Simple Summary
Giardia infections impact over 200 million people yearly, causing significant diarrheal disease. Wild animals, especially mammals and birds, host various Giardia types. Molecular tools have identified nine Giardia species, notably Giardia duodenalis, with eight genotypes. Birds are key Giardia carriers that spread the parasite. A global study targeting wild birds was performed to identify Giardia species using academic databases and precise search terms. While Giardia spans thirteen bird orders, it is genotyped in only seven orders. Different assemblages exist worldwide, with assemblage B linked to humans and many animals. Understanding G. duodenalis prevalence and its diverse assemblages in avian species is vital to assess zoonotic risks from the spread of this parasite.
Abstract
Globally, Giardia infections affect an estimated 280 million people each year, making it a significant cause of enteric diarrheal disease. Despite being categorized as a neglected disease, the prevalence of Giardia in wild animals varies, with both mammals and birds serving as hosts. Molecular tools have identified nine Giardia species, including Giardia duodenalis with eight genotypes. Birds, with their wide-ranging habitats and movement capabilities, are crucial disseminators of Giardia. This review aimed to identify Giardia species in wild birds globally, utilizing various academic databases with search terms such as “wild birds” and “Giardia” in combination with Boolean operators for comprehensive results. Giardia spp. have been identified in thirteen orders of wild birds; however, they have been genotyped in only seven orders. Giardia duodenalis assemblage A was identified in wild birds from the Americas; meanwhile, assemblages A, B, D, and F were identified from Europe, and B and E from Asia. Assemblage B is mainly associated with humans and a wide number of wild animals. Therefore, ascertaining the prevalence of G. duodenalis and its assemblages across diverse avian species and orders is crucial to assess the potential risk of zoonotic transmission associated with the dissemination of this protozoan.
1. Introduction
Giardia is an anaerobic flagellate protozoan that parasitizes a wide range of vertebrate hosts. Giardia species are characterized by their bilateral symmetry and the presence of two symmetrical nuclei in the trophozoites. The taxonomy of Giardia species has been debated for many years due to two problems: the first description in 1859 presented confusion with similar names, and later descriptions were based on the type of host rather than the parasite, leading to an excess of described species [1]. Strictly speaking, the first species described was Giardia duodenalis (previously known as intestinalis and lamblia), by Van Leeuwenhoek in 1681, a record that had been lost, and its first description had been attributed to Lamb in 1859 [2]. Subsequently, Giardia agilis was described in the intestine of amphibians by Kunstler in 1882 [3]. Historically, Giardia species were originally described using the morphology and biological data of the protozoan, and assuming a host specificity of every taxon, the latter of which is now known to be incorrect [3]. Subsequently, this classification was changed with the advance of molecular tools, and to date, nine species have been described within this genus: G. agilis; G. ardeae; G. psittaci; G. microti; G. muris; G. varani; G. duodenalis; and, recently, G. paramelis and G. cricetidarum [4].
Giardia duodenalis has eight genotypes according to an analysis of the sequences of several genes; these are known as assemblages, labelled from A to H [5]. Each assemblage consists of a large cluster in the phylogenetic tree, constructed from the sequences of every locus. Some of the assemblages have robust subdivisions in phylogenetic analyses, forming subclusters of well-supported sequences across multiple genetic loci, known as sub-assemblages. For example, within assemblage A, there are three common sub-assemblages, known as AI, AII, and AIII [6]. The genomes of the assemblages have been sequenced in recent years, revealing significant differences between them [7,8,9]. Assemblages A and B can infect non-human animals, as well as humans, with assemblage B being responsible for most infections, and some sub-assemblages present zoonotic potential [10]; AI and AIII are mainly found in animals, while AII is widely detected in humans [4,11]. Assemblage B is mainly associated with humans, livestock, and a wide number of wild animals (mainly mammals and birds), which raises public health concerns [12]. Meanwhile, assemblages C–H show higher host specificity [6]. Furthermore, wild animals can be infected with different G. duodenalis assemblages and sub-assemblages, making the determination of the role of wild species in the epidemiology of giardiasis difficult [6,13]. On the other hand, G. duodenalis assemblages C, D, E, and F have been sporadically isolated from humans; however, it is suggestive of potential zoonotic transmission [6,10].
- Life Cycle
The life cycle of Giardia is simple, including the proliferating trophozoite and the infective cyst stage [14]. Once the host ingests these cysts, these cysts reach the duodenum, where the release of trophozoites occurs. Then, these trophozoites replicate asexually through binary fission, and some of them differentiate into cysts, to later be disseminated through feces [15]. Giardia cysts are environmentally stable as they are tolerant to disinfectants and environmental degradation, and immediately infective [16]. In addition, Giardia cysts, which have four nuclei inside, have a low metabolic rate, which allows them to survive up to 12 weeks in cool and moist environments [17,18].
- Diagnosis of Giardia
The methods most commonly used for the diagnosis of Giardia in feces are light microscopy, immunodiagnostics, and molecular characterization. Until the year 2000, scanning electron microscopy (SEM) was the method of choice for distinguishing between Giardia species of mammalian and avian origin. This technique measured the length of the caudal flagellum, as well as symmetry and asymmetry, to differentiate among species. However, additional studies have raised doubts about the reliability of microscopy as a method for this distinction, as sample preparation and the age of trophozoite cultures can result in anomalies in the length of the caudal flagellum [19]. On the other hand, immunoassays continue to be important tools in the diagnosis of giardiasis with high sensitivity and specificity, but they do not provide the option to determine the species of the parasite as Polymerase Chain Reaction (PCR) assays do [3,5].
The molecular characterization of Giardia involves initial species classification, followed by genetic identification of their assemblage. To carry out this process, some specific loci serve as markers. The small ribosomal unit RNA (ssuRNA) gene and the elongation factor 1 (ef1a) gene are used to differentiate between species [20]. For assembly and sub-assembly identification, the most common approach is through multiloci genotyping (MLG), performed with the initial amplification of highly conserved genes, such as glutamate dehydrogenase (gdh), triose phosphate isomerase (tpi), and beta giardin (bg) [21]. Likewise, following PCR, restriction enzymes can be employed to assess polymorphisms within the amplified products [6]. The complete genome of the protozoan can also be sequenced from cysts and trophozoites obtained directly from fecal material [22].
