Protein Composition of the Bovine Herpesvirus 1.1 Virion

Abstract
1. Introduction
2. Materials and Methods
2.1. Cells and Virus
2.2. Virus Purification
2.3. Mass Spectrometry Sample Preparation
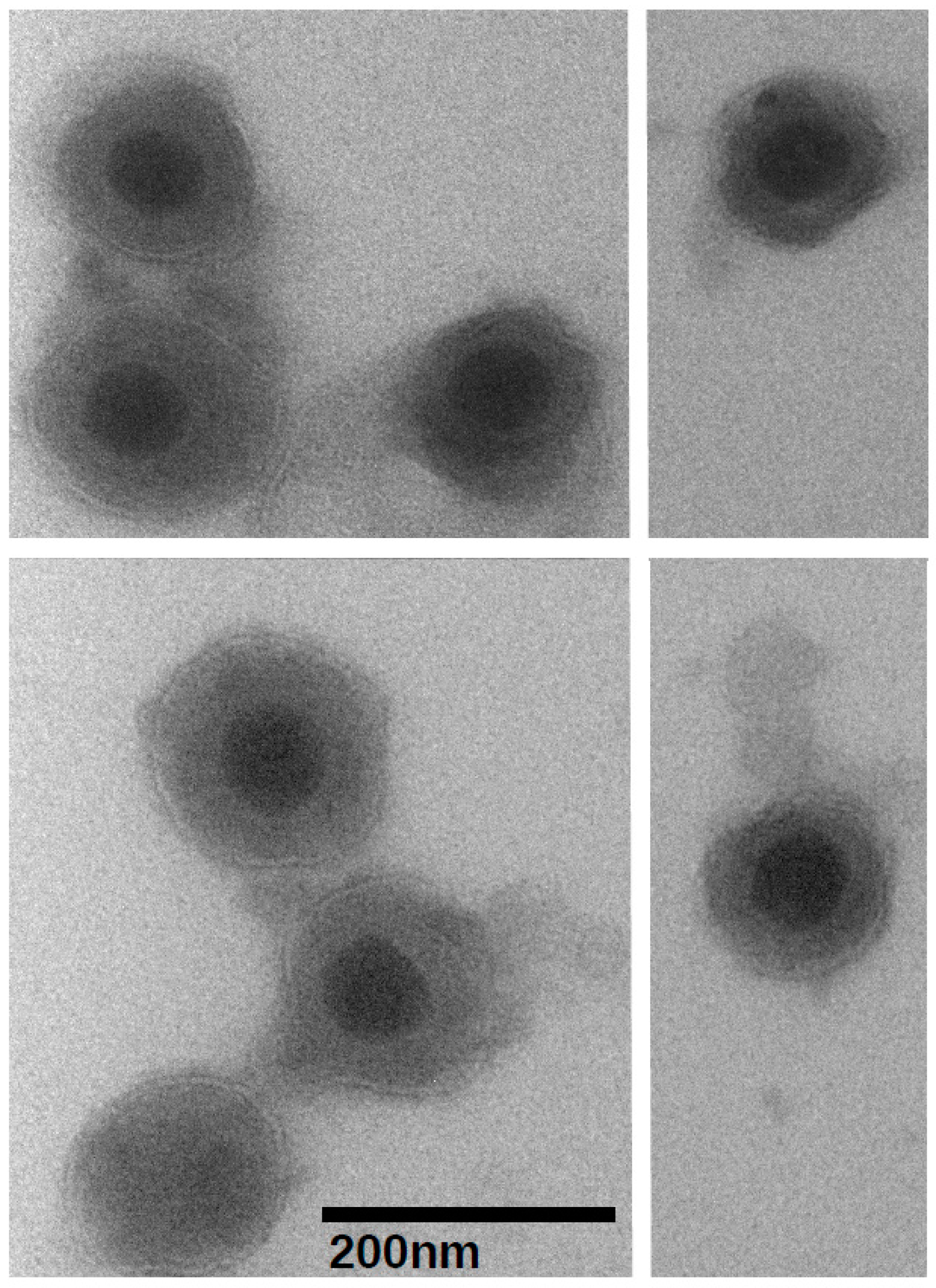
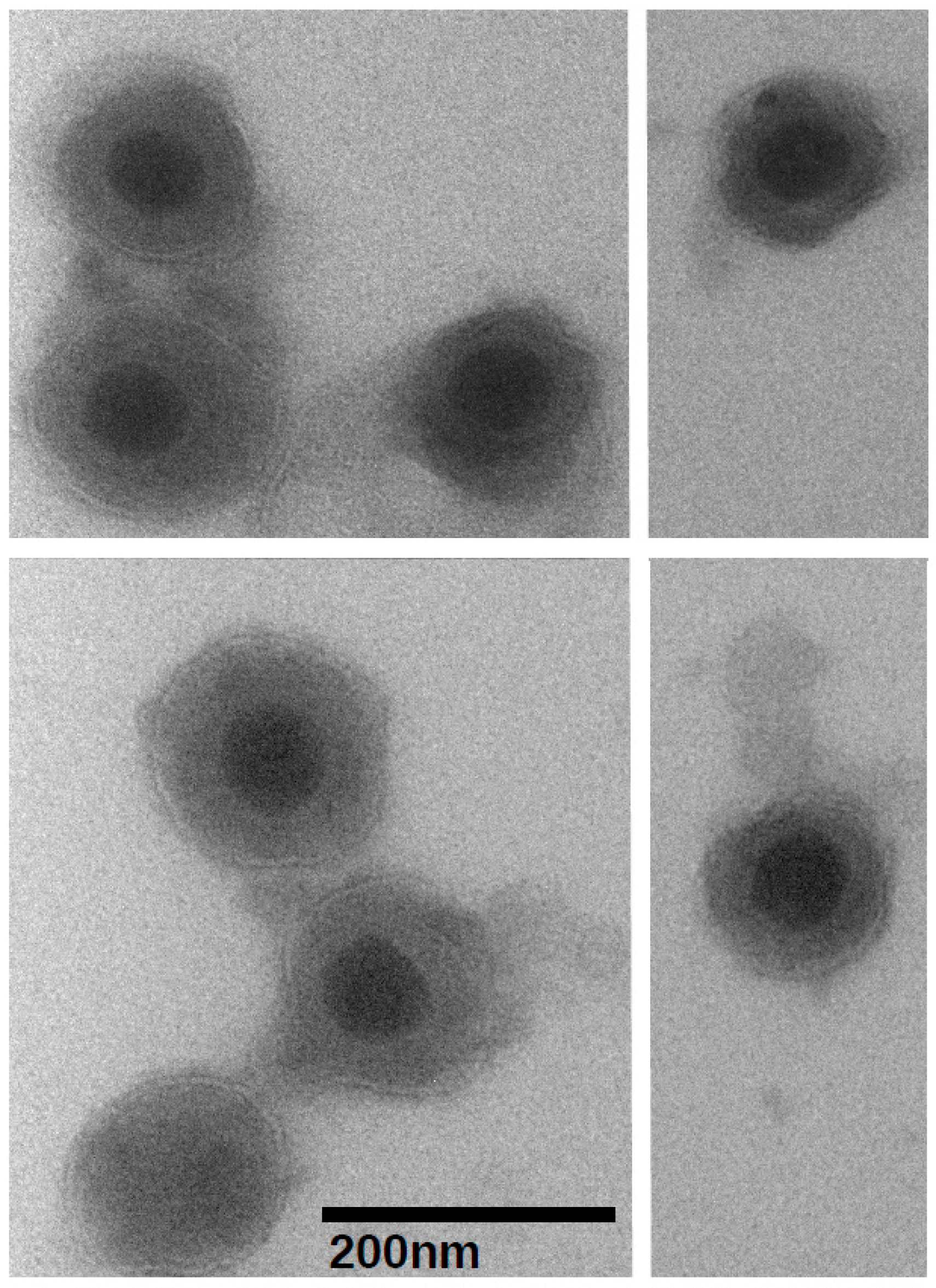
2.4. Transmission Electron Microscopy
2.5. Liquid Chromatography—Tandem Mass Spectrometry and Database Searches
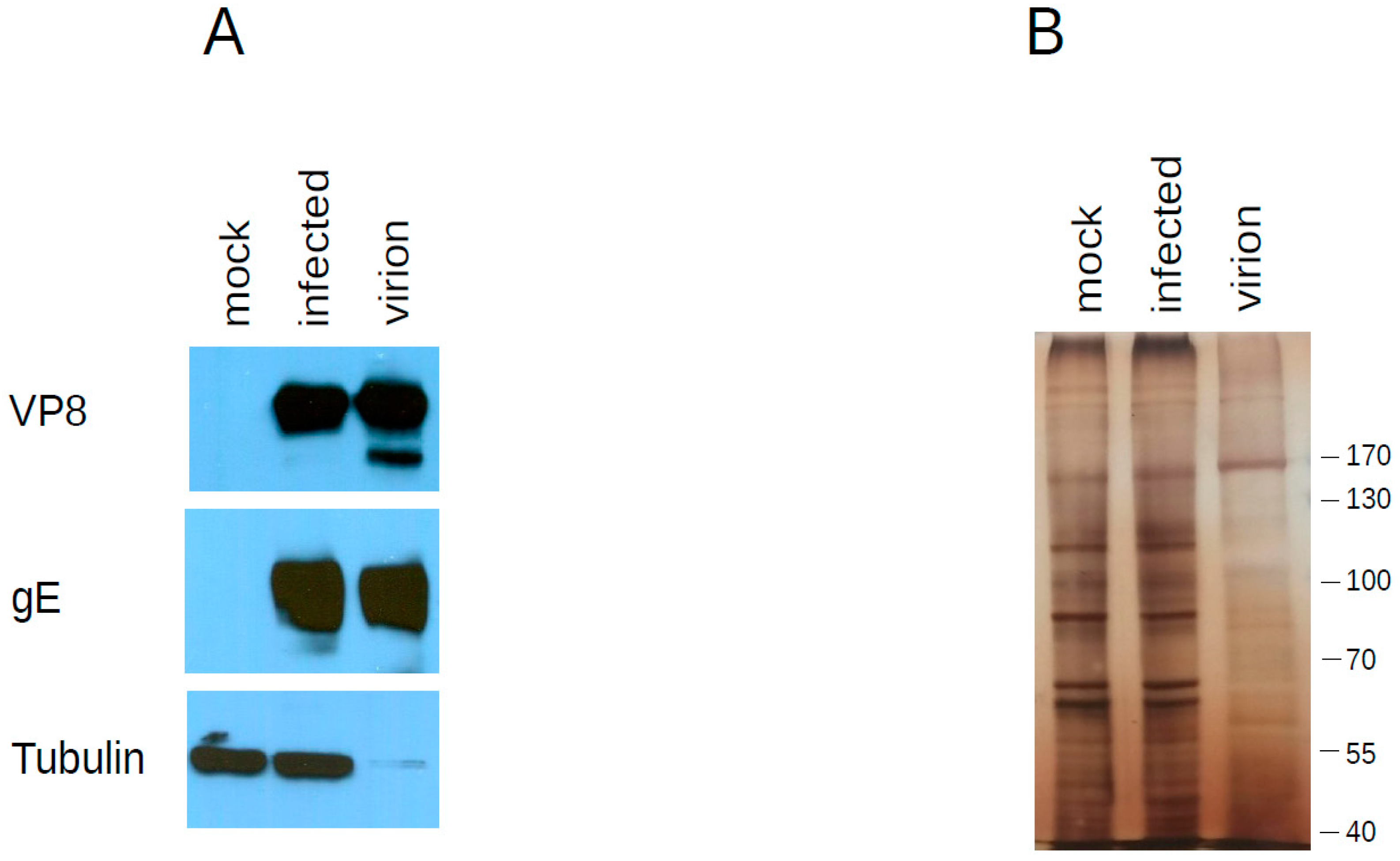
2.6. Protein Analysis (Western Blot and Silver Staining)
3. Results
3.1. Virion Purification and Imaging
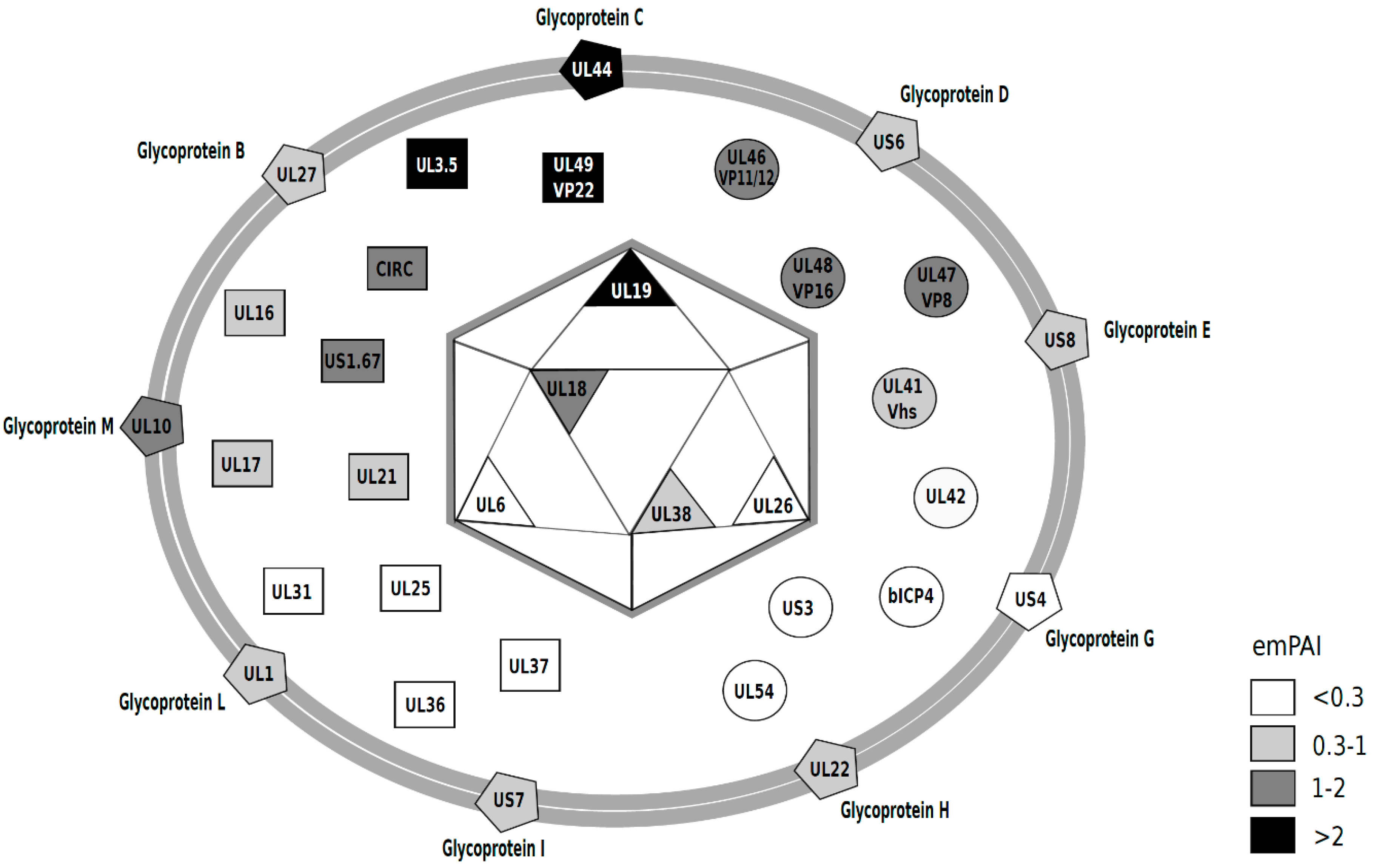
3.2. Viral Proteins Present in the Virion
3.3. Host Proteins Present in the Virion
4. Discussion
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Thiry, J.; Keuser, V.; Muylkens, B.; Meurens, F.; Gogev, S.; Vanderplasschen, A.; Thiry, E. Ruminant alphaherpesviruses related to bovine herpesvirus 1. Vet. Res. 2006, 37, 169–190. [Google Scholar] [CrossRef] [PubMed]
- Sausker, E.A.; Dyer, N.W. Seroprevalence of OHV-2, BVDV, BHV-1, and BRSV in ranch-raised bison (Bison bison). J. Vet. Diagn. Invest. 2002, 14, 68–70. [Google Scholar] [CrossRef] [PubMed]
- Scicluna, M.T.; Caprioli, A.; Saralli, G.; Manna, G.; Barone, A.; Cersini, A.; Cardeti, G.; Condoleo, R.U.; Autorino, G.L. Should the domestic buffalo (Bubalus bubalis) be considered in the epidemiology of Bovine Herpesvirus 1 infection? Vet. Microbiol. 2010, 143, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.M.; Whetstone, C.A.; Van der Maaten, M.J. Abortifacient property of bovine herpesvirus type 1 isolates that represent three subtypes determined by restriction endonuclease analysis of viral DNA. Am. J. Vet. Res. 1991, 52, 458–461. [Google Scholar] [PubMed]
- Metzler, A.E.; Matile, H.; Gassmann, U.; Engels, M.; Wyler, R. European isolates of bovine herpesvirus 1: A comparison of restriction endonuclease sites, polypeptides, and reactivity with monoclonal antibodies. Arch. Virol. 1985, 85, 57–69. [Google Scholar] [CrossRef] [PubMed]
- Edwards, S.; Newman, R.H.; White, H. The virulence of British isolates of bovid herpesvirus 1 in relationship to viral genotype. Br. Vet. J. 1991, 147, 216–231. [Google Scholar] [CrossRef]
- Jones, C. Regulation of Innate Immune Responses by Bovine Herpesvirus 1 and Infected Cell Protein 0 (bICP0). Viruses 2009, 1, 255–275. [Google Scholar] [CrossRef] [PubMed]
- Winkler, M.T.; Doster, A.; Jones, C. Bovine herpesvirus 1 can infect CD4(+) T lymphocytes and induce programmed cell death during acute infection of cattle. J. Virol. 1999, 73, 8657–8668. [Google Scholar] [PubMed]
- Hinkley, S.; Hill, A.B.; Srikumaran, S. Bovine herpesvirus-1 infection affects the peptide transport activity in bovine cells. Virus Res. 1998, 53, 91–96. [Google Scholar] [CrossRef]
- Nataraj, C.; Eidmann, S.; Hariharan, M.J.; Sur, J.H.; Perry, G.A.; Srikumaran, S. Bovine herpesvirus 1 downregulates the expression of bovine MHC class I molecules. Viral Immunol. 1997, 10, 21–34. [Google Scholar] [CrossRef] [PubMed]
- Koppers-Lalic, D.; Rijsewijk, F.A.M.; Verschuren, S.B.E.; van Gaans-van den Brink, J.A.M.; Neisig, A.; Ressing, M.E.; Neefjes, J.; Wiertz, E.J.H.J. The UL41-encoded virion host shutoff (vhs) protein and vhs- independent mechanisms are responsible for down-regulation of MHC class I molecules by bovine herpesvirus 1. J. Gen. Virol. 2001, 82, 2071–2081. [Google Scholar] [CrossRef] [PubMed]
- Henderson, G.; Zhang, Y.; Jones, C. The Bovine herpesvirus 1 gene encoding infected cell protein 0 (bICP0) can inhibit interferon-dependent transcription in the absence of other viral genes. J. Gen. Virol. 2005, 86, 2697–2702. [Google Scholar] [CrossRef] [PubMed]
