adejoro, D.O. Incidence of aflatoxins, fumonisins, trichothecenes and ochratoxins in Nigerian foods and possible intervention strategies. Food Sci. Qual. Manag. 2014, 31, 127–146. [Google Scholar]
- Park, J.; Kim, D.-H.; Moon, J.-Y.; An, J.-A.; Kim, Y.-W.; Chung, S.-H.; Lee, C. Distribution analysis of twelve mycotoxins in corn, and corn-derived products by LC-MS/MS to evaluate the carry-over ratio during wet-milling. Toxins 2018, 10, 319. [Google Scholar] [CrossRef]
- Eckard, S.; Wettstein, F.E.; Forrer, H.-R.; Vogelgsang, S. Incidence of Fusarium species and mycotoxins in silage maize. Toxins 2011, 3, 949–967. [Google Scholar] [CrossRef]
- Mihalache, O.A.; De Boevre, M.; Dellafiora, L.; De Saeger, S.; Moretti, A.; Pinson-Gadais, L.; Ponts, N.; Richard-Forget, F.; Susca, A.; Dall’Asta, C. The occurrence of non-regulated mycotoxins in foods: A systematic review. Toxins 2023, 15, 583. [Google Scholar] [CrossRef] [PubMed]
- Lemming, E.W.; Montano, M.A.; Schmidt, J.; Cramer, B.; Humpf, H.U.; Moraeus, L.; Olsen, M. Mycotoxins in blood and urine of Swedish adolescents-possible associations to food intake and other background characteristics. Mycotoxin Res. 2020, 36, 193–206. [Google Scholar] [CrossRef] [PubMed]
- Izzo, L.; Rodríguez-Carrasco, Y.R.; Tolosa, J.; Graziani, G.; Gaspari, A. Target analysis and retrospective screening of mycotoxins and pharmacologically active substances in milk using an ultra-high-performance liquid chromatography/high-resolution mass spectrometry approach. J. Dairy Sci. 2020, 103, 1250–1260. [Google Scholar] [CrossRef] [PubMed]
- Abia, W.A.; Warth, B.; Sulyok, M.; Krska, R.; Tchana, A.; Njobeh, P.B.; Turner, P.C.; Kouanfack, C.; Eyongetah, M.; Dutton, M.; et al. Bio-monitoring of mycotoxin exposure in Cameroon using a urinary multi-biomarker approach. Food Chem. Toxicol. 2013, 62, 927–934. [Google Scholar] [CrossRef]
- De Santis, B.; Debegnach, F.; Toscano, P.; Crisci, A.; Battilani, P.; Brera, C. Overall Exposure of European adult population to mycotoxins by statistically modelled biomonitoring data. Toxins 2021, 13, 695. [Google Scholar] [CrossRef]
- Schmidt, J.; Cramer, B.; Turner, P.C.; Stoltzfus, R.J.; Humphrey, J.H.; Smith, L.E.; Humpf, H.-U. Determination of urinary mycotoxin biomarkers using a sensitive online solid phase extraction-UHPLC-MS/MS method. Toxins 2021, 13, 418. [Google Scholar] [CrossRef]
- Tarazona, A.; Gómez, J.V.; Mateo, F.; Jiménez, M.; Romera, D.; Mateo, E.M. Study on mycotoxin contamination of maize kernels in Spain. Food Control 2020, 118, 107370. [Google Scholar] [CrossRef]
- EC1 Document 32006R1881; Commission Regulation (EC) No 1881/2006 of 19 December 2006 Setting Maximum Levels for Certain Contaminants in Foodstuffs. Official Journal of the European Union: Brussels, Belgium, 2006.
- EC2 Document 32002L0032; Commission Regulation Directive 2002/32/EC of the European Parliament and of the Council of 7 May 2002 on Undesirable Substances in Animal Feed. Official Journal of the European Union: Brussels, Belgium, 2002.
- EC3 Commission Regulation 2006/576/EC; Commission Recommendation of 17 August 2006 on the Presence of Deoxynivalenol, Zearalenone, Ochratoxin A, T-2 and HT-2 and Fumonisins in Products Intended for Animal Feeding. Official Journal of the European Union: Brussels, Belgium, 2006; Volume I. 229, pp. 7–9.
- Pereira, V.L.; Fernandes, J.O.; Cunha, S.C. Mycotoxins in cereals and related foodstuffs: A review on occurrence and recent methods of analysis. Trends Food Sci. Technol. 2014, 36, 96–136. [Google Scholar] [CrossRef]
- James, M., Ed.; Grand Geneva Resort. In Proceedings of the 47th Annual Maize Genetics Conference, Lake Geneva, WI, USA, 10–13 March 2005; p. 191. [Google Scholar]
- Hollick, J. Program and Abstracts. In Proceedings of the 48th Annual Maize Genetics Conference, Pacific Grobve, CA, USA, 9–12 March 2006. 209p. [Google Scholar]
- Brutnell, T. Program and Abstracts. In Proceedings of the 50th Annual Maize Genetics Conference, Marriott—Wardman Park, Washington, DC, USA, 27 February–1 March 2008. 207p. [Google Scholar]
- Sparks, E. Program and Abstracts. In Proceedings of the 64th Annual Maize Genetics Meeting, St. Louis, MO, USA, 31 March–3 April 2022. 222p. [Google Scholar]
- Moretti, A.; Pasquale, M.; Logrieco, A.F. Mycotoxin risks under a climate change scenario in Europe. Trends Food Sci. Technol. 2019, 84, 38–40. [Google Scholar] [CrossRef]
- Dövényi-Nagy, T.; Rácz, C.; Molnár, K.; Bakó, K.; Szláma, Z.; Jóźwiak, Á.; Farkas, Z.; Pócsi, I.; Dobos, A.C. Pre-harvest modelling and mitigation of aflatoxins in maize in a changing climatic environment, A review. Toxins 2020, 12, 768. [Google Scholar] [CrossRef] [PubMed]
- Tirado, M.C.; Clarke, R.; Jaykus, L.A.; McQuatters-Gollop, A.; Frank, J.M. Climate change and food safety: A review. Food Res. Int. 2010, 43, 1745–1765. [Google Scholar] [CrossRef]
- Ortega-Beltran, A.; Bandyopadhyay, R. Aflatoxin biocontrol in practice requires a multidisciplinary, long-term approach. Front. Sustain. Food Syst. 2023, 7, 1110964. [Google Scholar] [CrossRef]
- Ouadhene, M.A.; Ortega-Beltran, A.; Sanna, M.; Cotty, P.J.; Battilani, P. Multiple Year influences of the aflatoxin biocontrol product af-x1 on the A. flavus communities associated with maize production in Italy. Toxins 2023, 15, 184. [Google Scholar] [CrossRef]
- Liao, X.-Y. Physical resistance: Resistance: A different perspective on maize ear rot resistance. Plant Growth Regul. 2023, 100, 573–576. [Google Scholar] [CrossRef]
- Sampietro, D.A.; Vattuone, M.A.; Presello, D.A.; Fauguel, C.M.; Catala, C.A.N. The pericarp and its surface wax layer in maize kernels as resistance factors to fumonisin accumulation by Fusarium verticillioides. Crop Prot. 2009, 28, 196–200. [Google Scholar] [CrossRef]
- Sampietro, D.A.; Fauguel, C.M.; Vattuone, M.A.; Presello, D.A.; Catalán, C.A.N. Phenylpropanoids from maize pericarp: Resistance factors to kernel infection and fumonisin accumulation by Fusarium verticillioides. Eur. J. Plant Pathol. 2013, 135, 105–113. [Google Scholar] [CrossRef]
