
The Early Stages of Heart Development: Insights from Chicken Embryos


{kind=link}
{kind=link}
{kind=link}
Abstract
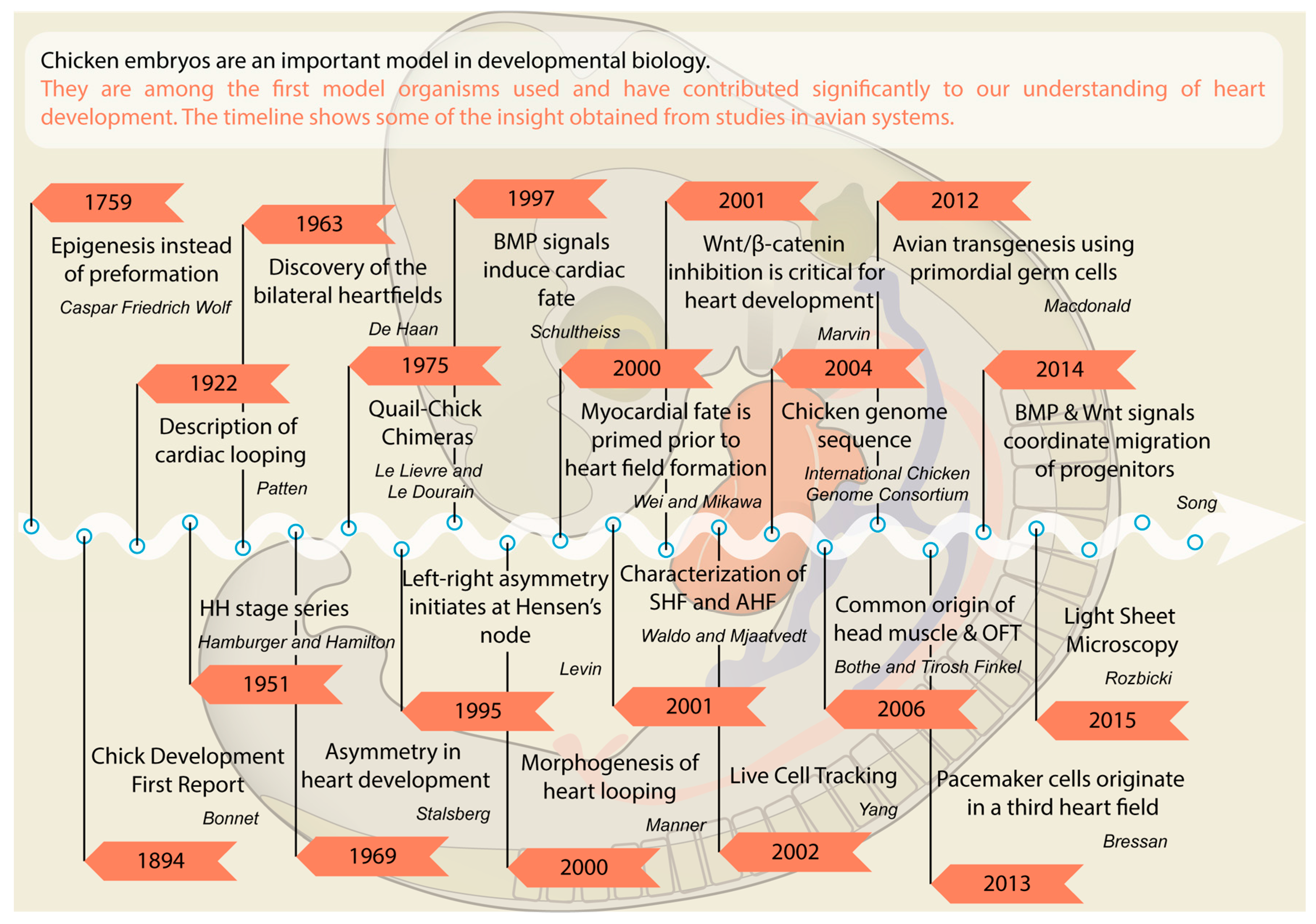
:1. Introduction
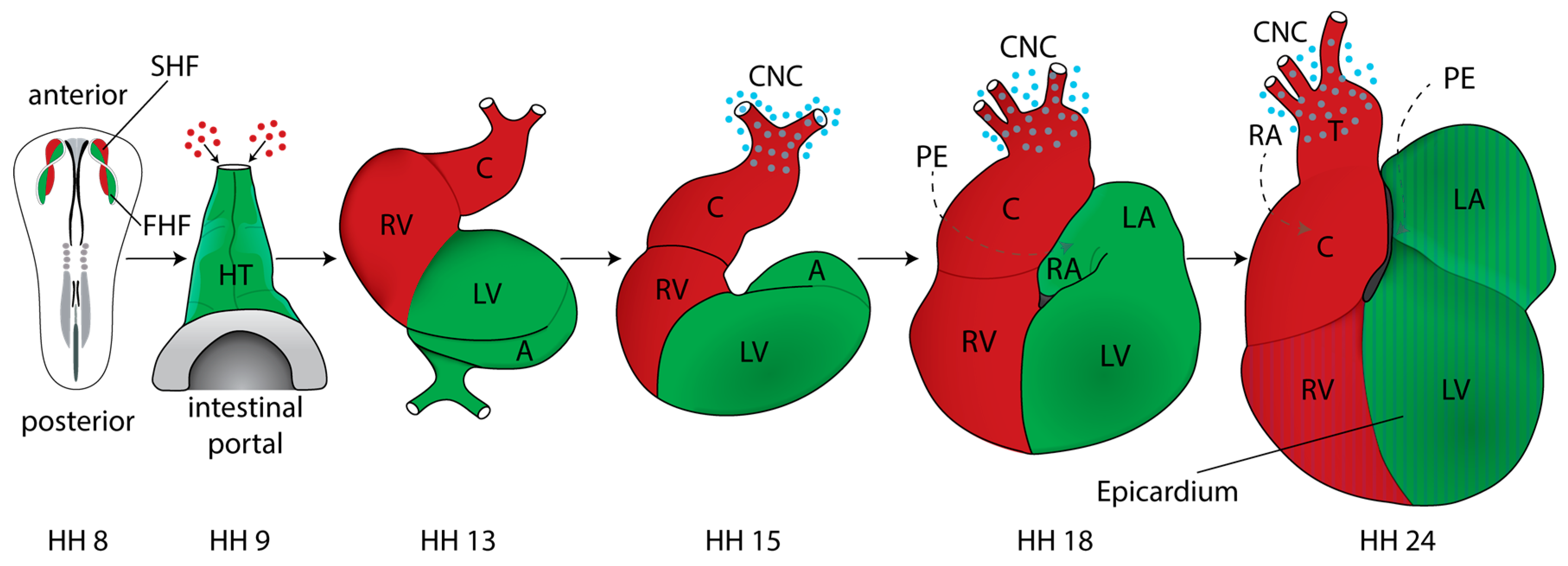
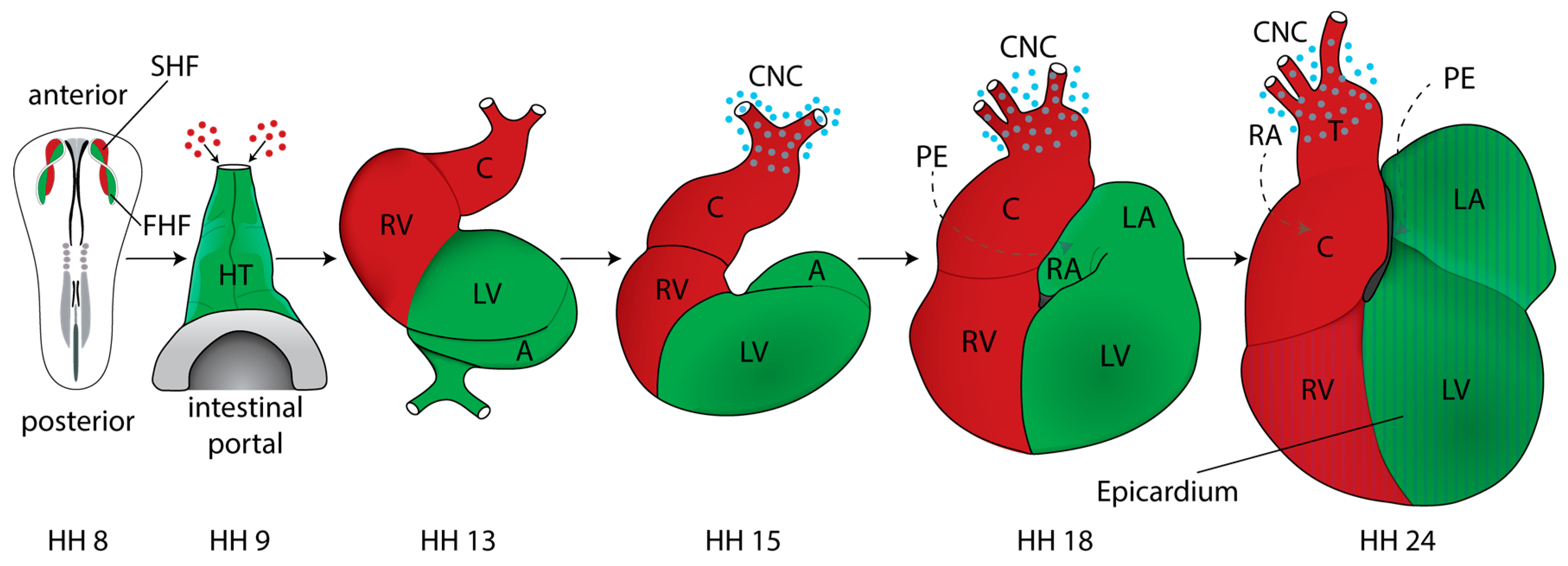
2. Cardiac Development and Morphogenesis
2.1. Mapping Studies and Characterization of Cardiogenic Fields
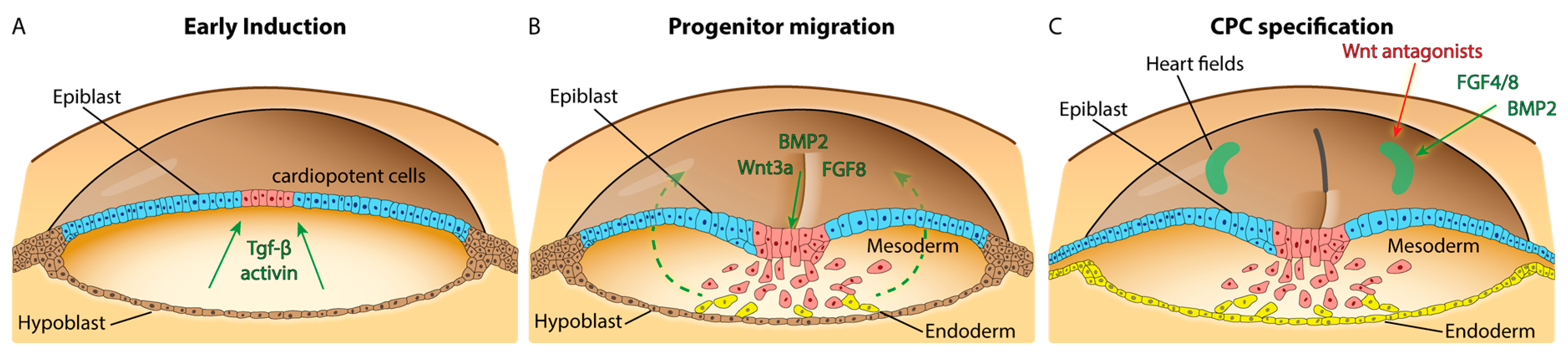
2.2. Pre-Gastrula and Gastrula Stages
2.2.1. Specification and Migration of Cardiac Progenitor Cells
2.2.2. Establishment of Left–Right Asymmetry
2.3. Discovery of Additional Heart Fields
2.4. Formation and Transformation of the Straight Heart Tube
2.5. Cardiac Neural Crest
2.6. Cardiac Chambers
2.7. The Proepicardium
3. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| AHF/SHF | anterior/secondary heart field |
| HH | Hamburger-Hamilton |
| NCC | neural crest cells |
| OFT | outflow tract |
| PC | pacemaker cell |
| PE | proepicardium |
References
- B, X. Charles Bonnet’s idea of the development of the chick. Science 1894, 23, 71–72. [Google Scholar] [CrossRef] [PubMed][Green Version]
- St Johnston, D. The renaissance of developmental biology. PLoS Biol. 2015, 13, e1002149. [Google Scholar] [CrossRef] [PubMed]
- Stern, C.D. The chick; a great model system becomes even greater. Dev. Cell 2005, 8, 9–17. [Google Scholar] [PubMed]
- Kain, K.H.; Miller, J.W.; Jones-Paris, C.R.; Thomason, R.T.; Lewis, J.D.; Bader, D.M.; Barnett, J.V.; Zijlstra, A. The chick embryo as an expanding experimental model for cancer and cardiovascular research. Dev. Dyn. 2014, 243, 216–228. [Google Scholar] [CrossRef] [PubMed]
- Stern, C.D. The chick embryo-past, present and future as a model system in developmental biology. Mech. Dev. 2004, 121, 1011–1013. [Google Scholar] [CrossRef] [PubMed]
- Le Douarin, N. A biological cell labeling technique and its use in experimental embryology. Dev. Biol. 1973, 30, 217–222. [Google Scholar] [CrossRef]
- Yang, X.; Dormann, D.; Münsterberg, A.E.; Weijer, C.J. Cell movement patterns during gastrulation in the chick are controlled by positive and negative chemotaxis mediated by fgf4 and fgf8. Dev. Cell 2002, 3, 425–437. [Google Scholar] [CrossRef]
- Rozbicki, E.; Chuai, M.; Karjalainen, A.I.; Song, F.; Sang, H.M.; Martin, R.; Knolker, H.J.; MacDonald, M.P.; Weijer, C.J. Myosin-ii-mediated cell shape changes and cell intercalation contribute to primitive streak formation. Nat. Cell Biol. 2015, 17, 397–408. [Google Scholar] [CrossRef] [PubMed]
- Grocott, T.; Thomas, P.; Münsterberg, A.E. Atlas toolkit: Fast registration of 3D morphological datasets in the absence of landmarks. Sci. Rep. 2016, 6, 20732. [Google Scholar] [CrossRef] [PubMed]
- Zamir, E.A.; Czirok, A.; Cui, C.; Little, C.D.; Rongish, B.J. Mesodermal cell displacements during avian gastrulation are due to both individual cell-autonomous and convective tissue movements. Proc. Natl. Acad. Sci. USA 2006, 103, 19806–19811. [Google Scholar] [CrossRef] [PubMed]
- Zamir, E.A.; Rongish, B.J.; Little, C.D. The ECM moves during primitive streak formation--computation of ECM versus cellular motion. PLoS Biol. 2008, 6, e247. [Google Scholar] [CrossRef] [PubMed]
- Veron, N.; Qu, Z.; Kipen, P.A.; Hirst, C.E.; Marcelle, C. Crispr mediated somatic cell genome engineering in the chicken. Dev. Biol. 2015, 407, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Cui, C.; Filla, M.B.; Jones, E.A.; Lansford, R.; Cheuvront, T.; Al-Roubaie, S.; Rongish, B.J.; Little, C.D. Embryogenesis of the first circulating endothelial cells. PLoS ONE 2013, 8, e60841. [Google Scholar] [CrossRef] [PubMed]