- Saira, K.; Zhou, Y.; Jones, C. The infected cell protein 0 encoded by bovine herpesvirus 1 (bICP0) associates with interferon regulatory factor 7 and consequently inhibits beta interferon promoter activity. J. Virol. 2009, 83, 3977–3981. [Google Scholar] [CrossRef] [PubMed]
- Saira, K.; Zhou, Y.; Jones, C. The infected cell protein 0 encoded by bovine herpesvirus 1 (bICP0) induces degradation of interferon response factor 3 and, consequently, inhibits beta interferon promoter activity. J. Virol. 2007, 81, 3077–3086. [Google Scholar] [CrossRef] [PubMed]
- Frizzo da Silva, L.; Gaudreault, N.; Jones, C. Cytoplasmic localized infected cell protein 0 (bICP0) encoded by bovine herpesvirus 1 inhibits β interferon promoter activity and reduces IRF3 (interferon response factor 3) protein levels. Virus Res. 2011, 160, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, L.F.; Sinani, D.; Jones, C. ICP27 protein encoded by bovine herpesvirus type 1 (bICP27) interferes with promoter activity of the bovine genes encoding beta interferon 1 (IFN-β1) and IFN-β3. Virus Res. 2012, 169, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Griffin, D.; Chengappa, M.M.; Kuszak, J.; McVey, D.S. Bacterial pathogens of the bovine respiratory disease complex. Vet. Clin. North Am. Food Anim. Pract. 2010, 26, 381–394. [Google Scholar] [CrossRef] [PubMed]
- Jones, C.; Chowdhury, S. Bovine herpesvirus type 1 (BHV-1) is an important cofactor in the bovine respiratory disease complex. Vet. Clin. North Am. Food Anim. Pract. 2010, 26, 303–321. [Google Scholar] [CrossRef] [PubMed]
- Jones, C.; Chowdhury, S. A review of the biology of bovine herpesvirus type 1 (BHV-1), its role as a cofactor in the bovine respiratory disease complex and development of improved vaccines. Anim. Health Res. Rev. 2007, 8, 187–205. [Google Scholar] [CrossRef] [PubMed]
- Schneider, M.J.; Tait, R.G.; Busby, W.D.; Reecy, J.M. An evaluation of bovine respiratory disease complex in feedlot cattle: Impact on performance and carcass traits using treatment records and lung lesion scores. J. Anim. Sci. 2009, 87, 1821–1827. [Google Scholar] [CrossRef] [PubMed]
- Ackermann, M.; Engels, M. Pro and contra IBR-eradication. Vet. Microbiol. 2006, 113, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Raaperi, K.; Orro, T.; Viltrop, A. Epidemiology and control of bovine herpesvirus 1 infection in Europe. Vet. J. 2014, 201, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Guarino, H.; Núñez, A.; Repiso, M.V.; Gil, A.; Dargatz, D.A. Prevalence of serum antibodies to bovine herpesvirus-1 and bovine viral diarrhea virus in beef cattle in Uruguay. Prev. Vet. Med. 2008, 85, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Campos, F.S.; Franco, A.C.; Hübner, S.O.; Oliveira, M.T.; Silva, A.D.; Esteves, P.A.; Roehe, P.M.; Rijsewijk, F.A.M. High prevalence of co-infections with bovine herpesvirus 1 and 5 found in cattle in southern Brazil. Vet. Microbiol. 2009, 139, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Van Drunen Littel-van den Hurk, S. Rationale and perspectives on the success of vaccination against bovine herpesvirus-1. Vet. Microbiol. 2006, 113, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Nardelli, S.; Farina, G.; Lucchini, R.; Valorz, C.; Moresco, A.; Dal Zotto, R.; Costanzi, C. Dynamics of infection and immunity in a dairy cattle population undergoing an eradication programme for Infectious Bovine Rhinotracheitis (IBR). Prev. Vet. Med. 2008, 85, 68–80. [Google Scholar] [CrossRef] [PubMed]
- Engel, E.A.; Song, R.; Koyuncu, O.O.; Enquist, L.W. Investigating the biology of alpha herpesviruses with MS-based proteomics. Proteomics 2015, 15, 1943–1956. [Google Scholar] [CrossRef] [PubMed]