- Importance of giardiasis in human, animal, and ecosystem health
Annually, about 280 million people worldwide suffer from symptomatic G. duodenalis infections [16,23]. Although Giardia infection is mainly associated with developing countries in Asia, Africa, and Latin America, the National Center for Emerging and Zoonotic Infectious Diseases (2014) reports giardiasis as the third leading cause of diarrheal disease from contaminated water in the United States. According to the Food and Agriculture Organization of the United Nations (FAO) and the World Health Organization (WHO), Giardia is the 11th most common cause of morbidity and mortality out of 24 foodborne parasites [24]. The full extent of the public health impact of this parasite is unclear due to variations in reporting and surveillance systems, differences in testing criteria, and different laboratory diagnostic methods [25].
Given that Giardia can be transmitted between wildlife and domestic animals and between humans and animals (zoonotic), gaining a deeper understanding of the present distribution patterns and possible transmission routes among wildlife is crucial for the health of humans, domestic animals, and wildlife alike [26].
In addition to the above considerations, the wide host range of Giardia plays an important role in the environmental dissemination of the parasite, contaminating food and water sources, contributing to maintaining the life cycle of the protozoan [6,10,21,27]. Two species of Giardia are responsible for causing avian giardiasis: G. ardeae and G. psittaci [10]. In addition to these two species, G. duodenalis has also been reported in wild birds, including the zoonotic assemblages A and B, as well as non-zoonotic assemblages D and F, and to be potentially zoonotic as assemblage E [12,28,29].
Birds have certain traits that facilitate the spread of pathogens, which could be more noticeable in birds adapted to anthropized landscapes. Some of these traits are the ability to travel long distances and inhabit aquatic and/or terrestrial ecosystems, and some birds may come into contact with human waste, for example, in landfills or household garbage; transit through areas near animal production farms; and be in environments highly contaminated by waste from other animal species, for example, wetlands and green areas contaminated with canine and feline feces [30]. Additionally, the transmission cycle can be maintained among wild species inhabiting the same ecosystem, promoting the parasite’s persistence in the environment and in hosts [31].
Giardia is a waterborne disease for which infection rates and distribution are projected to increase in response to global climate change scenarios [30]. Therefore, this protozoan has been identified as a priority for One Health, as it is a zoonotic disease and little is known about its prevalence in wild animals [32].
- Effects of Climate Change on the Distribution of Giardia and its Transmission
Several studies have focused on evaluating the prevalence, risk factors, and variables that influence the global occurrence of parasites such as Giardia [33]. Climate change can trigger ecological disruptions that alter the transmission mechanisms of pathogens. This phenomenon, known as the intersection of climate change with transmission dynamics or ecological fitting, may enable the emergence of parasites and diseases without requiring evolutionary changes in their capacity to utilize hosts [34]. This implies that parasites may colonize new hosts or increase prevalence, morbidity, and mortality in already known hosts. Climate change also promotes the movement and migration of wild species, carrying with them the pathogenic organisms that infect them [35]. For example, the northward expansion of Giardia from subarctic to Arctic environments has been documented in a variety of migratory and resident species [36,37].
Several studies have reported positive associations between environmental variables—many linked to the hydrological cycle, such as increased precipitation and temperature, permafrost melting, and coastal flooding—and the occurrence of giardiasis in humans and animals [31,38]. However, these variables may either promote the presence of Giardia or, conversely, reduce its prevalence depending on geographic location [31]. For instance, one study documented a consistent seasonal rise in giardiasis cases in temperate, developed countries during the summer months, possibly because higher temperatures prolong the persistence of cysts and oocysts in the environment [39,40]. Evidence suggests that the protozoan pathogen Giardia is particularly sensitive to environmental changes due to its environmentally mediated life cycle [39]. Consequently, further studies are needed to support epidemiological surveillance of this parasite in the context of climate change, as the full impacts of these changes—especially under extreme events—remain difficult to predict or project [38].
Birds play a key role as carriers of Giardia and other parasitic diseases, and their interaction with climate change is a major concern. Rising temperatures and the expansion of geographic areas favorable to the survival and transmission of Giardia may allow migratory birds to transport and disperse the parasite into new regions. Moreover, shifts in bird migration and movement in response to climate change could enhance the spread of Giardia along different migratory routes. These changes in bird migration patterns and distributions, combined with climate-induced ecosystem disruptions, may significantly influence the prevalence and incidence of Giardia across regions. This highlights the urgent need to investigate and monitor the interplay between Giardia-carrying birds and climate change in order to mitigate risks to both public and environmental health.
The aim of this literature review was to identify the presence of Giardia species and assemblages among various orders of wild birds worldwide, drawing upon the published scientific literature.
2. Materials and Methods
A comprehensive literature review was conducted from original scientific articles and case reports spanning the period from 1990 to March of 2024. This review focused on extracting information not only on the species of parasitized wild birds but also on the geographic regions or areas where Giardia occurrences have been documented.
A bibliographic search was performed using the following search engines and information sources: Google Scholar, ProQuest, PubMed, ScienceDirect, and Scopus. The search was performed using the keywords wild birds, Giardia, assemblage, avian, and birds, with the Boolean operators ‘’AND’’ and “OR”. Only scientific papers published in English were considered. Among these publications, original articles, short communications, and case reports were included. Reports of Giardia in domestic birds and other domestic or wild animals were not considered. Regardless of the article type, the selected references were given equal weight, provided they met the established inclusion and exclusion criteria.
A first search with the keywords without Boolean operators yielded a total of 14,200 results. Then, a search was performed using the Boolean operators separately with the keywords. The titles of the first 100 scientific articles were reviewed, and those unrelated to the research interests were excluded. Subsequently, selection was made by reading the abstract to determine whether the study presented results on Giardia positivity in wild birds (Figure 1).
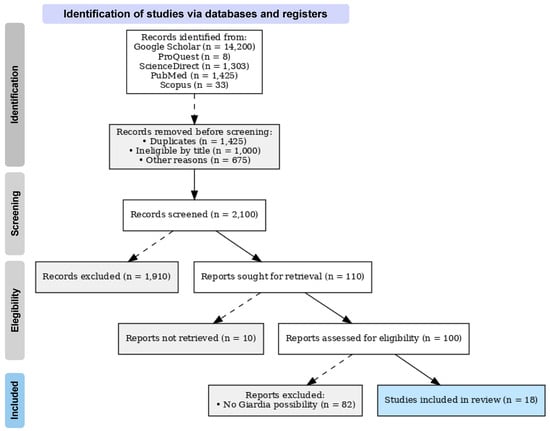
Figure 1.
Prisma flow diagram of the papers selected for this review. This diagram explains how the search was conducted and the academic articles selected for the review according to the inclusion and exclusion criteria.
