
Individual Contributions of Cardiac Ion Channels on Atrial Repolarization and Reentrant Waves: A Multiscale In-Silico Study


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Zero-Dimensional Computational Modeling
2.2. Two-Dimensional Computational Modeling
3. Results
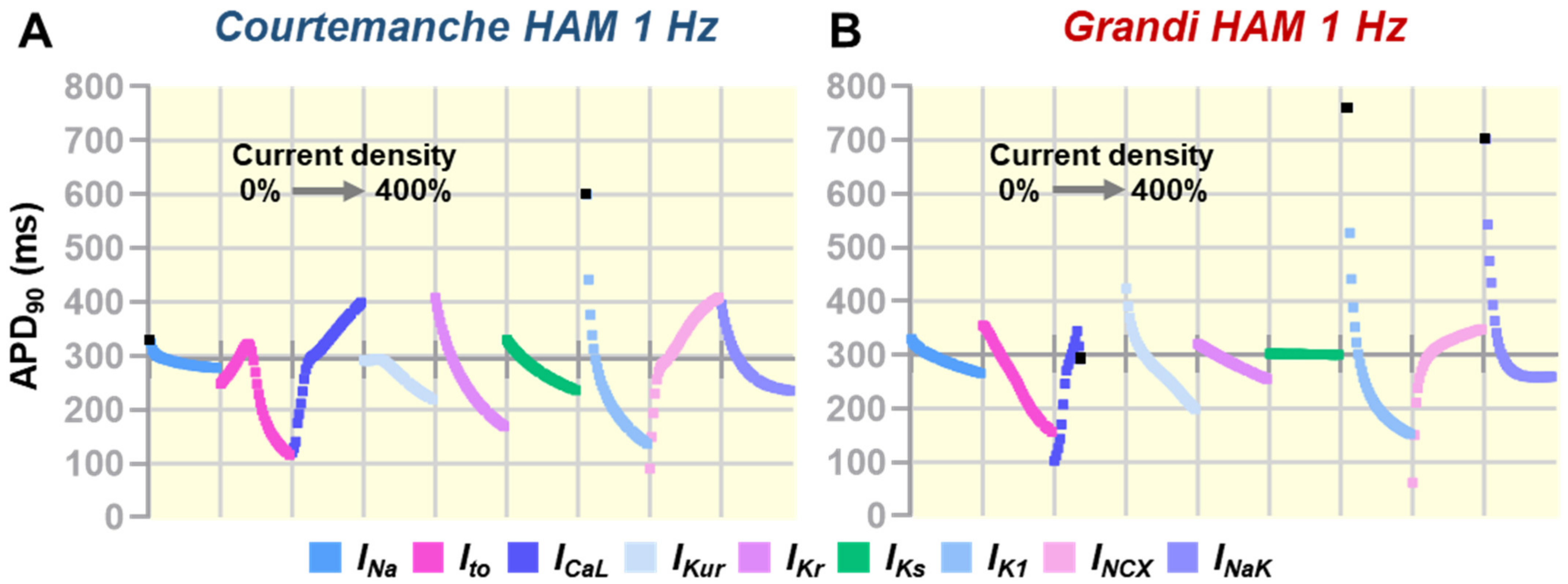
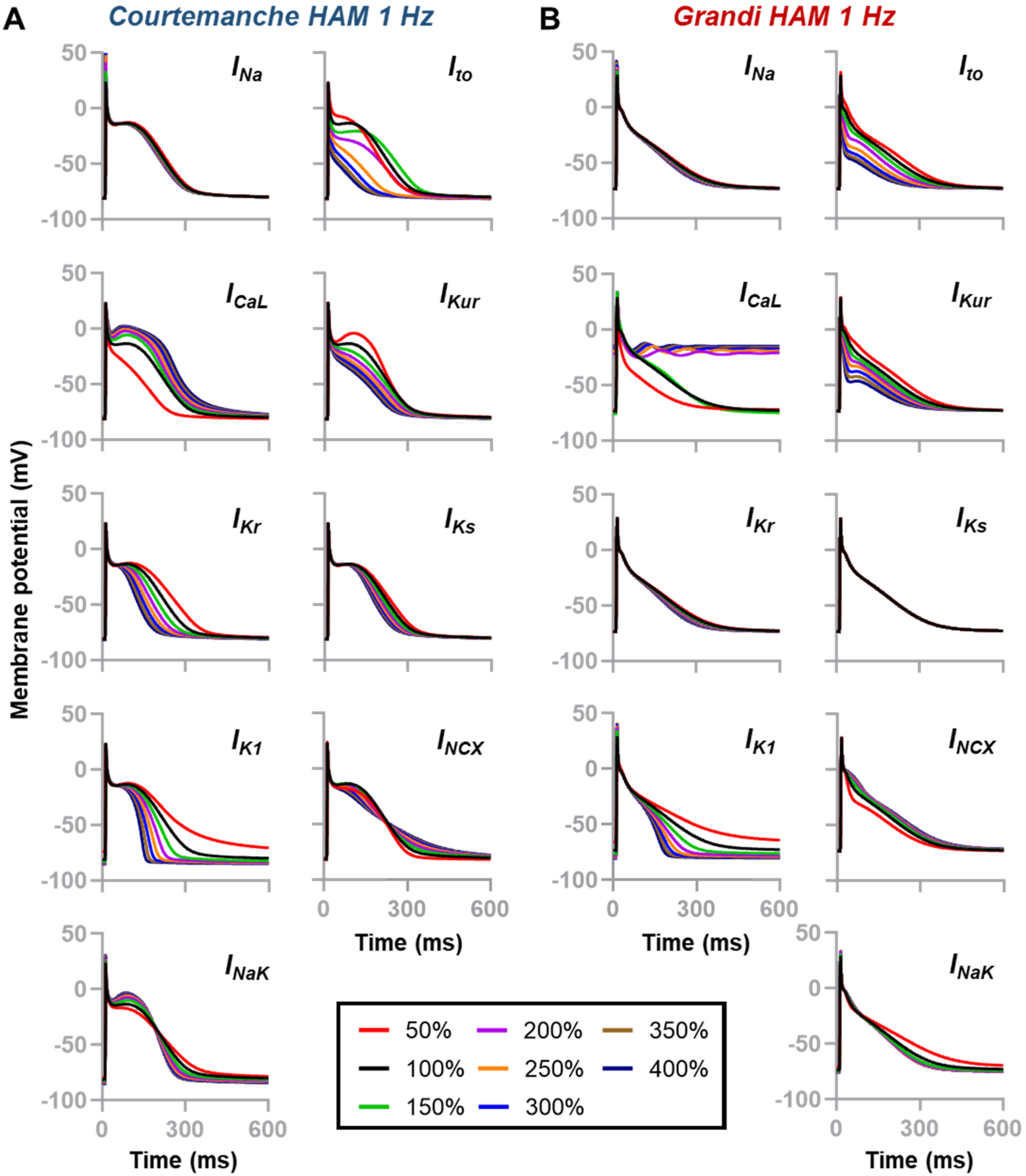
3.1. Individual Contribution of Ion Channels on Atrial Action Potential Repolarization
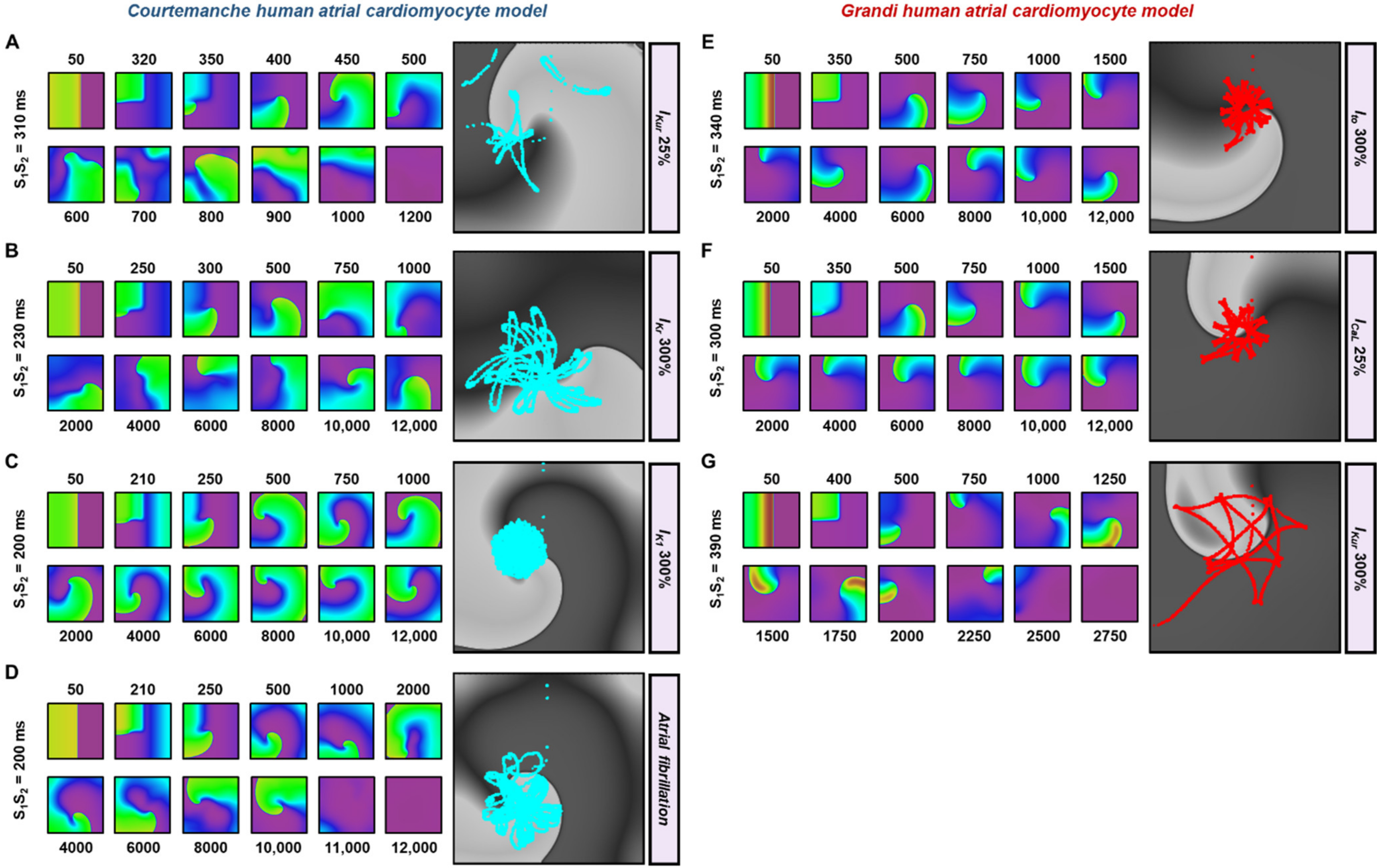
3.2. The Effects of Ionic Current Conductance Perturbation on Reentrant Spiral Waves
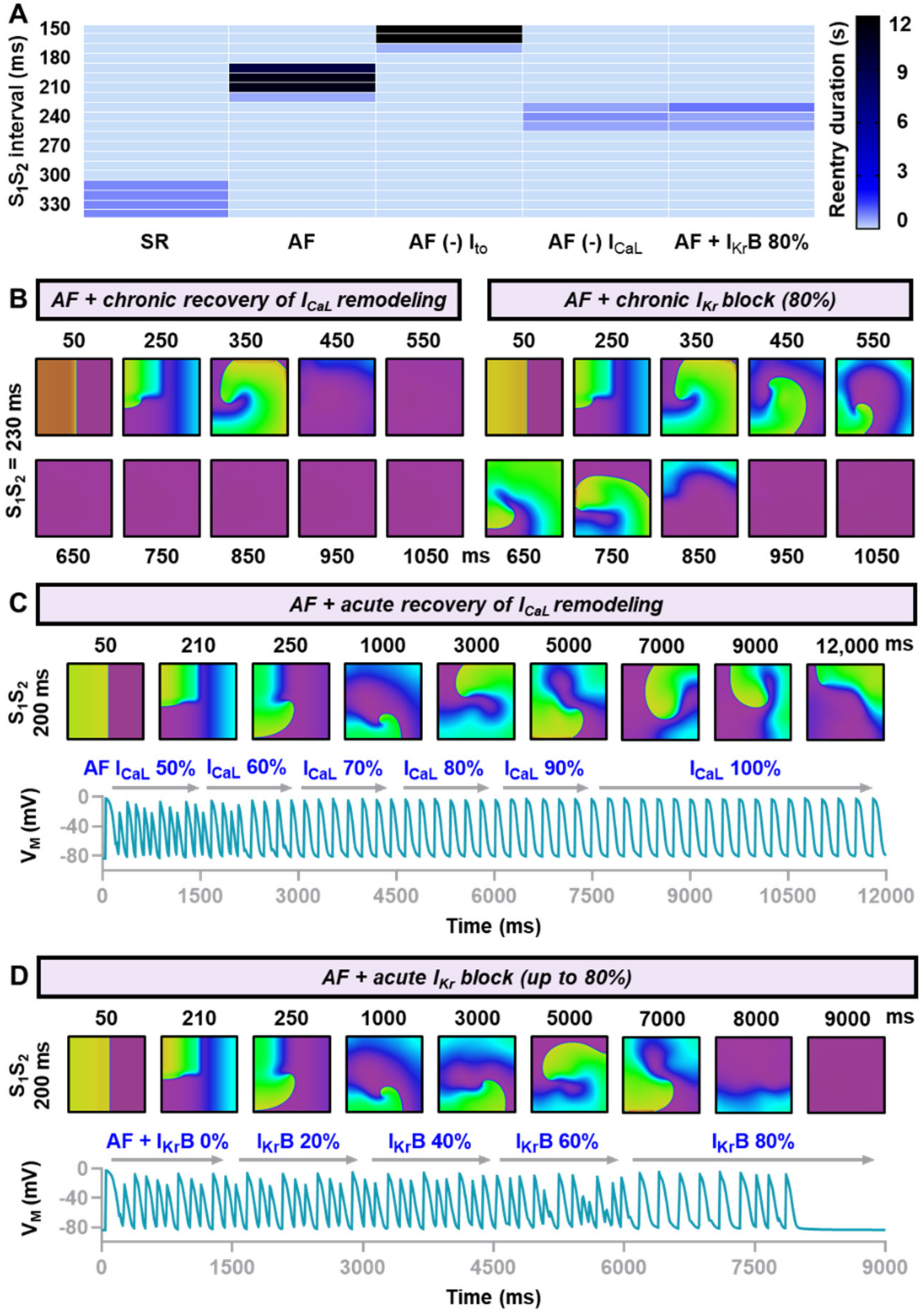
3.3. In-Silico Assessment of Antiarrhythmic Properties of Ionic Current Perturbations
4. Discussion
5. Conclusions
Supplementary Materials
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sutanto, H.; Lyon, A.; Lumens, J.; Schotten, U.; Dobrev, D.; Heijman, J. Cardiomyocyte calcium handling in health and disease: Insights from in vitro and in silico studies. Prog. Biophys. Mol. Biol. 2020, 157, 54–75. [Google Scholar] [CrossRef] [PubMed]
- Heijman, J.; Sutanto, H.; Crijns, H.; Nattel, S.; Trayanova, N.A. Computational models of atrial fibrillation: Achievements, challenges, and perspectives for improving clinical care. Cardiovasc. Res. 2021, 117, 1682–1699. [Google Scholar] [CrossRef]