- Leroy, B.; Gillet, L.; Vanderplasschen, A.; Wattiez, R. Structural Proteomics of Herpesviruses. Viruses 2016, 8, 50. [Google Scholar] [CrossRef] [PubMed]
- Takáč, T.; Vadovič, P.; Pechan, T.; Luptovčiak, I.; Šamajová, O.; Šamaj, J. Comparative proteomic study of Arabidopsis mutants mpk4 and mpk6. Sci. Rep. 2016, 6, 28306. [Google Scholar] [CrossRef] [PubMed]
- Loret, S.; Guay, G.; Lippé, R. Comprehensive characterization of extracellular herpes simplex virus type 1 virions. J. Virol. 2008, 82, 8605–8618. [Google Scholar] [CrossRef] [PubMed]
- Kramer, T.; Greco, T.M.; Enquist, L.W.; Cristea, I.M. Proteomic characterization of pseudorabies virus extracellular virions. J. Virol. 2011, 85, 6427–6441. [Google Scholar] [CrossRef] [PubMed]
- McGeoch, D.J.; Dalrymple, M.A.; Davison, A.J.; Dolan, A.; Frame, M.C.; McNab, D.; Perry, L.J.; Scott, J.E.; Taylor, P. The Complete DNA Sequence of the Long Unique Region in the Genome of Herpes Simplex Virus Type 1. J. Gen. Virol. 1988, 69, 1531–1574. [Google Scholar] [CrossRef] [PubMed]
- McGeoch, D.J.; Dolan, A.; Donald, S.; Rixon, F.J. Sequence determination and genetic content of the short unique region in the genome of herpes simplex virus type 1. J. Mol. Biol. 1985, 181, 1–13. [Google Scholar] [CrossRef]
- Schwyzer, M.; Ackermann, M. Molecular virology of ruminant herpesviruses. Vet. Microbiol. 1996, 53, 17–29. [Google Scholar] [CrossRef]
- Berard, A.R.; Coombs, K.M.; Severini, A. Quantification of the Host Response Proteome after Herpes Simplex Virus Type 1 Infection. J. Proteome Res. 2015, 14, 2121–2142. [Google Scholar] [CrossRef] [PubMed]
- Traylen, C.; Ramasubramanyan, S.; Zuo, J.; Rowe, M.; Almohammad, R.; Heesom, K.; Sweet, S.; Matthews, D.; Sinclair, A. Identification of Epstein-Barr Virus Replication Proteins in Burkitt’s Lymphoma Cells. Pathogens 2015, 4, 739–751. [Google Scholar] [CrossRef] [PubMed]
- Coombs, K.M.; Berard, A.; Xu, W.; Krokhin, O.; Meng, X.; Cortens, J.P.; Kobasa, D.; Wilkins, J.; Brown, E.G. Quantitative proteomic analyses of influenza virus-infected cultured human lung cells. J. Virol. 2010, 84, 10888–10906. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Sadygov, R.G.; Yates, J.R. A model for random sampling and estimation of relative protein abundance in shotgun proteomics. Anal. Chem. 2004, 76, 4193–4201. [Google Scholar] [CrossRef] [PubMed]
- Ishihama, Y.; Oda, Y.; Tabata, T.; Sato, T.; Nagasu, T.; Rappsilber, J.; Mann, M. Exponentially modified protein abundance index (emPAI) for estimation of absolute protein amount in proteomics by the number of sequenced peptides per protein. Mol. Cell. Proteom. 2005, 4, 1265–1272. [Google Scholar] [CrossRef] [PubMed]
- Lété, C.; Palmeira, L.; Leroy, B.; Mast, J.; Machiels, B.; Wattiez, R.; Vanderplasschen, A.; Gillet, L. Proteomic characterization of bovine herpesvirus 4 extracellular virions. J. Virol. 2012, 86, 11567–11580. [Google Scholar] [CrossRef] [PubMed]
- Varnum, S.M.; Streblow, D.N.; Monroe, M.E.; Smith, P.; Auberry, K.J.; Pasa-Tolic, L.; Wang, D.; Camp, D.G.; Rodland, K.; Wiley, S.; et al. Identification of proteins in human cytomegalovirus (HCMV) particles: The HCMV proteome. J. Virol. 2004, 78, 10960–10966. [Google Scholar] [CrossRef] [PubMed]
- Baldick, C.J.; Shenk, T. Proteins associated with purified human cytomegalovirus particles. Proteins Associated with Purified Human Cytomegalovirus Particles. J. Virol. 1996, 70, 6097–6105. [Google Scholar] [PubMed]
- Kattenhorn, L.M.; Mills, R.; Wagner, M.; Lomsadze, A.; Makeev, V.; Borodovsky, M.; Ploegh, H.L.; Kessler, B.M. Identification of proteins associated with murine cytomegalovirus virions. J. Virol. 2004, 78, 11187–11197. [Google Scholar] [CrossRef] [PubMed]
