GAT1 Gene, the GATA Transcription Activator, Regulates the Production of Higher Alcohol during Wheat Beer Fermentation by Saccharomyces cerevisiae

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Strains and Plasmids
2.2. Medium and Culture Conditions
2.3. Yeast Transformation and Screening
2.4. Quantitative Real-Time PCR (RT-qPCR)
2.5. Fermentation Trials
2.6. Statistical Analysis
3. Results
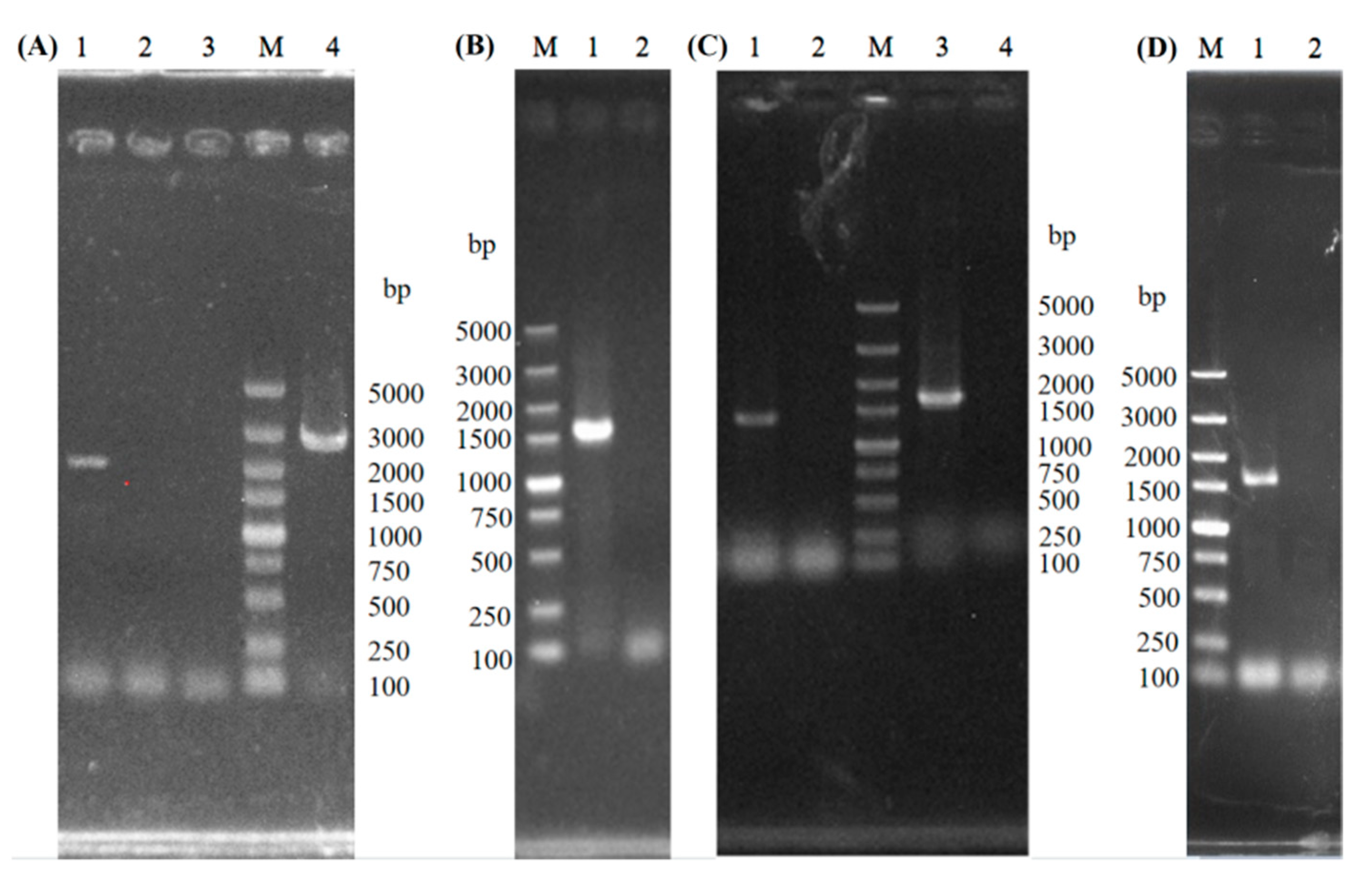
3.1. Construction of GAT1 Deleted Strains
3.2. Effects of GAT1 Deletion on Higher Alcohol Metabolism
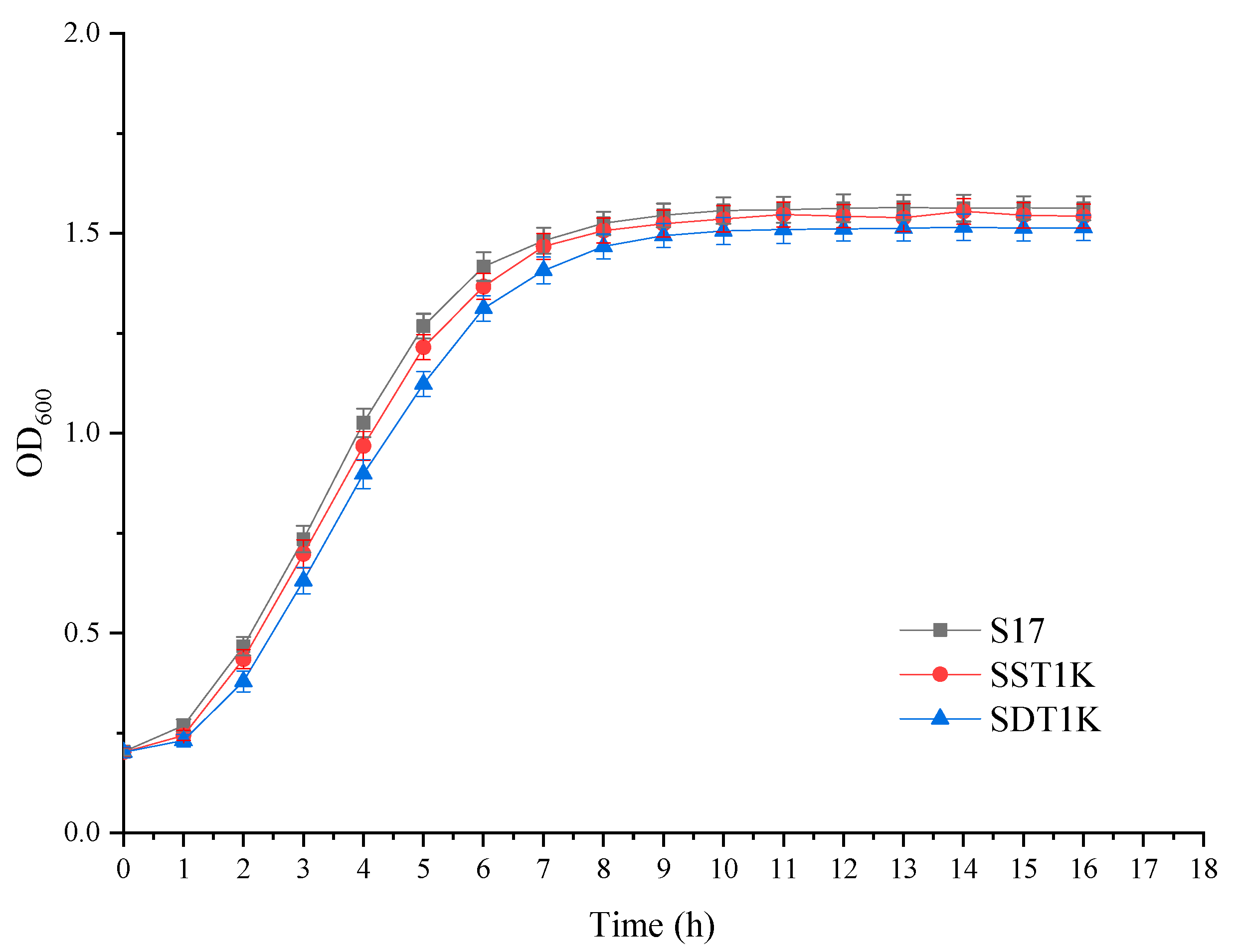
3.3. Effects of GAT1 Deletion on the Growth and Fermentation Performance of Yeast
3.4. mRNA Levels of Genes Related to Higher Alcohol Metabolism
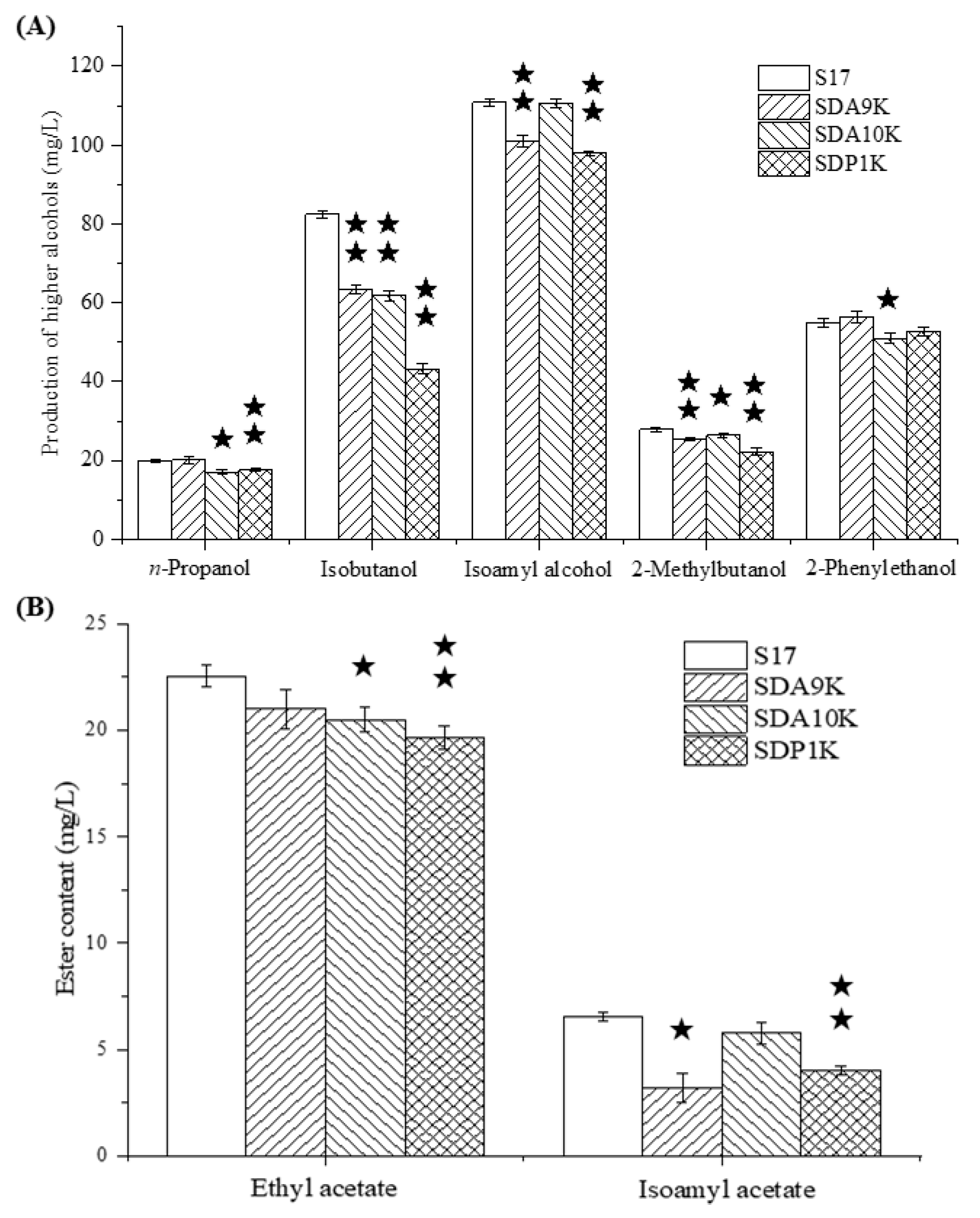
3.5. Effects of Deleting Target Genes of GAT1 on the Content of Higher Alcohols in Wheat Beer
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Faltermaier, A.; Waters, D.; Becker, T.; Arendt, E.; Gastl, M. Common wheat (Triticum aestivum L) and its use as a brewing cereal-a review. J. Inst. Brew. 2014, 120, 1–15. [Google Scholar] [CrossRef]
- Cui, Y.; Wang, A.; Zhang, Z.; Speers, R.A. Enhancing the levels of 4-vinylguaiacol and 4-vinylphenol in pilot-scale top-fermented wheat beers by response surface methodology. J. Inst. Brew. 2015, 121, 129–136. [Google Scholar] [CrossRef]
- Liguori, L.; De Francesco, G.; Russo, P.; Perretti, G.; Albanese, D.; Di Matteo, M. Quality attributes of low-alcohol top-fermented beers produced by membrane contactor. Food Bioprocess Technol. 2016, 9, 191–200. [Google Scholar] [CrossRef]
- Langos, D.; Granvogl, M.; Schieberle, P. Characterization of the key aroma compounds in two bavarian wheat beers by means of the sensomics approach. J. Agric. Food Chem. 2013, 61, 11303–11311. [Google Scholar] [CrossRef] [PubMed]
