ASIP Promoter Variants Predict the Sesame Coat Color in Shiba Inu Dogs

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples
2.2. Genotyping of ASIP Promoters
2.3. Sequencing the MC1R Gene
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kaelin, C.B.; Barsh, G.S. Genetics of pigmentation in dogs and cats. Annu. Rev. Anim. Biosci. 2013, 1, 125–156. [Google Scholar] [CrossRef] [PubMed]
- Schmutz, S.M.; Berryere, T.G. Genes affecting coat colour and pattern in domestic dogs: A review. Anim. Genet. 2007, 38, 539–549. [Google Scholar] [CrossRef] [PubMed]
- Ando, H.; Niki, Y.; Ito, M.; Akiyama, K.; Matsui, M.S.; Yarosh, D.B.; Ichihashi, M. Melanosomes are transferred from melanocytes to keratinocytes through the processes of packaging, release, uptake, and dispersion. J. Investig. Dermatol. 2012, 132, 1222–1229. [Google Scholar] [CrossRef] [Green Version]
- Tobin, D.J. Human hair pigmentation--biological aspects. Int. J. Cosmet. Sci. 2008, 30, 233–257. [Google Scholar] [CrossRef] [PubMed]
- Prota, G. Recent advances in the chemistry of melanogenesis in mammals. J. Investig. Dermatol. 1980, 75, 122–127. [Google Scholar] [CrossRef] [Green Version]
- Barsh, G.S. Regulation of Pigment Type Switching by Agouti, Melanocortin Signaling, Attractin, and Mahoganoid. In The Pigmentary System; Blackwell Publishing Ltd: Oxford, UK, 2006; pp. 395–409. [Google Scholar]
- Chandramohan, B.; Renieri, C.; La Manna, V.; La Terza, A. The alpaca agouti gene: Genomic locus, transcripts and causative mutations of eumelanic and pheomelanic coat color. Gene 2013, 521, 303–310. [Google Scholar] [CrossRef]
- Han, J.L.; Yang, M.; Yue, Y.J.; Guo, T.T.; Liu, J.B.; Niu, C.E.; Yang, B.H. Analysis of agouti signaling protein (ASIP) gene polymorphisms and association with coat color in Tibetan sheep (Ovis aries). Genet. Mol. Res. 2015, 14, 1200–1209. [Google Scholar] [CrossRef]
- Millar, S.E.; Miller, M.W.; Stevens, M.E.; Barsh, G.S. Expression and transgenic studies of the mouse agouti gene provide insight into the mechanisms by which mammalian coat color patterns are generated. Development 1995, 121, 3223–3232. [Google Scholar] [CrossRef]
