
Genomic Characterization and Phylogenetic Analysis of Five Avian Influenza H5N1 Subtypes from Wild Anser indicus in Yunnan, China
 ,
,  ,
,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
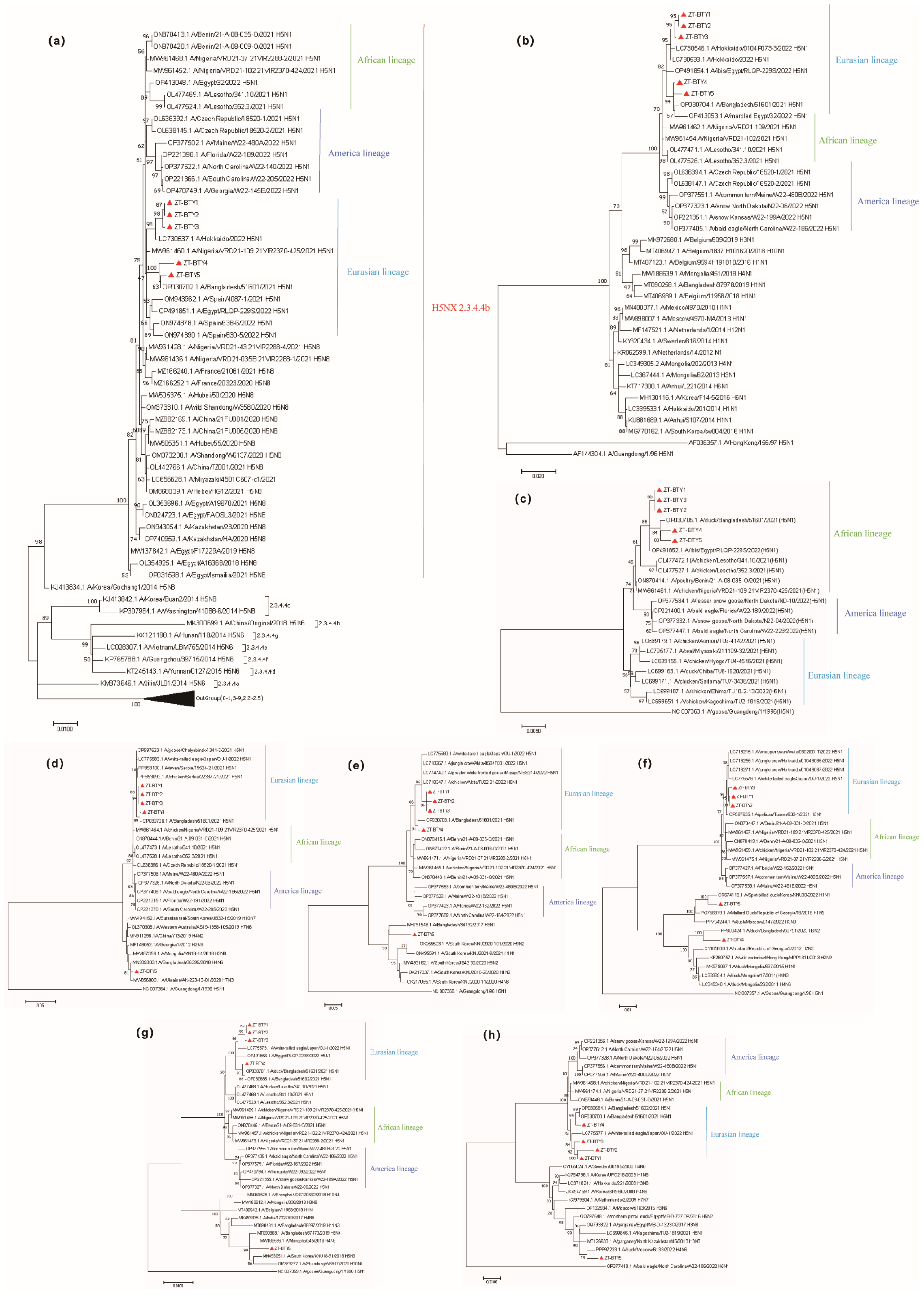
3. Results and Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Luo, W.; Tian, L.; Gan, Y.; Chen, E.; Shen, X.; Pan, J.; Irwin, D.M.; Chen, R.A.; Shen, Y. The fit of codon usage of human-isolated avian influenza A viruses to human. Infect. Genet. Evol. 2020, 81, 104181. [Google Scholar] [CrossRef] [PubMed]
- de Bruin, A.C.M.; Funk, M.; Spronken, M.I.; Gultyaev, A.P.; Fouchier, R.A.M.; Richard, M. Hemagglutinin Subtype Specificity and Mechanisms of Highly Pathogenic Avian Influenza Virus Genesis. Viruses 2022, 14, 1566. [Google Scholar] [CrossRef] [PubMed]
- Wille, M.; Holmes, E.C. The Ecology and Evolution of Influenza Viruses. Cold Spring Harb. Perspect. Med. 2020, 10, a038489. [Google Scholar] [CrossRef]
- Xu, X.; Subbarao Cox, N.J.; Guo, Y. Genetic characterization of the pathogenic influenza A/Goose/Guangdong/1/96 (H5N1) virus: Similarity of its hemagglutinin gene to those of H5N1 viruses from the 1997 outbreaks in Hong Kong. Virology 1999, 261, 15–19. [Google Scholar] [CrossRef]
- Cui, Y.; Li, Y.; Li, M.; Zhao, L.; Wang, D.; Tian, J.; Bai, X.; Ci, Y.; Wu, S.; Wang, F.; et al. Evolution and extensive reassortment of H5 influenza viruses isolated from wild birds in China over the past decade. Emerg. Microbes Infect. 2020, 9, 1793–1803. [Google Scholar] [CrossRef]
- Plaza, P.I.; Gamarra-Toledo, V.; Euguí, J.R.; Lambertucci, S.A. Recent Changes in Patterns of Mammal Infection with Highly Pathogenic Avian Influenza A(H5N1) Virus Worldwide. Emerg. Infect. Dis. 2024, 30, 444–452. [Google Scholar] [CrossRef]
- Burrough, E.R.; Magstadt, D.R.; Petersen, B.; Timmermans, S.J.; Gauger, P.C.; Zhang, J.; Siepker, C.; Mainenti, M.; Li, G.; Thompson, A.C.; et al. Highly Pathogenic Avian Influenza A(H5N1) Clade 2.3.4.4b Virus Infection in Domestic Dairy Cattle and Cats, United States, 2024. Emerg. Infect. Dis. 2024, 30, 1335–1343. [Google Scholar] [CrossRef]
- Peiris, J.S.; Yu, W.C.; Leung, C.W.; Cheung, C.Y.; Ng, W.F.; Nicholls, J.M.; Ng, T.K.; Chan, K.H.; Lai, S.T.; Lim, W.L.; et al. Re-emergence of fatal human influenza A subtype H5N1 disease. Lancet 2004, 363, 617–619. [Google Scholar] [CrossRef]
- Pulit-Penaloza, J.A.; Brock, N.; Belser, J.A.; Sun, X.; Pappas, C.; Kieran, T.J.; Basu Thakur, P.; Zeng, H.; Cui, D.; Frederick, J.; et al. Highly pathogenic avian influenza A(H5N1) virus of clade 2.3.4.4b isolated from a human case in Chile causes fatal disease and transmits between co-housed ferrets. Emerg. Microbes Infect. 2024, 13, 2332667. [Google Scholar] [CrossRef]