- Hazelwood, L.A.; Daran, J.; van Maris, A.J.A.; Pronk, J.T.; Dickinson, J.R. The Ehrlich pathway for fusel alcohol production: A century of research on Saccharomyces cerevisiae metabolism. Appl. Environ. Microbiol. 2008, 74, 2259–2266. [Google Scholar] [CrossRef]
- El-Dalatony, M.M.; Saha, S.; Govindwar, S.P.; Abou-Shanab, R.A.I.; Jeon, B. Biological conversion of amino acids to higher alcohols. Trends Biotechnol. 2019, 37, 855–869. [Google Scholar] [CrossRef] [PubMed]
- Kuroda, K.; Ueda, M. Cellular and molecular engineering of yeast Saccharomyces cerevisiae for advanced biobutanol production. FEMS Microbiol. Lett. 2016, 363, fnv247. [Google Scholar] [CrossRef]
- Milne, N.; van Maris, A.J.A.; Pronk, J.T.; Daran, J.M. Comparative assessment of native and heterologous 2-oxo acid decarboxylases for application in isobutanol production by Saccharomyces cerevisiae. Biotechnol. Biofuels 2015, 8, 1–15. [Google Scholar] [CrossRef]
- Park, S.H.; Kim, S.; Hahn, J.S. Improvement of isobutanol production in Saccharomyces cerevisiae by increasing mitochondrial import of pyruvate through mitochondrial pyruvate carrier. Appl. Microbiol. Biotechnol. 2016, 100, 7591–7598. [Google Scholar] [CrossRef]
- Shen, L.; Nishimura, Y.; Matsuda, F.; Ishii, J.; Kondo, A. Overexpressing enzymes of the Ehrlich pathway and deleting genes of the competing pathway in Saccharomyces cerevisiae for increasing 2-phenylethanol production from glucose. J. Biosci. Bioeng. 2016, 122, 34–39. [Google Scholar] [CrossRef]
- Yuan, J.; Mishra, P.; Ching, C.B. Engineering the leucine biosynthetic pathway for isoamyl alcohol overproduction in Saccharomyces cerevisiae. J. Ind. Microbiol. Biotechnol. 2017, 44, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Liu, Y.; Li, H.; Yang, Q.; Wu, Q.; Chen, S.; Tang, J.; Xu, Y. Modeling and regulation of higher alcohol production through the combined effects of the C/N ratio and microbial interaction. J. Agric. Food Chem. 2019, 67, 10694–10701. