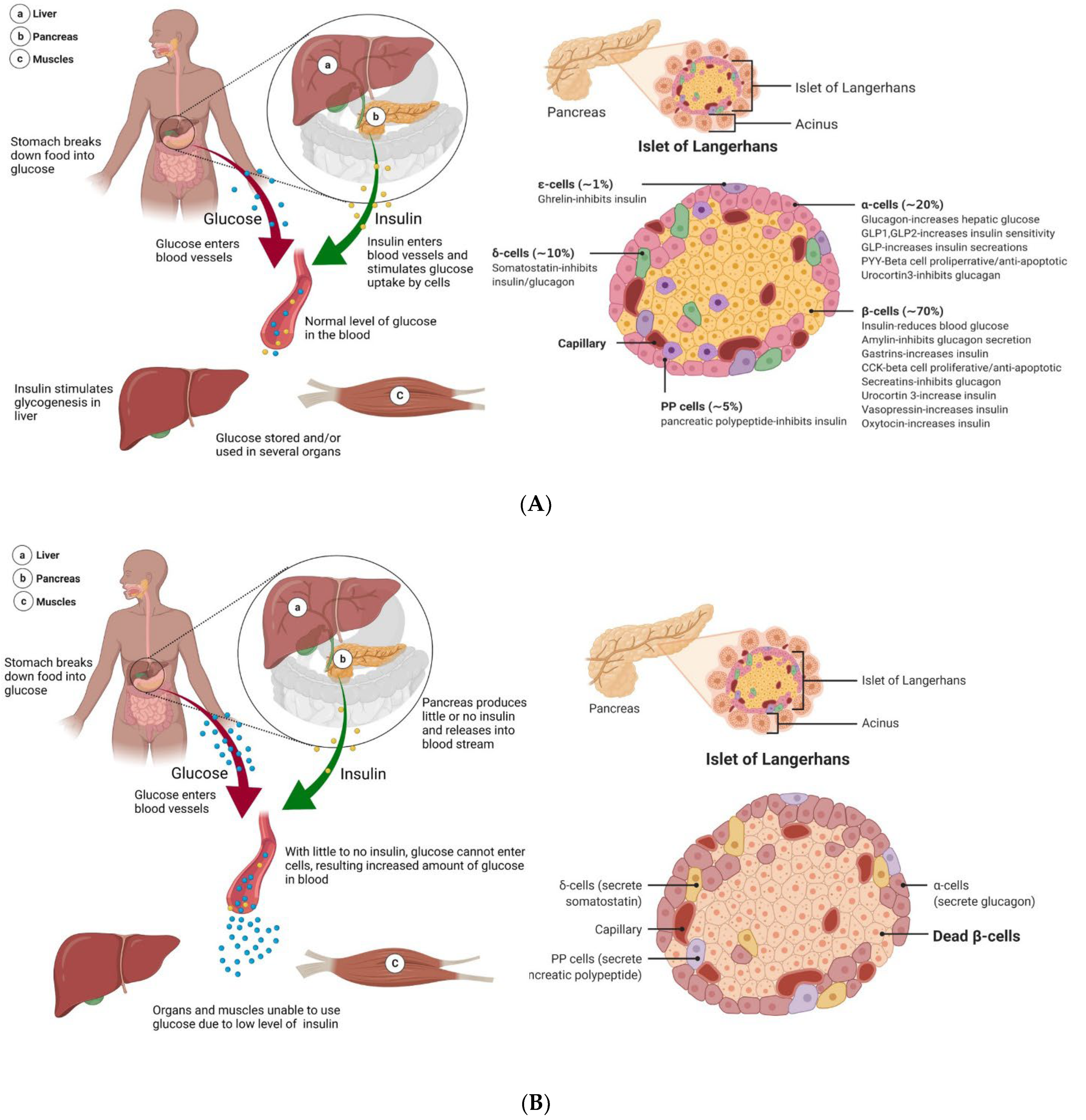
4.1. Healthy Blood Sugar Regulation
Diabetes is a disease in which metabolic processing and energy regulation malfunction. Energy in the body is provided through an intricate balance of food intake, breakdown, and uptake. In a healthy, non-diabetic system, the carbohydrates, lipids, and proteins from food are broken down into glucose, which is then delivered to cells via facilitated diffusion to be used for energy. Insulin is the vital gatekeeper in this process, regulating blood glucose levels (also known as blood sugar) by triggering cells to take up glucose from the blood (
Figure 2A) [
41,
42,
43]. This increase in membrane permeability is thought to be a result of the translocation of glucose receptors to the cell membrane [
42,
43]. The pancreas plays a key role in this process, as it aids in the digestion, absorption, utilization, and storage of energy substrates [
41,
44]. The pancreas is located close to other important organs in metabolism, lying below the stomach and extending from the spleen to the duodenum [
45]. The pancreas is structurally distinct, containing cells that perform exocrine (nutrient digestion) and endocrine functions (glucose homeostasis) [
44,
45]. Though endocrine cells, which make up clusters called the islets of Langerhans (or pancreatic islets), are vital in the regulation of blood sugar, they make up only 1–2% of the adult pancreas [
7,
44,
45,
46]. Each islet has a number of distinct cell types, which produce hormones that play different roles in blood sugar maintenance. This includes α-cells and δ-cells, which produce glucagon (which has a role opposite to that of insulin, raising blood glucose levels) and somatostatin (which regulates the secretion of other hormones from the pancreas), respectively (
Figure 2A) [
44,
45]. β-cells, which make up 65 to 80% of islets, produce insulin and amylin at a 100:1 ratio, which collaboratively works to prevent blood sugar spikes by triggering the uptake of glucose into cells and promoting satiety [
44,
45,
46,
47]. Each β-cell produces up to 1 million insulin molecules/minute (a number that is further elevated in response to eating) [
7,
46]. Insulin is considered a “key” in sugar regulation, allowing cells to uptake glucose by increasing the permeability of plasma membrane to glucose, inducing the conversion of glucose to glycogen in the liver, and promoting the production of fatty acids and proteins [
46]. Together, the cells of the pancreas and the hormones secreted regulate blood sugar, with the average, optimal 24 h glucose levels of 99 ± 7 mg/dL [
48]. In non-optimal, hypoglycemic conditions, blood sugar levels are below ideal levels, triggering the secretion of glucagon from α-cells, which promotes the breakdown of glycogen stored in the liver to glucose [
46]. When the body has high blood glucose content, called a hyperglycemic state, β-cells release insulin and amylin to reduce the blood sugar levels and decrease food intake [
46]. This system malfunctions in T1DM causing insulin levels drop, and blood sugar levels spike as cells are unable to take up glucose.