- Lee, D.H.; Bertran, K.; Kwon, J.H.; Swayne, D.E. Evolution, global spread, and pathogenicity of highly pathogenic avian influenza H5Nx clade 2.3.4.4. J. Vet. Sci. 2017, 18, 269–280. [Google Scholar] [CrossRef]
- Lazzari, S.; Stöhr, K. Avian influenza and influenza pandemics. Bull. World Health Organ. 2004, 82, 242. [Google Scholar] [PubMed]
- Global Consortium for H5N8 and Related Influenza Viruses. Role for migratory wild birds in the global spread of avian influenza H5N8. Science 2016, 354, 213–217. [Google Scholar] [CrossRef] [PubMed]
- Lawpoolsri, S.; Kaewkungwal, J.; Khamsiriwatchara, A.; Sovann, L.; Sreng, B.; Phommasack, B.; Kitthiphong, V.; Lwin Nyein, S.; Win Myint, N.; Dang Vung, N.; et al. Data quality and timeliness of outbreak reporting system among countries in Greater Mekong subregion: Challenges for international data sharing. PLoS Negl. Trop. Dis. 2018, 12, e0006425. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.; Yaqub, T.; Shahid, M.F.; Wong, F.Y.; Mukhtar, N.; Naeem, M.; Lam, P.; Jayakumar, J.; Smith, G.J.D.; Su, Y.C.F. Genetic Characterization of Highly Pathogenic Avian Influenza A(H5N8) Virus in Pakistani Live Bird Markets Reveals Rapid Diversification of Clade 2.3.4.4b Viruses. Viruses 2021, 13, 1633. [Google Scholar] [CrossRef]
- Yang, Q.; Xue, X.; Zhang, Z.; Wu, M.J.; Ji, J.; Wang, W.; Yin, H.; Li, S.; Dai, H.; Duan, B.; et al. Clade 2.3.4.4b H5N8 Subtype Avian Influenza Viruses Were Identified from the Common Crane Wintering in Yunnan Province, China. Viruses 2022, 15, 38. [Google Scholar] [CrossRef]
- Hoffmann, E.; Stech, J.; Guan, Y.; Webster, R.G.; Perez, D.R. Universal primer set for the full-length amplification of all influenza A viruses. Arch. Virol. 2001, 146, 2275–2289. [Google Scholar] [CrossRef]
- Wang, J.; Shah, T.; Zhou, J.; Long, X.; Wang, Y.; Chen, J.; Shi, M.; Shah, Z.; Wang, B.; Xia, X. Identification, Characterization, and Homology Analysis of a Novel Strain of the Crimean-Congo Hemorrhagic Fever Virus from Yunnan, China. Microorganisms 2024, 12, 1466. [Google Scholar] [CrossRef]
- Xu, W.; Berhane, Y.; Dubé, C.; Liang, B.; Pasick, J.; VanDomselaar, G.; Alexandersen, S. Epidemiological and Evolutionary Inference of the Transmission Network of the 2014 Highly Pathogenic Avian Influenza H5N2 Outbreak in British Columbia, Canada. Sci. Rep. 2016, 6, 30858. [Google Scholar] [CrossRef]
- Ye, H.; Zhang, J.; Sang, Y.; Shan, N.; Qiu, W.; Zhong, W.; Li, J.; Yuan, Z. Divergent Reassortment and Transmission Dynamics of Highly Pathogenic Avian Influenza A(H5N8) Virus in Birds of China During 2021. Front. Microbiol. 2022, 13, 913551. [Google Scholar] [CrossRef]
- Tian, J.; Li, M.; Bai, X.; Li, Y.; Wang, X.; Wang, F.; Shi, J.; Zeng, X.; Tian, G.; Li, Y. H5 low pathogenic avian influenza viruses maintained in wild birds in China. Vet. Microbiol. 2021, 263, 109268. [Google Scholar] [CrossRef]