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Bai, X.; Guo, X.; He, X. Regulation of crucial enzymes and transcription factors on 2-phenylethanol biosynthesis via Ehrlich pathway in Saccharomyces cerevisiae. J. Ind. Microbiol. Biotechnol. 2017, 44, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Cooper, T.G. Transmitting the signal of excess nitrogen in Saccharomyces cerevisiae from the Tor proteins to the GATA factors: Connecting the dots. FEMS Microbiol. Rev. 2002, 26, 223–238. [Google Scholar] [CrossRef]
- Lee, K.; Hahn, J.S. Interplay of Aro80 and GATA activators in regulation of genes for catabolism of aromatic amino acids in Saccharomyces cerevisiae. Mol. Microbiol. 2013, 88, 1120–1134. [Google Scholar] [CrossRef]
- Cain, N.E.; Kaiser, C.A.; Riezman, H. Transport activity-dependent intracellular sorting of the yeast general amino acid permease. Mol. Biol. Cell 2011, 22, 1919–1929. [Google Scholar] [CrossRef]
- Iraqui, I.; Vissers, S.; André, B.; Urrestarazu, A. Transcriptional induction by aromatic amino acids in Saccharomyces cerevisiae. Mol. Cell Biol. 1999, 19, 3360–3371. [Google Scholar] [CrossRef]
- Güldener, U.; Heck, S.; Fiedler, T.; Beinhauer, J.; Hegemann, J.H. A new efficient gene disruption cassette for repeated use in budding yeast. Nucleic Acids Res. 1996, 24, 2519–2524. [Google Scholar] [CrossRef]
- Lu, J.; Dong, J.; Wu, D.; Chen, Y.; Guo, X.; Shi, Y.; Sun, X.; Xiao, D. Construction of recombinant industrial brewer’s yeast with lower diacetyl production and proteinase A activity. Eur. Food Res. Technol. 2012, 235, 951–961. [Google Scholar] [CrossRef]
