
Navigating the Collagen Jungle: The Biomedical Potential of Fiber Organization in Cancer

 ,
,
Abstract
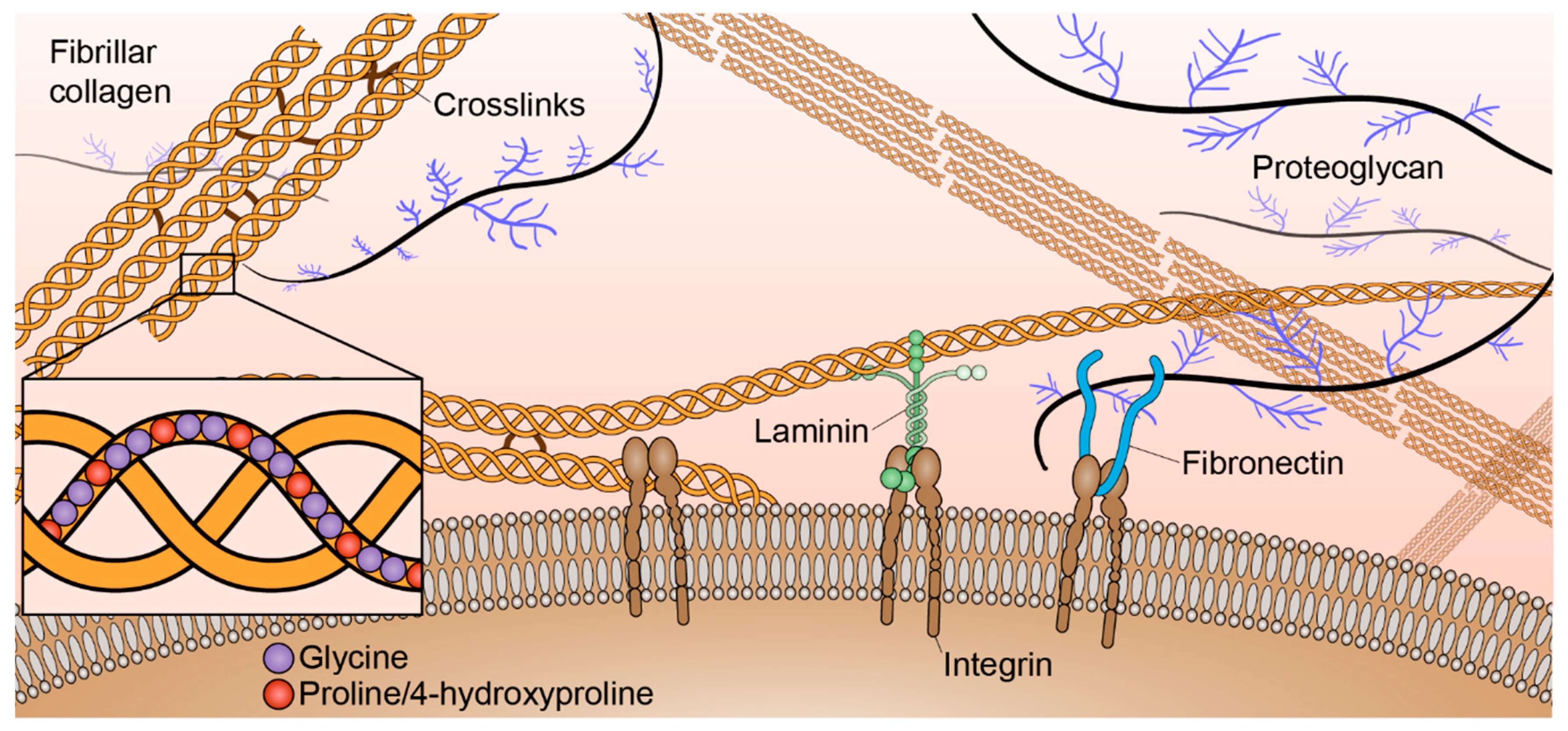
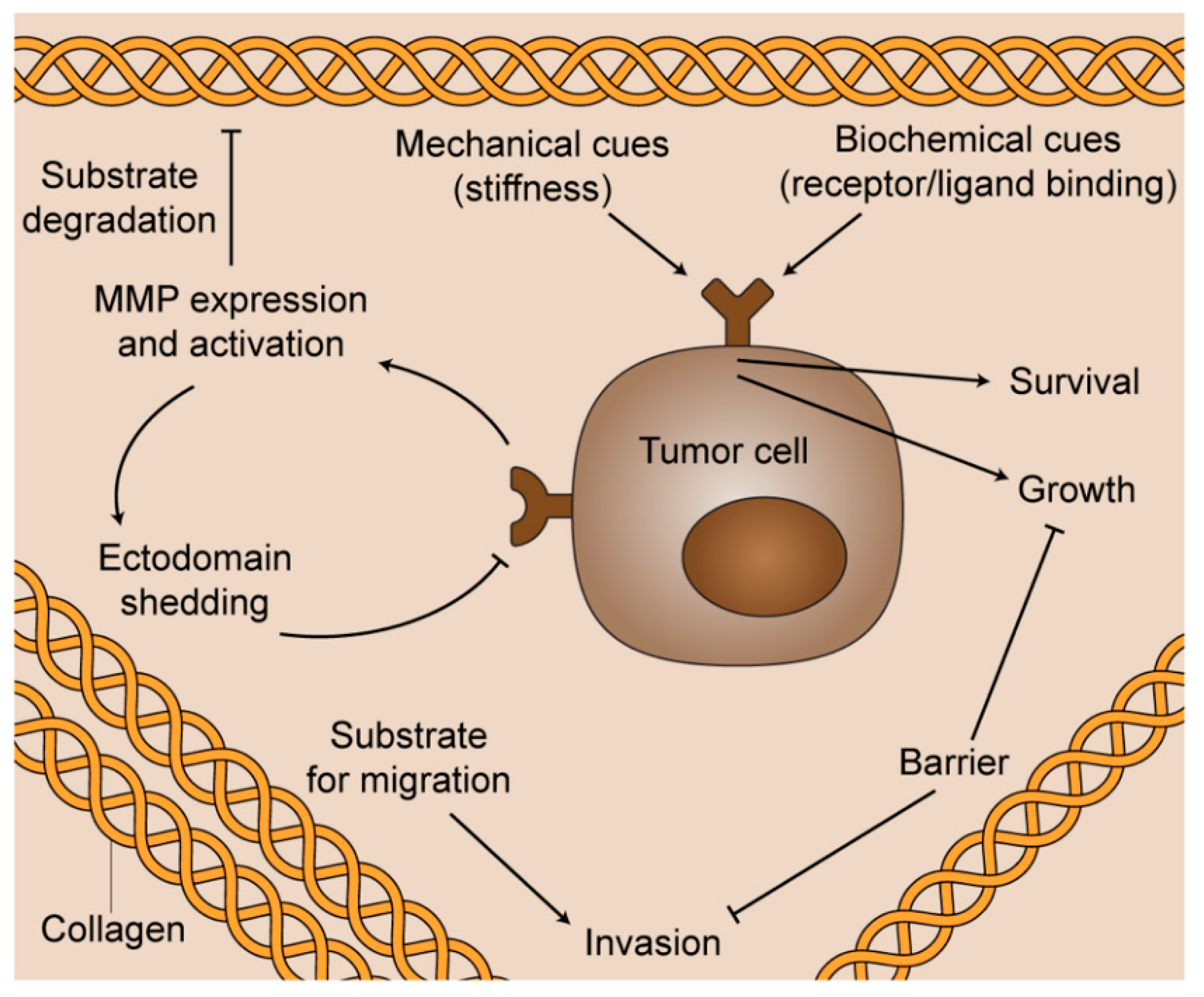
1. Introduction
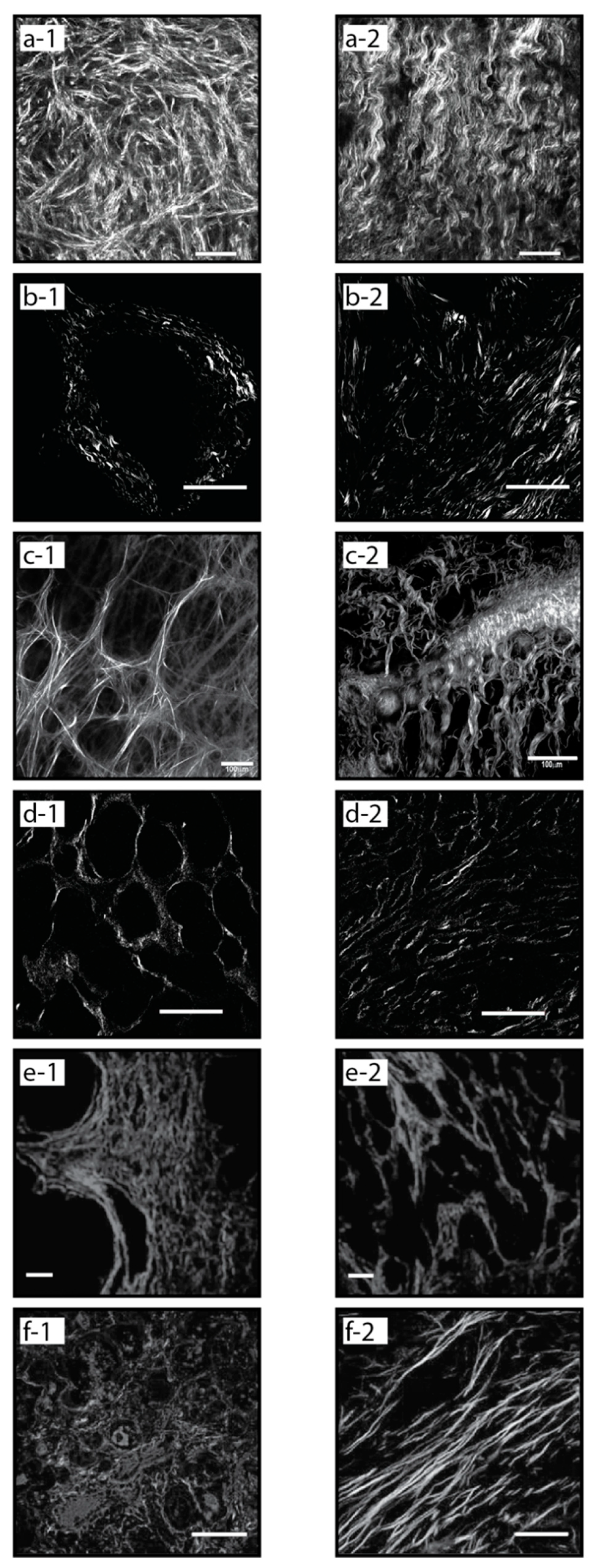
2. Collagen Fiber Imaging and Quantification
2.1. Histological Staining
2.2. Second Harmonic Generation Microscopy
2.3. Liquid Crystal-Based Polarizing Microscope (LC-PolScope)
2.4. Computational Methods
3. Collagen Fiber Organization as a Candidate Cancer Biomarker
3.1. Tissue Diagnosis and Grading
3.2. Patient Prognosis
4. Clinical Implications
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ricard-Blum, S. The Collagen Family. Cold Spring Harb. Perspect. Biol. 2011, 3, a004978. [Google Scholar] [CrossRef] [PubMed]
- Brinckmann, J. Collagens at a Glance. Top. Curr. Chem. 2005, 247, 1–6. [Google Scholar] [CrossRef]
- Gautieri, A.; Vesentini, S.; Redaelli, A.; Buehler, M.J. Hierarchical Structure and Nanomechanics of Collagen Microfibrils from the Atomistic Scale Up. Nano Lett. 2011, 11, 757–766. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharjee, A.; Bansal, M. Collagen Structure: The Madras Triple Helix and the Current Scenario. IUBMB Life 2005, 57, 161–172. [Google Scholar] [CrossRef] [PubMed]
- Makareeva, E.; Han, S.; Vera, J.C.; Sackett, D.L.; Holmbeck, K.; Phillips, C.L.; Visse, R.; Nagase, H.; Leikin, S. Carcinomas Contain a Matrix Metalloproteinase-Resistant Isoform of Type I Collagen Exerting Selective Support to Invasion. Cancer Res. 2010, 70, 4366–4374. [Google Scholar] [CrossRef]
- Mouw, J.K.; Ou, G.; Weaver, V.M. Extracellular Matrix Assembly: A Multiscale Deconstruction. Nat. Rev. Mol. Cell Biol. 2014, 15, 771–785. [Google Scholar] [CrossRef]