- Sutanto, H.; Laudy, L.; Clerx, M.; Dobrev, D.; Crijns, H.; Heijman, J. Maastricht antiarrhythmic drug evaluator (MANTA): A computational tool for better understanding of antiarrhythmic drugs. Pharmacol. Res. 2019, 148, 104444. [Google Scholar] [CrossRef] [PubMed]
- Sutanto, H.; Cluitmans, M.J.M.; Dobrev, D.; Volders, P.G.A.; Bebarova, M.; Heijman, J. Acute effects of alcohol on cardiac electrophysiology and arrhythmogenesis: Insights from multiscale in silico analyses. J. Mol. Cell Cardiol. 2020, 146, 69–83. [Google Scholar] [CrossRef] [PubMed]
- Boyle, P.M.; Zahid, S.; Trayanova, N.A. Towards personalized computational modelling of the fibrotic substrate for atrial arrhythmia. Europace 2016, 18, iv136–iv145. [Google Scholar] [CrossRef]
- Courtemanche, M.; Ramirez, R.J.; Nattel, S. Ionic mechanisms underlying human atrial action potential properties: Insights from a mathematical model. Am. J. Physiol. 1998, 275, H301–H321. [Google Scholar] [CrossRef] [PubMed]
- Grandi, E.; Pandit, S.V.; Voigt, N.; Workman, A.J.; Dobrev, D.; Jalife, J.; Bers, D.M. Human atrial action potential and Ca2+ model: Sinus rhythm and chronic atrial fibrillation. Circ. Res. 2011, 109, 1055–1066. [Google Scholar] [CrossRef] [Green Version]
- Clerx, M.; Collins, P.; De Lange, E.; Volders, P.G. Myokit: A simple interface to cardiac cellular electrophysiology. Prog. Biophys. Mol. Biol. 2016, 120, 100–114. [Google Scholar] [CrossRef] [PubMed]