- Ollmann, M.M.; Lamoreux, M.L.; Wilson, B.D.; Barsh, G.S. Interaction of Agouti protein with the melanocortin 1 receptor in vitro and in vivo. Genes Dev. 1998, 12, 316–330. [Google Scholar] [CrossRef] [Green Version]
- Lu, D.; Willard, D.; Patel, I.R.; Kadwell, S.; Overton, L.; Kost, T.; Luther, M.; Chen, W.; Woychik, R.P.; Wilkison, W.O.; et al. Agouti protein is an antagonist of the melanocyte-stimulating-hormone receptor. Nature 1994, 371, 799–802. [Google Scholar] [CrossRef]
- Vrieling, H.; Duhl, D.M.; Millar, S.E.; Miller, K.A.; Barsh, G.S. Differences in dorsal and ventral pigmentation result from regional expression of the mouse agouti gene. Proc. Natl. Acad. Sci. USA 1994, 91, 5667–5671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willis, M.B. Genetics of the Dog; Howell Book House: New York, NY, USA, 1989. [Google Scholar]
- Bannasch, D.L.; Kaelin, C.B.; Letko, A.; Loechel, R.; Hug, P.; Jagannathan, V.; Henkel, J.; Roosje, P.; Hytonen, M.K.; Lohi, H.; et al. Dog colour patterns explained by modular promoters of ancient canid origin. Nat. Ecol. Evol. 2021, 5, 1415–1423. [Google Scholar] [CrossRef] [PubMed]
- Berryere, T.G.; Kerns, J.A.; Barsh, G.S.; Schmutz, S.M. Association of an Agouti allele with fawn or sable coat color in domestic dogs. Mamm. Genome 2005, 16, 262–272. [Google Scholar] [CrossRef] [PubMed]
- Dreger, D.L.; Schmutz, S.M. A SINE insertion causes the black-and-tan and saddle tan phenotypes in domestic dogs. J. Hered. 2011, 102 (Suppl. 1), S11–S18. [Google Scholar] [CrossRef] [PubMed]
- Dreger, D.L.; Parker, H.G.; Ostrander, E.A.; Schmutz, S.M. Identification of a mutation that is associated with the saddle tan and black-and-tan phenotypes in Basset Hounds and Pembroke Welsh Corgis. J. Hered. 2013, 104, 399–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindblad-Toh, K.; Wade, C.M.; Mikkelsen, T.S.; Karlsson, E.K.; Jaffe, D.B.; Kamal, M.; Clamp, M.; Chang, J.L.; Kulbokas, E.J., 3rd; Zody, M.C.; et al. Genome sequence, comparative analysis and haplotype structure of the domestic dog. Nature 2005, 438, 803–819. [Google Scholar] [CrossRef]