4.2. Mechanisms in T1DM
Type 1 diabetes is a multifaceted disorder produced by a complex interplay of environmental and genetic variables, the former of which includes several susceptibility genes. The discovery of genes that confer vulnerability to T1DM would help to clarify pathogenetic processes in the establishment and progression of the disease, paving the way for the development of effective approaches for disease prevention and management. Genome-wide linkage analysis and genome-wide association studies (GWAS) and genome-wide linkage analysis have recently contributed significantly to our understanding of the role of genetics in the onset and progression of T1DM. Nearly 60 risk regions all over the human genome were found by GWAS, which is designated by single nucleotide polymorphisms that confer genetic susceptibility to T1DM [
49,
50]. Several studies claimed that 50% of the risk factors for T1DM are genetic [
51]. When researchers first began investigating the causes of T1DM, genetic risk factors were focused on as the sole trigger, as susceptibility to developing T1DM is influenced by carrying specific high-risk haplotypes of class II human leukocyte antigens (HLA), such as HLA-DR (DR3/4) and HLA-DQ (DQ8) [
4,
52,
53]. Over 50 non-human leukocyte antigen (HLA) regions and HLA genes were discovered in T1DM using genomic screening [
51]. HLA-DQ and HLA-DR, in particular, are closely linked to T1DM. Haplotypes of class II molecules antigens can substantially increase or decrease the binding properties of associated autoantigens, resulting in T1DM development [
54,
55]. The impact of HLA on T1DM risk is supported by a twin concordance rate of up to 70% and 8–10% of risk shared between siblings [
56]. HLA genetic susceptibility contributes to the loss of tolerance of β-cells antigens due to T-cell mediated islet cell death, insulin deficiency, and elevated blood glucose levels, suggesting that these genes operate at the β-cell level and play a crucial role in the pathogenesis of T1DM [
3,
57]. Despite initial thoughts, genetic predisposition alone cannot explain the incidence of T1DM, as recent work has identified that HLA loci account for 50% of familial T1DM [
4,
6,
53,
56,
58]. Now, T1DM is considered an intricate interaction and balance between the genome, metabolism, and gut microbiome diversity [
4,
6,
53,
59,
60,
61]. There are also a number of environmental influences, including diet, vitamin D deficiency, infections, pollutants, and oxidative stress, associated with the increasing presentation of type 1 diabetes, with β-cell stress potentially providing a mechanistic underpinning [
6,
7,
56,
59,
60]. In addition, comorbidities are often found in those diagnosed with T1DM, as the majority of T1DM patients (60%) are obese or overweight, while many others have hyperlipidemia and hypertension [
62]. Comorbidities can occur as complications of T1DM, with individuals with poorly regulated T1DM having a 10 times higher chance of dying from cardiovascular disease than healthy individuals [
4]. Though T1DM is slightly more common in males than females, females are more likely to experience complications of T1DM such as hypoglycemia, vascular events, and premature lethality in response to T1DM than males [
6,
62,
63].
The mechanisms of T1DM begin well before full diabetes, as T-cells attack insulin-producing β-cells (
Figure 2B) [
6,
7,
53]. During this prediabetes phase, the pancreas can maintain normal glucose processing. However, the amount of insulin that can be secreted in the body is reliant both upon the absolute number of β-cells and β-cell function [
46]. As the immune-mediated destruction of β-cells continues, this puts increasing demand and thus limits the function of the remaining β-cells. The body enters a state of hyperglycemia, with excessive levels of unused glucose in the bloodstream. In a healthy system, protective immune activated inflammation is terminated after a threat is eliminated, but the autoimmunity of T1DM leads to chronic inflammation, further promoting disease progression, β-cell apoptosis, and influencing insulin resistance [
64,
65]. Though some patients suffer from a complete loss of functional β-cells, measurable levels of C-peptide, which is the biologically inactive peptide formed when β-cells convert proinsulin to insulin, indicate that some T1DM patients can produce insulin; nevertheless, the functional β-cell mass is still decreased to the level of insulin-dependency [
2,
7]. In the 100 years since the discovery of insulin, knowledge of the T1DM pathology process, improved treatment options, and novel diagnostic measures were developed.