- Ma, L.; Shi, Y.; Du, L.; Tang, P.; Xiao, D. Reduced production of higher alcohols by Saccharomyces cerevisiae in red wine fermentation by simultaneously overexpressing BAT1 and deleting BAT2. J. Agric. Food Chem. 2017, 65, 6936–6942. [Google Scholar] [CrossRef]
- Sun, Z.; Wang, M.; Wang, Y.; Xing, S.; Hong, K.; Chen, Y.; Guo, X.; Xiao, D. Identification by comparative transcriptomics of core regulatory genes for higher alcohol production in a top-fermenting yeast at different temperatures in beer fermentation. Appl. Microbiol. Biotechnol. 2019, 103, 4917–4929. [Google Scholar] [CrossRef]
- Gietz, R.; Schiestl, R. Large-scale high-efficiency yeast transformation using the LiAc/SS carrier DNA/PEG method. Nat. Protoc. 2007, 2, 38–41. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Chen, S.; Wang, J.; Zhang, C.; Shi, Y.; Guo, X.; Chen, Y.; Xiao, D. Genetic engineering to alter carbon flux for various higher alcohol productions by Saccharomyces cerevisiae for Chinese Baijiu fermentation. Appl. Microbiol. Biotechnol. 2018, 102, 1783–1795. [Google Scholar] [CrossRef] [PubMed]
- Chiva, R.; Baiges, I.; Mas, A.; Guillamon, J.M. The role of GAP1 gene in the nitrogen metabolism of Saccharomyces cerevisiae during wine fermentation. J. Appl. Microbiol. 2009, 107, 235–244. [Google Scholar] [CrossRef]
- Kneen, M.M.; Stan, R.; Yep, A.; Tyler, R.P.; Saehuan, C.; McLeish, M.J. Characterization of a thiamin diphosphate-dependent phenylpyruvate decarboxylase from Saccharomyces cerevisiae. FEBS J. 2011, 278, 1842–1853. [Google Scholar] [CrossRef] [PubMed]
- Dickinson, J.R.; Harrison, S.J.; Dickinson, J.A.; Hewlins, M.J.E. An investigation of the metabolism of isoleucine to active amyl alcohol in Saccharomyces cerevisiae. J. Biol. Chem. 2000, 275, 10937–10942. [Google Scholar] [CrossRef] [PubMed]
- Dickinson, J.R.; Salgado, L.E.; Hewlins, M.J. The catabolism of amino acids to long chain and complex alcohols in Saccharomyces cerevisiae. J. Biol. Chem. 2003, 278, 8028–8034. [Google Scholar] [CrossRef]