To complete the bibliographic search, the Connected Papers tool (https://www.connectedpapers.com/ (accessed on 10 September 2024)) was used to identify the most important authors related to the topic of interest and to refine the bibliographic search. In this search, 41 bibliographic references were identified, which were also reviewed considering the inclusion and exclusion criteria (Figure S1). After the screening, 7 references were considered, which coincide with the initial search. The list of references identified by Connected Papers can be found in the Supplementary Materials.
In this search, the study by Reboredo-Fernandez et al. [28] was used as the base reference, since the publication contained more information on Giardia in wild birds (Figure S1). From this reference, a cluster map was constructed, showing the relationships between the main authors on the topic of interest (https://www.connectedpapers.com/main/45bf46e5aad407c2d5ca9a22cea672b863ccaa66/graph?utm_source=share_popup&utm_medium=email&utm_campaign=share_graph (accessed on 17 June 2025)).
The search was filtered, and inconclusive studies on Giardia positivity were excluded. Finally, 18 scientific articles provided data related to the variables of interest.
The variables analyzed to achieve the general objectives of this study were as follows: order, bird species, Giardia species or assemblage, country, year, and author. A descriptive analysis was performed to summarize the information regarding the variables. The graph presented in this article illustrates the distribution of reports by geographic area (country), along with the predominant avian order represented in each area.
3. Results
The analysis of the scientific literature resulted in the identification of Giardia spp. in 32 species and 13 orders of wild birds. According to the published literature, birds belonging to the order Anseriformes are the most frequently reported hosts worldwide (seven reports in six countries) and have the highest number of infected species (twelve), followed by Charadriiformes and Psittaciformes (two reports and three host species each (Figure 2a, Table 1).
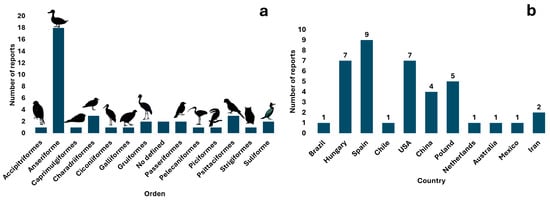
Figure 2.
Graphical representation of the number of reports identified by order of wild birds (a) and by country (b).

Table 1.
Information obtained from the bibliographic search ordered by the variables under study.
These reports were found mainly in Europe (twenty-one reports) (six references and seventeen host species) and North America (eight reports) (four references and eight host species). In the case of G. duodenalis and its genotyping, assemblage A is reported in America (Brazil); assemblages A, B, D, and F in Europe (Spain and Hungary); and assemblages B and E in Asia (China). Spain has the most orders of birds carrying identified G. duodenalis (four orders: Anseriformes, Accipitriformes, Galliformes, and Passeriformes), from which G. duodenalis has been genotyped, followed by China (three orders: Anseriformes, Charadriiformes, and Suliformes) (Figure 2b and Figure 3). According to the literature search and bibliometric analysis, the first genotyping study of G. duodenalis was conducted by Plutzer and Tomo in 2009 (see Supplementary Figure S1). A list of published studies on Giardia in wild birds is provided in Table 1. Two studies reported the presence of Giardia in waterfowl feces but did not define the species of birds that were carriers or the orders to which they belong: Cano et al. [30] identified G. duodenalis assemblage B IV, and Shemshadi et al. [51] reported G. duodenalis. A list of published studies on Giardia in wild birds is provided in Table 1.
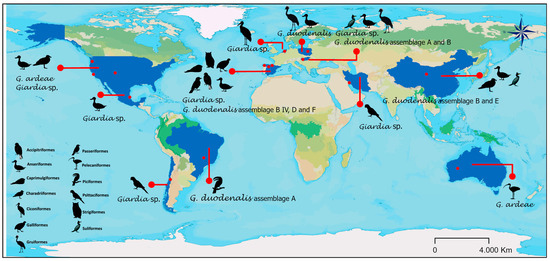
Figure 3.
Global distribution of Giardia species and genotypes of Giardia duodenalis in wild birds.
4. Discussion
This study offers an analysis of the global presence of Giardia in wild birds, delineating the species and orders harboring the protozoan and highlighting studies where genotyping of G. duodenalis has been accomplished. Birds serve as sentinels, and detecting pathogens within this taxon holds significant importance for epidemiological surveillance [32]. For G. ardeae, the findings are limited to two bird species, with only two studies reported up to the search date [42,45]. However, these studies date back to the nineties, and recent studies have not reported the presence of G. ardeae. This may be due to the fact that initially, Giardia species were differentiated microscopically based on the difference in the length of the caudal flagellum. However, subsequent studies have confirmed that this is not an appropriate technique for discriminating between Giardia species [19,42].
On the other hand, wild birds harboring Giardia spp. have been reported in Asia, Europe, North America, South America, and Oceania, with highly variable prevalences, ranging from 3% to 49% [26,32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52]. Most bird orders reported as carriers of Giardia share the commonality that they are marine or freshwater species. Several studies agree that both wild and captive birds carrying this protozoan are waterfowl [26,32,44,50]. This finding coincides with those of previous reports indicating the contamination of surface water with pathogens probably of avian origin, such as Giardia, that have been found in the water source [52].
Prevalences vary by continent, but it is important to consider that these data are specific to some countries and primarily focused on birds of the Anseriformes order. However, Giardia has also been identified in birds of other orders and families, which may be exposed to carrying the parasite. In this context, further studies are needed to gather evidence of Giardia presence in other bird species, establish prevalences, and assess risk factors that may be associated with Giardia presence in avian species.
Wild birds share their habitat with other domestic or exotic bird species and mammals, which could potentially act as hosts for Giardia [53,54]. In the case of invasive birds, such as pigeons, which have a wide worldwide distribution and may be free-ranging or kept as pets, the prevalences of Giardia spp. could reach up to 52% [55]. Unfortunately, not all reports identify the Giardia species present in pigeons, but one study identified G. duodenalis assemblage E, which is considered the most representative genotype among ruminants and has a zoonotic potential. Additionally, this assemblage has been identified in other wild birds in the Anseriformes, Charadriiformes, and Suliformes orders [56,57]. However, studies genotyping Giardia in birds to determine if other potentially zoonotic assemblages are present or to establish which assemblage may be dominant in birds are lacking [58].