4.3. Role of EVs in Pancreas and Islet β-Cell Transplantation
The continuous glucose monitoring systems, unfortunately, are not capable of rapid responses to glycemic variability as might be produced by illness, stress, or a simple unannounced meal [
2]. Moreover, though insulin administration offers some regulatory abilities, many patients develop chronic complications or become disabled by refractory hypoglycemia [
66,
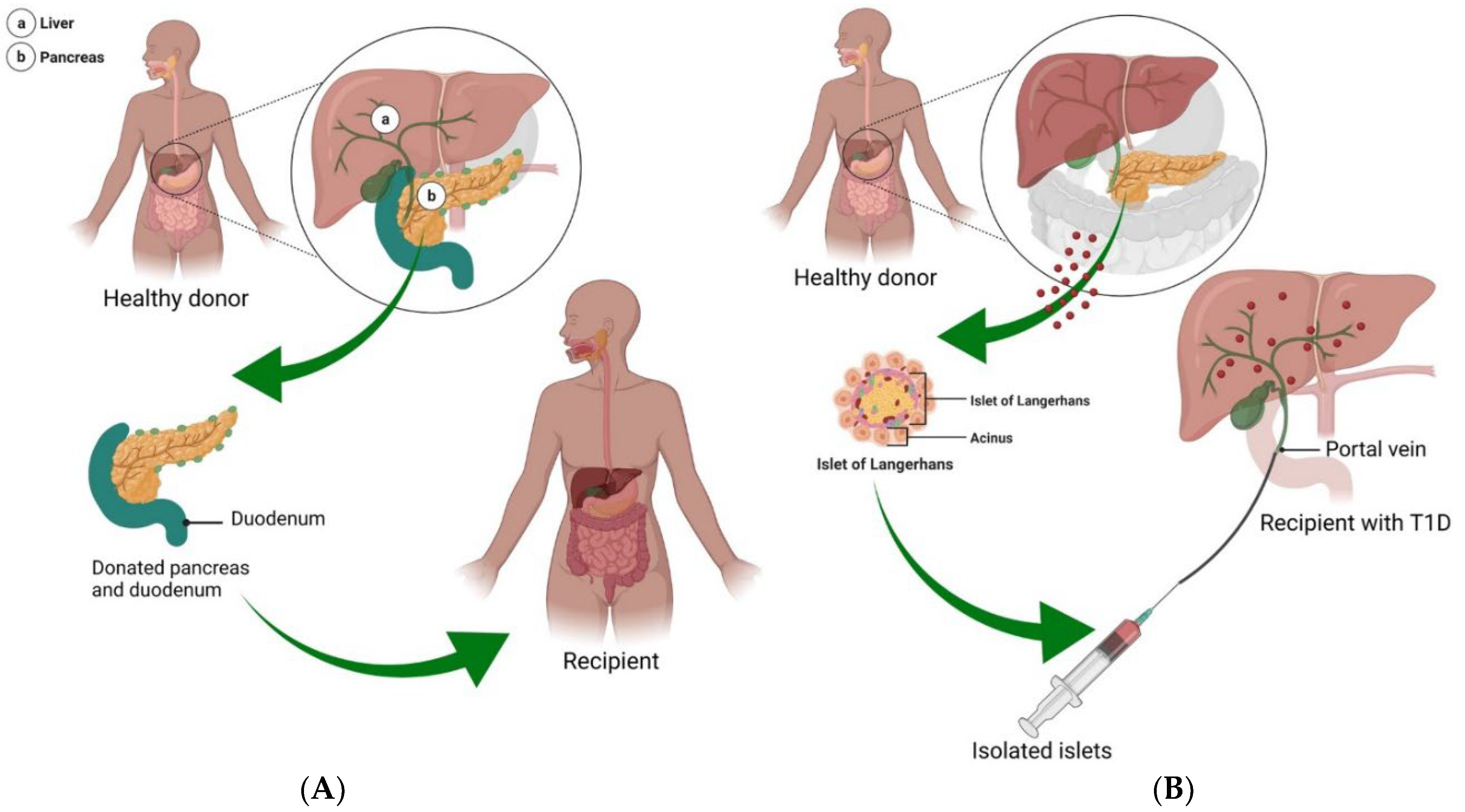
67]. An alternative treatment option is the transplantation of the pancreas or islet β-cells to prevent or slow the complications and progression of T1DM, improving longevity and quality of life (
Figure 3A). Though pancreas transplantations continued to evolve and develop over time (for the full record, see [
66]), the number of pancreas transplants are dropping. This is due to inherent surgery risk and improvements in other treatment options, with a 20% decrease in transplants in the USA between 2005 and 2014, transplants peaking in 2004 [
44,
68]. Transplantations as treatments for T1DM are generally only considered in patients who exhibit a history of hard to control diabetes, defined as ≥1 episode of severe hypoglycemia per year, frequent and severe metabolic or microvascular complications, and problems with exogenous insulin therapy [
69,
70,
71,
72]. As such, pancreas transplantations are typically only performed 20 years after diabetes diagnosis [
69]. Transplanting a pancreas can fully replace the β-cell mass and as such, is considered the only “near-cure” for T1DM, while transplanting islet β-cells can only partially restore the population of insulin-secreting cells needed for insulin and glucose regulation [
66]. The first documented pancreas transplants were performed in the 1960s–1970s on uremic diabetic patients in combination with a kidney transplant [
44,
48,
66]. Early surgeries found that transplanting a pancreas alone led to a high rate of organ rejection, while cases in which a pancreas and a kidney were simultaneously transplanted had minimal organ failure (though were susceptible to numerous other issues accompanying surgery such as thrombosis or infection) [
66]. Recent reports indicate that this graft survival trend has continued, with 70% graft survival in patients with simultaneous pancreas and kidney transplant, and only ~50% for pancreas in isolation [
2,
44,
68].