- Balic, A.; Garcia-Morales, C.; Vervelde, L.; Gilhooley, H.; Sherman, A.; Garceau, V.; Gutowska, M.W.; Burt, D.W.; Kaiser, P.; Hume, D.A.; et al. Visualisation of chicken macrophages using transgenic reporter genes: Insights into the development of the avian macrophage lineage. Development 2014, 141, 3255–3265. [Google Scholar] [CrossRef] [PubMed]
- Macdonald, J.; Taylor, L.; Sherman, A.; Kawakami, K.; Takahashi, Y.; Sang, H.M.; McGrew, M.J. Efficient genetic modification and germ-line transmission of primordial germ cells using piggybac and tol2 transposons. Proc. Natl. Acad. Sci. USA 2012, 109, E1466–E1472. [Google Scholar] [CrossRef] [PubMed]
- Plein, A.; Fantin, A.; Ruhrberg, C. Neural crest cells in cardiovascular development. Curr. Top. Dev. Biol. 2015, 111, 183–200. [Google Scholar] [PubMed]
- Hamburger, V.; Hamilton, H.L. A series of normal stages in the development of the chick embryo. J. Morphol. 1951, 88, 49–92. [Google Scholar] [CrossRef] [PubMed]
- Martinsen, B.J. Reference guide to the stages of chick heart embryology. Dev. Dyn. 2005, 233, 1217–1237. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Sanchez, C.; Puelles, L.; Garcia-Martinez, V.; Rodriguez-Gallardo, L. Morphological and molecular analysis of the early developing chick requires an expanded series of primitive streak stages. J. Morphol. 2005, 264, 105–116. [Google Scholar] [CrossRef] [PubMed]
- DeHaan, R.L. Organization of the cardiogenic plate in the early chick embryo. Acta Embryol. Morphol. Exp. 1963, 6, 26–38. [Google Scholar]
- Garcia-Martinez, V.; Schoenwolf, G.C. Primitive-streak origin of the cardiovascular system in avian embryos. Dev. Biol. 1993, 159, 706–719. [Google Scholar] [CrossRef] [PubMed]
- Bellairs, R. The primitive streak. Anat. Embryol. (Berl.) 1986, 174, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Rosenquist, G.C. Location and movements of cardiogenic cells in the chick embryo: The heart-forming portion of the primitive streak. Dev. Biol. 1970, 22, 461–475. [Google Scholar] [CrossRef]
- Psychoyos, D.; Stern, C.D. Fates and migratory routes of primitive streak cells in the chick embryo. Development 1996, 122, 1523–1534. [Google Scholar] [PubMed]
- Stalsberg, H.; DeHaan, R.L. The precardiac areas and formation of the tubular heart in the chick embryo. Dev. Biol. 1969, 19, 128–159. [Google Scholar] [CrossRef]
- Rosenquist, G.C.; DeHaan, R.L. Migration of precardiac cells in the chick embryo: A radioautographic study; Carnegie Institution of Washington: Washington, DC, USA, 1966. [Google Scholar]
- DeRuiter, M.C.; Poelmann, R.E.; VanderPlas-de Vries, I.; Mentink, M.M.; Gittenberger-de Groot, A.C. The development of the myocardium and endocardium in mouse embryos. Fusion of two heart tubes? Anat. Embryol. (Berl.) 1992, 185, 461–473. [Google Scholar] [CrossRef] [PubMed]
- Colas, J.F.; Lawson, A.; Schoenwolf, G.C. Evidence that translation of smooth muscle alpha-actin mrna is delayed in the chick promyocardium until fusion of the bilateral heart-forming regions. Dev. Dyn. 2000, 218, 316–330. [Google Scholar] [CrossRef]
- Le Lievre, C.S.; Le Douarin, N.M. Mesenchymal derivatives of the neural crest: Analysis of chimaeric quail and chick embryos. J. Embryol. Exp. Morphol. 1975, 34, 125–154. [Google Scholar] [PubMed]
- Hatada, Y.; Stern, C.D. A fate map of the epiblast of the early chick embryo. Development 1994, 120, 2879–2889. [Google Scholar] [PubMed]