To date, molecular studies support the zoonotic potential of some G. duodenalis assemblages, but further studies are required to understand the infection dynamics of the protozoan in different animal species and humans. As highlighted in this literature review, most genotyping studies have been conducted in the European continent, leaving a gap in our understanding of what may be occurring in continents such as Africa and the Americas, particularly given the abundance and richness of bird species. For example, in South America, all reported cases are restricted to Brazil (G. duodenalis assemblage A) [27]. It highlights the need for research to explore how various species act as reservoirs for G. duodenalis, considering human, animal, and ecosystem health. This research would help clarify species interactions and the impact of human activities on the protozoan’s ecology and suggest ways to prevent and control related diseases [59,60]. A recent study that analyzed reports of giardiasis in Latin America concluded that it still represents a neglected public health problem [61].
Wildlife, such as wild birds, can act as hosts for these pathogens, aiding in their transmission among themselves, humans, and the environment, underscoring the complex One Health issue presented by these zoonotic agents [61]. Efforts should focus on improving water, sanitation, and hygiene practices to reduce disease transmission. Additionally, addressing the impact of climatic variability, land-use changes, and social factors in giardiasis is crucial, emphasizing the need for global public health interventions to prevent future disease burden, particularly in regions with limited resources [41]. Therefore, adopting a One Health approach is essential for addressing zoonotic diseases and mitigating the challenges posed by this pathogen.
5. Conclusions
In this literature review, we gathered data on the presence of Giardia and its assemblages across various orders of wild birds worldwide. While Giardia has predominantly been identified in Anseriformes, molecular characterization has largely been restricted to birds from European countries. Therefore, an investigation of the circulation of Giardia assemblages and an assessment of whether birds could serve as potential disseminators of this protozoan, as well as the implications for zoonotic risk, are essential.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/vetsci12090911/s1, Figure S1: Graphical summary of the main authors reporting on Giardia and Giardia species in birds, created using the Connected Papers tool. Supplementary reference: Include all reference from Connected Papers search. Supplementary references [62,63,64,65,66,67,68,69,70,71,72,73,74,75,76,77,78,79,80,81,82,83,84,85,86,87,88,89,90,91,92,93,94].
Author Contributions
Conceptualization, D.E.; methodology, D.E.; validation, D.E., formal analysis, D.E.; investigation, D.E., P.O.-R. and C.L.-A.; resources, D.E.; writing—original draft preparation, D.E.; writing—review and editing, D.E., P.O.-R. and C.L.-A. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Agencia Nacional de Investigación y Desarrollo (ANID) through the Fondecyt Initiation Project grant number 11240245.
Institutional Review Board Statement
This study was conducted in accordance with the Declaration of Helsinki and approved by the Ethics Committee of Universidad San Sebastián (protocol code 05-24, 3 April 2024).
Data Availability Statement
The original contributions presented in this study are included in the article/Supplementary Materials. Further inquiries can be directed to the corresponding author.
Acknowledgments
The authors would like to thank Felipe Orellana and Victor Aguilar for the map design, as well as Vicerrectoría de Investigación y Doctorados VRID from Universidad San Sebastián.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of this study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Wielinga, C.; Williams, A.; Monis, P.; Thompson, R.C.A. Proposed Taxonomic Revision of Giardia duodenalis. Infect. Genet. Evol. 2023, 111, 105430. [Google Scholar] [CrossRef]
- Ortega, Y.R.; Adam, R.D. Giardia: Overview and Update. Clin. Infect. Dis. 1997, 25, 545–549. [Google Scholar] [CrossRef]
- Ryan, U.M.; Feng, Y.; Fayer, R.; Xiao, L. Taxonomy and Molecular Epidemiology of Cryptosporidium and Giardia—A 50 Year Perspective (1971–2021). Int. J. Parasitol. 2021, 51, 1099–1119. [Google Scholar] [CrossRef]
- Adam, R.D. Giardia duodenalis: Biology and Pathogenesis. Clin. Microbiol. Rev. 2021, 34, e00024-19. [Google Scholar] [CrossRef]
- Ryan, U.; Zahedi, A. Molecular Epidemiology of Giardiasis from a Veterinary Perspective. In Advances in Parasitology; Academic Press: Cambridge, MA, USA, 2019; Volume 106, pp. 209–254. ISBN 9780128177204. [Google Scholar]
- Feng, Y.; Xiao, L. Zoonotic Potential and Molecular Epidemiology of Giardia Species and Giardiasis. Clin. Microbiol. Rev. 2011, 24, 110–140. [Google Scholar] [CrossRef]
- Morrison, H.G.; McArthur, A.G.; Gillin, F.D.; Aley, S.B.; Adam, R.D.; Olsen, G.J.; Best, A.A.; Cande, W.Z.; Chen, F.; Cipriano, M.J.; et al. Genomic Minimalism in the Early Diverging Intestinal Parasite Giardia lamblia. Science 2007, 317, 1921–1926. [Google Scholar] [CrossRef]