- Qu, Z.; Xie, F.; Garfinkel, A.; Weiss, J.N. Origins of spiral wave meander and breakup in a two-dimensional cardiac tissue model. Ann. Biomed. Eng. 2000, 28, 755–771. [Google Scholar] [CrossRef] [PubMed]
- Wilhelms, M.; Hettmann, H.; Maleckar, M.M.; Koivumaki, J.T.; Dossel, O.; Seemann, G. Benchmarking electrophysiological models of human atrial myocytes. Front. Physiol. 2012, 3, 487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yue, L.; Feng, J.; Li, G.R.; Nattel, S. Characterization of an ultrarapid delayed rectifier potassium channel involved in canine atrial repolarization. J. Physiol. 1996, 496, 647–662. [Google Scholar] [CrossRef] [PubMed]
- Aguilar, M.; Feng, J.; Vigmond, E.; Comtois, P.; Nattel, S. Rate-Dependent Role of IKur in Human Atrial Repolarization and Atrial Fibrillation Maintenance. Biophys. J. 2017, 112, 1997–2010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Fermini, B.; Nattel, S. Delayed rectifier outward current and repolarization in human atrial myocytes. Circ. Res. 1993, 73, 276–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, X.Y.; Diness, J.G.; Brundel, B.J.; Zhou, X.B.; Naud, P.; Wu, C.T.; Huang, H.; Harada, M.; Aflaki, M.; Dobrev, D.; et al. Role of