- Mesterházy, Á.; Kovács, G., Jr.; Kovács, K. Breeding Resistance for Fusarium Ear Rot (FER) in Corn. In Proceedings of the 18th International Conference on Maize and Sorghum, Genetics and Breeding, Eucarpia, Beograd, Acta Biologica Yugoslavia, Serija F. Genetika, Belgrade, Yugoslavia, 4–9 June 2000; Volume 32, pp. 495–505. [Google Scholar]
- Araújo, S.; Júnior, O.; Viteri, L.; Aguiar, R.; Oliveira, E.; Santos, G. Efficiency of inoculation methods for genotypes selection in corn ear rot disease studies. Rev. Cienc. Agrícolas 2022, 39, 160–173. [Google Scholar] [CrossRef]
- Miedaner, T.; Boeven, A.L.G.-C.; Gaikpa, D.S.; Kistner, M.B.; Grote, C.P. Genomics-assisted breeding for quantitative disease resistances in small-grain cereals and maize. Int. J. Mol. Sci. 2020, 21, 9717. [Google Scholar] [CrossRef]
- McIntosh, R.; Wellings, C.R.; Park, R.F. Wheat Rusts: An Atlas of Resistance Genes; CSIRO Publications: East Melbourne, Australia, 1995; ISBN 0-7923-3430-2. [Google Scholar]
- Winzeler, M.; Mesterházy, Á.; Park, R.F.; Bartos, P.; Csosz, M.; Goyeau, H.; Ittu, M.; Jones, E.; Loschenberger, F.; Manninger, K.; et al. Resistance of European winter wheat germplasm to leaf rust. Agronomie 2000, 20, 783–792. [Google Scholar] [CrossRef]
- Munkvold, G.P. Epidemiology of Fusarium diseases and their mycotoxins in maize ears. Eur. J. Plant Pathol. 2003, 109, 705–713. [Google Scholar] [CrossRef]
- Munkvold, G.P. Cultural and genetic approaches to managing mycotoxins in maize. Ann. Rev. Phytopathol. 2003, 41, 99–116. [Google Scholar] [CrossRef] [PubMed]
- Hung, H.-Y.; Holland, J.B. Diallel analysis of resistance to Fusarium ear rot and fumonisin contamination in maize. Crop Sci. 2012, 52, 2173–2181. [Google Scholar] [CrossRef]
- Munkvold, G.P. Crop management practices to minimize the risk of mycotoxin contamination in temperate zone maize. In Mycotoxin Reduction in Grain Chains; Leslie, J.F., Logrieco, A.F., Eds.; John Wiley and Sons, Inc.: Ames, IA, USA; Chichester, UK; Oxford, UK; Blackwell Publishing: Danver, MA, USA, 2014; pp. 59–77, 352. [Google Scholar]
- Santiago, R.; Cao, A.; Malvar, R.A.; Reid, L.M.; Butron, A. Assessment of corn resistance to fumonisin accumulation in a broad collection of inbred lines. Field Crop. Res. 2013, 149, 193–202. [Google Scholar] [CrossRef]
- Mesterhazy, A.; Szabó, B.; Szél, S.; Nagy, Z.; Berényi, A.; Tóth, B. Novel insights into the inheritance of Gibberella ear rot (GER), deoxynivalenol (DON) accumulation, and DON production. Toxins 2022, 14, 583. [Google Scholar] [CrossRef]
- Miller, S.S.; Reid, L.M.; Harris, L.J. Colonization of maize silks by Fusarium graminearum, the causative organism of Gibberella ear rot. Can. J. Bot. 2007, 85, 369–376. [Google Scholar] [CrossRef]
- Reid, L.M.; Hamilton, R.I. Effects of inoculation position, timing, macroconidial concentration and irrigation on resistance of maize to F. graminearum infection through kernels. Can. J. Plant Path. 1996, 18, 279–285. [Google Scholar] [CrossRef]
- Giomi, G.M.; Kreff, E.D.; Iglesias, J.; Fauguel, C.M.; Fernandez, M.; Oviedo, M.S.; Presello, D.A. Quantitative trait loci for Fusarium and Gibberella ear rot resistance in Argentinian maize germplasm. Euphytica 2016, 211, 287–294. [Google Scholar] [CrossRef]
- Brauner, P.C.; Melchinger, A.E.; Schrag, T.A.; Utz, H.F.; Schipprack, W.; Kessel, B.; Ouzunova, M.; Miedaner, T. Low validation rate of quantitative trait loci for Gibberella ear rot resistance in European maize. Theor. Appl. Genet. 2017, 130, 175–186. [Google Scholar] [CrossRef]
- Yuan, J.; Ali, M.L.; Taylor, J.; Liu, J.; Sun, G.; Liu, W.; Masilimany, P.; Gulati, A.; Sakhuja, G.A.; Pauls, K.P. A guanylyl cyclase-like gene is associated with Gibberella ear rot resistance in maize (Zea mays L.). Theor. Appl. Genet. 2008, 116, 465–479. [Google Scholar] [CrossRef]
- Martin, M.; Miedaner, T.; Dhillon, B.S.; Ufermann, U.; Kessel, B.; Ouzunova, M.; Schipprack, W.; Melchinger, A.E. Colocalization of QTL for Gibberella ear rot resistance and low mycotoxin contamination in early European maize. Crop Sci. 2011, 51, 1935–1945. [Google Scholar] [CrossRef]
- Yang, T.; Stoopen, G.; Yalpani, N.; Vervoort, J.; Vos, R.; Voster, A.; Francel, W.A.; Verstappen, F.W.A.; Bouwmeester, H.J.; Maarten, A.; et al. Metabolic engineering of geranic acid in maize to achieve fungal resistance is compromised by novel glycosylation patterns. Metab. Eng. 2011, 13, 414–425. [Google Scholar] [CrossRef] [PubMed]
- Kant, P.; Liu, W.-Z.; Pauls, K.P. PDC1, a corn defensin peptide expressed in Escherichia coli and Pichia pastoris inhibits growth of Fusarium graminearum. Peptides 2009, 30, 1593–1599. [Google Scholar] [CrossRef] [PubMed]
- Szabó-Hevér, Á.; Lehoczki-Krsjak, S.; Tóth, B.; Purnhauser, L.; Buerstmayr, H.; Steiner, B.; Mesterházy, Á. Identification and validation of Fusarium head blight and Fusarium damaged kernel QTL in the Frontana/Remus DH mapping population. Can. J. Plant Pathol. 2012, 34, 224–238. [Google Scholar] [CrossRef]
- Szabó-Hevér, Á.; Lehoczki-Krsjak, S.; Varga, M.; Purnhauser, L.; Pauk, J.; Lantos, C.; Mesterházy, Á. 2014 Differential influence of QTL linked to Fusarium head blight, Fusarium-damaged kernel, deoxynivalenol contents and associated morphological traits in a Frontana-derived. Wheat population. Euphytica 2014, 200, 9–26. [Google Scholar] [CrossRef]
- Desjardins, A.E.; Munkvold, G.P.; Plattner, R.D.; Proctor, R.H. Fum1—A gene required for fumonisin biosynthesis but not for maize ear rot and ear infection by Gibberella moniliformis in field tests. Mol. Plant–Microbe Interact. 2002, 15, 1157–1164. [Google Scholar] [CrossRef] [PubMed]
- Balconi, C.; Berardo, N.; Locatelli, S.; Lanzanova, C.; Torri, A.; Redaelli, R. Evaluation of ear rot (Fusarium verticillioides) resistance and fumonisin accumulation in Italian maize inbred lines. Phytopath. Mediterr. 2014, 53, 14–26. [Google Scholar] [CrossRef]
- Desjardins, A.E.; Plattner, R.D.; Nelsen, T.; Leslie, J. Genetic analysis of fumonisin production and virulence of Gibberella fujikuroi mating population A (Fusarium moniliforme) on maize (Zea mays) seedlings. Appl. Environ. Microbiol. 1995, 61, 79–86. [Google Scholar] [CrossRef]