- Kooyman, F.N.J.; Wagenaar, J.A.; Zomer, A. Whole-Genome Sequencing of Dog-Specific Assemblages C and D of Giardia duodenalis from Single and Pooled Cysts Indicates Host-Associated Genes. Microb. Genom. 2019, 5, e000302. [Google Scholar] [CrossRef] [PubMed]
- Adam, R.D.; Dahlstrom, E.W.; Martens, C.A.; Bruno, D.P.; Barbian, K.D.; Ricklefs, S.M.; Hernandez, M.M.; Narla, N.P.; Patel, R.B.; Porcella, S.F.; et al. Genome Sequencing of Giardia lamblia Genotypes A2 and B Isolates (DH and GS) and Comparative Analysis with the Genomes of Genotypes A1 and E (WB and Pig). Genome Biol. Evol. 2013, 5, 2498–2511. [Google Scholar] [CrossRef] [PubMed]
- Ryan, U.; Cacciò, S.M. Zoonotic Potential of Giardia. Int. J. Parasitol. 2013, 43, 943–956. [Google Scholar] [CrossRef] [PubMed]
- Procesi, I.G.; Di Filippo, M.M.; De Liberato, C.; Lombardo, A.; Brocherel, G.; Perrucci, S.; Di Cave, D.; Berrilli, F. Giardia duodenalis in Wildlife: Exploring Genotype Diversity in Italy and across Europe. Pathogens 2022, 11, 105. [Google Scholar] [CrossRef]
- Jian, Y.; Zhang, X.; Li, X.; Schou, C.; Charalambidou, I.; Ma, L.; Karanis, P. Occurrence of Cryptosporidium and Giardia in Wild Birds from Qinghai Lake on the Qinghai-Tibetan Plateau, China. Parasitol. Res. 2021, 120, 615–628. [Google Scholar] [CrossRef] [PubMed]
- Hulme, P.E. Invasive Species Challenge the Global Response to Emerging Diseases. Trends Parasitol. 2014, 30, 267–270. [Google Scholar] [CrossRef] [PubMed]
- Adam, R.D. Biology of Giardia lamblia. Clin. Microbiol. Rev. 2001, 14, 447–475. [Google Scholar] [CrossRef]
- Barash, N.R.; Maloney, J.G.; Singer, S.M.; Dawson, S.C. Giardia Alters Commensal Microbial Diversity throughout the Murine Gut. Infect. Immun. 2017, 85, 10–1128. [Google Scholar] [CrossRef]
- Einarsson, E.; Ma’ayeh, S.; Svärd, S.G. An Up-Date on Giardia and Giardiasis. Curr. Opin. Microbiol. 2016, 34, 47–52. [Google Scholar] [CrossRef]
- Olson, M.E.; Goh, J.; Phillips, M.; Guselle, N.; McAllister, T.A. Giardia Cyst and Cryptosporidium Oocyst Survival in Water, Soil, and Cattle Feces. J. Environ. Qual. 1999, 28, 1991–1996. [Google Scholar] [CrossRef]
- Coffey, C.M.; Collier, S.A.; Gleason, M.E.; Yoder, J.S.; Kirk, M.D.; Richardson, A.M.; Fullerton, K.E.; Benedict, K.M. Evolving Epidemiology of Reported Giardiasis Cases in the United States, 1995–2016. Clin. Infect. Dis. 2021, 72, 764–770. [Google Scholar] [CrossRef]
- McDonnell, P.A.; Upcroft, J.; Upcroft, P.; Buret, A. Morphological identification markers for distinguishing avian from mammalian Giardia species—Do they need reconsideration? Eur. J. Protistol. 2001, 37, 273–280. [Google Scholar] [CrossRef]
- Ballweber, L.R.; Xiao, L.; Bowman, D.D.; Kahn, G.; Cama, V.A. Giardiasis in Dogs and Cats: Update on Epidemiology and Public Health Significance. Trends Parasitol. 2010, 26, 180–189. [Google Scholar] [CrossRef] [PubMed]
- Cacciò, S.M.; Beck, R.; Lalle, M.; Marinculic, A.; Pozio, E. Multilocus Genotyping of Giardia duodenalis Reveals Striking Differences between Assemblages A and B. Int. J. Parasitol. 2008, 38, 1523–1531. [Google Scholar] [CrossRef]
- Hanevik, K.; Bakken, R.; Brattbakk, H.R.; Saghaug, C.S.; Langeland, N. Whole Genome Sequencing of Clinical Isolates of Giardia lamblia. Clin. Microbiol. Infect. 2015, 21, 192.e1–192.e3. [Google Scholar] [CrossRef]
- Yason, J.A.D.L.; Rivera, W.L. Genotyping of Giardia duodenalis Isolates among Residents of Slum Area in Manila, Philippines. Parasitol. Res. 2007, 101, 681–687. [Google Scholar] [CrossRef] [PubMed]
- FAO/WHO. Multicriteria-Based Ranking for Risk Management of Food-Borne Parasites. Microbiological Risk Assessment Series No. 23. Food and Agriculture Organization of the United Nations/World Health Organization. FAO Headquarters, Rome, Italy. 2014. Available online: http://apps.who.int/iris/bitstream/10665/112672/1/9789241564700_eng.pdf (accessed on 9 September 2025).
- Horton, B.; Bridle, H.; Alexander, C.L.; Katzer, F. Giardia duodenalis in the UK: Current Knowledge of Risk Factors and Public Health Implications. Parasitology 2019, 146, 413–424. [Google Scholar] [CrossRef]
- Jenkins, E.J.; Simon, A.; Bachand, N.; Stephen, C. Wildlife parasites in a One Health world. Trends Parasitol. 2015, 31, 174–180. [Google Scholar] [CrossRef]
- Ebani, V.V.; Guardone, L.; Bertelloni, F.; Perrucci, S.; Poli, A.; Mancianti, F. Survey on the Presence of Bacterial and Parasitic Zoonotic Agents in the Feces of Wild Birds. Vet. Sci. 2021, 8, 171. [Google Scholar] [CrossRef]
- Reboredo-Fernandez, A.; Ares-Mazás, E.; Caccio, S.M.; Gómez-Couso, H. Occurrence of Giardia and Cryptosporidium in Wild Birds in Galicia (Northwest Spain). Parasitology 2015, 142, 917–925. [Google Scholar] [CrossRef] [PubMed]
- da Cunha, M.J.R.; Cury, M.C.; Santín, M. Molecular Identification of Enterocytozoon Bieneusi, Cryptosporidium, and Giardia in Brazilian Captive Birds. Parasitol. Res. 2017, 116, 487–493. [Google Scholar] [CrossRef]
- Cano, L.; de Lucio, A.; Bailo, B.; Cardona, G.A.; Muadica, A.S.O.; Lobo, L.; Carmena, D. Identification and Genotyping of Giardia spp. and Cryptosporidium spp. Isolates in Aquatic Birds in the Salburua Wetlands, Álava, Northern Spain. Vet. Parasitol. 2016, 221, 144–148. [Google Scholar] [CrossRef] [PubMed]