small-conductance calcium-activated potassium channels in atrial electrophysiology and fibrillation in the dog. Circulation 2014, 129, 430–440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartos, D.C.; Grandi, E.; Ripplinger, C.M. Ion Channels in the Heart. Compr. Physiol. 2015, 5, 1423–1464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, C.; Wiedmann, F.; Voigt, N.; Zhou, X.B.; Heijman, J.; Lang, S.; Albert, V.; Kallenberger, S.; Ruhparwar, A.; Szabo, G.; et al. Upregulation of K2P3.1 K+ Current Causes Action Potential Shortening in Patients With Chronic Atrial Fibrillation. Circulation 2015, 132, 82–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez, C.; Corrias, A.; Bueno-Orovio, A.; Davies, M.; Swinton, J.; Jacobson, I.; Laguna, P.; Pueyo, E.; Rodriguez, B. The Na+/K+ pump is an important modulator of refractoriness and rotor dynamics in human atrial tissue. Am. J. Physiol. Heart Circ. Physiol. 2012, 302, H1146–H1159. [Google Scholar] [CrossRef] [PubMed]
- Burashnikov, A.; Belardinelli, L.; Antzelevitch, C. Inhibition of IKr potentiates development of atrial-selective INa block leading to effective suppression of atrial fibrillation. Heart Rhythm 2015, 12, 836–844. [Google Scholar] [CrossRef]