As an alternative to pancreas transplant, islet β-cell transplantation is a low-risk procedure, with donor islets infused into the liver via the portal vein (
Figure 3B) [
6,
73]. Though islet transplants are practiced internationally, this procedure is still considered experimental in the USA [
74]. Though islet transplantation does not replace the entire β-cell mass, it is less invasive than a pancreas transplant while still providing the potential to restore normal glycemic function, re-establish hypoglycemia awareness, and minimize severe hypoglycemic responses [
2,
67]. Islet transplants were first attempted in the 1970s, with very low success rates [
71]. The first fruitful islet transplantations occurred in the late 1980s and 1990s, with the ability to consistently establish endogenous insulin secretion via transplant taking place in the 2000s [
48]. The principal indications for β-cell replacement therapy are to treat a small subset of insulin-dependent T1DM and T2DM patients with severe or uncontrollable hypoglycemia [
6,
75]. An estimated 50% of transplant recipients maintain insulin independence one to two years post islet transplant [
45,
73,
76]. A recent approach in islet delivery is to encapsulate islets to protect them from the immunological environment, allowing greater survival and function [
2]. Islet cells may be transplanted in combination with hormones or growth factors such as glucagon-like peptide-1 to augment insulin secretion and inhibit glucagon release [
56,
77,
78].
There are several pieces of evidence that indicate that EVs may increase the survival and efficacy of pancreatic islet transplantation [
79]. The communication between β-cells and endothelial cells is imperative for islet transplantation because it helps the body re-grow new blood vessels. Figliolini et al. showed that EVs derived from human islets could be taken up by endothelial cells inside the islets and help with angiogenesis and engraftment, according to in vitro experiments [
80]. Poor graft vascularization limits the success of islet transplantation. Cantaluppi et al. investigated that EVs secreted by endothelial progenitor cells promoted human islet vascularization. EVs stimulated migration, proliferation, cell survival, and organization in vessel-like structures in islet endothelial cells [
81]. EVs derived from endothelial progenitor cells increased insulin secretion, survival, and angiogenesis of islets implanted in severe combined immunodeficiency (SCID) mice after incorporation into islet endothelium and β-cells [
81]. EVs can facilitate islet transplantation through immunomodulatory effects in addition to enhancing revascularization. Human bone marrow MSCs and their EVs have been shown to be capable of suppressing immunological responses and delivering small RNAs. As a result, they may be able to improve islet transplantation by delivering small RNAs that enhance islet function while preventing immunological rejection. EVs produced from MSCs are shown to promote islet transplantation by promoting regulatory T cell function and suppressing peripheral blood mononuclear cell proliferation, according to a study conducted by Wen et al. [
82]. The therapeutic cargo contained within EVs must be appropriately studied for health and safety issues. In conclusion, exosomes may portray an enthralling therapy in the context of islet transplantation, not only for enhancing revascularization but also for stimulating graft tolerance.
4.4. EVs and T1DM Pathogenesis
T1DM is a complex, multifactorial disease that is heavily influenced by cross-talk between metabolically active tissues. EVs are thought to be a major mediator of cellular communication and have been found to be increasingly important in disease pathogenesis, as exposure of materials from cells in crisis affects the survival of recipient cells [
38,
83,
84]. For example, EVs from obese or diabetic mouse models can induce insulin resistance and glucose intolerance, while EVs from healthy lean mice can reverse these symptoms [
85,
86]. EVs are shown to contain hormones and autoantigens that have a key role in insulin sensitivity and the initiation of the strong, detrimental autoimmune response in T1DM, including insulin receptor substrate 1, GAD65, IA-2, and proinsulin [
87,
88,
89,
90,
91,
92]. The injection of EVs containing autoantigens was found to increase cytokine release, stimulate Toll-like receptors, and facilitate graft rejection [
90,
93]. EVs containing high levels of autoantigens are released as the pancreas undergoes excess stress, as increased levels of glucose or treatment with proinflammatory cytokines were found to drastically increase EV secretion [
87,
89,
91]. Interestingly, macrophage-derived EVs are able to activate memory T-cells [
94]. In a similar fashion, EVs isolated from insulinomas can induce the secretion of inflammatory cytokines in diabetes mouse models through the MyD88-dependent pathway, which is downstream of inflammatory signals of Toll-like receptors and interleukin-1. Activation of MyD88 can trigger T-cell proliferation and activation, further stimulating detrimental autoimmune response in T1DM [
88,
95]. EVs also participate in a negative feedback loop of cytokine production, which further drives macrophage activation [
96].
Micro-RNAs (miRNAs) play key regulatory functions in various aspects of blood sugar regulation, such as embryonic development in pancreatic islets, maintenance of α- and β-cell phenotypes, insulin secretion, and exocytosis [
84]. The miRNA content of EVs can play a major role in propagating T1DM pathology. EVs secreted from cells exposed to cytokines led to apoptosis when delivered to β-cells, an effect that can be blocked through the inactivation of miRNA activity through inhibition of Ago2 (a component of the RISC complex that is essential for miRNA action) [
83]. Increased EV expression of miR-21, miR-34a, miR-146a, and miR-29 as triggered by exposure to cytokines early in disease contributes to the cytokine-induced β-cell dysfunction [
53]. The release of miR-375 and miR-148a-3p in EVs are associated with β-cell death [
97]. Similarly, EV-contained miR-142-3p/5p and miR-155 lead to the upregulation of cytokines in pancreatic islets β-cells, with in vitro inhibition of these miRNAs able to reduce apoptosis rate in primary rat islet cells [
90]. When splenocytes isolated from diabetes-prone mice are exposed to β-cell EVs, there is a miR-29b-dependent increase in the production and secretion of TNF-α [
90]. Further understanding of how the contents of EVs are modified in disease states can give insight into the spread of inflammation and pathology in T1DM.