- Yin, S.; Zhou, H.; Xiao, X.; Lang, T.; Liang, J.; Wang, C. Improving 2-Phenylethanol Production via Ehrlich Pathway Using Genetic Engineered Saccharomyces cerevisiae Strains. Curr. Microbiol. 2015, 70, 762–767. [Google Scholar] [CrossRef]
- Kim, B.; Cho, B.; Hahn, J. Metabolic engineering of Saccharomyces cerevisiae for the production of 2-phenylethanol via Ehrlich pathway. Biotechnol. Bioeng. 2014, 111, 115–124. [Google Scholar] [CrossRef]
- Chen, H.; Huang, H.; Li, X.; Tong, S.; Niu, L.; Teng, M. Crystallization and preliminary X-ray diffraction analysis of ARO9, an aromatic aminotransferase from Saccharomyces cerevisiae. Protein Pept. Lett. 2009, 16, 450. [Google Scholar] [CrossRef]
- Iraqui, I.; Vissers, S.; Cartiaux, M.; Urrestarazu, A. Characterisation of Saccharomyces cerevisiae ARO8 and ARO9 genes encoding aromatic aminotransferases I and II reveals a new aminotransferase subfamily. Mol. Gen. Genet. 1998, 257, 238–248. [Google Scholar] [CrossRef] [PubMed]
- Pires, E.J.; Teixeira, J.A.; Brányik, T.; Vicente, A.A. Yeast: The soul of beer’s aroma—A review of flavour-active esters and higher alcohols produced by the brewing yeast. Appl. Microbiol. Biotechnol. 2014, 98, 1937–1949. [Google Scholar] [CrossRef] [PubMed]
- Procopio, S.; Qian, F.; Becker, T. Function and regulation of yeast genes involved in higher alcohol and ester metabolism during beverage fermentation. Eur. Food Res. Technol. 2011, 233, 721–729. [Google Scholar] [CrossRef]
- Vanderhaegen, B.; Neven, H.; Coghe, S.; Verstrepen, K.J.; Verachtert, H.; Derdelinckx, G. Evolution of Chemical and Sensory Properties during Aging of Top-Fermented Beer. J. Agric. Food Chem. 2003, 51, 6782–6790. [Google Scholar] [CrossRef]