- Shibata, E.F.; Drury, T.; Refsum, H.; Aldrete, V.; Giles, W. Contributions of a transient outward current to repolarization in human atrium. Am. J. Physiol. 1989, 257, H1773–H1781. [Google Scholar] [CrossRef]
- Fermini, B.; Wang, Z.; Duan, D.; Nattel, S. Differences in rate dependence of transient outward current in rabbit and human atrium. Am. J. Physiol. 1992, 263, H1747–H1754. [Google Scholar] [CrossRef]
- Ni, H.; Zhang, H.; Grandi, E.; Narayan, S.M.; Giles, W.R. Transient outward K+ current can strongly modulate action potential duration and initiate alternans in the human atrium. Am. J. Physiol. Heart Circ. Physiol. 2019, 316, H527–H542. [Google Scholar] [CrossRef] [PubMed]
- Pandit, S.V.; Jalife, J. Rotors and the dynamics of cardiac fibrillation. Circ. Res. 2013, 112, 849–862. [Google Scholar] [CrossRef] [Green Version]
- Malhotra, R.; Bilchick, K.C.; DiMarco, J.P. Usefulness of pharmacologic conversion of atrial fibrillation during dofetilide loading without the need for electrical cardioversion to predict durable response to therapy. Am. J. Cardiol. 2014, 113, 475–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Donahue, J.K. The Use of Gene Therapy for Ablation of Atrial Fibrillation. Arrhythm. Electrophysiol. Rev. 2014, 3, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Fermini, B.; Nattel, S. Rapid and slow components of delayed rectifier current in human atrial myocytes. Cardiovasc. Res. 1994, 28, 1540–1546. [Google Scholar] [CrossRef] [PubMed]