- LaComb, R.; Nadiarnykh, O.; Campagnola, P.J. Quantitative Second Harmonic Generation Imaging of the Diseased State Osteogenesis Imperfecta: Experiment and Simulation. Biophys. J. 2008, 94, 4504–4514. [Google Scholar] [CrossRef]
- Provenzano, P.P.; Eliceiri, K.W.; Campbell, J.M.; Inman, D.R.; White, J.G.; Keely, P.J. Collagen Reorganization at the Tumor-Stromal Interface Facilitates Local Invasion. BMC Med. 2006, 4. [Google Scholar] [CrossRef]
- Nadiarnykh, O.; LaComb, R.B.; Brewer, M.A.; Campagnola, P.J. Alterations of the Extracellular Matrix in Ovarian Cancer Studied by Second Harmonic Generation Imaging Microscopy. BMC Cancer 2010, 10. [Google Scholar] [CrossRef]
- Hanley, C.J.; Noble, F.; Ward, M.; Bullock, M.; Drifka, C.; Mellone, M.; Manousopoulou, A.; Johnston, H.E.; Hayden, A.; Thirdborough, S.; et al. A Subset of Myofibroblastic Cancer-Associated Fibroblasts Regulate Collagen Fiber Elongation, Which Is Prognostic in Multiple Cancers. Oncotarget 2015, 7, 6159–6174. [Google Scholar] [CrossRef]
- Provenzano, P.P.; Inman, D.R.; Eliceiri, K.W.; Trier, S.M.; Keely, P.J. Contact Guidance Mediated Three-Dimensional Cell Migration Is Regulated by Rho/ROCK-Dependent Matrix Reorganization. Biophys. J. 2008, 95, 5374–5384. [Google Scholar] [CrossRef] [PubMed]
- Riching, K.M.; Cox, B.L.; Salick, M.R.; Pehlke, C.; Riching, A.S.; Ponik, S.M.; Bass, B.R.; Crone, W.C.; Jiang, Y.; Weaver, A.M.; et al. 3D Collagen Alignment Limits Protrusions to Enhance Breast Cancer Cell Persistence. Biophys. J. 2014, 107, 2546–2558. [Google Scholar] [CrossRef] [PubMed]
- Salmon, H.; Franciszkiewicz, K.; Damotte, D.; Dieu-Nosjean, M.C.; Validire, P.; Trautmann, A.; Mami-Chouaib, F.; Donnadieu, E. Matrix Architecture Defines the Preferential Localization and Migration of T Cells into the Stroma of Human Lung Tumors. J. Clin. Investig. 2012, 122, 899–910. [Google Scholar] [CrossRef] [PubMed]