- AbuBakar, U.; Amrani, L.; Kamarulzaman, F.A.; Karsani, S.A.; Hassandarvish, P.; Khairat, J.E. Avian Influenza Virus Tropism in Humans. Viruses 2023, 15, 833. [Google Scholar] [CrossRef]
- Mostafa, A.; Abdelwhab, E.M.; Mettenleiter, T.C.; Pleschka, S. Zoonotic Potential of Influenza A Viruses: A Comprehensive Overview. Viruses 2018, 10, 497. [Google Scholar] [CrossRef]
- Yang, Z.Y.; Wei, C.J.; Kong, W.P.; Wu, L.; Xu, L.; Smith, D.F.; Nabel, G.J. Immunization by avian H5 influenza hemagglutinin mutants with altered receptor binding specificity. Science 2007, 317, 825–828. [Google Scholar] [CrossRef]
- Lee, C.Y.; An, S.H.; Choi, J.G.; Lee, Y.J.; Kim, J.H.; Kwon, H.J. Rank orders of mammalian pathogenicity-related PB2 mutations of avian influenza A viruses. Sci. Rep. 2020, 10, 5359. [Google Scholar] [CrossRef]
- Duong, B.T.; Bal, J.; Sung, H.W.; Yeo, S.J.; Park, H. Molecular Analysis of the Avian H7 Influenza Viruses Circulating in South Korea during 2018-2019: Evolutionary Significance and Associated Zoonotic Threats. Viruses 2021, 13, 2260. [Google Scholar] [CrossRef]
- Chakraborty, C.; Bhattacharya, M. Evolution and mutational landscape of highly pathogenic avian influenza strain A(H5N1) in the current outbreak in the USA and global landscape. Virology 2024, 600, 110246. [Google Scholar] [CrossRef]
- Suttie, A.; Deng, Y.M.; Greenhill, A.R.; Dussart, P.; Horwood, P.F.; Karlsson, E.A. Inventory of molecular markers affecting biological characteristics of avian influenza A viruses. Virus Genes 2019, 55, 739–768. [Google Scholar] [CrossRef]
- Pu, J.; Sun, H.; Qu, Y.; Wang, C.; Gao, W.; Zhu, J.; Sun, Y.; Bi, Y.; Huang, Y.; Chang, K.C.; et al. M Gene Reassortment in H9N2 Influenza Virus Promotes Early Infection and Replication: Contribution to Rising Virus Prevalence in Chickens in China. J. Virol. 2017, 91, e02055-16. [Google Scholar] [CrossRef]
- Atim, G.; Tugume, T.; Ukuli, Q.A.; Erima, B.; Mubiru, A.; Kibuuka, H.; Mworozi, E.; McKenzie, P.; Turner, J.C.M.; Walker, D.; et al. Genetic Evolution of Avian Influenza A (H9N2) Viruses Isolated from Domestic Poultry in Uganda Reveals Evidence of Mammalian Host Adaptation, Increased Virulence and Reduced Sensitivity to Baloxavir. Viruses 2022, 14, 2074. [Google Scholar] [CrossRef]
- Nao, N.; Kajihara, M.; Manzoor, R.; Maruyama, J.; Yoshida, R.; Muramatsu, M.; Miyamoto, H.; Igarashi, M.; Eguchi, N.; Sato, M.; et al. A Single Amino Acid in the M1 Protein Responsible for the Different Pathogenic Potentials of H5N1 Highly Pathogenic Avian Influenza Virus Strains. PLoS ONE 2015, 10, e0137989. [Google Scholar] [CrossRef]