- Dreger, D.L.; Anderson, H.; Donner, J.; Clark, J.A.; Dykstra, A.; Hughes, A.M.; Ekenstedt, K.J. Atypical Genotypes for Canine Agouti Signaling Protein Suggest Novel Chromosomal Rearrangement. Genes 2020, 11, 739. [Google Scholar] [CrossRef]
- Kerns, J.A.; Newton, J.; Berryere, T.G.; Rubin, E.M.; Cheng, J.F.; Schmutz, S.M.; Barsh, G.S. Characterization of the dog Agouti gene and a nonagoutimutation in German Shepherd Dogs. Mamm. Genome 2004, 15, 798–808. [Google Scholar] [CrossRef]
- Letko, A.; Ammann, B.; Jagannathan, V.; Henkel, J.; Leuthard, F.; Schelling, C.; Carneiro, M.; Drogemuller, C.; Leeb, T. A deletion spanning the promoter and first exon of the hair cycle-specific ASIP transcript isoform in black and tan rabbits. Anim. Genet. 2020, 51, 137–140. [Google Scholar] [CrossRef]
- Dreger, D.L.; Hooser, B.N.; Hughes, A.M.; Ganesan, B.; Donner, J.; Anderson, H.; Holtvoigt, L.; Ekenstedt, K.J. True Colors: Commercially-acquired morphological genotypes reveal hidden allele variation among dog breeds, informing both trait ancestry and breed potential. PLoS ONE 2019, 14, e0223995. [Google Scholar] [CrossRef] [Green Version]
- Shiba Inu Pedigree DataBase. Available online: http://www.shiba-pedigree.ru (accessed on 21 October 2021).
- Lis, J.T. Fractionation of DNA fragments by polyethylene glycol induced precipitation. Methods Enzymol. 1980, 65, 347–353. [Google Scholar] [CrossRef] [PubMed]
- Anderson, H.; Honkanen, L.; Ruotanen, P.; Mathlin, J.; Donner, J. Comprehensive genetic testing combined with citizen science reveals a recently characterized ancient MC1R mutation associated with partial recessive red phenotypes in dog. Canine Med. Genet. 2020, 7, 16. [Google Scholar] [CrossRef] [PubMed]
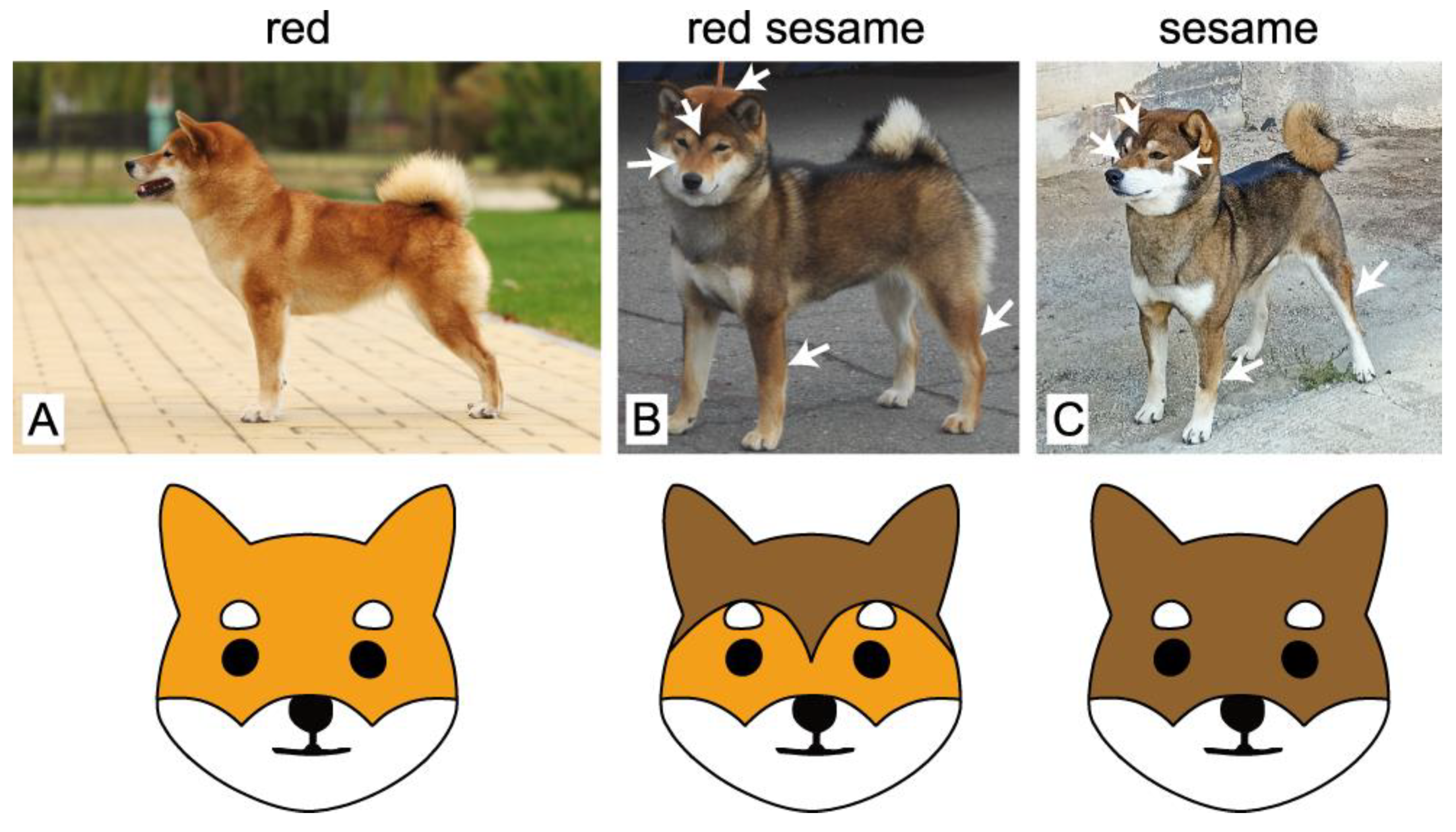




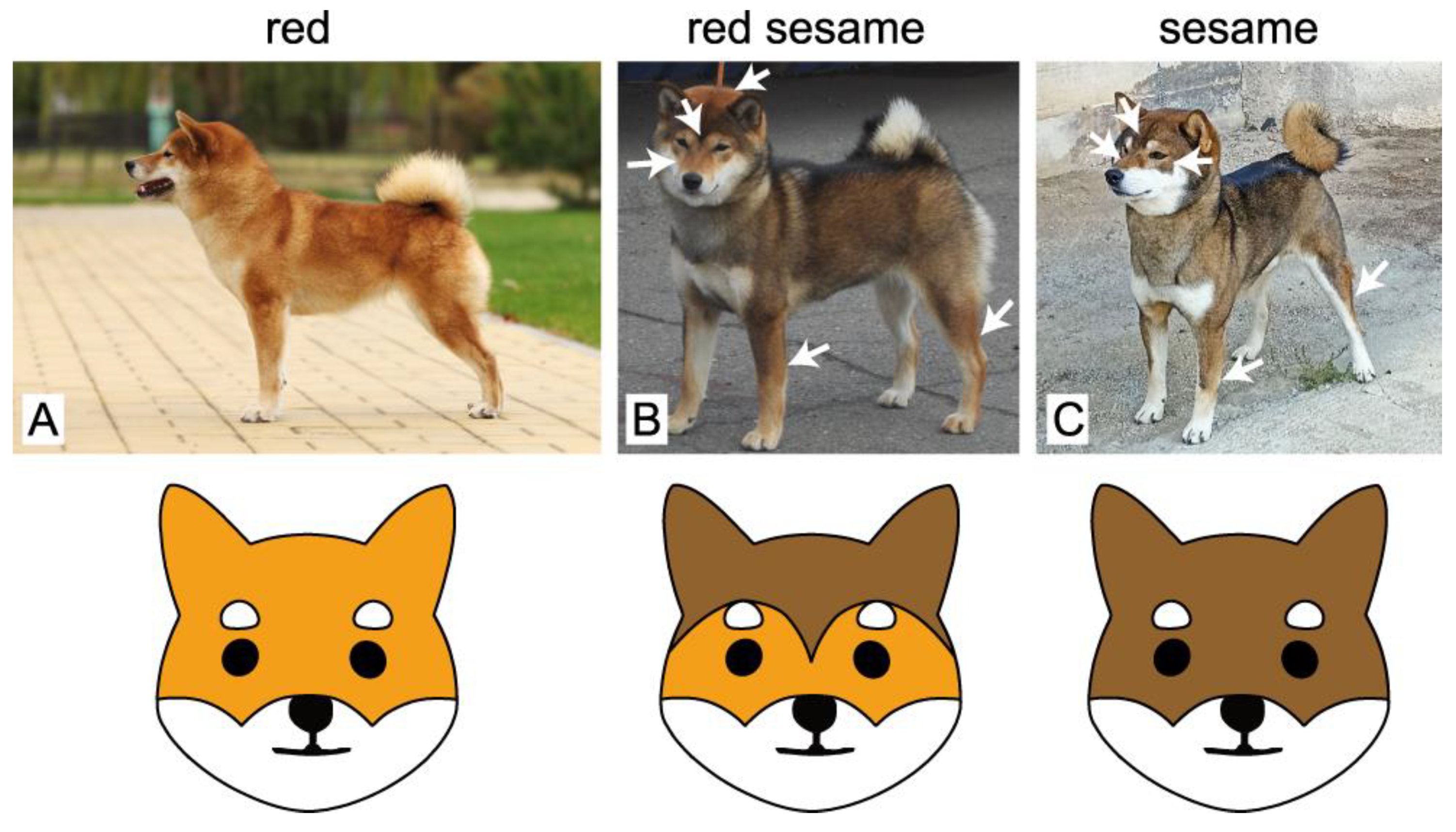{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Allele Name | Causative Promoter Haplotype & | Associated Genetic Variant # | Reference § |
|---|---|---|---|
| Dominant yellow (allele Ay) | DY: VP1-HCP1 | chr24:23393510T and chr24:23393514A | [15] |