- Shi, Q.; Ghosh, R.P.; Engelke, H.; Rycroft, C.H.; Cassereau, L.; Sethian, J.A.; Weaver, V.M.; Liphardt, J.T. Rapid Disorganization of Mechanically Interacting Systems of Mammary Acini. Proc. Natl. Acad. Sci. USA 2014, 111, 658–663. [Google Scholar] [CrossRef]
- Wozniak, M.A.; Desai, R.; Solski, P.A.; Der, C.J.; Keely, P.J. ROCK-Generated Contractility Regulates Breast Epithelial Cell Differentiation in Response to the Physical Properties of a Three-Dimensional Collagen Matrix. J. Cell Biol. 2003, 163, 583–595. [Google Scholar] [CrossRef]
- Ma, X.; Schickel, M.E.; Stevenson, M.D.; Sarang-Sieminski, A.L.; Gooch, K.J.; Ghadiali, S.N.; Hart, R.T. Fibers in the Extracellular Matrix Enable Long-Range Stress Transmission between Cells. Biophys. J. 2013, 104, 1410–1418. [Google Scholar] [CrossRef]
- Levental, K.R.; Yu, H.; Kass, L.; Lakins, J.N.; Egeblad, M.; Erler, J.T.; Fong, S.F.T.; Csiszar, K.; Giaccia, A.; Weninger, W.; et al. Matrix Crosslinking Forces Tumor Progression by Enhancing Integrin Signaling. Cell 2009, 139, 891–906. [Google Scholar] [CrossRef]
- Gehler, S.; Baldassarre, M.; Lad, Y. Filamin A–Β1 Integrin Complex Tunes Epithelial Cell Response to Matrix Tension. Mol. Biol. Cell 2009, 20, 3224–3238. [Google Scholar] [CrossRef]
- Navab, R.; Strumpf, D.; To, C.; Pasko, E.; Kim, K.S.; Park, C.J.; Hai, J.; Liu, J.; Jonkman, J.; Barczyk, M.; et al. Integrin A11β1 Regulates Cancer Stromal Stiffness and Promotes Tumorigenicity and Metastasis in Non-Small Cell Lung Cancer. Oncogene 2015, 1–10. [Google Scholar] [CrossRef]