- Ladd, A.N.; Yatskievych, T.A.; Antin, P.B. Regulation of avian cardiac myogenesis by activin/tgfbeta and bone morphogenetic proteins. Dev. Biol. 1998, 204, 407–419. [Google Scholar] [CrossRef] [PubMed]
- Yatskievych, T.A.; Ladd, A.N.; Antin, P.B. Induction of cardiac myogenesis in avian pregastrula epiblast: The role of the hypoblast and activin. Development 1997, 124, 2561–2570. [Google Scholar] [PubMed]
- Matsui, H.; Ikeda, K.; Nakatani, K.; Sakabe, M.; Yamagishi, T.; Nakanishi, T.; Nakajima, Y. Induction of initial cardiomyocyte alpha-actin--smooth muscle alpha-actin--in cultured avian pregastrula epiblast: A role for nodal and bmp antagonist. Dev. Dyn. 2005, 233, 1419–1429. [Google Scholar] [CrossRef] [PubMed]
- Tam, P.P.; Parameswaran, M.; Kinder, S.J.; Weinberger, R.P. The allocation of epiblast cells to the embryonic heart and other mesodermal lineages: The role of ingression and tissue movement during gastrulation. Development 1997, 124, 1631–1642. [Google Scholar] [PubMed]
- Wei, Y.; Mikawa, T. Fate diversity of primitive streak cells during heart field formation in ovo. Dev. Dyn. 2000, 219, 505–513. [Google Scholar] [CrossRef]
- Schultheiss, T.M.; Xydas, S.; Lassar, A.B. Induction of avian cardiac myogenesis by anterior endoderm. Development 1995, 121, 4203–4214. [Google Scholar] [PubMed]
- Schultheiss, T.M.; Burch, J.B.; Lassar, A.B. A role for bone morphogenetic proteins in the induction of cardiac myogenesis. Genes Dev. 1997, 11, 451–462. [Google Scholar] [CrossRef] [PubMed]
- Andree, B.; Duprez, D.; Vorbusch, B.; Arnold, H.H.; Brand, T. Bmp-2 induces ectopic expression of cardiac lineage markers and interferes with somite formation in chicken embryos. Mech. Dev. 1998, 70, 119–131. [Google Scholar] [CrossRef]
- Schlange, T.; Andree, B.; Arnold, H.H.; Brand, T. Bmp2 is required for early heart development during a distinct time period. Mech. Dev. 2000, 91, 259–270. [Google Scholar] [CrossRef]
- Alsan, B.H.; Schultheiss, T.M. Regulation of avian cardiogenesis by fgf8 signaling. Development 2002, 129, 1935–1943. [Google Scholar] [PubMed]
- Lee, K.H.; Evans, S.; Ruan, T.Y.; Lassar, A.B. Smad-mediated modulation of yy1 activity regulates the bmp response and cardiac-specific expression of a gata4/5/6-dependent chick nkx2.5 enhancer. Development 2004, 131, 4709–4723. [Google Scholar] [CrossRef] [PubMed]
- Marvin, M.J.; Di Rocco, G.; Gardiner, A.; Bush, S.M.; Lassar, A.B. Inhibition of wnt activity induces heart formation from posterior mesoderm. Genes Dev. 2001, 15, 316–327. [Google Scholar] [CrossRef] [PubMed]
- Schneider, V.A.; Mercola, M. Wnt antagonism initiates cardiogenesis in Xenopus laevis. Genes Dev. 2001, 15, 304–315. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Villalba, A.; Hoppler, S.; van den Hoff, M.J. Wnt signaling in the heart fields: Variations on a common theme. Dev. Dyn. 2016, 245, 294–306. [Google Scholar] [CrossRef] [PubMed]
- Lickert, H.; Kutsch, S.; Kanzler, B.; Tamai, Y.; Taketo, M.M.; Kemler, R. Formation of multiple hearts in mice following deletion of beta-catenin in the embryonic endoderm. Dev. Cell 2002, 3, 171–181. [Google Scholar] [CrossRef]
- Tzahor, E. Wnt/beta-catenin signaling and cardiogenesis: Timing does matter. Dev. Cell 2007, 13, 10–13. [Google Scholar] [CrossRef] [PubMed]