| Shaded yellow (allele Ays) £ | SY: VP2-HCP1 | chr24:23393510T and chr24:23393514A | [15] |
| Wild type (allele aw) | AG: VP2-HCP2 | NA | |
| Saddle tan (allele asa) | BS: VP1-HCP4 | SINE insertion at chr24:23365297 | [16] |
| Black and tan (allele at) | BB: VP2-HCP3-5 | SINE insertion at chr24:23365297 and chr24:23252755-70dup | [16] [17] |
| Primer Pair * | Product Size (bp) in the ASIP Promoter Haplotypes | ||||||
|---|---|---|---|---|---|---|---|
| VP1 | VP2 | HCP1 | HCP2 | HCP3 | HCP4 | HCP5 | |
| 5′-AGCATGCTTATGTGGCATGT-3′ 5′-CGCTCTTTCAATGTGATTGG-3′ | 475 | 300 | - | - | - | - | - |
| 5′-TTAAAAAGTGAAAGTGAAAAGATAACCC-3′ 5′-CAAAATTCTGGGTGGGCTAA-3′ | - | - | 600 | 400 | 780 | - | - |
| 5′-TTAAAAAGTGAAAGTGAAAAGATAACCC-3′ 5′-TCAATGGAAATGGCAGAACA-3′ | - | - | - | - | - | 860 | - |
| 5′-GATTGAAAGCCAAAGGGTGA-3′ 5′-AGAGCAGGCCAGGTTTTACA-3′ | - | - | 300 | 300 | 300 | 300 | 400 |
| Coat Color | Ay/Ay | Ay/Ays | Ay/at | Ay/aw | Ays/at | aw/at |
|---|---|---|---|---|---|---|
| red | 26 | 7 | 6 | 3 | - | - |
| red sesame | - | - | - | - | 11 | - |
| sesame | - | - | - | - | - | 3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Belyakin, S.N.; Maksimov, D.A.; Pobedintseva, M.A.; Laktionov, P.P.; Voronova, D. ASIP Promoter Variants Predict the Sesame Coat Color in Shiba Inu Dogs. Vet. Sci. 2022, 9, 222. https://doi.org/10.3390/vetsci9050222
Belyakin SN, Maksimov DA, Pobedintseva MA, Laktionov PP, Voronova D. ASIP Promoter Variants Predict the Sesame Coat Color in Shiba Inu Dogs. Veterinary Sciences. 2022; 9(5):222. https://doi.org/10.3390/vetsci9050222
Chicago/Turabian StyleBelyakin, Stepan N., Daniil A. Maksimov, Maria A. Pobedintseva, Petr P. Laktionov, and Dinara Voronova. 2022. "ASIP Promoter Variants Predict the Sesame Coat Color in Shiba Inu Dogs" Veterinary Sciences 9, no. 5: 222. https://doi.org/10.3390/vetsci9050222
APA StyleBelyakin, S. N., Maksimov, D. A., Pobedintseva, M. A., Laktionov, P. P., & Voronova, D. (2022). ASIP Promoter Variants Predict the Sesame Coat Color in Shiba Inu Dogs. Veterinary Sciences, 9(5), 222. https://doi.org/10.3390/vetsci9050222