- Plodinec, M.; Loparic, M.; Monnier, C.A.; Obermann, E.C.; Zanetti-Dallenbach, R.; Oertle, P.; Hyotyla, J.T.; Aebi, U.; Bentires-Alj, M.; Lim, R.Y.H.; et al. The Nanomechanical Signature of Breast Cancer. Nat. Nanotechnol. 2012, 7, 757–765. [Google Scholar] [CrossRef]
- Tung, J.C.; Barnes, J.M.; Desai, S.R.; Sistrunk, C.; Conklin, M.W.; Schedin, P.; Eliceiri, K.W.; Keely, P.J.; Seewaldt, V.L.; Weaver, V.M. Tumor Mechanics and Metabolic Dysfunction. Free Radic. Biol. Med. 2015, 79, 269–280. [Google Scholar] [CrossRef] [PubMed]
- McConnell, J.C.; O’Connell, O.V.; Brennan, K.; Weiping, L.; Howe, M.; Joseph, L.; Knight, D.; O’Cualain, R.; Lim, Y.; Leek, A.; et al. Increased Peri-Ductal Collagen Micro-Organization May Contribute to Raised Mammographic Density. Breast Cancer Res. BCR 2016, 18, 5. [Google Scholar] [CrossRef] [PubMed]
- Stylianopoulos, T.; Martin, J.D.; Chauhan, V.P.; Jain, S.R.; Diop-Frimpong, B.; Bardeesy, N.; Smith, B.L.; Ferrone, C.R.; Hornicek, F.J.; Boucher, Y.; et al. Causes, Consequences, and Remedies for Growth-Induced Solid Stress in Murine and Human Tumors. Proc. Natl. Acad. Sci. USA 2012, 109, 15101–15108. [Google Scholar] [CrossRef]
- Jacobetz, M.A.; Chan, D.S.; Neesse, A.; Bapiro, T.E.; Cook, N.; Frese, K.K.; Feig, C.; Nakagawa, T.; Caldwell, M.E.; Zecchini, H.I.; et al. Hyaluronan Impairs Vascular Function and Drug Delivery in a Mouse Model of Pancreatic Cancer. Gut 2012. [Google Scholar] [CrossRef] [PubMed]
- Gaggioli, C.; Hooper, S.; Hidalgo-Carcedo, C.; Grosse, R.; Marshall, J.F.; Harrington, K.; Sahai, E. Fibroblast-Led Collective Invasion of Carcinoma Cells with Differing Roles for RhoGTPases in Leading and Following Cells. Nat. Cell Biol. 2007, 9, 1392–1400. [Google Scholar] [CrossRef]
- Yang, N.; Mosher, R.; Seo, S.; Beebe, D.; Friedl, A. Syndecan-1 in Breast Cancer Stroma Fibroblasts Regulates Extracellular Matrix Fiber Organization and Carcinoma Cell Motility. Am. J. Pathol. 2011, 178, 325–335. [Google Scholar] [CrossRef]