- Noseda, M.; Peterkin, T.; Simoes, F.C.; Patient, R.; Schneider, M.D. Cardiopoietic factors: Extracellular signals for cardiac lineage commitment. Circ. Res. 2011, 108, 129–152. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Sanchez, C.; Franco, D.; Bonet, F.; Garcia-Lopez, V.; Aranega, A.; Garcia-Martinez, V. Negative fgf8-bmp2 feed-back is regulated by mir-130 during early cardiac specification. Dev. Biol. 2015, 406, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Sanchez, C.; Franco, D.; Bonet, F.; Garcia-Lopez, V.; Aranega, A.; Garcia-Martinez, V. Reciprocal repression between fgf8 and mir-133 regulates cardiac induction through bmp2 signaling. Data in brief 2015, 5, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Yue, Q.; Wagstaff, L.; Yang, X.; Weijer, C.; Münsterberg, A. Wnt3a-mediated chemorepulsion controls movement patterns of cardiac progenitors and requires rhoa function. Development 2008, 135, 1029–1037. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kaarbo, M.; Crane, D.I.; Murrell, W.G. Rhoa is highly up-regulated in the process of early heart development of the chick and important for normal embryogenesis. Dev. Dyn. 2003, 227, 35–47. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; McColl, J.; Camp, E.; Kennerley, N.; Mok, G.F.; McCormick, D.; Grocott, T.; Wheeler, G.N.; Münsterberg, A.E. Smad1 transcription factor integrates bmp2 and wnt3a signals in migrating cardiac progenitor cells. Proc. Natl. Acad. Sci. USA 2014, 111, 7337–7342. [Google Scholar] [CrossRef] [PubMed]
- Klaus, A.; Saga, Y.; Taketo, M.M.; Tzahor, E.; Birchmeier, W. Distinct roles of wnt/beta-catenin and bmp signaling during early cardiogenesis. Proc. Natl. Acad. Sci. USA 2007, 104, 18531–18536. [Google Scholar] [CrossRef] [PubMed]
- Udan, R.S.; Piazza, V.G.; Hsu, C.W.; Hadjantonakis, A.K.; Dickinson, M.E. Quantitative imaging of cell dynamics in mouse embryos using light-sheet microscopy. Development 2014, 141, 4406–4414. [Google Scholar] [CrossRef] [PubMed]
- Ramsdell, A.F. Left-right asymmetry and congenital cardiac defects: Getting to the heart of the matter in vertebrate left-right axis determination. Dev. Biol. 2005, 288, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Levin, M.; Johnson, R.L.; Stern, C.D.; Kuehn, M.; Tabin, C. A molecular pathway determining left-right asymmetry in chick embryogenesis. Cell 1995, 82, 803–814. [Google Scholar] [CrossRef]
- Mendes, R.V.; Martins, G.G.; Cristovao, A.M.; Saude, L. N-cadherin locks left-right asymmetry by ending the leftward movement of Hensen’s node cells. Dev. Cell 2014, 30, 353–360. [Google Scholar] [CrossRef] [PubMed]
- Collignon, J.; Varlet, I.; Robertson, E.J. Relationship between asymmetric nodal expression and the direction of embryonic turning. Nature 1996, 381, 155–158. [Google Scholar] [CrossRef] [PubMed]
- Meyers, E.N.; Martin, G.R. Differences in left-right axis pathways in mouse and chick: Functions of fgf8 and shh. Science 1999, 285, 403–406. [Google Scholar] [CrossRef] [PubMed]
- Schlueter, J.; Brand, T. Left-right axis development: Examples of similar and divergent strategies to generate asymmetric morphogenesis in chick and mouse embryos. Cytogenet. Genome Res. 2007, 117, 256–267. [Google Scholar] [CrossRef] [PubMed]
- Logan, M.; Pagan-Westphal, S.M.; Smith, D.M.; Paganessi, L.; Tabin, C.J. The transcription factor pitx2 mediates situs-specific morphogenesis in response to left-right asymmetric signals. Cell 1998, 94, 307–317. [Google Scholar] [CrossRef]
- Tessari, A.; Pietrobon, M.; Notte, A.; Cifelli, G.; Gage, P.J.; Schneider, M.D.; Lembo, G.; Campione, M. Myocardial pitx2 differentially regulates the left atrial identity and ventricular asymmetric remodeling programs. Circ. Res. 2008, 102, 813–822. [Google Scholar] [CrossRef] [PubMed]