4.5. EVs as Novel Biomarkers in T1DM
Several lines of evidence suggest that many factors induce T1DM, including genetic predisposing factors, non-infectious environment agents, endogenous antigens, exogenous infectious pathogens, and physiological stress [
4,
6,
56,
59,
60]. As such, the underlying autoimmune process of T1DM precedes the onset of clinical symptoms. Though the early stage of T1DM, called prediabetes, is asymptomatic, it provides an excellent advantage for early prognosis and prevention of disease [
98]. Currently, susceptibility genes (high-risk HLA genes; HLA-DR (DR3/4) and HLA-DQ (DQ8)) and hallmark islet autoantibodies (GAD65, IA-2, insulin, and ZnT8) are the gold-standard approaches for the detection of T1DM. Notably, a clinical study has revealed that HLA typing identified only 10% of study subjects with high T1DM risk while the remaining 90% displayed no autoimmunity, and less than 50% of cases were identified by a combination of genetic markers [
52,
53]. Further, a major limitation of diagnostic autoantibodies is that they appear relatively late during disease progression when almost 90% of β-cells are lost [
99,
100]. Additionally, a subset of individuals who showed seropositivity for autoantibodies will not develop clinical diabetes [
101]. Therefore, current diagnostic tools do not meet the need for early diagnosis, creating a great demand for the identification of a suitable biomarker for early risk prediction.
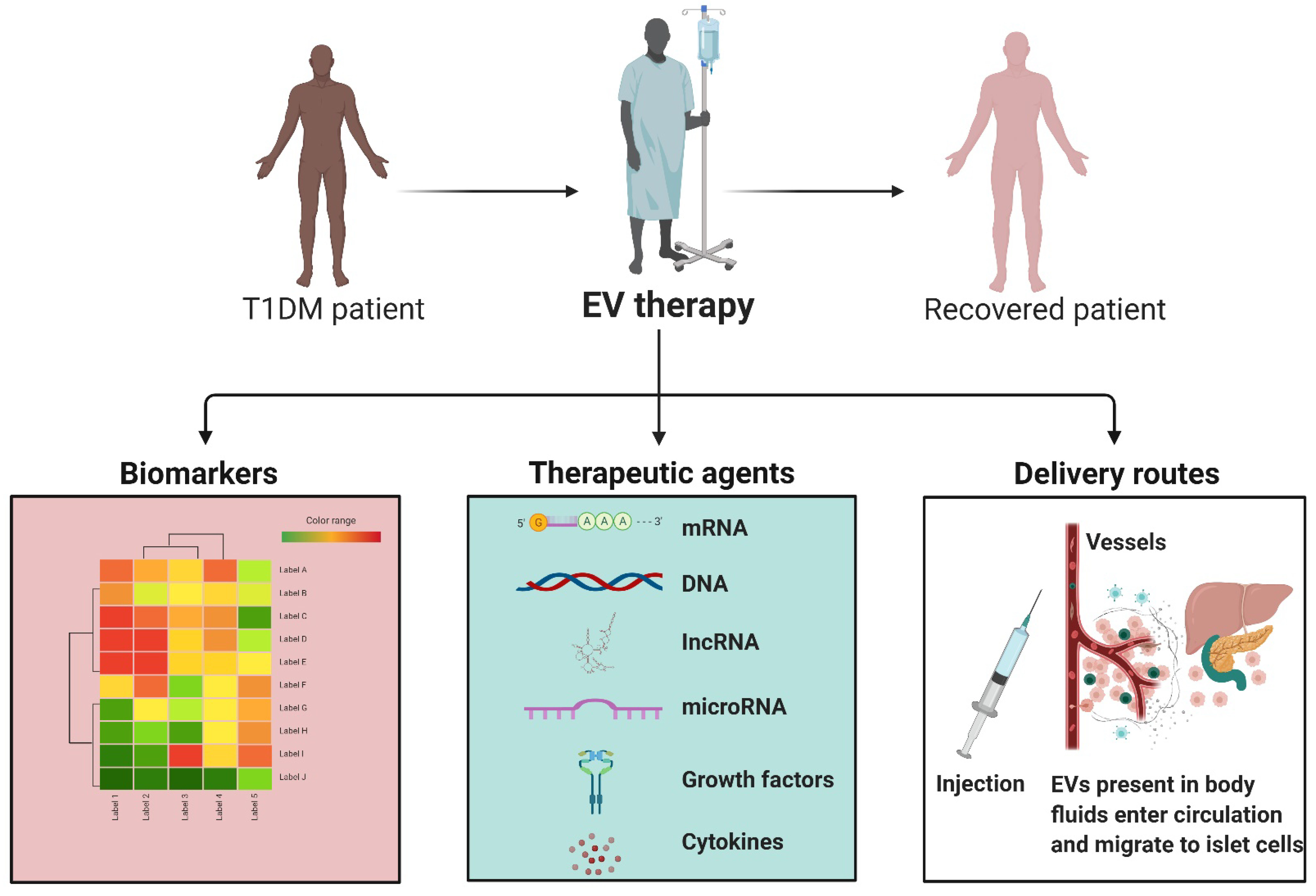
In recent years, several studies documented the potential utility of EV-based biomarkers in ailments such as liver disease, neurodegenerative diseases, and cardiac disorders [
102,
103,
104,
105,
106,
107]. EVs have easy accessibility from bodily fluids and molecular and structural stabilities, making them an excellent candidate for harboring biomarkers in clinical settings [
108]. In the context of T1DM, EVs drawn from blood can offer a new understanding of the status of sugar regulation (
Table 4). Evidence indicates that EVs secreted by human pancreatic insulin-producing β-cells may trigger and accelerate disease progression via stimulating autoimmune responses [
88,
109,
110]. Studies also implicated that the severity of T1DM and its pathophysiological conditions, such as inflammation, can alter the yield, content, and composition of EVs released from β-cells, information that can be utilized in the discovery of biomarkers [
47,
83,
108,
111,
112,
113,
114,
115,
116]. A study by Garcia-Contreas and colleagues found that T1DM patients had reduced plasma EV size and concentration compared to healthy controls [
108]. A recent report of EV RNA profiling of human islets from T1DM patients revealed modified EV RNA expression in response to proinflammatory cytokine treatment. Treating islets with proinflammatory cytokines mimics the chronic inflammation found in T1DM and results in the differential expression of RNAs associated with necrosis, insulin secretion, calcium signaling, and apoptosis, emphasizing the toxic effect of inflammation on islet survival [
117]. Krishnan and colleagues identified miR-155-5p as the most upregulated miRNA, while miR-4485 was the most downregulated [
117]. Significantly, islet-derived EVs act as antigen carriers, including autoantibodies GAD65, ZnT8, and β-cell glucose transporter 2, which are released in response to cellular stress, and serve as markers of T1DM progression [
87,
95,
97,
114,
118,
119,
120]. Importantly, islet EVs were found to specifically activate memory T-cell responses in diabetic patients, but not healthy controls [
119].