- Van Hemert, C.; Ballweber, L.R.; Sinnett, D.R.; Atwood, T.C.; Fischbach, A.; Gustine, D.D.; Pabilonia, K.L. Giardia and Cryptosporidium in resident wildlife species in Arctic Alaska. Food Waterborne Parasitol. 2023, 32, e00206. [Google Scholar] [CrossRef]
- Salubi, E.A.; Gizaw, Z.; Schuster-Wallace, C.J.; Pietroniro, A. Climate change and waterborne diseases in temperate regions: A systematic review. J. Water Health 2025, 23, 58–78. [Google Scholar] [CrossRef]
- Dietrich, J.; Hammerl, J.A.; Johne, A.; Kappenstein, O.; Loeffler, C.; Nöckler, K.; Rosner, B.; Spielmeyer, A.; Szabo, I.; Richter, M.H. Impact of climate change on foodborne infections and intoxications. J. Health Monit. 2023, 8 (Suppl. 3), 78. [Google Scholar] [CrossRef] [PubMed]
- Thompson, R.A. Parasite zoonoses and wildlife: One health, spillover and human activity. Int. J. Parasitol. 2013, 43, 1079–1088. [Google Scholar] [CrossRef] [PubMed]
- Davidson, R.; Simard, M.; Kutz, S.J.; Kapel, C.M.; Hamnes, I.S.; Robertson, L.J. Arctic parasitology: Why should we care? Trends Parasitol. 2011, 27, 239–245. [Google Scholar] [CrossRef]
- Robertson, L.J.; Debenham, J.J. Cryptosporidiosis and Giardiosis in the Arctic: Increasing Threats in a Warmer World? In Arctic One Health: Challenges for Northern Animals and People; Springer International Publishing: Cham, Switzerland, 2022; pp. 339–362. [Google Scholar] [CrossRef]
- Tape, K.D.; Clark, J.A.; Jones, B.M.; Kantner, S.; Gaglioti, B.V.; Grosse, G.; Nitze, I. Expanding beaver pond distribution in Arctic Alaska, 1949 to 2019. Sci. Rep. 2022, 12, 7123. [Google Scholar] [CrossRef]
- Britton, E.; Hales, S.; Venugopal, K.; Baker, M.G. The impact of climate variability and change on cryptosporidiosis and giardiasis rates in New Zealand. J. Water Health 2010, 8, 561–571. [Google Scholar] [CrossRef]
- Lal, A.; Baker, M.G.; Hales, S.; French, N.P. Potential effects of global environmental changes on cryptosporidiosis and giardiasis transmission. Trends Parasitol. 2013, 29, 83–90. [Google Scholar] [CrossRef]
- Jenkins, E.J.; Castrodale, L.J.; de Rosemond, S.J.; Dixon, B.R.; Elmore, S.A.; Gesy, K.M.; Hoberg, E.P.; Polley, L.; Schurer, J.M.; Simard, M.; et al. Tradition and transition: Parasitic zoonoses of people and animals in Alaska, northern Canada, and Greenland. Adv. Parasitol. 2013, 82, 33–204. [Google Scholar] [CrossRef]
- Talazadeh, F.; Razijalali, M.H.; Roshanzadeh, N.; Davoodi, P. Survey on the gastrointestinal parasites in Passeriformes and Psittaciformes with a focus on zoonotic parasites: Survey on the gastrointestinal parasites in Passeriformes and Psittaciformes with a focus on zoonotic parasites. J. Hell. Vet. Med. Soc. 2023, 74, 6237–6245. [Google Scholar] [CrossRef]
- McRoberts, K.M.; Meloni, B.P.; Morgan, U.M.; Marano, R.; Binz, N.; Erlandsen, S.L.; Halse, S.A.; Thompson, R.C.A. Morphological and Molecular Characterization of Giardia Isolated from the Straw-Necked Ibis (Threskiornis spinicollis) in Western Australia. J. Parasitol. 1996, 82, 711. [Google Scholar] [CrossRef] [PubMed]
- Alarcón-Zapata, A.M.; Romero-Salas, D.; Chaparro-Gutiérrez, J.J.; González-Hernández, M.; Ojeda-Chi, M.; Serrano-Solís, A. Frequency of Giardia spp. and Cryptosporidium spp. in domestic and captive wild animals in the north of Veracruz, Mexico. Pak. Vet. J. 2023, 43, 814–818. [Google Scholar] [CrossRef]
- Kuhn, R.C.; Rock, C.M.; Oshima, K.H. Occurrence of Cryptosporidium and Giardia in Wild Ducks along the Rio Grande River Valley in Southern New Mexico. Appl. Env. Microbiol. 2002, 68, 161–165. [Google Scholar] [CrossRef] [PubMed]
- Erlandsen, S.L.; Bemrick, W.J.; Wells, C.L.; Feely, D.E.; Knudson, L.; Campbell, S.R.; van Keulen, H.; Jarroll, E.L. Axenic culture and characterization of Giardia ardeae from the great blue heron (Ardea herodias). J. Parasitol. 1990, 76, 717–724. [Google Scholar] [CrossRef]
- Oates, S.C.; Miller, M.A.; Hardin, D.; Conrad, P.A.; Melli, A.; Jessup, D.A.; Dominik, C.; Roug, A.; Tinker, M.T.; Miller, W.A. Prevalence, environmental loading, and molecular characterization of Cryptosporidium and Giardia isolates from domestic and wild animals along the Central California Coast. Appl. Environ. Microbiol. 2012, 78, 8762–8772. [Google Scholar] [CrossRef]
- Sandoval-Rodríguez, A.; Marcone, D.; Alegría-Morán, R.; Larraechea, M.; Yévenes, K.; Fredes, F.; Briceño, C. Cryptosporidium spp. and Giardia spp. in Free-Ranging Introduced Monk Parakeets from Santiago, Chile. Animals 2021, 11, 801. [Google Scholar] [CrossRef]
- Plutzer, J.; Tomor, B. The Role of Aquatic Birds in the Environmental Dissemination of Human Pathogenic Giardia duodenalis Cysts and Cryptosporidium Oocysts in Hungary. Parasitol. Int. 2009, 58, 227–231. [Google Scholar] [CrossRef]
- Franssen, F.F.J.; Hooimeijer, J.; Blankenstein, B.; Houwers, D.J. Giardiasis in a white stork in The Netherlands. J. Wildl. Dis. 2000, 36, 764–766. [Google Scholar] [CrossRef][Green Version]
- Majewska, A.C.; Graczyk, T.K.; Słodkowicz-Kowalska, A.; Tamang, L.; Jędrzejewski, S.; Zduniak, P.; Solarczyk, P.; Nowosad, A.; Nowosad, P. The Role of Free-Ranging, Captive, and Domestic Birds of Western Poland in Environmental Contamination with Cryptosporidium parvum Oocysts and Giardia lamblia Cysts. Parasitol. Res. 2009, 104, 1093–1099. [Google Scholar] [CrossRef]