- Schlueter, J.; Brand, T. A right-sided pathway involving fgf8/snai1 controls asymmetric development of the proepicardium in the chick embryo. Proc. Natl. Acad. Sci. USA 2009, 106, 7485–7490. [Google Scholar] [CrossRef] [PubMed]
- Schulte, I.; Schlueter, J.; Abu-Issa, R.; Brand, T.; Manner, J. Morphological and molecular left-right asymmetries in the development of the proepicardium: A comparative analysis on mouse and chick embryos. Dev. Dyn. 2007, 236, 684–695. [Google Scholar] [CrossRef] [PubMed]
- de la Cruz, M.V.; Sanchez Gomez, C.; Arteaga, M.M.; Arguello, C. Experimental study of the development of the truncus and the conus in the chick embryo. J. Anat. 1977, 123, 661–686. [Google Scholar] [PubMed]
- Arguello, C.; de la Cruz, M.V.; Gomez, C.S. Experimental study of the formation of the heart tube in the chick embryo. J. Embryol. Exp. Morphol. 1975, 33, 1–11. [Google Scholar] [PubMed]
- de La Cruz, M.V.; Sanchez-Gomez, C.; Palomino, M.A. The primitive cardiac regions in the straight tube heart (stage 9) and their anatomical expression in the mature heart: An experimental study in the chick embryo. J. Anat. 1989, 165, 121–131. [Google Scholar] [PubMed]
- Kelly, R.G.; Brown, N.A.; Buckingham, M.E. The arterial pole of the mouse heart forms from fgf10-expressing cells in pharyngeal mesoderm. Dev. Cell 2001, 1, 435–440. [Google Scholar] [CrossRef]
- Mjaatvedt, C.H.; Nakaoka, T.; Moreno-Rodriguez, R.; Norris, R.A.; Kern, M.J.; Eisenberg, C.A.; Turner, D.; Markwald, R.R. The outflow tract of the heart is recruited from a novel heart-forming field. Dev. Biol. 2001, 238, 97–109. [Google Scholar] [CrossRef] [PubMed]
- Waldo, K.L.; Kumiski, D.H.; Wallis, K.T.; Stadt, H.A.; Hutson, M.R.; Platt, D.H.; Kirby, M.L. Conotruncal myocardium arises from a secondary heart field. Development 2001, 128, 3179–3188. [Google Scholar] [PubMed]
- Camp, E.; Dietrich, S.; Münsterberg, A. Fate mapping identifies the origin of SHF/AHFprogenitors in the chick primitive streak. PLoS ONE 2012, 7, e51948. [Google Scholar] [CrossRef] [PubMed]
- Nathan, E.; Monovich, A.; Tirosh-Finkel, L.; Harrelson, Z.; Rousso, T.; Rinon, A.; Harel, I.; Evans, S.M.; Tzahor, E. The contribution of islet1-expressing splanchnic mesoderm cells to distinct branchiomeric muscles reveals significant heterogeneity in head muscle development. Development 2008, 135, 647–657. [Google Scholar] [CrossRef] [PubMed]
- Lescroart, F.; Kelly, R.G.; Le Garrec, J.F.; Nicolas, J.F.; Meilhac, S.M.; Buckingham, M. Clonal analysis reveals common lineage relationships between head muscles and second heart field derivatives in the mouse embryo. Development 2010, 137, 3269–3279. [Google Scholar] [CrossRef] [PubMed]
- Milgrom-Hoffman, M.; Harrelson, Z.; Ferrara, N.; Zelzer, E.; Evans, S.M.; Tzahor, E. The heart endocardium is derived from vascular endothelial progenitors. Development 2011, 138, 4777–4787. [Google Scholar] [CrossRef] [PubMed]
- Misfeldt, A.M.; Boyle, S.C.; Tompkins, K.L.; Bautch, V.L.; Labosky, P.A.; Baldwin, H.S. Endocardial cells are a distinct endothelial lineage derived from flk1+ multipotent cardiovascular progenitors. Dev. Biol. 2009, 333, 78–89. [Google Scholar] [CrossRef] [PubMed]
- Ferdous, A.; Caprioli, A.; Iacovino, M.; Martin, C.M.; Morris, J.; Richardson, J.A.; Latif, S.; Hammer, R.E.; Harvey, R.P.; Olson, E.N.; et al. Nkx2–5 transactivates the ets-related protein 71 gene and specifies an endothelial/endocardial fate in the developing embryo. Proc. Natl. Acad. Sci. USA 2009, 106, 814–819. [Google Scholar] [CrossRef] [PubMed]