- Warfield, C.Y.; Davis, R.M. Importance of husk covering on the susceptibility of corn hybrids to Fusarium ear rot. Plant Dis. 1996, 80, 208–210. [Google Scholar] [CrossRef]
- Giomi, G.M.; Sampietro, D.A.; Velazco, J.G.; Iglesias, J.; Fernández, M.; Oviedo, M.S.; Presello, D.A. Map overlapping of QTL for resistance to Fusarium ear rot and associated traits in maize. Euphytica 2021, 217, 81. [Google Scholar] [CrossRef]
- Lanubile, A.; Maschietto, V.; De Leonardis, S.; Battilani, P.; Paciolla, C.; Marocco, A. defense responses to mycotoxin-producing fungi Fusarium proliferatum, F. subglutinans, and Aspergillus flavus in kernels of susceptible and resistant maize genotypes. Mol. Plant-Micr. Interact. 2015, 28, 546–557. [Google Scholar] [CrossRef] [PubMed]
- Almeida, R.N.; Vivas, M.; dos Santos Junior, D.R.; Saluci, J.C.G.; Cazadini Carlos, M.; Santos, J.S.; Teixeira do Amaral Júnior, A.; Scapim, C.A. Combining abilities analysis for ear rot resistance in popcorn hybrids development. Rev. Ceres Viçosa 2021, 68, 61–70. [Google Scholar] [CrossRef]
- Morales, L.; Zila, C.T.; Moreta Mejía, D.E.; Montoya Arbelaez, M.; Balint-Kurti, P.J.; Holland, J.B.; Nelson, R.J. Diverse Components of Resistance to Fusarium verticillioides Infection and Fumonisin Contamination in Four Maize Recombinant Inbred Families. Toxins 2019, 11, 86. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Ding, J.; Li, H.; Li, Z.; Sun, X.; Li, J.; Wang, R.; Dai, X.; Dong, H.; Song, W.; et al. Detection and verification of quantitative trait loci for resistance to Fusarium ear rot in maize. Mol. Breed. 2012, 30, 1649–1656. [Google Scholar] [CrossRef]
- Agostini, R.B.; Postigo, A.; Rius, S.P.; Rech, G.E.; Campos-Bermudez, V.A.; Vargas, W.A. Long-lasting primed state in maize plants: Salicylic acid and steroid signaling pathways as key players in the early activation of immune responses in silks. Mol. Plant-Microb. Interact. 2019, 32, 95–106. [Google Scholar] [CrossRef] [PubMed]
- Perez-Brito, D.; Jeffers, D.; de Leon, G.; Khairallah, D.; Cortes-Cruz, M.; Velázquez, C.G.; Azpíroz Rivero, S.; Srinivasan, G. QTL mapping of Fusarium moniliforme ear rot resistance in highland maize, Mexico. Agrocencia 2001, 35, 181–196. [Google Scholar]
- Drepper, W.J.; Renfro, B.L. Comparison of methods for inoculation of ears and stalks of maize with Fusarium moniliforme. Plant Dis. 1990, 74, 952–956. [Google Scholar] [CrossRef]
- Santiago, R.; Cao, A.; Malvar, R.A.; Butrón, A. Genomics of maize resistance to Fusarium ear rot and fumonisin contamination. Toxins 2020, 12, 431. [Google Scholar] [CrossRef]
- Coan, M.M.D.; Senhorinho, H.J.C.; Pinto, R.J.B.; Scapim, C.A.; Tessmann, D.J.; Williams, W.P.; Warburton, M.L. Genome-wide association study of resistance to ear rot by Fusarium verticillioides in a tropical field maize and popcorn core collection. Crop Sci. 2018, 58, 564–578. [Google Scholar] [CrossRef]
- Zila, C.T.; Samayoa, L.F.; Santiago, R.; Butrón, A.; Holland, J.B. A genome-wide association study reveals genes associated with Fusarium Ear Rot resistance in a maize core diversity panel. G3 Genes Genomes Genet. 2013, 3, 2014–2095. [Google Scholar] [CrossRef]
- Shin, J.-H.; Kim, J.-E.; Malapi-Wight, M.; Choi, Y.-E.; Shaw, B.D.; Shim, W.-B. Protein phosphatase 2A regulatory subunits perform distinct functional roles in the maize pathogen Fusarium verticillioides. Mol. Plant Path. 2013, 14, 518–529. [Google Scholar] [CrossRef] [PubMed]
- Small, I.M.; Flett, B.C.; Marasas, W.F.O.; McLeod, A.; Viljoen, A. Use of resistance elicitors to reduce Fusarium ear rot and fumonisin accumulation in maize. Crop Prot. 2012, 41, 10–16. [Google Scholar] [CrossRef]
- Mu, C.; Gao, J.; Zhou, Z.; Wang, Z.; Sun, X.; Zhang, X.; Dong, H.; Han, Y.; Li, X.; Wu, Y.; et al. Genetic analysis of cob resistance to F. verticillioides: Another step towards the protection of maize from ear rot. Theor. Appl. Genet. 2019, 132, 1049–1059. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Choi, Y.E.; Zou, X.; Xu, J.R. The FvMK1 mitogen-activated protein kinase gene regulates conidiation, pathogenesis, and fumonisin production in Fusarium verticillioides. Fungal Genet. Biol. 2011, 48, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Czembor, E.; Waskiewicz, A.; Piechota, U.; Puchta, M.; Czembor, J.H.; Stepien, Ł. Differences in ear rot resistance and Fusarium verticillioides—Produced fumonisin contamination between Polish currently and historically used maize inbred lines. Front. Microbiol. 2019, 10, 449. [Google Scholar] [CrossRef]
- Christensen, S.A.; Nemchenko, A.; Park, Y.-S.; Borrego, E.; Huang, P.-C.; Schmelz, E.A.; Kunze, S.; Feussner, I.; Yalpani, N.; Meeley, R.; et al. The novel monocot-specific 9-lipoxygenase zmlox12 is required to mount an effective jasmonate-mediated defense against Fusarium verticillioides in maize. MPMI 2014, 27, 1263–1276. [Google Scholar] [CrossRef]
- Gnonlonfin, G.J.B.; Hell, K.; Adjovi, Y.; Fandohan, P.; Koudande, D.O.; Mensah, G.A.; Sanni, A.; Brimer, L. A review on aflatoxin contamination and its implications in the developing world: A Sub-Saharan African perspective. Crit. Rev. Food Sci. Nutr. 2013, 53, 349–365. [Google Scholar] [CrossRef] [PubMed]
- Womack, E.D.; Williams, W.P.; Windham, G.L.; Xu, W. Mapping quantitative trait loci associated with resistance to aflatoxin accumulation in maize inbred Mp719. Front. Microbiol. 2020, 11, 45. [Google Scholar] [CrossRef]
- Xu, F.; Baker, R.C.; Whitaker, T.B.; Luo, H.; Zhao, Y.; Stevenson, A.; Boesch, C.J.; Zhang, G. Review of good agricultural practices for smallholder maize farmers to minimize aflatoxin contamination. World Mycotoxin J. 2022, 15, 171–186. [Google Scholar] [CrossRef]
- Kumar, P.; Mahato, D.K.; Kamle, M.; Mohanta, T.K.; Kang, S.G. Aflatoxins: A global concern for food safety, human health, and their management. Front. Microbiol. 2017, 7, 2170. [Google Scholar] [CrossRef]
- Available online: https://www.krugerseed.com/en-us/agronomy-library/aflatoxin-in-corn.html (accessed on 12 October 2023).