MiRNAs are crucial in blood sugar regulation, and a diverse population of miRNAs can be found within EVs [
84]. As such, EV miRNAs are considered attractive candidates for biomarkers to diagnose T1DM or determine the success of a treatment plan [
84,
121,
122]. A number of miRNAs are consistently upregulated in T1DM, including miR-21-5p, miR-100-5p, miR-150-5p, miR-181a-5p, miR-210-5p, and miR-375, many of which can be found in EVs [
53,
122]. Work studying young, newly diagnosed T1DM patients identified distinct EV miRNAs modifications compared to healthy controls, with downregulated levels of miR-195 and miR-455, while miR185 was upregulated [
115]. A study employed miRNA microarray and qPCR analysis to characterize plasma-derived EVs and identified a distinct miRNA profile found in patients with long-duration T1DM compared with healthy controls with seven differentially expressed miRNAs [
123]. Distinctions in EV miRNA content were also found in children with urinary EV miR-424 and miR-218, and serum EV miR-21-5p was found to be elevated in children with T1DM compared to healthy controls [
114,
124,
125]. Of vital importance, research has suggested that miRNAs such as miR-21-5p have detrimental impacts in T1DM and can be linked with β-cell inflammation and apoptosis [
126,
127,
128,
129]. Work studying young, newly diagnosed T1DM patients identified distinct EV miRNA modifications compared to healthy controls, with downregulated levels of miR-195 and miR-455, while miR185 was upregulated [
115]. Importantly, certain EV miRNAs are elevated in distinct timeframes and conditions; for example, work within a mouse model of diabetes found that miR-375 was elevated two weeks before the onset of diabetes and was thought to be a specific marker of deteriorating β-cell health and function [
53,
83,
84]. Studies reported elevated levels of β-cell miR-21-5p in EVs in response to inflammatory cytokines treatments, both in animal models of type 1 diabetes and in vitro, while total levels are decreased [
95,
126,
127,
128,
129].
There are a number of complications that can occur in diabetic patients, including diabetic nephropathy and retinopathy, a kidney disease that and complication damaging the blood vessels in the eye, respectively. Recent studies found that EVs and the miRNA content within EVs can aid in the early diagnoses of these complications [
84,
85,
130,
131,
132,
133,
134,
135]. Barutta and colleagues determined that miRNA-145 within urinary EVs is elevated in diabetic patients with diabetic nephropathy [
130], while Kalani et al. demonstrated that increased levels of Wilm’s tumor-1 protein within urinary EVs could predict a decline in renal function [
131]. Further, peroxisome proliferator-activated receptor-γ within EVs increases with disease severity of diabetic retinopathy, while miR-222 is decreased [
132]. Furthermore, certain populations of EVs have significantly elevated levels of creatine in response to hyperglycemic events in T1DM patients [
133].
In transplantation, the primary target time frame for both biomarkers and therapeutics is immediately after the graft to prevent surgical complications and graft rejection [
136]. In the instance of transplantation, the organ or cells release donor-specific EVs that are involved in immune recognition and often influence rejection or graft tolerance; in the case of graft rejection, there is a significant decrease in transplant islet EV signal, as well as changes in EV miRNA and proteomic profiles, displaying distinct biomarker potential [
137,
138,
139]. Notably, miRNAs in EVs can be beneficial in identifying successful T1DM treatments, as in many cases of malfunctioning transplantations, there are distinct upregulated and dysregulated miRNA and protein profiles that can be identified in EVs up to 5 years post-transplantation [
138,
140]. Additionally, studies identified that islet endothelial cells (IECs) and β-cells secrete a subset of miRNAs in EVs that take part in cross-talk between IECs and β-cells, thus promoting angiogenesis, a process limited in T1DM and inflammatory conditions, and often a limiting factor in transplantation efficacy [
80,
81,
82,
83,
140]. As a marker of vascularization, these miRNAs can identify early problems in transplantation. Time course studies found that EVs from islet cells contain varying miRNA in response to inflammation and hypoxia, with miR-29b-3p and miR-216a-5p released in EVs 6 h after hypoxia and cytokine exposure, while miR-375 and miR-148a-3p were secreted 24 h after exposure [
91]. Importantly, these stress conditions replicate negative conditions found in diabetic mice following islet transplantation [
91]. Cumulatively, this evidence suggests that EVs and EV miRNAs are promising biomarkers for early T1DM prognosis and development, as well as conditions following pancreas or islet transplant.
Table 4.
Studies utilizing EVs as biomarkers in diabetes.
Table 4.