- Dyer, L.A.; Kirby, M.L. The role of secondary heart field in cardiac development. Dev. Biol. 2009, 336, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Vincent, S.D.; Buckingham, M.E. How to make a heart: The origin and regulation of cardiac progenitor cells. Curr. Top. Dev. Biol. 2010, 90, 1–41. [Google Scholar] [PubMed]
- Buckingham, M.; Meilhac, S.; Zaffran, S. Building the mammalian heart from two sources of myocardial cells. Nature reviews. Genetics 2005, 6, 826–835. [Google Scholar] [CrossRef] [PubMed]
- Hutson, M.R.; Zeng, X.L.; Kim, A.J.; Antoon, E.; Harward, S.; Kirby, M.L. Arterial pole progenitors interpret opposing fgf/bmp signals to proliferate or differentiate. Development 2010, 137, 3001–3011. [Google Scholar] [CrossRef] [PubMed]
- Tirosh-Finkel, L.; Zeisel, A.; Brodt-Ivenshitz, M.; Shamai, A.; Yao, Z.; Seger, R.; Domany, E.; Tzahor, E. Bmp-mediated inhibition of fgf signaling promotes cardiomyocyte differentiation of anterior heart field progenitors. Development 2010, 137, 2989–3000. [Google Scholar] [CrossRef] [PubMed]
- Bothe, I.; Dietrich, S. The molecular setup of the avian head mesoderm and its implication for craniofacial myogenesis. Dev. Dyn. 2006, 235, 2845–2860. [Google Scholar] [CrossRef] [PubMed]
- Tirosh-Finkel, L.; Elhanany, H.; Rinon, A.; Tzahor, E. Mesoderm progenitor cells of common origin contribute to the head musculature and the cardiac outflow tract. Development 2006, 133, 1943–1953. [Google Scholar] [CrossRef] [PubMed]
- Bothe, I.; Tenin, G.; Oseni, A.; Dietrich, S. Dynamic control of head mesoderm patterning. Development 2011, 138, 2807–2821. [Google Scholar] [CrossRef] [PubMed]
- Grifone, R.; Kelly, R.G. Heartening news for head muscle development. Trends in genetics : TIG 2007, 23, 365–369. [Google Scholar] [CrossRef] [PubMed]
- Diogo, R.; Kelly, R.G.; Christiaen, L.; Levine, M.; Ziermann, J.M.; Molnar, J.L.; Noden, D.M.; Tzahor, E. A new heart for a new head in vertebrate cardiopharyngeal evolution. Nature 2015, 520, 466–473. [Google Scholar] [CrossRef] [PubMed]
- Bressan, M.; Liu, G.; Mikawa, T. Early mesodermal cues assign avian cardiac pacemaker fate potential in a tertiary heart field. Science 2013, 340, 744–748. [Google Scholar] [CrossRef] [PubMed]
- Kamino, K.; Hirota, A.; Fujii, S. Localization of pacemaking activity in early embryonic heart monitored using voltage-sensitive dye. Nature 1981, 290, 595–597. [Google Scholar] [CrossRef] [PubMed]
- Needham, J.; Hughes, A. A history of embryology, 2nd ed.; Cambridge University Press: London, UK, 1959. [Google Scholar]
- Patten, B.M. The formation of the cardiac loop in the chick. Am. J. Anat. 1922, 30, 373–397. [Google Scholar] [CrossRef]
- Varner, V.D.; Taber, L.A. Not just inductive: A crucial mechanical role for the endoderm during heart tube assembly. Development 2012, 139, 1680–1690. [Google Scholar] [CrossRef] [PubMed]
- DeHaan, R.L. Cardia bifida and the development of pacemaker function in the early chick heart. Dev. Biol. 1959, 1, 586–602. [Google Scholar] [CrossRef]
- Kitajima, S.; Takagi, A.; Inoue, T.; Saga, Y. Mesp1 and mesp2 are essential for the development of cardiac mesoderm. Development 2000, 127, 3215–3226. [Google Scholar] [PubMed]
- Matsui, T.; Raya, A.; Kawakami, Y.; Callol-Massot, C.; Capdevila, J.; Rodriguez-Esteban, C.; Izpisua Belmonte, J.C. Noncanonical wnt signaling regulates midline convergence of organ primordia during zebrafish development. Genes Dev. 2005, 19, 164–175. [Google Scholar] [CrossRef] [PubMed]