- Willcox, M.C.; Davis, G.L.; Warburton, M.L.; Windham, G.L.; Abbas, H.K.; Betrán, J.; Holland, J.B.; Williams, W.P. Confirming quantitative trait loci for aflatoxin resistance from Mp313E in different genetic backgrounds. Mol. Breed. 2013, 32, 15–26. [Google Scholar] [CrossRef]
- Wahl, N.; Murray, S.C.; Isakeit, T.; Krakowsky, M.; Windham, G.L.; Williams, W.P.; Guo, B.; Ni, X.; Knoll, J.; Xu, W.; et al. Identification of resistance to aflatoxin accumulation and yield potential in maize hybrids in the Southeast Regional Aflatoxin Trials (SERAT). Crop Sci. 2017, 57, 202–215. [Google Scholar] [CrossRef]
- Abbas, H.K.; Cartwright, R.D.; Xie, W.; Shier, W.T. Aflatoxin and fumonisin contamination of corn (maize, Zea mays) hybrids in Arkansas. Crop Prot. 2006, 25, 1–9. [Google Scholar] [CrossRef]
- Darrah, L.L.; Zuber, M.S. Yield and the Genetic Control of Aflatoxin in Maize. In Aflatoxin in Maize: Proceedings of the Workshop; Zuber, M.S., Lillehoj, E.B., Renfro, B.L., Eds.; CIMMYT: México-Veracruz, El Batan, Mexico, 1987; pp. 236–249. [Google Scholar]
- Chen, Z.-Y.; Rajasekaran, K.; Brown, R.L.; Sayler, R.J.; Bhatnagar, D. Discovery, and confirmation of genes/proteins associated with maize aflatoxin resistance. World Mycotoxin J. 2015, 8, 211–224. [Google Scholar] [CrossRef]
- Baisakh, N.; Da Silva, E.A.; Pradhan, A.K.; Rajasekaran, K. Comprehensive meta-analysis of QTL and gene expression studies identify candidate genes associated with resistance in maize. Front. Plant Sci. 2023, 14, 1214907. [Google Scholar] [CrossRef]
- Warburton, M.L.; Jeffers, D.; Smith, J.S.; Scapim, C.; Uhdre, R.; Thrash, A.; Williams, W.P. Comparative analysis of multiple GWAS results identifies metabolic pathways associated with resistance to A. flavus infection and aflatoxin accumulation in maize. Toxins 2022, 14, 738. [Google Scholar] [CrossRef] [PubMed]
- Paul, C.; Naidoo, G.; Forbes, A.; Mikkilineni, V.; White, D.; Rocheford, T. Quantitative trait loci for low aflatoxin production in two related maize populations. Theor. Appl. Genet. 2003, 107, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Williams, W.P.; Krakowsky, M.D.; Scully, B.T.; Brown, R.L.; Menkir, A.; Warburton, M.L.; Windham, G.L. Identifying and developing maize germplasm with resistance to accumulation of aflatoxins. World Mycotoxin J. 2015, 8, 193–209. [Google Scholar] [CrossRef]
- Williams, W.P. Breeding for resistance to aflatoxin accumulation in maize. Mycotoxin Res. 2006, 22, 27–32. [Google Scholar] [CrossRef]
- Ojiambo, P.S.; Battilani, P.; Jeffrey, W.; Cary, J.F.; Burt, H.; Blum, B.H.; Carbone, I. Cultural and genetic approaches to manage aflatoxin contamination: Recent insights provide opportunities for improved control. Phytopathology 2018, 108, 1024–1037. [Google Scholar] [CrossRef]
- Bertagna, F.A.B.; Kuki, M.C.; Neto, H.Z.; Tessmann, D.J.; Pinto, R.J.B.; Scapim, C.A.; Williams, W.P.; Warburton, M. Association mapping and pathway analysis of ear rot disease caused by Aspergillus flavus in a panel of tropical maize germplasm. Crop Sci. 2021, 61, 4128–4138. [Google Scholar] [CrossRef]
- Hawkins, L.K.; Mylroie, J.E.; Oliveira, D.A.; Smith, J.S.; Ozkan, S.; Windham, G.L.; Williams, W.P.; Warburton, M.L. Characterization of the maize chitinase genes and their effect on Aspergillus flavus and aflatoxin accumulation resistance. PLoS ONE 2015, 10, e0126185. [Google Scholar] [CrossRef]
- Warburton, M.L.; Brooks, T.D.; Krakowsky, M.D.; Shan, X.; Windham, G.L.; Williams, W.P. Identification, and mapping of new sources of resistance to aflatoxin accumulation in maize. Crop Sci. 2009, 49, 1403–1408. [Google Scholar] [CrossRef]
- Musungu, B.M.; Bhatnagar, D.; Brown, R.L.; Payne, G.A.; O’Brian, G.; Fakhoury, A.M.; Geisler, M.A. Network approach of gene co-expression in the Zea mays/Aspergillus flavus pathosystem to map host/pathogen interaction pathways. Front. Genet. 2016, 7, 206. [Google Scholar] [CrossRef]
- Omolehin, O.; Raruang, Y.; Hu, D.; Han, Z.-Q.; Wei, Q.; Wang, K.; Rajasekaran, K.; Cary, J.W.; Chen, Z.-Y. Resistance to aflatoxin accumulation in maize mediated by host-induced silencing of the Aspergillus flavus alkaline protease (alk) gene. J. Fungi 2021, 7, 904. [Google Scholar] [CrossRef] [PubMed]
- Thakare, D.; Zhang, J.; Wing, R.A.; Cotty, P.J.; Schmidt, M.A. Aflatoxin-free transgenic maize using host-induced gene silencing. Sci. Adv. 2017, 3, e1602382. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.-Y.; Brown, R.L.; Damann, K.E.; Cleveland, T.E. Identification of maize kernel endosperm proteins associated with resistance to aflatoxin contamination by Aspergillus flavus. Phytopathology 2007, 97, 1094–1103. [Google Scholar] [CrossRef] [PubMed]