- Shemshadi, B.; Ranjbar-Bahadori, S.; Faghihzadeh-Gorgi, S. Occurrence of parasitic protozoa in wild waterfowl in southern coastal Caspian Sea lagoons. Iran. J. Vet. Med. 2014, 8, 261–267. [Google Scholar]
- Papini, R.; Girivetto, M.; Marangi, M.; Mancianti, F.; Giangaspero, A. Endoparasite Infections in Pet and Zoo Birds in Italy. Sci. World J. 2012, 2012, 253127. [Google Scholar] [CrossRef]
- Abe, N.; Makino, I.; Kojima, A. Molecular Characterization of Giardia psittaci by Multilocus Sequence Analysis. Infect. Genet. Evol. 2012, 12, 1710–1716. [Google Scholar] [CrossRef] [PubMed]
- Boehm, A.B.; Soller, J.A. Refined Ambient Water Quality Thresholds for Human-Associated Fecal Indicator HF183 for Recreational Waters with and without Co-Occurring Gull Fecal Contamination. Microb. Risk Anal. 2020, 16, 100139. [Google Scholar] [CrossRef]
- Appelbee, A.J.; Frederick, L.M.; Heitman, T.L.; Olson, M.E. Prevalence and Genotyping of Giardia duodenalis from Beef Calves in Alberta, Canada. Vet. Parasitol. 2003, 112, 289–294. [Google Scholar] [CrossRef]
- Li, J.; Qin, H.; Li, X.; Zhang, L. Role of Rodents in the Zoonotic Transmission of Giardiasis. One Health 2023, 16, 100500. [Google Scholar] [CrossRef]
- Durrani, A.Z.; Saleem, M.H.; Ahmad, H.A.; Ali, M.H.; Chaudhary, M.; Usman, M. Bacterial and Parasitic Profiling of Native Pigeons in District Lahore-Pakistan. In Proceedings of the 2020 17th International Bhurban Conference on Applied Sciences and Technology (IBCAST), Islamabad, Pakistan, 14–18 January 2020; IEEE: Piscataway, NJ, USA, 2020; pp. 191–194. [Google Scholar]
- Dong, H.; Cheng, R.; Li, X.; Li, J.; Chen, Y.; Ban, C.; Zhang, X.; Liu, F.; Zhang, L. Molecular Identification of Cryptosporidium spp., Enterocytozoon bieneusi, and Giardia duodenalis in Captive Pet Birds in Henan Province, Central China. J. Eukaryot. Microbiol. 2021, 68, e12839. [Google Scholar] [CrossRef]
- Zahedi, A.; Field, D.; Ryan, U. Molecular Typing of Giardia duodenalis in Humans in Queensland—First Report of Assemblage E. Parasitology 2017, 144, 1154–1161. [Google Scholar] [CrossRef]
- Egan, S.; Barbosa, A.D.; Feng, Y.; Xiao, L.; Ryan, U. The Risk of Wild Birds Contaminating Source Water with Zoonotic Cryptosporidium and Giardia Is Probably Overestimated. Sci. Total Environ. 2024, 912, 169032. [Google Scholar] [CrossRef]
- Sarria-Guzmán, Y.; Chávez-Romero, Y.; Bernal, J.E.; González-Jiménez, F.E.; Serrano-Silva, N.; Fusaro, C. Molecular Identification of Giardia spp. in Latin America: An Updated Systematic Review on Reports from 2017 to 2021. J. Infect. Dev. Ctries. 2022, 16, 392–401. [Google Scholar] [CrossRef]
- Bomfim, T.C.B.; Gomes, R.S.; Huber, F.; Couto, M.C.M. The importance of poultry in environmental dissemination of Cryptosporidium spp. Open Vet. Sci. J. 2013, 7, 12–17. [Google Scholar] [CrossRef][Green Version]
- Qi, M.; Wang, R.; Ning, C.; Li, X.; Zhang, L.; Jian, F.; Sun, Y.; Xiao, L. Cryptosporidium spp. in pet birds: Genetic diversity and potential public health significance. Exp. Parasitol. 2011, 128, 336–340. [Google Scholar] [CrossRef]
- Wang, R.; Wang, F.; Zhao, J.; Qi, M.; Ning, C.; Zhang, L.; Xiao, L. Cryptosporidium spp. in quails (Coturnix coturnix japonica) in Henan, China: Molecular characterization and public health significance. Vet. Parasitol. 2012, 187, 534–537. [Google Scholar] [CrossRef]
- Nakamura, A.A.; Simões, D.; Antunes, R.; Silva, D.C.D.; Meireles, M. Molecular characterization of Cryptosporidium spp. from fecal samples of birds kept in captivity in Brazil. Vet. Parasitol. 2009, 166, 47–51. [Google Scholar] [CrossRef]
- Antunes, R.; Simões, D.; Nakamura, A.A.; Meireles, M. Natural Infection with Cryptosporidium galli in Canaries (Serinus canaria), in a Cockatiel (Nymphicus hollandicus), and in Lesser Seed-Finches (Oryzoborus angolensis) from Brazil. Avian Dis. 2008, 52, 702–705. [Google Scholar] [CrossRef]
- Nakamura, A.A.; Homem, C.G.; Silva, A.J.D.D.; Meireles, M. Diagnosis of gastric cryptosporidiosis in birds using a duplex real-time PCR assay. Vet. Parasitol. 2014, 205, 7–13. [Google Scholar] [CrossRef]
- Li, J.; Lin, X.; Zhang, L.; Qi, N.; Liao, S.; Lv, M.; Wu, C.; Sun, M. Molecular characterization of Cryptosporidium spp. in domestic pigeons (Columba livia domestica) in Guangdong Province, Southern China. Parasitol. Res. 2015, 114, 2237–2241. [Google Scholar] [CrossRef]
- Nakamura, A.A.; Meireles, M. Cryptosporidium infections in birds-a review Infecção por Cryptosporidium em aves-uma revisão. Rev. Bras. Parasitol. Vet. 2015, 24, 253–267. [Google Scholar] [CrossRef]
- Santana, B.N.; Kurahara, B.; Nakamura, A.A.; Camargo, V.D.S.; Ferrari, E.D.; Silva, G.S.D.; Nagata, W.; Meireles, M. Detection and characterization of Cryptosporidium species and genotypes in three chicken production systems in Brazil using different molecular diagnosis protocols. Prev. Vet. Med. 2018, 151, 73–78. [Google Scholar] [CrossRef]
- Sevá, A.P.; Funada, M.R.; Richtzenhain, L.; Guimarães, M.B.; Souza, S.; Allegretti, L.; Sinhorini, J.; Duarte, V.V.; Soares, R. Genotyping of Cryptosporidium spp. from free-living wild birds from Brazil. Vet. Parasitol. 2011, 175, 27–32. [Google Scholar] [CrossRef]