- Manner, J. Cardiac looping in the chick embryo: A morphological review with special reference to terminological and biomechanical aspects of the looping process. Anat. Rec. 2000, 259, 248–262. [Google Scholar] [CrossRef]
- Manner, J. The anatomy of cardiac looping: A step towards the understanding of the morphogenesis of several forms of congenital cardiac malformations. Clin. Anat. 2009, 22, 21–35. [Google Scholar] [CrossRef] [PubMed]
- de la Cruz, M.; Sanchez-Gomez, C. Straight tube heart. Primitive cardiac cavities vs. Primitive cardiac segments. In Living Morphogenesis of the Heart; de la Cruz, M., Markwald, R., Eds.; Birkhäuser Boston: Boston, MA, USA, 1998; pp. 85–98. [Google Scholar]
- Goenezen, S.; Rennie, M.Y.; Rugonyi, S. Biomechanics of early cardiac development. Biomech. Model. Mechanobiol. 2012, 11, 1187–1204. [Google Scholar] [CrossRef] [PubMed]
- Le Douarin, N.M. The avian embryo as a model to study the development of the neural crest: A long and still ongoing story. Mech. Dev. 2004, 121, 1089–1102. [Google Scholar] [CrossRef] [PubMed]
- Kirby, M.L.; Gale, T.F.; Stewart, D.E. Neural crest cells contribute to normal aorticopulmonary septation. Science 1983, 220, 1059–1061. [Google Scholar] [CrossRef] [PubMed]
- Waldo, K.; Miyagawa-Tomita, S.; Kumiski, D.; Kirby, M.L. Cardiac neural crest cells provide new insight into septation of the cardiac outflow tract: Aortic sac to ventricular septal closure. Dev. Biol. 1998, 196, 129–144. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Ruest, L.B. Analysis of neural crest cell fate during cardiovascular development using cre-activated lacz/beta-galactosidase staining. Methods Mol. Biol. 2012, 843, 125–138. [Google Scholar] [PubMed]
- Escot, S.; Blavet, C.; Hartle, S.; Duband, J.L.; Fournier-Thibault, C. Misregulation of sdf1-cxcr4 signaling impairs early cardiac neural crest cell migration leading to conotruncal defects. Circ. Res. 2013, 113, 505–516. [Google Scholar] [CrossRef] [PubMed]
- Sierro, F.; Biben, C.; Martinez-Munoz, L.; Mellado, M.; Ransohoff, R.M.; Li, M.; Woehl, B.; Leung, H.; Groom, J.; Batten, M.; et al. Disrupted cardiac development but normal hematopoiesis in mice deficient in the second cxcl12/sdf-1 receptor, cxcr7. Proc. Natl. Acad. Sci. USA 2007, 104, 14759–14764. [Google Scholar] [CrossRef] [PubMed]
- Bressan, M.; Yang, P.B.; Louie, J.D.; Navetta, A.M.; Garriock, R.J.; Mikawa, T. Reciprocal myocardial-endocardial interactions pattern the delay in atrioventricular junction conduction. Development 2014, 141, 4149–4157. [Google Scholar] [CrossRef] [PubMed]
- Bonet, F.; Duenas, A.; Lopez-Sanchez, C.; Garcia-Martinez, V.; Aranega, A.E.; Franco, D. Mir-23b and mir-199a impair epithelial-to-mesenchymal transition during atrioventricular endocardial cushion formation. Dev. Dyn. 2015, 244, 1259–1275. [Google Scholar] [CrossRef] [PubMed]
- Samsa, L.A.; Yang, B.; Liu, J. Embryonic cardiac chamber maturation: Trabeculation, conduction, and cardiomyocyte proliferation. Am. J. Med. Genet. C Semin. Med. Genet. 2013, 163C, 157–168. [Google Scholar] [CrossRef] [PubMed]
- Moorman, A.F.; Christoffels, V.M. Cardiac chamber formation: Development, genes, and evolution. Physiol. Rev. 2003, 83, 1223–1267. [Google Scholar] [CrossRef] [PubMed]