- Gaymard, A.; Charles-Dufant, A.; Sabatier, M.; Cortay, J.C.; Frobert, E.; Picard, C.; Casalegno, J.S.; Rosa-Calatrava, M.; Ferraris, O.; Valette, M.; et al. Impact on antiviral resistance of E119V, I222L and R292K substitutions in influenza A viruses bearing a group 2 neuraminidase (N2, N3, N6, N7 and N9). J. Antimicrob. Chemother. 2016, 71, 3036–3045. [Google Scholar] [CrossRef]
- Tang, J.; Gao, R.; Liu, L.; Zhang, S.; Liu, J.; Li, X.; Fang, Q.; Feng, Z.; Xu, C.; Huang, W.; et al. Substitution of I222L-E119V in neuraminidase from highly pathogenic avian influenza H7N9 virus exhibited synergistic resistance effect to oseltamivir in mice. Sci. Rep. 2021, 11, 16293. [Google Scholar] [CrossRef]
- Paradis, E.G.; Pinilla, L.T.; Holder, B.P.; Abed, Y.; Boivin, G.; Beauchemin, C.A. Impact of the H275Y and I223V Mutations in the Neuraminidase of the 2009 Pandemic Influenza Virus In Vitro and Evaluating Experimental Reproducibility. PLoS ONE 2015, 10, e0126115. [Google Scholar] [CrossRef]
- Rey-Carrizo, M.; Barniol-Xicota, M.; Ma, C.; Frigolé-Vivas, M.; Torres, E.; Naesens, L.; Llabrés, S.; Juárez-Jiménez, J.; Luque, F.J.; DeGrado, W.F.; et al. Easily accessible polycyclic amines that inhibit the wild-type and amantadine-resistant mutants of the M2 channel of influenza A virus. J. Med. Chem. 2014, 57, 5738–5747. [Google Scholar] [CrossRef]
- Yin, Y.; Li, W.; Chen, R.; Wang, X.; Chen, Y.; Cui, X.; Lu, X.; Irwin, D.M.; Shen, X.; Shen, Y. Random forest algorithm reveals novel sites in HA protein that shift receptor binding preference of the H9N2 avian influenza virus. Virol. Sin. 2024; in press. [Google Scholar] [CrossRef]
- Sun, X.; Belser, J.A.; Pulit-Penaloza, J.A.; Brock, N.; Kieran, T.J.; Pappas, C.; Zeng, H.; Tumpey, T.M.; Maines, T.R. Dissecting the role of the HA1-226 leucine residue in the fitness and airborne transmission of an A(H9N2) avian influenza virus. J. Virol. 2024, 98, e0092824. [Google Scholar] [CrossRef]
- Gabriel, G.; Czudai-Matwich, V.; Klenk, H.D. Adaptive mutationsin the H5N1polymerase complex. Virus Res. 2013, 178, 53–62. [Google Scholar] [CrossRef]
- Zhang, X.; Xu, G.; Wang, C.; Jiang, M.; Gao, W.; Wang, M.; Sun, H.; Sun, Y.; Chang, K.C.; Liu, J.; et al. Enhanced pathogenicity and neurotropism of mouse-adapted H10N7 influenza virus are mediated by novel PB2 and NA mutations. J. Gen. Virol. 2017, 98, 1185–1195. [Google Scholar] [CrossRef]
- Jiao, P.; Tian, G.; Li, Y.; Deng, G.; Jiang, Y.; Liu, C.; Liu, W.; Bu, Z.; Kawaoka, Y.; Chen, H. A single-amino-acid substitution in the NS1 protein changes the pathogenicity of H5N1 avian influenza viruses in mice. J. Virol. 2008, 82, 1146–1154. [Google Scholar] [CrossRef]

{kind=link}
| Protein | Amino Acid/Motif | Phenotypic Characteristic | ZT-BTY-1 | ZT-BTY-2 | ZT-BTY-3 | ZT-BTY-4 | ZT-BTY-5 |
|---|---|---|---|---|---|---|---|
| HA | N158D | Lack of glycosylation sites to promote airborne transmission to ferrets and humans [35] | N | N | N | D | D |