- Severi, S.; Corsi, C.; Cerbai, E. From in vivo plasma composition to in vitro cardiac electrophysiology and in silico virtual heart: The extracellular calcium enigma. Philos. Trans. A Math Phys. Eng. Sci. 2009, 367, 2203–2223. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sutanto, H. Individual Contributions of Cardiac Ion Channels on Atrial Repolarization and Reentrant Waves: A Multiscale In-Silico Study. J. Cardiovasc. Dev. Dis. 2022, 9, 28. https://doi.org/10.3390/jcdd9010028
Sutanto H. Individual Contributions of Cardiac Ion Channels on Atrial Repolarization and Reentrant Waves: A Multiscale In-Silico Study. Journal of Cardiovascular Development and Disease. 2022; 9(1):28. https://doi.org/10.3390/jcdd9010028
Chicago/Turabian StyleSutanto, Henry. 2022. "Individual Contributions of Cardiac Ion Channels on Atrial Repolarization and Reentrant Waves: A Multiscale In-Silico Study" Journal of Cardiovascular Development and Disease 9, no. 1: 28. https://doi.org/10.3390/jcdd9010028
APA StyleSutanto, H. (2022). Individual Contributions of Cardiac Ion Channels on Atrial Repolarization and Reentrant Waves: A Multiscale In-Silico Study. Journal of Cardiovascular Development and Disease, 9(1), 28. https://doi.org/10.3390/jcdd9010028