- Morris, R.A.; Damon, B.; Mironov, V.; Kasyanov, V.; Ramamurthi, A.; Moreno-Rodriguez, R.; Trusk, T.; Potts, J.D.; Goodwin, R.L.; Davis, J.; et al. Periostin Regulates Collagen Fibrillogenesis and the Biomechanical Properties of Connective Tissues. J. Cell. Biochem. 2007, 101, 695–711. [Google Scholar] [CrossRef]
- Chen, J.; Chen, Z.; Chen, M.; Li, D.; Li, Z.; Xiong, Y.; Dong, J.; Li, X. Role of Fibrillar Tenascin-C in Metastatic Pancreatic Cancer. Int. J. Oncol. 2009, 34, 1029–1036. [Google Scholar] [CrossRef]
- Velling, T.; Risteli, J.; Wennerberg, K.; Mosher, D.F.; Johansson, S. Polymerization of Type I and III Collagens Is Dependent on Fibronectin and Enhanced by Integrins alpha 11beta 1 and alpha 2beta 1. J. Biol. Chem. 2002, 277, 37377–37381. [Google Scholar] [CrossRef]
- McDonald, J.A.; Kelley, D.G.; Broekelmann, T.J. Role of Fibronectin in Collagen Deposition: Fab’ to the Gelatin-Binding Domain of Fibronectin Inhibits Both Fibronectin and Collagen Organization in Fibroblast Extracellular Matrix. J. Cell Biol. 1982, 92, 485–492. [Google Scholar] [CrossRef]
- Sottile, J.; Shi, F.; Rublyevska, I.; Chiang, H.-Y.; Lust, J.; Chandler, J. Fibronectin-Dependent Collagen I Deposition Modulates the Cell Response to Fibronectin. Am. J. Physiol. Cell Physiol. 2007, 293, 1934–1946. [Google Scholar] [CrossRef] [PubMed]
- Kadler, K.E.; Hill, A.; Canty-Laird, E.G. Collagen Fibrillogenesis: Fibronectin, Integrins, and Minor Collagens as Organizers and Nucleators. Curr. Opin. Cell Biol. 2008, 20, 495–501. [Google Scholar] [CrossRef] [PubMed]