Studies utilizing EVs as biomarkers in diabetes.
| Study | Experimental Subject | Collected Specimen | EV Isolation | EV Size | MSC EV Characterization | Biomarkers | Important Finding from the Studies |
|---|
| [141] | Rat | Blood | UC | NA | WB, FC | eNOS and caveolin-1 | Decreased levels of eNOS and overexpression of caveolin-1 may serve as a biomarker for vascular injury |
| [117] | Human | Human islet cell | UC | <150 nm | EM, WB | piRNAs, snoRNAs, tRNAs, and lncRNAs | EV miRNAs may consider as potential circulating biomarkers for T1DM |
| [139] | Human | Plasma samples | SEC, UC | NA | WB | Islet endocrine hormone proteins and mRNAs | Circulating transplant islet-specific EVs has the potential to be a diagnostic tool for recurrent autoimmune T1DM after islet transplantation |
| [137] | Mice and Human | Human plasma and urine samples | SEC, UC | 30–200 nm | NTA, EM, WB | miRNA | EV miRNAs as biomarkers for monitoring immune rejection |
| [142] | Human | Urine samples | UC | NA | WB | Water channel aquaporins | Role of water channel aquaporins AQP5 and AQP2 as novel biomarkers to help in classifying the clinical stage of diabetic nephropathy |
| [133] | Human | Urine samples | UC | 40–100 nm | EM, NTA | Urinary podocyte | Analysis of urinary podocyte MPs may act as an early biomarker of glomerular injury in uncomplicated T1DM |
| [143] | Human | Plasma samples | UC | NA | NA | Cytokines and angiogenic factors | EVs isolated from plasma shows upregulated levels of cytokines and angiogenic agents in diabetic patients. |
| [134] | Human | Urine samples | UC, Filtration | NA | WB | Cystatin B and altered protease profiles | Enhanced cystatin B and altered protease profiles in EVs isolated from urine may act as biomarkers of kidney dysfunction in T1DM |
| [144] | Rat and Human | Kidney tissue and Urine samples | UC | NA | WB | Regucalcin | Lower levels of urinary exosomal regucalcin may act as a biomarker of diabetic kidney disease |
| [130] | Human | Urine samples | UC | <100 nm | EM, NTA, WB | miRNAs | EV miR-145 may act as a biomarker of T1DM |
| [131] | Human | Urine samples | UC | NA | WB | WT1 protein | Elevated expression of EV WT1 protein may act as a biomarker of T1DM |
4.6. Paracrine Effects of EVs on Islet Cells
As carriers of vital proteins, miRNAs, and other contents, EVs have natural therapeutic benefits in various diseases, including T1DM (
Table 5) [
145]. Research found that the administration of EVs can help preserve organs in donor transplantation and aid in minimizing autoimmunity and cytokine release, thus minimizing immune rejection, improving viability and long-term survival [
63,
82,
145,
146,
147]. Specifically, EVs derived from mesenchymal stem cells (MSCs) were found to suppress the development of T-cells and slow disease development [
63,
114,
145]. EVs naturally contain a number of valuable proteins and miRNAs but can also be used to deliver select cargoes, either through modification of the parent cells or through specific loading of therapeutic content [
148,
149]. Wen et al. found that cell modifications in which EVs delivered plasmids to silence Fas and miR-375 in human islets improved resilience against inflammatory cytokines and suppressed immune reaction [
82]. Application of MSC EVs, through the delivery and upregulation of vascular endothelial growth factor (VEGF), promoted insulin secretion in islets, as well as downregulated pro-apoptotic genes, upregulated pro-survival factors, and increased angiogenesis, thus targeting primary causes of graft loss [
80,
150]. MSC-derived EVs were found to be protective in diseases, including T1DM, serving to block apoptosis, a major issue in graft survival, through the inhibition of the phosphorylation of p38 [
151]. In a mouse model of diabetes induced using streptozotocin, the delivery of bone-marrow MSC-derived EVs aided in the regeneration of islet cells [
147,
152]. EVs from adipocyte-derived stem cells can promote angiogenesis and modulate the immune response, attenuating T1DM pathology [
95]. Delivery of EVs derived from endothelial progenitor cells can enhance human islet vascularization, promoting insulin secretion and islet survival [
81], and it is hypothesized that EV delivery early in T1DM may help restore a patient’s original islet cells [
153]. Adipocyte-derived stem cell EVs modulate the immune response and promote proangiogenic properties, a vital aid in transplant survival [
95]. EVs derived from bone-marrow-MSCs have shown the ability to ameliorate insulin resistance developed through aging and T2DM through the delivery of miR-29b [
154].
In addition, EVs from a number of cell types, including MSCs, can have therapeutic impacts on the many complications that surround T1DM, including cognitive deficits, cardiac dysfunction, microvascular complications, and even transplantation rejection [
155,
156,
157,
158]. EVs loaded with miR-222 injected into rabbits with T1DM, and diabetic retinopathy showed attenuation of retinal degeneration [
159]. Diabetic nephropathy was improved through the delivery of EVs derived from MSCs and adipose-derived stem cells by induction of autophagy [
85,
160,
161] Though a number of studies regarding EV therapeutics use a heterogeneous population of EVs, a study by McGuinness and team (2016) found that microvesicles specifically stimulated long-term recovery and function in diabetic mice, improving glucose, insulin, and glucagon levels compared to the smaller EV population exosomes [
162].