- Vidick, S.; Leroy, B.; Palmeira, L.; Machiels, B.; Mast, J.; François, S.; Wattiez, R.; Vanderplasschen, A.; Gillet, L. Proteomic characterization of murid herpesvirus 4 extracellular virions. PLoS One 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Okazaki, K.; Matsuzaki, T.; Sugahara, Y.; Okada, J.; Hasebe, M.; Iwamura, Y.; Ohnishi, M.; Kanno, T.; Shimizu, M.; Honda, E. BHV-1 adsorption is mediated by the interaction of glycoprotein gIII with heparinlike moiety on the cell surface. Virology 1991, 181, 666–670. [Google Scholar] [CrossRef]
- Campadelli-Fiume, G.; Cocchi, F.; Menotti, L.; Lopez, M. The novel receptors that mediate the entry of herpes simplex viruses and animal alphaherpesviruses into cells. Rev. Med. Virol. 2000, 10, 305–319. [Google Scholar] [CrossRef]
- Connolly, S.A.; Whitbeck, J.J.; Rux, A.H.; Krummenacher, C.; van Drunen Littel-van den Hurk, S.; Cohen, G.H.; Eisenberg, R.J. Glycoprotein D homologs in herpes simplex virus type 1, pseudorabies virus, and bovine herpes virus type 1 bind directly to human HveC(nectin-1) with different affinities. Virology 2001, 280, 7–18. [Google Scholar] [CrossRef] [PubMed]
- Alves, L.D.; Pereira, A.A.F.; van Drunen Littel-van den Hurk, S. Bovine herpesvirus glycoprotein D: A review of its structural characteristics and applications in vaccinology. Vet. Res. 2014, 45, 111. [Google Scholar] [CrossRef] [PubMed]
- Baranowski, E.; Keil, G.; Lyaku, J.; Rijsewijk, F.A.M.; Van Oirschot, J.T.; Pastoret, P.P.; Thiry, E. Structural and functional analysis of bovine herpesvirus 1 minor glycoproteins. Vet. Microbiol. 1996, 53, 91–101. [Google Scholar] [CrossRef]
- Foster, T.P.; Melancon, J.M.; Olivier, T.L.; Kousoulas, K.G. Herpes simplex virus type 1 glycoprotein K and the UL20 protein are interdependent for intracellular trafficking and trans-Golgi network localization. J. Virol. 2004, 78, 13262–13277. [Google Scholar] [CrossRef] [PubMed]
- Melancon, J.M.; Fulmer, P.A.; Kousoulas, K.G. The herpes simplex virus UL20 protein functions in glycoprotein K (gK) intracellular transport and virus-induced cell fusion are independent of UL20 functions in cytoplasmic virion envelopment. Virol. J. 2007, 4. [Google Scholar] [CrossRef] [PubMed]
- Ward, P.L.; Campadelli-Fiume, G.; Avitabile, E.; Roizman, B. Localization and putative function of the UL20 membrane protein in cells infected with herpes simplex virus 1. J. Virol. 1994, 68, 7406–7417. [Google Scholar] [PubMed]
- Fuchs, W.; Klupp, B.G.; Granzow, H.; Mettenleiter, T.C. The UL20 gene product of pseudorabies virus functions in virus egress. J. Virol. 1997, 71, 5639–5646. [Google Scholar] [PubMed]
- Koppers-Lalic, D.; Reits, E.A.J.; Ressing, M.E.; Lipinska, A.D.; Abele, R.; Koch, J.; Marcondes Rezende, M.; Admiraal, P.; van Leeuwen, D.; Bienkowska-Szewczyk, K.; et al. Varicelloviruses avoid T cell recognition by UL49.5-mediated inactivation of the transporter associated with antigen processing. Proc. Natl. Acad. Sci. USA 2005, 102, 5144–5149. [Google Scholar] [CrossRef] [PubMed]
- Robinson, K.E.; Meers, J.; Gravel, J.L.; McCarthy, F.M.; Mahony, T.J. The essential and non-essential genes of Bovine herpesvirus 1. J. Gen. Virol. 2008, 89, 2851–2863. [Google Scholar] [CrossRef] [PubMed]
- Fraefel, C.; Ackermann, M.; Schwyzer, M. Identification of the bovine herpesvirus 1 circ protein, a myristylated and virion-associated polypeptide which is not essential for virus replication in cell culture. J. Virol. 1994, 68, 8082–8088. [Google Scholar] [PubMed]