- Scott, G.E.; Zummo, N. Host-Plant Resistance: Screening Techniques. In Aflatoxin in Maize: Proceedings of the Workshop; Zuber, M.S., Lillehoj, E.B., Renfro, B.L., Eds.; CIMMYT: México-Veracruz, El Batan, Mexico, 1987; pp. 221–233. [Google Scholar]
- King, S.B.; Scott, G.E. Screening maize single crosses for resistance to preharvest infection of kernels by Aspergillus flavus. Phytopathology 1982, 72, 942. [Google Scholar]
- Kelley, R.Y.; Williams, W.P.; Mylroie, J.E.; Boykin, D.L.; Harper, J.W.; Windham, G.L.; Ankala, A.; Shan, X. Identification of maize genes associated with host plant resistance or susceptibility to Aspergillus flavus infection and aflatoxin accumulation. PLoS ONE 2012, 7, e36892. [Google Scholar] [CrossRef]
- Miedaner, T.; Löffler, M.; Bolduan, C.; Kessel, B.; Ouzunova, M.; Mirdita, V.; Melchinger, A.E. Genetic variation for resistance and mycotoxin content of European maize inoculated with Fusarium graminearum and F. verticillioides. Cereal Res. Commun. 2008, 36 (Suppl. B), 45–48. [Google Scholar] [CrossRef]
- Ge, C.-X.; Tang, C.-X.; Zhu, Y.-X.; Wang, G.-F. Genome-wide identification of the maize 2OGD superfamily genes and their response to Fusarium verticillioides and Fusarium graminearum. Gene 2021, 764, 145078. [Google Scholar] [CrossRef] [PubMed]
- Akohoue, F.; Miedaner, T. Meta-analysis and co-expression analysis revealed stable QTL and candidate genes conferring resistances to Fusarium and Gibberella ear rots while reducing mycotoxin contamination in maize. Front. Plant Sci. 2022, 13, 1050891. [Google Scholar] [CrossRef] [PubMed]
- Lanubile, A.; Pasini, L.; Lo Pinto, M.; Battilani, P.; Prandini, A.; Marocco, A. Evaluation of broad spectrum a.sources of resistance to Fusarium verticillioides and advanced maize breeding lines. World Mycotox. J. 2013, 4, 43–51. [Google Scholar] [CrossRef]
- Presello, D.A.; Pereyra, A.O.; Iglesias, J.; Faugue, C.M.; Sampietro, D.A.; Eyherabide, G.H. Responses to selection of S5 inbreds for broad-based resistance to ear rots and grain mycotoxin contamination caused by Fusarium spp. in maize. Euphytica 2011, 178, 23–29. [Google Scholar] [CrossRef]
- Mesterházy, Á.; Kovács, K. Breeding corn against fusarial stalk rot, ear rot and seedling blight. Acta Phytopath. Acad. Sci. Hung. 1986, 21, 231–249. [Google Scholar]
- Akello, J.; Ortega-Beltran, A.; Katati, B.; Atehnkeng, J.; Augusto, J.; Mwila, C.M.; Mahuku, G.; Chikoye, D.; Bandyopadhyay, R. Prevalence of aflatoxin- and fumonisin-producing fungi associated with cereal crops grown in Zimbabwe, and their associated risks in a climate change scenario. Foods 2021, 10, 287. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA); Maggiore, A.; Afonso, A.; Barrucci, F.; De Sanctis, G. Climate change as a driver of emerging risks for food and feed safety, plant, animal health and nutritional quality. EFSA Support. Publ. 2020, 17, 1881E. [Google Scholar] [CrossRef]
- Zanon, M.S.A.; Pena, G.; ·Yerkovich, N.; ·Bossa, M.; ·Chiotta, M.L.; Chulze, S.N. Aflatoxins and fumonisins in maize under a climate change scenario. Biocontrol strategies at the pre-harvest stage. Eur. J. Plant Pathol. 2023, 167, 551–567. [Google Scholar] [CrossRef]
- Yu, J.; Hennessy, D.A.; Tack, J.; Wu, F. Climate change will increase aflatoxin presence in US Corn. Environ. Res. Lett. 2022, 17, 054017. [Google Scholar] [CrossRef]
- Van der Fels-Klerx, H.J.; Vermeulen, L.C.; Gavai, A.K.; Liu, C. Climate change impacts on aflatoxin B1 in maize and aflatoxin M1 in milk: A case study of maize grown in Eastern Europe and imported to the Netherlands. PLoS ONE 2019, 14, e0218956. [Google Scholar] [CrossRef]
- Palumbo, R.; Gonçalves, A.; Gkrillas, A.; Logrieco, A.; Dorne, J.-L.; Dall’Asta, C.; Venâncio, A.; Battilani, P. Mycotoxins in maize: Mitigation actions, with a chain management approach. Phytopath. Mediterr. 2020, 59, 5–28. [Google Scholar] [CrossRef]
- Kawasaki, K. Two harvests are better than one: Double cropping as a strategy for climate change adaptation Am. J. Agric. Econ. 2018, 101, 172–192. [Google Scholar] [CrossRef]
- Cui, X.-M.; Xie, W. Adapting Agriculture to climate change through growing season adjustments: Evidence from corn in China. Amer. J. Agron Econ. 2023, 105, 249–272. [Google Scholar] [CrossRef]
- Mesterházy, Á. Resistance of corn to Fusarium ear rot and its relation to seedling resistance. Phytopath. Z. 1982, 103, 218–231. [Google Scholar] [CrossRef]
- Young, H.C., Jr. The toothpick method of inoculating corn for ear and stalk rots. Phytopathology 1943, 33, 16. [Google Scholar]
- Reid, L.M.; Nicol, R.W.; Ouellet, T.; Savard, M.; Miller, J.D.; Young, J.C.; Stewart, D.W.; Schaafsma, A.W. Interaction of Fusarium graminearum and F. moniliforme in maize ears: Disease progress, fungal biomass, and mycotoxin accumulation. Phytopathology 1999, 89, 1028–1037. [Google Scholar] [CrossRef] [PubMed]