- Junqueira, L.C.U.; Bignolas, G.; Brentani, R.R. Picrosirius Staining plus Polarization Microscopy, a Specific Method for Collagen Detection in Tissue Sections. Histochem. J. 1979, 11, 447–455. [Google Scholar] [CrossRef]
- Drifka, C.R.; Loeffler, A.G.; Mathewson, K.; Mehta, G.; Keikhosravi, A.; Liu, Y.; Lemancik, S.; Ricke, W.A.; Weber, S.M.; Kao, W.J.; et al. Comparison of Picrosirius Red Staining with Second Harmonic Generation Imaging for the Quantification of Clinically Relevant Collagen Fiber Features in Histopathology Samples. J. Histochem. Cytochem. 2016, 64, 519–529. [Google Scholar] [CrossRef] [PubMed]
- Lattouf, R.; Younes, R.; Lutomski, D.; Naaman, N.; Godeau, G.; Senni, K.; Changotade, S. Picrosirius Red Staining: A Useful Tool to Appraise Collagen Networks in Normal and Pathological Tissues. J. Histochem. Cytochem. 2014, 62, 751–758. [Google Scholar] [CrossRef]
- Hwang, J.; Huang, Y.; Burwell, T.J.; Peterson, N.C.; Connor, J.; Weiss, S.J.; Yu, S.M.; Li, Y. In Situ Imaging of Tissue Remodeling with Collagen Hybridizing Peptides. ACS Nano 2017, 11, 9825–9835. [Google Scholar] [CrossRef]
- Li, Y.; Yu, S.M. In Situ Detection of Degraded and Denatured Collagen via Triple Helical Hybridization: New Tool in Histopathology. In Collagen: Methods and Protocols; Sagi, I., Afratis, N.A., Eds.; Springer: New York, NY, USA, 2019; pp. 135–144. ISBN 978-1-4939-9095-5. [Google Scholar]
- Brisson, B.K.; Mauldin, E.A.; Lei, W.; Vogel, L.K.; Power, A.M.; Lo, A.; Dopkin, D.; Khanna, C.; Wells, R.G.; Puré, E.; et al. Type III Collagen Directs Stromal Organization and Limits Metastasis in a Murine Model of Breast Cancer. Am. J. Pathol. 2015, 185, 1471–1486. [Google Scholar] [CrossRef]
- Keikhosravi, A.; Bredfeldt, J.S.; Sagar, A.K.; Eliceiri, K.W. Chapter 28—Second-harmonic generation imaging of cancer. In Methods in Cell Biology; Waters, J.C., Wittman, T., Eds.; Quantitative Imaging in Cell Biology; Academic Press: Cambridge, MA, USA, 2014; Volume 123, pp. 531–546. [Google Scholar]