- Ahn, B.C.; Kim, S.; Zhang, Y.; Charvat, R.A.; O’Callaghan, D.J. The early UL3 gene of equine herpesvirus-1 encodes a tegument protein not essential for replication or virulence in the mouse. Virology 2011, 420, 20–31. [Google Scholar] [CrossRef] [PubMed]
- Sato, H.; Pesnicak, L.; Cohen, J.I. Varicella-zoster virus open reading frame 2 encodes a membrane phosphoprotein that is dispensable for viral replication and for establishment of latency. J. Virol. 2002, 76, 3575–3578. [Google Scholar] [CrossRef] [PubMed]
- Osterrieder, N.; Neubauer, A.; Brandmüller, C.; Kaaden, O.R.; O’Callaghan, D.J. The equine herpesvirus 1 IR6 protein that colocalizes with nuclear lamins is involved in nucleocapsid egress and migrates from cell to cell independently of virus infection. J. Virol. 1998, 72, 9806–9817. [Google Scholar] [PubMed]
- Campos, F.S.; Paim, W.P.; Silva, A.G.; Santos, R.N.; Firpo, R.M.; Scheffer, C.M.; Finoketti, F.; Franco, A.C.; Roehe, P.M. A Spontaneous Deletion of the US1.67/US2 Genomic Region on the Bovine Herpesvirus 1 Strain Cooper. Genome Announc. 2016, 4. [Google Scholar] [CrossRef] [PubMed]
- Khattar, S.K.; van Drunen Littel-van den Hurk, S.; Babiuk, L.A.; Tikoo, S.K. Identification and transcriptional analysis of a 3’-coterminal gene cluster containing UL1, UL2, UL3, and UL3.5 open reading frames of bovine herpesvirus-1. Virology 1995, 213, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Schikora, B.; Lu, Z.; Kutish, G.F.; Rock, D.; Magyar, G.; Letchworth, G.J. The bovine herpesvirus type 1 UL3.5 open reading frame encodes a virion structural protein. Virology 1998, 240, 76–82. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Fuchs, W.; Mettenleiter, T.C. DNA sequence and transcriptional analysis of the UL1 to UL5 gene cluster of infectious laryngotracheitis virus. J. Gen. Virol. 1996, 77, 2221–2229. [Google Scholar] [CrossRef] [PubMed]
- Telford, E.A.; Watson, M.S.; McBride, K.; Davison, A.J. The DNA sequence of equine herpesvirus-1. Virology 1992, 189, 304–316. [Google Scholar] [CrossRef]
- Dean, H.J.; Cheung, A.K. A 3’ coterminal gene cluster in pseudorabies virus contains herpes simplex virus UL1, UL2, and UL3 gene homologs and a unique UL3.5 open reading frame. J. Virol. 1993, 67, 5955–5961. [Google Scholar] [PubMed]
- Davison, A.J.; Scott, J.E. The complete DNA sequence of varicella-zoster virus. J. Gen. Virol. 1986, 67, 1759–1816. [Google Scholar] [CrossRef] [PubMed]
- Lam, N.; Letchworth, G.J. Bovine herpesvirus 1 U(L)3.5 interacts with bovine herpesvirus 1 alpha-transinducing factor. J. Virol. 2000, 74, 2876–2884. [Google Scholar] [CrossRef] [PubMed]
- Lam, N.; Letchworth, G. A derivative of bovine herpesvirus 1 (BoHV-1) UL3.5 lacking the last forty amino acids inhibits replication of BoHV-1. Arch. Virol. 2004, 149, 2295–2306. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, W.; Klupp, B.G.; Granzow, H.; Rziha, H.J.; Mettenleiter, T.C. Identification and characterization of the pseudorabies virus UL3.5 protein, which is involved in virus egress. J. Virol. 1996, 70, 3517–3527. [Google Scholar] [PubMed]
- Fuchs, W.; Granzow, H.; Klupp, B.G.; Karger, A.; Michael, K.; Maresch, C.; Klopfleisch, R.; Mettenleiter, T.C. Relevance of the interaction between alphaherpesvirus UL3.5 and UL48 proteins for virion maturation and neuroinvasion. J. Virol. 2007, 81, 9307–9318. [Google Scholar] [CrossRef] [PubMed]