- Li, W.; Wang, J.; Zhang, C.; Ma, H.; Xiao, D. Regulation of Saccharomyces cerevisiae genetic engineering on the production of acetate esters and higher alcohols during Chinese Baijiu fermentation. J. Ind. Microbiol. Biotechnol. 2017, 44, 949–960. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Qi, Y.; Ma, H.; Li, W.; Dai, L.; Xiao, D. Decreased production of higher alcohols by Saccharomyces cerevisiae for Chinese rice wine fermentation by deletion of Bat aminotransferases. J. Ind. Microbiol. Biotechnol. 2015, 42, 617–625. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains or Plasmids | Relevant Characteristic | References or Source |
|---|---|---|
| Strains | ||
| S17, CICC1929 | Wild-type top-fermenting industrial brewer’s strain | This study |
| E. coli DH5α | Host of plasmid | Stratagene |
| Transformants | ||
| SST1 | S-17GAT1(n-1)/ΔGAT1::loxP-KanMX-loxP | This study |
| SST1K | S-17GAT1(n-1)/ΔGAT1::loxP::loxP | This study |
| SDT1 | S-17GAT1(n-2)/ΔGAT1::loxP-KanMX-loxP | This study |
| SDT1K | S-17GAT1(n-2)/ΔGAT1::loxP::loxP | This study |
| SSP1 | S-17GAP1(n-1)/ΔGAP1::loxP-KanMX-loxP | This study |
| SSP1K | S-17GAP1(n-1)/ΔGAP1::loxP::loxP | This study |
| SDP1 | S-17GAP1(n-2)/ΔGAP1::loxP-KanMX-loxP | This study |
| SDP1K | S-17GAP1(n-2)/ΔGAP1::loxP::loxP | This study |
| SSA9 | S-17ARO9(n-1)/ΔARO9::loxP-KanMX-loxP | This study |
| SSA9K | S-17ARO9(n-1)/ΔARO9::loxP::loxP | This study |
| SDA9 | S-17ARO9(n-2)/ΔARO9::loxP-KanMX-loxP | This study |
| SDA9K | S-17ARO9(n-2)/ΔARO9::loxP::loxP | This study |
| SSA10 | S-17ARO10(n-1)/ΔARO10:: loxP-KanMX-loxP | This study |
| SSA10K | S-17ARO10(n-1)/ΔARO10::loxP::loxP | This study |
| SDA10 | S-17ARO10(n-2)/ΔARO10::loxP-KanMX-loxP | This study |
| SDA10K | S-17ARO10(n-2)/ΔARO10::loxP::loxP | This study |
| Plasmids | ||
| pUG6 | Kan r, containing loxP-KanMX-loxP disruption cassette | [18] |