- Mikawa, T.; Gourdie, R.G. Pericardial mesoderm generates a population of coronary smooth muscle cells migrating into the heart along with ingrowth of the epicardial organ. Dev. Biol. 1996, 174, 221–232. [Google Scholar] [CrossRef] [PubMed]
- Reese, D.E.; Mikawa, T.; Bader, D.M. Development of the coronary vessel system. Circ. Res. 2002, 91, 761–768. [Google Scholar] [CrossRef] [PubMed]
- Kattan, J.; Dettman, R.W.; Bristow, J. Formation and remodeling of the coronary vascular bed in the embryonic avian heart. Dev. Dyn. 2004, 230, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Olivey, H.E.; Compton, L.A.; Barnett, J.V. Coronary vessel development: The epicardium delivers. Trends Cardiovasc. Med. 2004, 14, 247–251. [Google Scholar] [CrossRef] [PubMed]
- Manner, J.; Schlueter, J.; Brand, T. Experimental analyses of the function of the proepicardium using a new microsurgical procedure to induce loss-of-proepicardial-function in chick embryos. Dev. Dyn. 2005, 233, 1454–1463. [Google Scholar] [CrossRef] [PubMed]
- Perez-Pomares, J.M.; Phelps, A.; Sedmerova, M.; Wessels, A. Epicardial-like cells on the distal arterial end of the cardiac outflow tract do not derive from the proepicardium but are derivatives of the cephalic pericardium. Dev. Dyn. 2003, 227, 56–68. [Google Scholar] [CrossRef] [PubMed]
- Wilting, J.; Buttler, K.; Schulte, I.; Papoutsi, M.; Schweigerer, L.; Manner, J. The proepicardium delivers hemangioblasts but not lymphangioblasts to the developing heart. Dev. Biol. 2007, 305, 451–459. [Google Scholar] [CrossRef] [PubMed]
- Combs, M.D.; Braitsch, C.M.; Lange, A.W.; James, J.F.; Yutzey, K.E. Nfatc1 promotes epicardium-derived cell invasion into myocardium. Development 2011, 138, 1747–1757. [Google Scholar] [CrossRef] [PubMed]
- Schlueter, J.; Brand, T. Subpopulation of proepicardial cells is derived from the somatic mesoderm in the chick embryo. Circ. Res. 2013, 113, 1128–1137. [Google Scholar] [CrossRef] [PubMed]
- Ishii, Y.; Garriock, R.J.; Navetta, A.M.; Coughlin, L.E.; Mikawa, T. Bmp signals promote proepicardial protrusion necessary for recruitment of coronary vessel and epicardial progenitors to the heart. Dev. Cell 2010, 19, 307–316. [Google Scholar] [CrossRef] [PubMed]
- Hatcher, C.J.; Diman, N.Y.; Kim, M.S.; Pennisi, D.; Song, Y.; Goldstein, M.M.; Mikawa, T.; Basson, C.T. A role for tbx5 in proepicardial cell migration during cardiogenesis. Physiol. Genomics 2004, 18, 129–140. [Google Scholar] [CrossRef] [PubMed]
- Katz, T.C.; Singh, M.K.; Degenhardt, K.; Rivera-Feliciano, J.; Johnson, R.L.; Epstein, J.A.; Tabin, C.J. Distinct compartments of the proepicardial organ give rise to coronary vascular endothelial cells. Dev. Cell 2012, 22, 639–650. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wittig, J.G.; Münsterberg, A. The Early Stages of Heart Development: Insights from Chicken Embryos. J. Cardiovasc. Dev. Dis. 2016, 3, 12. https://doi.org/10.3390/jcdd3020012
Wittig JG, Münsterberg A. The Early Stages of Heart Development: Insights from Chicken Embryos. Journal of Cardiovascular Development and Disease. 2016; 3(2):12. https://doi.org/10.3390/jcdd3020012
Chicago/Turabian StyleWittig, Johannes G., and Andrea Münsterberg. 2016. "The Early Stages of Heart Development: Insights from Chicken Embryos" Journal of Cardiovascular Development and Disease 3, no. 2: 12. https://doi.org/10.3390/jcdd3020012
APA StyleWittig, J. G., & Münsterberg, A. (2016). The Early Stages of Heart Development: Insights from Chicken Embryos. Journal of Cardiovascular Development and Disease, 3(2), 12. https://doi.org/10.3390/jcdd3020012