- Mar, E.C.; Patel, P.C.; Huang, E.S. Human cytomegalovirus-associated DNA polymerase and protein kinase activities. J. Gen. Virol. 1981, 57, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Lobanov, V.A.; Maher-Sturgess, S.L.; Snider, M.G.; Lawman, Z.; Babiuk, L.A.; van Drunen Littel-van den Hurk, S. A UL47 gene deletion mutant of bovine herpesvirus type 1 exhibits impaired growth in cell culture and lack of virulence in cattle. J. Virol. 2010, 84, 445–458. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Afroz, S.; Brownlie, R.; Snider, M.; van Drunen Littel-van den Hurk, S. Regulation and function of phosphorylation on VP8, the major tegument protein of bovine herpesvirus 1. J. Virol. 2015, 89, 4598–4611. [Google Scholar] [CrossRef] [PubMed]
- Labiuk, S.L.; Babiuk, L.A.; van Drunen Littel-van den Hurk, S. Major tegument protein VP8 of bovine herpesvirus 1 is phosphorylated by viral US3 and cellular CK2 protein kinases. J. Gen. Virol. 2009, 90, 2829–2839. [Google Scholar] [CrossRef] [PubMed]
- Michael, K.; Klupp, B.G.; Karger, A.; Mettenleiter, T.C. Efficient incorporation of tegument proteins pUL46, pUL49, and pUS3 into pseudorabies virus particles depends on the presence of pUL21. J. Virol. 2007, 81, 1048–1051. [Google Scholar] [CrossRef] [PubMed]
- Dry, I.; Haig, D.M.; Inglis, N.F.; Imrie, L.; Stewart, J.P.; Russell, G.C. Proteomic analysis of pathogenic and attenuated alcelaphine herpesvirus 1. J. Virol. 2008, 82, 5390–5397. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.; Fraser, N.W. Temporal association of the herpes simplex virus genome with histone proteins during a lytic infection. J. Virol. 2008, 82, 3530–3537. [Google Scholar] [CrossRef] [PubMed]
- Kutluay, S.B.; Triezenberg, S.J. Role of chromatin during herpesvirus infections. Biochim. Biophys. Acta. Gen. Subj. 2009, 1790, 456–466. [Google Scholar] [CrossRef] [PubMed]
- Kent, J.R.; Zeng, P.; Atanasiu, D.; Gardner, J.; Fraser, N.W.; Berger, S.L. During Lytic Infection Herpes Simplex Virus Type 1 Is Associated with Histones Bearing Modifications That Correla. J. Virol. 2004, 78, 10178–10186. [Google Scholar] [CrossRef]
- Bechtel, J.T.; Winant, R.C.; Ganem, D. Host and Viral Proteins in the Virion of Kaposi’s Sarcoma-associated Herpesvirus. J. Virol. 2005, 79, 4952–4964. [Google Scholar] [CrossRef] [PubMed]
- Michel, B.; Leroy, B.; Stalin Raj, V.; Lieffrig, F.; Mast, J.; Wattiez, R.; Vanderplasschen, A.F.; Costes, B. The genome of cyprinid herpesvirus 3 encodes 40 proteins incorporated in mature virions. J. Gen. Virol. 2010, 91, 452–462. [Google Scholar] [CrossRef] [PubMed]
- Sodeik, B.; Ebersold, M.W.; Helenius, A. Microtubule-mediated transport of incoming herpes simplex virus 1 capsids to the nucleus. J. Cell Biol. 1997, 136, 1007–1021. [Google Scholar] [CrossRef] [PubMed]
- Ogawa-Goto, K.; Tanaka, K.; Gibson, W.; Moriishi, E.; Miura, Y.; Kurata, T.; Irie, S.; Sata, T. Microtubule network facilitates nuclear targeting of human cytomegalovirus capsid. J. Virol. 2003, 77, 8541–8547. [Google Scholar] [CrossRef] [PubMed]
- Frampton, A.R.; Uchida, H.; von Einem, J.; Goins, W.F.; Grandi, P.; Cohen, J.B.; Osterrieder, N.; Glorioso, J.C. Equine herpesvirus type 1 (EHV-1) utilizes microtubules, dynein, and ROCK1 to productively infect cells. Vet. Microbiol. 2010, 141, 12–21. [Google Scholar] [CrossRef]
- Hammonds, T.R.; Denyer, S.P.; Jackson, D.E.; Irving, W.L. Studies to show that with podophyllotoxin the early replicative stages of herpes simplex virus type 1 depend upon functional cytoplasmic microtubules. J. Med. Microbiol. 1996, 45, 167–172. [Google Scholar] [CrossRef] [PubMed]
- Brzozowska, A.; Rychłowski, M.; Lipińska, A.D.; Bieńkowska-Szewczyk, K. Point mutations in BHV-1 Us3 gene abolish its ability to induce cytoskeletal changes in various cell types. Vet. Microbiol. 2010, 143, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Harms, J.S.; Ren, X.; Oliveira, S.C.; Splitter, G.A. Distinctions between bovine herpesvirus 1 and herpes simplex virus type 1 VP22 tegument protein subcellular associations. J. Virol. 2000, 74, 3301–3312. [Google Scholar] [CrossRef] [PubMed]