- Reid, L.M.; Zhu, X. Screening Corn for Resistance to Common Diseases in Canada 2005; Agriculture and Agri-Food Canada: Burnaby, BC, Canada, 2005; 26p, ISBN 0-662-40347-9. [Google Scholar]
- Munkvold, G.P.; McGee, D.C.; Carleton, W.M. Importance of different pathways for maize kernel infection by Fusarium moniliforme. Phytopathology 1997, 87, 209–217. [Google Scholar] [CrossRef]
- Presello, D.A.; Reid, L.M.; Butler, G.; Mather, D.E. Pedigree selection for Gibberella ear rot resistance in maize populations. Euphytica 2005, 143, 1–8. [Google Scholar] [CrossRef]
- Papst, C.; Zellner, J.; Venkataratnam, S.; Eder, J. Fusarium—Problematik bei Kornermais (Zea mays L.). (Fusarium problems in grain maize). Gesunde Pflanzen 2007, 59, 7–16. [Google Scholar] [CrossRef]
- Poulsen, H.M.; Ridao, A.D.C.; Castaño, F. Comparación Entre Técnicas de Inoculación de Fusarium verticillioides en Espigas de Maíz. RIA/Trabajos en Prensa, 2013, RIA/1-7. Available online: https://www.researchgate.net/publication/260837506 (accessed on 16 October 2021).
- Oswald, I.P.; Seefelder, W.; Speijers, G.; Stroka, J. Masked mycotoxins: A review. Mol. Nutr. Food Res. 2013, 57, 165–186. [Google Scholar] [CrossRef]
- Zhang, Z.; Nie, D.; Fan, K.; Yang, J.; Guo, W.; Meng, J.; Zhao, Z.; Han, Z. A systematic review of plant-conjugated masked mycotoxins: Occurrence, toxicology, and metabolism. Crit. Rev. Food Sci. Nutr. 2020, 60, 1523–1537. [Google Scholar] [CrossRef] [PubMed]
- Mesterhazy, A.; Varga, M.; Tóth, B.; Kótai, C.; Bartók, T.; Véha, A.; Ács, K.; Vágvölgyi, C.; Lehoczki-Krsjak, S. Reduction of deoxynivalenol (DON) contamination by improved fungicide use in wheat. Part 2. Farm scale tests with different nozzle types and updating the integrated approach. Eur. J. Plant Pathol. 2018, 151, 1–20. [Google Scholar] [CrossRef]
- Mesterhazy, A. Chemical control of Fusarium head blight of wheat, a comprehensive view. In Mycotoxin Reduction in Grain Chains: A Practical Guide; Logrieco, A.F., Visconti, A., Eds.; Blackwell-Wiley: Ames, IA, USA; Chichester, UK; Oxford UK, 2014; pp. 232–247, 352. [Google Scholar]
- Mesterhazy, A.; Szieberth, D.; Szabó, B.; Berényi, A.; Tóth, B. Mycotoxin contamination of maize (Zea mays L.) samples in Hungary, 2012–2017. Cereal Res. Comm. 2022, 50, 1055–1063. [Google Scholar] [CrossRef]
- Gaikpa, D.S.; Miedaner, T. Genomics-assisted breeding for ear rot resistances and reduced mycotoxin contamination in maize: Methods, advances, and prospects. Theor. Appl. Genet. 2019, 132, 2721–2739. [Google Scholar] [CrossRef]
- Leggieri, M.C.; Toscano, P.; Battilani, P. Predicted Aflatoxin B1 Increase in Europe due to climate change: Actions and reactions at global level. Toxins 2021, 13, 292. [Google Scholar] [CrossRef]
- Medina, A.; Rodriguez, A.; Magan, N. Effect of climate change on Aspergillus flavus and aflatoxin B1 production. Front. Microbiol. 2014, 5, 348. [Google Scholar] [CrossRef]
- Pandey, A.K.; Samota, M.K.; Kumar, A.; Silva, A.S.; Dubey, N.K. Fungal mycotoxins in food commodities: Present status and future concerns. Front. Sustain. Food Syst. 2023, 7, 1162595. [Google Scholar] [CrossRef]



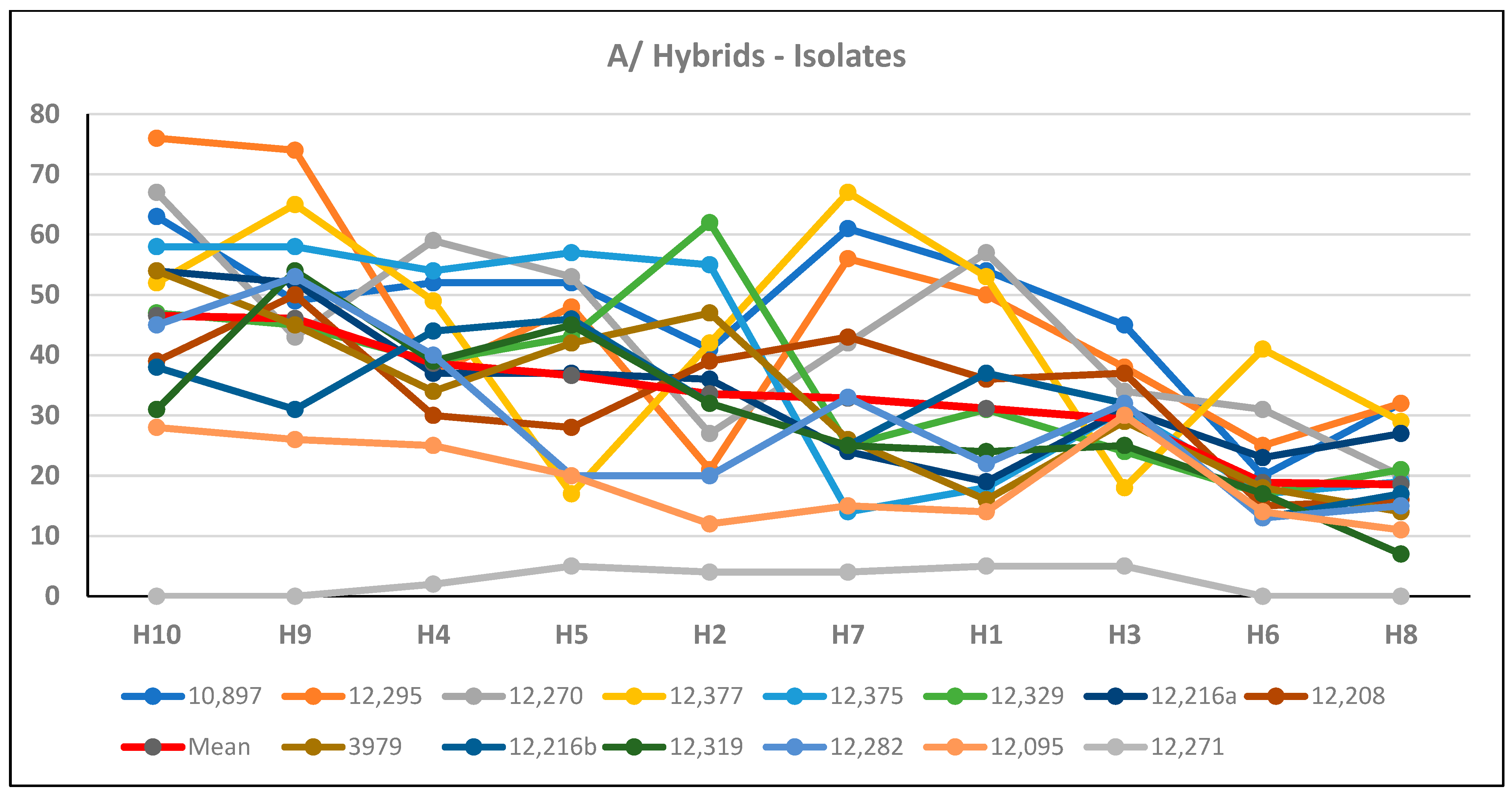{kind=link}
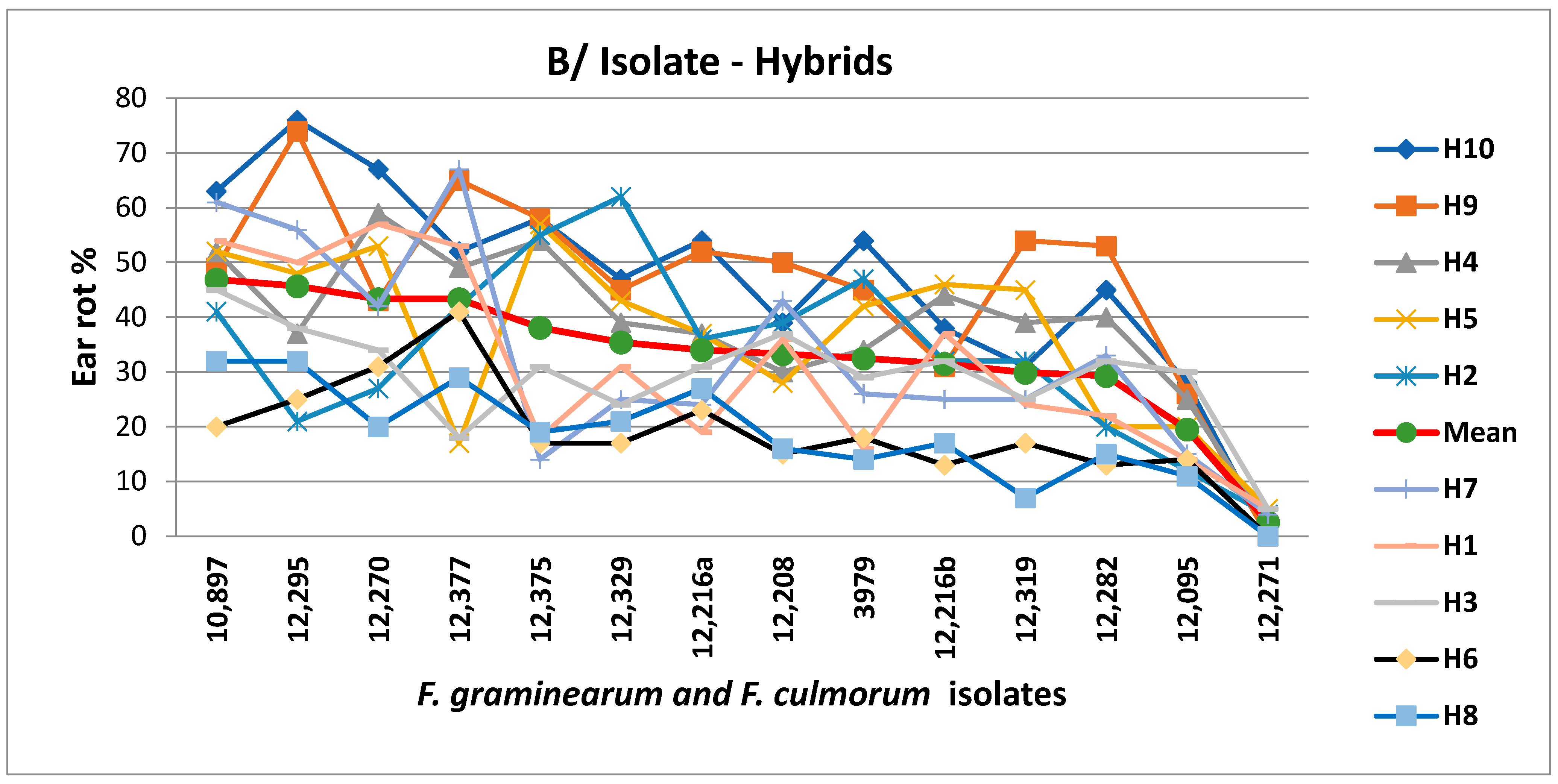{kind=link}
| Authors | Ref. No. | Pathogen | Inoculation | No. of Isolates/mixture | Conidium conc./mL | Aggressiveness | Evaluation | Severity% min–max | DON mg/kg | FUM mg/kg | AFB1 mg/kg |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Afolabi et al., 2007 | [14] | F. vert. | Art. silk | 1 | 1 × 106 | No | 2.3–57 | 1–53 | |||
| Araujo et al., 2022 | [15] | F. vert. | Silk + kernel | 1 | 5 × 105 | No | Scale 1–9 | 2.5–6.5 | No | ||
| Bernardi et al., 2018 | [16] | F. vert | Art. kernel | 1 | 1 × 106 | No | Not tested | No | No | ||
| Bolduan et al., 2009 | [17] | F. vert. | Art. silk | 1 | 1 × 106 | No | % | 1.6–39.5 | 0.05–58 | ||
| Butron et al., 2019 | [18] | F. vert | Art. silk | 1 | 1 × 106 | No | Scale 1–7 | No | |||
| Butron et al., 2019 | [18] | F. vert | Art. kernel | 1 | No data | No data | Scale 1–7 | No | No | ||
| Cao et al., 2014 | [19] | F. vert. | Natural | - | - | - | % disinfect. | 4.6–35.3 | No | ||
| Cao et al., 2014 | [19] | F. vert. | Natural | - | - | - | %, no disinfect. | 67–84 | No | ||
| Cao et al., 2014 | [20] | F. vert | Art. silk | 1 | 2.5 × 105 | No | Scale 1–9 | No | 4.1–5.9 | ||
| Cao et al., 2014 | [20] | F. vert | Art. kernel | 1 | 2.5 × 106 | No | Scale 1–9 | No | 2.6–9.6 | ||
| Cao et al., 2022 | [21] | F. vert. | Art. kernel | 1 | 1 × 106 | No | Scale 1–8 | No | No | ||
| Chen et al., 2016 | [22] | F. vert. | Art. | 1 | 5 × 106 | No | Scale 1–7 | 0–74 | No | ||
| Chiuraise et al., 2016 | [23] | F. vert. | No data | No data | No data | No data | Scale 1–7 | 2–25 | 22–48 | ||