- Baroudi, D.; Khelef, D.; Goucem, R.; Adjou, K.; Adamu, H.; Zhang, H.W.; Xiao, L. Common occurrence of zoonotic pathogen Cryptosporidium meleagridis in broiler chickens and turkeys in Algeria. Vet. Parasitol. 2013, 196, 334–340. [Google Scholar] [CrossRef]
- Ryan, U. Cryptosporidium in birds, fish and amphibians. Exp. Parasitol. 2010, 124, 113–120. [Google Scholar] [CrossRef]
- Cunha, M.J.D.D.; Cury, M.; Santín, M. Widespread presence of human-pathogenic Enterocytozoon bieneusi genotypes in chickens. Vet. Parasitol. 2016, 217, 108–112. [Google Scholar] [CrossRef]
- Makino, I.; Abe, N.; Reavill, D. Cryptosporidium Avian Genotype III as a Possible Causative Agent of Chronic Vomiting in Peach-Faced Lovebirds (Agapornis roseicollis). Avian Dis. 2010, 54, 1102–1107. [Google Scholar] [CrossRef]
- Abe, N.; Makino, I. Multilocus genotypic analysis of Cryptosporidium isolates from cockatiels, Japan. Parasitol. Res. 2010, 106, 1491–1497. [Google Scholar] [CrossRef]
- Ryan, U.; Xiao, L.; Read, C.; Sulaiman, I.; Monis, P.; Lal, A.; Fayer, R.; Pavlásek, I. A Redescription of Cryptosporidium galli pavlasek, 1999 (apicomplexa: Cryptosporidiidae) from birds. J. Parasitol. 2003, 89, 809–813. [Google Scholar] [CrossRef]
- Holubová, N.; Sak, B.; Hořčičková, M.; Hlásková, L.; Květoňová, D.; Menchaca, S.; Mcevoy, J.; Kváč, M. Cryptosporidium avium n. sp. (Apicomplexa: Cryptosporidiidae) in birds. Parasitol. Res. 2016, 115, 2243–2251. [Google Scholar] [CrossRef]
- Graczyk, T.; Majewska, A.; Schwab, K. The role of birds in dissemination of human waterborne enteropathogens. Trends Parasitol. 2008, 24, 55–59. [Google Scholar] [CrossRef]
- Wang, L.; Xue, X.; Li, J.Q.; Zhou, Q.J.; Yu, Y.; Du, A. Cryptosporidiosis in broiler chickens in Zhejiang Province, China: Molecular characterization of oocysts detected in fecal samples. Parasite 2014, 21, 36. [Google Scholar] [CrossRef]
- Wang, R.; Jian, F.; Sun, Y.; Hu, Q.; Zhu, J.; Wang, F.; Ning, C.; Zhang, L.; Xiao, L. Large-scale survey of Cryptosporidium spp. in chickens and Pekin ducks (Anas platyrhynchos) in Henan, China: Prevalence and molecular characterization. Avian Pathol. 2010, 39, 447–451. [Google Scholar] [CrossRef]
- Chelladurai, J.J.; Clark, M.; Kváč, M.; Holubová, N.; Khan, E.; Stenger, B.L.S.; Giddings, C.; Mcevoy, J. Cryptosporidium galli and novel Cryptosporidium avian genotype VI in North American red-winged blackbirds (Agelaius phoeniceus). Parasitol. Res. 2016, 115, 1901–1906. [Google Scholar] [CrossRef]
- Zhang, X.X.; Zhang, N.; Zhao, G.H.; Zhao, Q.; Zhu, X.Q. Prevalence and Genotyping of Cryptosporidium Infection in Pet Parrots in North China. BioMed Res. Int. 2015, 2015, 549798. [Google Scholar] [CrossRef]
- Silva, D.C. Avaliação fÍsica, Epidemiológica e Molecular da Infecção por Cryptosporidium spp. em passeriformes. Master’s Thesis, Universidade Estadual Paulista, Sao Paulo, Brazil, 2009. [Google Scholar]
- El-Ghany, W.A.A. Avian Cryptosporidiosis: A significant parasitic disease of public health hazard. Slov. Vet. Res. 2022, 59, 5. [Google Scholar] [CrossRef]
- Meireles, M.; Soares, R.; Santos, M.M.A.B.D.; Gennari, S. Biological studies and molecular characterization of a Cryptosporidium isolate from ostriches (Struthio camelus). J. Parasitol. 2006, 92, 623–626. [Google Scholar] [CrossRef]
- Wang, K.; Gazizova, A.; Wang, Y.; Zhang, K.; Zhang, Y.; Chang, Y.; Cui, Y.; Zhang, Y.; Zhang, S.M.; Zhang, L. First Detection of Cryptosporidium spp. in Migratory Whooper Swans (Cygnus cygnus) in China. Microorganisms 2019, 8, 6. [Google Scholar] [CrossRef]
- Paulo, S. Determinação da Ocorrência de Cryptosporidium galli em Amostras Fecais de Aves por meio da PCR em Tempo Real. Doctoral Dissertation, Universidade de São Paulo, São Paulo, Brazil, 2013. [Google Scholar]
- Laatamna, A.E.; Holubová, N.; Sak, B.; Kváč, M. Cryptosporidium meleagridis and C. baileyi (Apicomplexa) in domestic and wild birds in Algeria. Folia Parasitol. 2017, 64, 018. [Google Scholar] [CrossRef]
- Silva, D.C.; Homem, C.G.; Nakamura, A.A.; Teixeira, W.; Perri, S.; Meireles, M. Physical, epidemiological, and molecular evaluation of infection by Cryptosporidium galli in Passeriformes. Parasitol. Res. 2010, 107, 271–277. [Google Scholar] [CrossRef]
- Helmy, Y.A.; Krücken, J.; Abdelwhab, E.S.M.; Samson-Himmelstjerna, G.V.; Hafez, H. Molecular diagnosis and characterization of Cryptosporidium spp. in turkeys and chickens in Germany reveals evidence for previously undetected parasite species. PLoS ONE 2017, 12, e0177150. [Google Scholar] [CrossRef]
- Ng, J.; Pavlásek, I.; Ryan, U. Identification of Novel Cryptosporidium Genotypes from Avian Hosts. Appl. Environ. Microbiol. 2006, 72, 7548–7553. [Google Scholar] [CrossRef]
- Máca, O.; Pavlásek, I. First finding of spontaneous infections with Cryptosporidium baileyi and C. meleagridis in the red-legged partridge Alectoris rufa from an aviary in the Czech Republic. Vet. Parasitol. 2015, 209, 164–168. [Google Scholar] [CrossRef]
- Gomes, R.S.; Huber, F.; Silva, S.; Bomfim, T.C.B. Cryptosporidium spp. parasitize exotic birds that are commercialized in markets, commercial aviaries, and pet shops. Parasitol. Res. 2011, 110, 1363–1370. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).