- Wolfstein, A.; Nagel, C.-H.; Radtke, K.; Döhner, K.; Allan, V.J.; Sodeik, B. The Inner Tegument Promotes Herpes Simplex Virus Capsid Motility along Microtubules in vitro. Traffic 2006, 7, 227–237. [Google Scholar] [CrossRef]
- Bresnahan, W.A.; Shenk, T. A subset of viral transcripts packaged within human cytomegalovirus particles. Science 2000, 288, 2373–2376. [Google Scholar] [CrossRef] [PubMed]
- Sciortino, M.T.; Suzuki, M.; Taddeo, B.; Roizman, B. RNAs extracted from herpes simplex virus 1 virions: Apparent selectivity of viral but not cellular RNAs packaged in virions. J. Virol. 2001, 75, 8105–8116. [Google Scholar] [CrossRef] [PubMed]
- Anderson, N.L.; Anderson, N.G. The Human Plasma Proteome. Mol. Cell. Proteom. 2002, 1, 845–867. [Google Scholar] [CrossRef] [PubMed]
- Steel, L.F.; Trotter, M.G.; Nakajima, P.B.; Mattu, T.S.; Gonye, G.; Block, T. Efficient and Specific Removal of Albumin from Human Serum Samples. Mol. Cell. Proteom. 2003, 2, 262–270. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.X.; Chong, J.M.; Wu, L.; Yuan, Y. Virion proteins of Kaposi’s sarcoma-associated herpesvirus. J. Virol. 2005, 79, 800–811. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
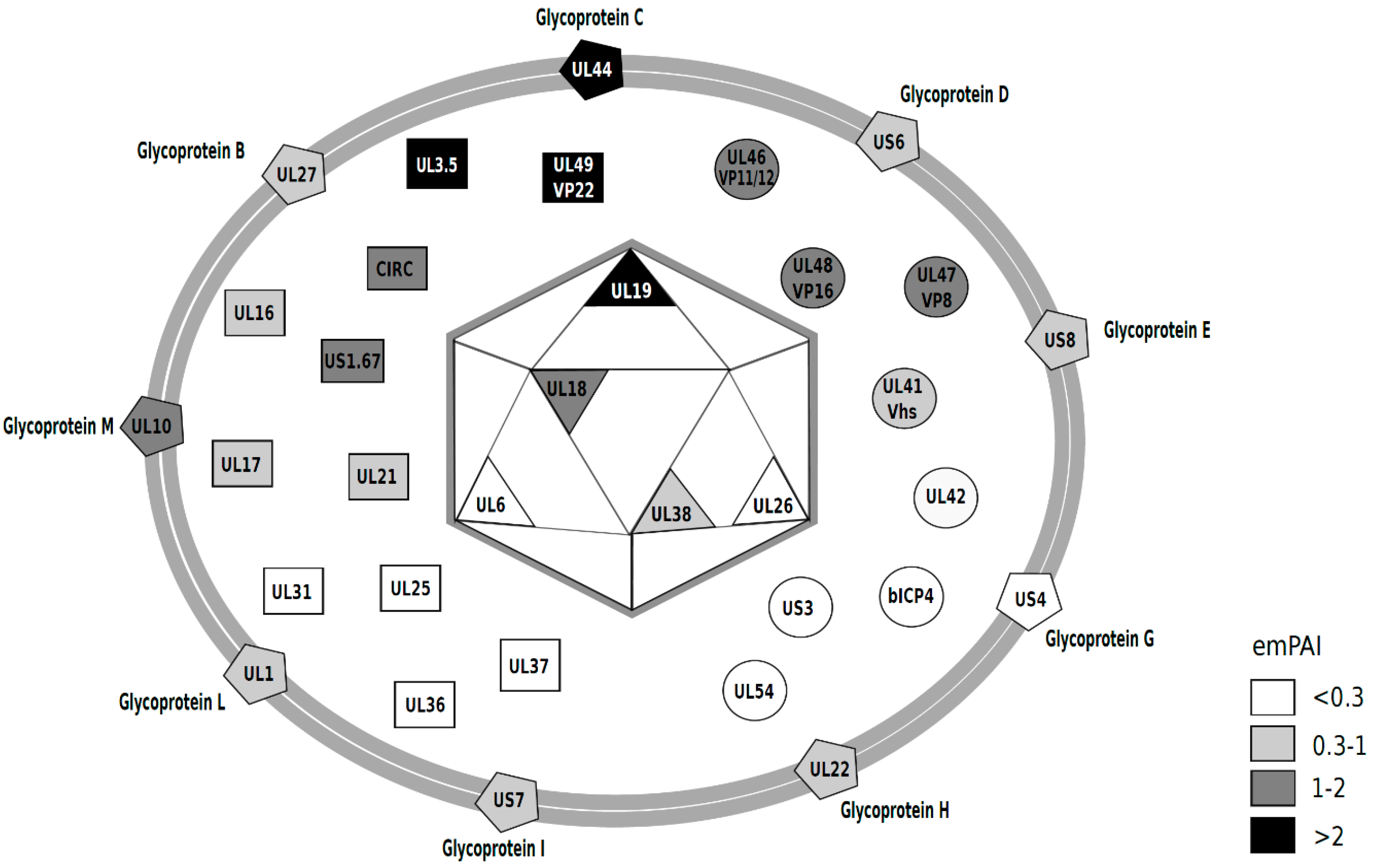{kind=link}
| BHV-1.1 Protein | Description/Alternative Name | Function | Size (aa) | MW (kDa) | # of Peptides a | # of Spectra a | # Unique Peptides a | % Coverage | emPAI | NCBI Accession |
|---|---|---|---|---|---|---|---|---|---|---|
| Capsid | ||||||||||
| UL6 | Capsid portal protein | DNA encapsidation | 692 | 75 | 4 | 12 | 4 | 6.4 | 0.245 | AFV53413 |
| UL18 | Capsid triplex subunit 2 | Capsid morphogenesis | 316 | 33.2 | 7 | 34 | 7 | 24.1 | 1.637 | AFV53401 |
| UL19 | Major capsid protein | Capsid morphogenesis | 1389 | 150 | 32 | 174 | 32 | 26.0 | 2.073 | AFV53400 |
| UL26 | Capsid scaffolding protein | Scaffold/serine protease | 622 | 63.7 | 2 | 11 | 2 | 4.5 | 0.274 | AFV53392 |
| UL38 | Capsid Triplex Subunit 1 | Capsid morphogenesis | 475 | 50 | 4 | 14 | 4 | 12.8 | 0.374 | AFV53380 |
| Envelope | ||||||||||
| UL1 | Glycoprotein L, gL | Cell entry/cell-to-cell spread | 158 | 17 | 3 | 12 | 3 | 17.7 | 0.874 | AFV53419 |
| UL10 | Glycoprotein M, gM | Virion morphogenesis/membrane fusion | 411 | 42.4 | 5 | 26 | 5 | 11.7 | 1.683 | AFV53409 |
| UL22 | Glycoprotein H, gH | Cell entry/cell-to-cell spread | 842 | 88.3 | 9 | 44 | 9 | 11.3 | 0.688 | AFV53397 |
| UL27 | Glycoprotein B, gB | Cell entry/cell-to-cell spread | 928 | 101.9 | 13 | 83 | 1 | 14.8 | 0.778 | AFV53391 |
| UL44 | Glycoprotein C, gC | Cell attachment | 521 | 55.4 | 13 | 113 | 13 | 29.9 | 3.786 | AFV53374 |
| US4 | Glycoprotein G, gG | Cell-to-cell spread | 444 | 46.6 | 1 | 9 | 1 | 3.4 | 0.116 | AFV53429 |
| US6 | Glycoprotein D, gD | Cell attachment | 417 | 44.9 | 3 | 35 | 3 | 9.4 | 0.551 | AFV53430 |
| US7 | Glycoprotein I, gI | Cell-to-cell spread | 382 | 39.6 | 3 | 12 | 3 | 9.4 | 0.389 | AFV53431 |
| US8 | Glycoprotein E, gE | Cell-to-cell spread | 575 | 61.2 | 3 | 22 | 3 | 7.3 | 0.413 | AFV53432 |
| Tegument | ||||||||||
| UL3.5 | protein V57 | Cell-to-cell spread | 126 | 13.4 | 6 | 77 | 6 | 56.3 | 6.499 | AFV53416 |
| Circ | Myristylated tegument protein | Unknown | 246 | 26.1 | 4 | 14 | 4 | 24.0 | 1.512 | AFV53363 |
| UL16 | Tegument protein UL16 | Possible virion morphogenesis | 343 | 36.4 | 6 | 29 | 6 | 20.1 | 0.823 | AFV53404 |
| UL17 | DNA Packaging Tegument Protein | DNA encapsidation/capsid transport | 703 | 72.6 | 7 | 33 | 7 | 11.7 | 0.455 | AFV53403 |