| pSH-Zeocin | Zeo r, Cre recombinant enzyme expression vector | [19] |
| Strains | Weight Loss of CO2 (g) | Ethanol (% v/v, 20 °C) | Residual Sugar (g/L) | Residual FAN (mg/L) | Real Fermentation Degree (%) |
|---|---|---|---|---|---|
| S17 | 5.77 ± 0.058 | 4.04 ± 0.0208 | 5.93 ± 0.058 | 141.88 ± 0.235 | 68.37 ± 0.112 |
| SST1K | 5.70 ± 0.000 | 4.03 ± 0.0451 | 6.03 ± 0.058 | 158.67 ± 0.359 ★★ | 68.25 ± 0.190 |
| SDT1K | 5.77 ± 0.058 | 4.04 ± 0.0513 | 5.90 ± 0.000 | 182.04 ± 0.136 ★★ | 68.22 ± 0.378 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.-P.; Liu, L.; Wang, X.-S.; Hong, K.-Q.; Zhang, L.-H.; Sun, Z.-G.; Xiao, D.-G. GAT1 Gene, the GATA Transcription Activator, Regulates the Production of Higher Alcohol during Wheat Beer Fermentation by Saccharomyces cerevisiae. Bioengineering 2021, 8, 61. https://doi.org/10.3390/bioengineering8050061
Wang Y-P, Liu L, Wang X-S, Hong K-Q, Zhang L-H, Sun Z-G, Xiao D-G. GAT1 Gene, the GATA Transcription Activator, Regulates the Production of Higher Alcohol during Wheat Beer Fermentation by Saccharomyces cerevisiae. Bioengineering. 2021; 8(5):61. https://doi.org/10.3390/bioengineering8050061
Chicago/Turabian StyleWang, Ya-Ping, Lin Liu, Xue-Shan Wang, Kun-Qiang Hong, Li-Hua Zhang, Zhong-Guan Sun, and Dong-Guang Xiao. 2021. "GAT1 Gene, the GATA Transcription Activator, Regulates the Production of Higher Alcohol during Wheat Beer Fermentation by Saccharomyces cerevisiae" Bioengineering 8, no. 5: 61. https://doi.org/10.3390/bioengineering8050061
APA StyleWang, Y.-P., Liu, L., Wang, X.-S., Hong, K.-Q., Zhang, L.-H., Sun, Z.-G., & Xiao, D.-G. (2021). GAT1 Gene, the GATA Transcription Activator, Regulates the Production of Higher Alcohol during Wheat Beer Fermentation by Saccharomyces cerevisiae. Bioengineering, 8(5), 61. https://doi.org/10.3390/bioengineering8050061