| Gesterio et al., 2021 | [24] | F. vert | Not given | No | 1 × 106 | No | No | 0.02–0.2 | |||
| Guche et al., 2022 | [25] | F. vert | Art. | 1 | 1 × 106 | No | 3.5–50 | 2.37–179 ppb | |||
| Guo et al., 2016 | [26] | F. vert. | Natural | No data | Not appl. | No | 0.3–5 | ||||
| Henry et al., 2009 | [27] | F. vert | Art. kernel | 1 | 9 × 107 | No | cm2 | 1–9 | 2–48 | ||
| Ivic et al., 2008 | [28] | F. vert. | Art. kernel | 7 | 3 × 106 | No | Scale 1–7 | 2.29–5.76 | No | ||
| Lanubile et al., 2014 | [29] | F. vert. | Art. | 1 | 3.5 × 106 | No | No data | No | |||
| Lanubile et al., 2014 | [30] | F. vert. | Art. kernel | 1 | 3.5 × 106 | Tested | No | No | |||
| Lanzanova | [31] | F. vert. | 2 | 1 × 106 | No | No inf. Grains | 4.7–97/cob | 4.4–141 | |||
| Loeffler et al., 2010 | [32] | F. vert. | Art. silk | 1 | 1 ×105 | No | % | No | |||
| Santiago et al., 2013 | [33] | F. vert | Art. kernel | 1, 2 mL | 1 × 106 | Old data | Scale 1–7 | 1.3–6.04 | 0.1–1.5 | ||
| Schjoth et al. | [34] | F. vert. | Art. silk | 6 | 1 × 106 | FB content | Scale 1–7 | No | 60–1083 ppb | ||
| Wu et al., 2020 | [35] | F. vert | Art. silk | 1 | 1 × 106 | No | Scale 1–7 | No | |||
| Zila et al., 2014 | [36] | F. vert | Art. silk | Not given | 2 × 106 | No | No | ||||
| Abbas et al., 2012 | [37] | A. flavus | Art. kernel | 1 × 106 | No | No data | No | 1–3008 | |||
| Abbas et al., 2012 | [37] | A. flavus | Natural | Not appl. | Not appl. | Not appl. | No data | No | 0–289 | ||
| Chiuraise et al., 2016 | [23] | A. flavus | No data | No data | No data | No | Scale 1–7 | 1.5–40 | 1.9–32 | ||
| Guche et al., 2022 | [25] | A. flavus | Art. | 1 | 2 × 106 | No | 5–71 | 12–30 × 103 | |||
| Guo et al., 2016 | [26] | A. flavus | Silk | 1 | 4 × 106 | No | 52–1524 | ||||
| Henry et al., 2009 | [27] | A. flavus | Art. kernel | 1 | 3 × 108 | No | cm2 | 1–9 | 18–10,592, mg/kg * | ||
| Henry et al., 2012 | [38] | A. flavus | Art. kernel | 1 | 3 × 106 | No | 0–9 | 0.3–7.3 | 0–776 | ||
| Williams et al., 2014 | [39] | A. flavus | Art. kernel | 1 | 3 × 108 | No | No | No | 0–119 | ||
| Ali et al. 2005 | [40] | F. gram | Art. kernel | No | 2.5 × 106 | No | Scale 1–7 | No | No | ||
| Ali et al., 2005 | [40] | F. gram. | Art. silk | No | 2.5 × 106 | No | Scale 1–7 | No | No | ||
| Bolduan et al., 2009 | [17] | F. gram | Art. silk | 1, IFA66 | 1 ×105 | No | % | 22–100 | 0–3072 | ||
| Gaikpa et al., 2021 | [41] | F. gram. | Art. silk | 1 | 1.5 × 104 | No | % | 0–90 | No | ||
| Galiano-Carnerio et al., 2021 | [42] | F. gram | Art. silk Br | 3 | 5 × 104 | No | n.d. | No | |||
| Galiano-Carnerio et al., 2021 | [42] | F. gram | Art. silk De | 1 | 1.5 × 104 | No | % | 0–68 | No | ||
| Kebede et al., 2016 | [43] | F. gram | Art. silk | 2 | No | No | % | 8–92 | No | ||
| Kebede et al., 2016 | [43] | F. gram | Art. kernel | 2 | No | No | % | 12–82 | No | ||
| Loeffler et al., 2010 | [32] | F. gram | Art. silk | 1 | No | No | % | 28–83 | 49–1009 | ||
| Reid et al., 1996 | [44] | F. gram. | Art. silk | No | Unknown | No | Scale 1–7 | No | Yes | ||
| Reid, Hamilton 1996 | [45] | F. gram | Art. kernel | No | 0–2 × 106 | No | Scale 1–7 | No | No | ||
| Zhou et al., 2021 | [46] | F. gram. | Art. kernel | 1 | 1 × 106 | No | Scale 1–7 | 2.2–8 | No | ||
| Schaafsma et al., 1997 | [47] | F. gram. | Silk | 1 | 1 × 105 | No | Scale 1–7 | S–MR | 1.4–4 | ||
| Schaafsma et al., 1997 | [47] | F. gram | PIN | 1 | 1 × 105 | No | Scale 1–7 | S–MR | 2–4.6 | ||
| Schaafsma et al., 1993 | [48] | Fg, Fv, Fm | Silk | 2 + 2 + 21 | No data | No | Scale 1–7 | 1.7–4.6 | 1–15.4 | 0–11.5 | |
| Item | Yield MMT | Y * Total % | Y ** Harv. % |
|---|---|---|---|
| Total capacity | 1590 | 100 | |
| Total harvested | 1060 | 66.7 | 100 |
| Preharvest loss (biotic, abiotic, 3% for harvest) 33% | 530 | 33.3 | - |
| Mycotoxin contamination at harvest | 106 | 6.7 | 10 |
| Storage waste | 212 | 13.3 | 20 |
| Consumer and other waste | 138 | 8.7 | 13 |
| Total waste | 986 | 62.0 | 43 |
| Total grain used | 604 | 38.0 | 57 |
| Fusarium spp. | Mycotoxins a |
|---|---|
| F. acuminatum | T2, MON, HT2, DAS, MAS, NEO, BEA |
| F. anthophilum | BEA |
| F. avenaceum | MON, BEA |
| F. cerealis | NIV, FUS, ZEN, ZOH |
| F. chlamydosporum | MON |
| F. culmorum | DON, ZEN, NIV, FUS, ZOH, AcDONs |
| F. equiseti | ZEN, ZOH, MAS, DAS, NIV, DAcNIV, FUS, FUC, BEA |
| F. graminearum | DON, ZEN, NIV, FUS, AcDONs, DAcDON, DAcNIV |
| F. heterosporum | ZEN, ZOH |
| F. nygamai | BEA, FB1, FB2 |
| F. oxysporum | MON, BEA |
| F. poae | DAS, NIV, FUS, MAS, T2, HT2, NEO, BEA |
| F. proliferatum | FB1, BEA, MON, FUP, FB2, |
| F. sambucinum | DAS, T2, NEO, ZEN, MAS, BEA |
| F. semitectum | ZEN, BEA |
| F. sporotrichioides | T2, HT2, NEO, MAS, DAS |
| F. subglutinans | BEA, MON, FUP |