| UL21 | Tegument protein | Virion morphogenesis | 578 | 60.2 | 5 | 23 | 5 | 9.5 | 0.433 | AFV53398 |
| UL25 | DNA Packaging Tegument Protein | DNA encapsidatoin | 598 | 63 | 3 | 9 | 3 | 7.2 | 0.184 | AFV53394 |
| UL31 | Nuclear egress lamina protein | Nuclear egress | 376 | 40.9 | 1 | 2 | 1 | 2.1 | 0.116 | AFV53387 |
| UL36 | Large tegument protein | Capsid transport | 3291 | 336 | 10 | 47 | 10 | 3.4 | 0.147 | AFV53382 |
| UL37 | Tegument protein | Virion morphogenesis | 1034 | 106 | 7 | 22 | 7 | 8.1 | 0.254 | AFV53381 |
| UL49 | Tegument protein VP22 | Virion morphogenesis | 258 | 26.8 | 8 | 106 | 8 | 43.0 | 6.356 | AFV53370 |
| US1.67 | Unknown | 243 | 27.1 | 4 | 16 | 4 | 15.6 | 1.154 | AFV53426 | |
| Tegument (enzymatic and regulatory proteins) | ||||||||||
| bICP4 | IE transactivator protein | Transcriptional regulator | 1386 | 141 | 2 | 2 | 2 | 1.7 | 0.032 | AFV53424 |
| UL41 | Virion host shutoff protein, Vhs | Cellular mRNA degradation | 459 | 50 | 5 | 28 | 5 | 11.3 | 0.557 | AFV53377 |
| UL42 | DNA polymerase processivity subunit | DNA replication | 408 | 42.6 | 3 | 11 | 3 | 8.1 | 0.28 | AFV53376 |
| UL46 | Tegument protein VP11/12 | Possible gene regulation | 748 | 78.6 | 11 | 86 | 11 | 22.9 | 1.572 | AFV53373 |
| UL47 | Tegument protein VP8 | Possible gene regulation | 741 | 80.5 | 13 | 132 | 13 | 18.2 | 1.88 | AFV53372 |
| UL48 | Trans-inducing factor bTIF/VP16 | Gene regulation/virion morphogenesis | 507 | 53.1 | 6 | 42 | 6 | 10.1 | 1.154 | AFV53371 |
| UL54 | Multifunctional regulator, bICP27 | Gene regulation; RNA metobolism | 400 | 43.3 | 1 | 3 | 1 | 2.3 | 0.11 | AFV53364 |
| US3 | US3 kinase | serine/threonine kinase | 468 | 50.2 | 1 | 3 | 1 | 3.8 | 0.101 | AFV53428 |
| Host Protein | UniProt Accession | # of Positive Samples | # of Peptides a |
|---|---|---|---|
| Proteins detected in purified virions | |||
| Histone H4 | P62803 | 3 | 9,9,1 |
| Histone H2A.V | Q32LA7 | 2 | 2,1,0 |
| Histone H2B type 1 | P62808 | 2 | 1,1,0 |
| 60S ribosomal protein L7 | Q58DT1 | 2 | 0,1,1 |
| 60S ribosomal protein L8 | Q3T0S6 | 2 | 3,1,0 |
| 60S ribosomal protein L14 | Q3T0U2 | 2 | 3,1,0 |
| 40S ribosomal protein S6 | Q5E995 | 2 | 2,2,0 |
| Tubulin beta-5 chain | Q2KJD0 | 2 | 3,3,0 |
| Proteins that appeared in both virions and mock-infected preparations | |||
| Serotransferrin | Q29443 | 3 | 1,1,1 |
| Serum albumin | P02769 | 3 | 7,3,2 |
| Pancreatic trypsin inhibitor | P00974 | 3 | 5,6,3 |
| Alpha-2-HS-glycoprotein | P12763 | 3 | 1,8,1 |
| Annexin A2 * | P04272 | 2 | 1,2,0 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barber, K.A.; Daugherty, H.C.; Ander, S.E.; Jefferson, V.A.; Shack, L.A.; Pechan, T.; Nanduri, B.; Meyer, F. Protein Composition of the Bovine Herpesvirus 1.1 Virion. Vet. Sci. 2017, 4, 11. https://doi.org/10.3390/vetsci4010011
Barber KA, Daugherty HC, Ander SE, Jefferson VA, Shack LA, Pechan T, Nanduri B, Meyer F. Protein Composition of the Bovine Herpesvirus 1.1 Virion. Veterinary Sciences. 2017; 4(1):11. https://doi.org/10.3390/vetsci4010011
Chicago/Turabian StyleBarber, Kaley A., Hillary C. Daugherty, Stephanie E. Ander, Victoria A. Jefferson, Leslie A. Shack, Tibor Pechan, Bindu Nanduri, and Florencia Meyer. 2017. "Protein Composition of the Bovine Herpesvirus 1.1 Virion" Veterinary Sciences 4, no. 1: 11. https://doi.org/10.3390/vetsci4010011
APA StyleBarber, K. A., Daugherty, H. C., Ander, S. E., Jefferson, V. A., Shack, L. A., Pechan, T., Nanduri, B., & Meyer, F. (2017). Protein Composition of the Bovine Herpesvirus 1.1 Virion. Veterinary Sciences, 4(1), 11. https://doi.org/10.3390/vetsci4010011