| T160A | A | A | A | A | A | ||
| S128P | Enhanced sialic acid A-2, 6-galactose receptor tropism [36] | P | P | P | P | P | |
| S137A | A | A | A | A | A | ||
| Q226L | Q | Q | Q | Q | Q | ||
| S227R | R | R | R | R | R | ||
| G228S | G | G | G | G | G | ||
| NA | E119V | Neuraminidase inhibitor resistance [31,32,33] | E | E | E | E | E |
| R152K | R | R | R | R | R | ||
| H275Y | H | H | H | H | H | ||
| R293K | R | R | R | R | R | ||
| N295S | N | N | N | N | N | ||
| I117T | I | I | I | I | I | ||
| PB2 | L89V | Increased virus polymerase activity [25] | V | V | V | V | V |
| E627K | Mammalian tropism [37,38] | E | E | E | E | E | |
| M631L | M | M | M | M | M | ||
| D701N | D | D | D | D | D | ||
| PB1 | L473V | Increased virus polymerase activity and replication in mammalian cell lines [27] | V | V | V | V | V |
| L598P | L | L | L | L | L | ||
| PA | N383D | Increased virus polymerase activity and replication in duck and mammalian cell lines [27] | D | D | D | D | D |
| N409S | S | S | S | S | S | ||
| K497R | K | K | K | K | K | ||
| S515T | T | T | T | T | T | ||
| NP | M105V | Increased virus pathogenicity in chickens [29] | V | V | V | V | V |
| I109T | I | I | I | I | I | ||
| M1 | N30D | Increased virus pathogenicity in mice [29,30] | D | D | D | D | D |
| I43M | M | M | M | M | M | ||
| T215A | A | A | A | A | A | ||
| M2 | L26F | M2 ion channel inhibitor resistance [34] | L | L | L | L | L |
| S31N | S | S | S | S | S | ||
| NS | P42S | Enhanced virus pathogenicity in mice [25,29] | S | S | S | S | S |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, L.; Wang, R.; Liu, Q.; Shah, T.; Zhou, J.; Zhao, W.; Wang, Y.; Deng, L.; Wang, B. Genomic Characterization and Phylogenetic Analysis of Five Avian Influenza H5N1 Subtypes from Wild Anser indicus in Yunnan, China. Vet. Sci. 2025, 12, 280. https://doi.org/10.3390/vetsci12030280
Yang L, Wang R, Liu Q, Shah T, Zhou J, Zhao W, Wang Y, Deng L, Wang B. Genomic Characterization and Phylogenetic Analysis of Five Avian Influenza H5N1 Subtypes from Wild Anser indicus in Yunnan, China. Veterinary Sciences. 2025; 12(3):280. https://doi.org/10.3390/vetsci12030280
Chicago/Turabian StyleYang, Lingsi, Rui Wang, Qi Liu, Taif Shah, Jiuxuan Zhou, Wenhua Zhao, Yixuan Wang, Lulu Deng, and Binghui Wang. 2025. "Genomic Characterization and Phylogenetic Analysis of Five Avian Influenza H5N1 Subtypes from Wild Anser indicus in Yunnan, China" Veterinary Sciences 12, no. 3: 280. https://doi.org/10.3390/vetsci12030280
APA StyleYang, L., Wang, R., Liu, Q., Shah, T., Zhou, J., Zhao, W., Wang, Y., Deng, L., & Wang, B. (2025). Genomic Characterization and Phylogenetic Analysis of Five Avian Influenza H5N1 Subtypes from Wild Anser indicus in Yunnan, China. Veterinary Sciences, 12(3), 280. https://doi.org/10.3390/vetsci12030280