Finally, many pancreas and islet transplant recipients receive treatments to aid in successful transplantation that, if loaded into EVs, may be better protected and enhance impact. For example, IL-2 receptor antibodies to provide better early posttransplant protection of islets and coadministration of a TNF-α inhibitor can lead to a higher probability of insulin independence, two cargos that may benefit from protective EV delivery [
67,
78]. The overexpression and delivery of insulin-like growth factor 1 and protein 3 gamma not only enhance the proliferation of β-cells but demonstrate the ability to regenerate and restore β-cells [
56]. The restoration and regeneration of pancreatic islet cells are regulated in part through EV interaction with the pancreatic and duodenal homeobox-1 pathway, which is vital in pancreatic cell differentiation [
163,
164]. Further, MSC-EVs at the time of islet transplantation are shown to improve islet functions through the delivery of necessary nutrients and thus transplant tolerance [
147,
165]. The delivery of EV-loaded cargos is not limited to T1DM. Glucagon-like peptide-1 is a hormone commonly used in T2DM to augment insulin secretion, aiding in glycemic index and insulin independence, can be loaded into EVs for better, more protected distribution [
77].
Table 5.
EVs as therapeutics in T1DM.
Table 5.
EVs as therapeutics in T1DM.
| Study | MSC Source | MSC EV Isolation | EV Size | MSC EV Characterization | Model Species/Cells | Intervention(s), Route, and Dose | Important Finding from the Studies |
|---|
| [166] | Bone marrow-derived MSCs | NA | NA | NA | Rat model of T1DM | Tail vein injection | EV miR-145 secreted by bone morrow MSCs shows neurorestorative effects in diabetic rats with stroke |
| [167] | Human urine-derived stem cells | UC 30% sucrose/D2O cushion | 50–100 nm | Flow cytometry, WB, NTA | Adult male Sprague Dawley (SD) rats | Weekly with 100 μg of EV dissolved in PBS to a final volume of 200 μL via the tail vein | Reduction of the urine volume and urinary microalbumin excretion; prevention of podocyte and tubular epithelial cell apoptosis in diabetic rats |
| [152] | Bone marrow-derived MSCs | UC | 100 nm | FC, EM | Albino female rat (STZ-induced rat model of T1DM) | Intraperitoneal injection | EVs derived from MSCs showed therapeutic and regenerative effects upon the pancreatic islet cells |
| [168] | Human umbilical cord blood-derivedendothelial progenitor cells | UC | 50–60 nm | Tunable resistive pulse sensing analysis, EM, WB, NTA | Adult male SD rats | Subcutaneous injection with EVs 2 × 1010 or 1 × 1011 particles, dissolved in 200 μL of PBS | Increased angiogenesis through Erk1/2 signaling |
| [169] | Human umbilical cord blood-derivedendothelial progenitor cells | UC | 40 to 80 nm | EM, WB | Adult male SD rats | Subcutaneous injection at wound sites with 100 μL PBS or EPC-EVs (100 μg) around the wounds | Enhanced wound healing by regulating vascular endothelial cells function |
| [170] | Adipose derived-MSCs | ExoQuick | 200 nm | EM, WB | 10-week-old SD rats | Intraperitoneal injection | Adipose tissue-derived MSC EVs enhanced the erectile function in diabetic rats |
| [155] | Bone-marrow MSCs | UC | NA | EM, WB | Streptozotocin-Induced Diabetic Nude Mouse Model | Intracerebroventricular administration PKH-labeled EVs (5 µg in 10 µL aCSF) | Improvement of cognitive impairments by repairing damaged neurons and astrocytes |
| [171] | Cardiomyocytes overexpressing HSP70 MSCs | UC | 50–60 nm | EM, WB, NTA | Streptozotocin-Induced Diabetic Nude Mouse Model | Intraperitonial injections | Enhancement of Hsp20 in cardiomyocytes can give protection in diabetic hearts via the release of EVs |
| [172] | Human bone marrow-derived MSCs | ExoQuick, Precipitation | NA | NA | Male C57BL/6J mice | Intravenous injections | Attenuation of renal fibrosis |
| [173] | Human fibrocytes | UC | 50–100 nm | EM, WB, NTA, FC | Diabetic B6.Leprdb/db mice (11–12 weeks old) | 200 μL of PBS containing 0, 5 or 50 μg EV; 40 μL subcutaneously injected around the wounds sites; 40 μL were directly applied to the wound beds | Enhanced proliferation and migration of diabetic keratinocyte; increased wound closure |
| [174] | Mouse serum | ExoQuick | NA | NA | C57BL/6J mice, RIP-CreER mice, and Rosa26-GNZ mice | Intraperitoneal and intravenous injections | Increased pancreatic beta-cell proliferation |
| [82] | Human bone marrow MSCs | Total exosome isolation reagent (Invitrogen) | NA | NA | NSG Mice (NOD scid gamma) | Intraperitoneal infusion | Inhibition of immune rejection |
| [81] | Endothelial progenitor cells isolated from PBMCs | UC | NA | FC, EM | Immunodeficient (SCID) Mice | Subcutaneous implantation | Enhancement of neo-angiogenesis of human pancreatic islets |
| [175] | Adipose-derived MSCs | UC | 40–100 nm | FC, EM, DLS, SEM | 6- to 8-week old C57BL/6 male mice | Intraperitoneal injection | Enhanced regulatory T-cell population without change in the proliferation index of lymphocytes |
| [110] | Human MSCs | UC | NA | NTA | NA | NA | MSC-derived MVs suppress inflammatory T-cell responses in the islet antigens through the promotion of regulatory dendritic cells in T1DM |
| [80] | Human islets | UC | 54–256 nm | FC, EM, WB, SEM | NA | NA | Human islet-derived EVs participate in beta cell-endothelium cross-talk and the neoangiogenesis process |
| [163] | Menstrual blood-derived MSC | Exo-spin kit | 30–150 µm | AFM, FE-SEM, WB | Male Wistar Rats (STZ-induced rat model of T1DMM) | Intravenous injection | EVs isolated from stem cells may regenerate beta-cells of islets through the Pdx-1 pathway |




{kind=link}
{kind=link}
{kind=link}
{kind=link}