| F. tricinctum | MON, BEA |
| F. verticillioides | FB1, FB2, FB3 |
| Hybrid | Rates between Toxin (mg/kg)/Ear Rot% | Mean | Variance | ||
|---|---|---|---|---|---|
| DON/GER% | FB1/FER% | AFB1/AER% | |||
| Sy Talisman * | ** 0.554 | 4.500 | 0.151 | 1.74 | 5.77 |
| Armagnac | 1.277 | 3.783 | 0.367 | 1.81 | 3.13 |
| SY Zephir | 0.837 | 4.730 | 0.263 | 1.94 | 5.91 |
| Konfites | 0.605 | 5.612 | 0.413 | 2.21 | 8.69 |
| DKC 4541 | 1.983 | 2.484 | 2.967 | 2.48 | 0.24 |
| Konfites | 1.788 | 5.721 | 0.154 | 2.55 | 8.19 |
| Illango | 2.345 | 4.943 | 0.622 | 2.64 | 4.73 |
| Kleopatras | 2.119 | 4.114 | 2.552 | 2.93 | 1.10 |
| P9415 | 1.534 | 2.300 | 5.573 | 3.14 | 4.60 |
| P9718E | 4.621 | 3.102 | 3.100 | 3.61 | 0.77 |
| Koregraf | 1.848 | 5.881 | 3.733 | 3.82 | 4.07 |
| P0725 | 1.796 | 9.156 | 0.557 | 3.84 | 21.61 |
| ES Lagoon | 1.967 | 9.719 | 0.891 | 4.19 | 23.20 |
| DKC 5830 | 2.753 | 6.250 | 4.100 | 4.37 | 3.11 |
| ES Harmonium | 3.272 | 9.130 | 0.842 | 4.41 | 18.15 |
| Sy Zoan | 2.725 | 14.170 | 0.322 | 5.74 | 54.76 |
| Valkür | 2.871 | 15.500 | 0.136 | 6.17 | 67.17 |
| Korimbos | 5.302 | 40.500 | 7.863 | 17.89 | 385.11 |
| Mean | 2.23 | 8.42 | 1.92 | 4.19 | 34.46 |
| Max/Min rate | 8.69 | 17.61 | 57.93 | ||
| Hybrid | Ear Rot Severity | Toxin Content | ER Severity | Toxin Content Check | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Fg % | Fv % | Afl % | DON mg/kg | FUM B1+B2 mg/kg | Afla ppb | F. Check | Asp. Check | DON mg/kg | FUM B1+B2 mg/kg | Afla ppb | FAO No. | |
| Konfites | 10.15 | 0.61 | 0.35 | 18.15 | 3.49 | 53 | 0.21 | 0.000 | 1.70 | 1.88 | 2 | 430 |
| ES Harmonium | 11.49 | 0.23 | 0.15 | 37.60 | 2.10 | 62 | 0.25 | 0.000 | 0.13 | 0.81 | 4 | 380 |
| Sy Talisman | 13.50 | 0.40 | 0.27 | 7.48 | 1.80 | 71 | 0.27 | 0.000 | 0.72 | 0.54 | 2 | 250 |
| Korimbos | 14.35 | 0.14 | 0.12 | 76.08 | 5.67 | 44 | 0.08 | 0.000 | 3.49 | 0.30 | 408 | 575 |
| P0725 | 15.91 | 0.45 | 0.11 | 28.58 | 4.12 | 613 | 0.17 | 0.005 | 0.00 | 0.12 | 794 | 560 |
| Koregraf | 16.62 | 0.42 | 0.24 | 30.72 | 2.47 | 37 | 0.10 | 0.000 | 0.00 | 0.30 | 352 | 410 |
| DKC 5830 | 17.08 | 0.52 | 0.44 | 47.02 | 3.25 | 245 | 0.17 | 0.000 | 2.18 | 5.63 | 0 | 560 |
| ES Lagoon | 19.76 | 0.32 | 0.24 | 38.87 | 3.11 | 896 | 0.19 | 0.005 | 0.00 | 2.23 | 20 | 460 |
| Armagnac | 19.79 | 0.23 | 0.11 | 25.27 | 0.87 | 98 | 0.13 | 0.005 | 2.33 | 1.58 | 9 | 490 |
| Illango | 19.94 | 0.53 | 0.36 | 46.75 | 2.62 | 1068 | 0.18 | 0.008 | 0.05 | 0.75 | 1143 | 530 |
| P9718E | 21.86 | 0.49 | 0.25 | 101.01 | 1.52 | 638 | 0.09 | 0.000 | 0.17 | 1.17 | 3 | 390 |
| Kathedralis | 22.10 | 0.67 | 0.58 | 13.38 | 3.76 | 361 | 0.21 | 0.000 | 0.27 | 1.73 | 0 | 490 |
| DKC 4541 | 24.59 | 1.61 | 0.54 | 48.77 | 4.00 | 174 | 0.30 | 0.005 | 3.59 | 3.06 | 0 | 370 |
| Valkür | 28.78 | 0.10 | 0.05 | 82.64 | 1.55 | 205 | 0.05 | 0.000 | 1.28 | 0.81 | 0 | 731 |
| P9415 | 32.04 | 0.60 | 0.28 | 49.14 | 1.38 | 38 | 0.24 | 0.000 | 0.58 | 2.32 | 0 | 350 |
| SY Zephir | 33.92 | 0.37 | 0.19 | 28.38 | 1.75 | 160 | 0.16 | 0.005 | 0.63 | 2.36 | 4 | 390 |
| Kleopatras | 35.02 | 0.44 | 0.10 | 74.22 | 1.81 | 310 | 0.16 | 0.000 | 0.35 | 2.31 | 114 | 630 |
| Sy Zoan | 35.36 | 0.47 | 0.16 | 96.35 | 6.66 | 1258 | 0.18 | 0.008 | 0.00 | 1.65 | 0 | 560 |
| Mean | 21.79 | 0.48 | 0.25 | 47.24 | 2.88 | 352 | 0.17 | 0.002 | 0.97 | 1.64 | 159 | |
| LSD 5% | 8.60 | 0.55 | 0.15 | 0.08 | ||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mesterhazy, A. Food Safety Aspects of Breeding Maize to Multi-Resistance against the Major (Fusarium graminearum, F. verticillioides, Aspergillus flavus) and Minor Toxigenic Fungi (Fusarium spp.) as Well as to Toxin Accumulation, Trends, and Solutions—A Review. J. Fungi 2024, 10, 40. https://doi.org/10.3390/jof10010040
Mesterhazy A. Food Safety Aspects of Breeding Maize to Multi-Resistance against the Major (Fusarium graminearum, F. verticillioides, Aspergillus flavus) and Minor Toxigenic Fungi (Fusarium spp.) as Well as to Toxin Accumulation, Trends, and Solutions—A Review. Journal of Fungi. 2024; 10(1):40. https://doi.org/10.3390/jof10010040
Chicago/Turabian StyleMesterhazy, Akos. 2024. "Food Safety Aspects of Breeding Maize to Multi-Resistance against the Major (Fusarium graminearum, F. verticillioides, Aspergillus flavus) and Minor Toxigenic Fungi (Fusarium spp.) as Well as to Toxin Accumulation, Trends, and Solutions—A Review" Journal of Fungi 10, no. 1: 40. https://doi.org/10.3390/jof10010040
APA StyleMesterhazy, A. (2024). Food Safety Aspects of Breeding Maize to Multi-Resistance against the Major (Fusarium graminearum, F. verticillioides, Aspergillus flavus) and Minor Toxigenic Fungi (Fusarium spp.) as Well as to Toxin Accumulation, Trends, and Solutions—A Review. Journal of Fungi, 10(1), 40. https://doi.org/10.3390/jof10010040