- Provenzano, P.P.; Eliceiri, K.W.; Keely, P.J. Shining New Light on 3D Cell Motility and the Metastatic Process. Trends Cell Biol. 2009, 19, 638–648. [Google Scholar] [CrossRef]
- Provenzano, P.P.; Eliceiri, K.W.; Keely, P.J. Multiphoton Microscopy and Fluorescence Lifetime Imaging Microscopy (FLIM) to Monitor Metastasis and the Tumor Microenvironment. Clin. Exp. Metastasis 2009, 26. [Google Scholar] [CrossRef]
- Campagnola, P.J. Second Harmonic Generation Imaging Microscopy: Applications to Diseases Diagnostics. Anal. Chem. 2011, 83, 3224–3231. [Google Scholar] [CrossRef]
- Campagnola, P.J.; Loew, L.M. Second-Harmonic Imaging Microscopy for Visualizing Biomolecular Arrays in Cells, Tissues and Organisms. Nat. Biotechnol. 2003, 21, 1356–1360. [Google Scholar] [CrossRef] [PubMed]
- Condeelis, J.; Segall, J.E. Intravital Imaging of Cell Movement in Tumours. Nat. Rev. Cancer 2003, 3, 921–930. [Google Scholar] [CrossRef] [PubMed]
- Drifka, C.R.; Tod, J.; Loeffler, A.G.; Liu, Y.; Thomas, G.J.; Eliceiri, K.W.; Kao, W.J. Periductal Stromal Collagen Topology of Pancreatic Ductal Adenocarcinoma Differs from That of Normal and Chronic Pancreatitis. Mod. Pathol. 2015, 28, 1470–1480. [Google Scholar] [CrossRef] [PubMed]
- Conklin, M.W.; Eickhoff, J.C.; Riching, K.M.; Pehlke, C.A.; Eliceiri, K.W.; Provenzano, P.P.; Friedl, A.; Keely, P.J. Aligned Collagen Is a Prognostic Signature for Survival in Human Breast Carcinoma. Am. J. Pathol. 2011, 178. [Google Scholar] [CrossRef]
- Drifka, C.R.; Eliceiri, K.W.; Weber, S.M.; Kao, W.J. A Bioengineered Heterotypic Stroma-Cancer Microenvironment Model to Study Pancreatic Ductal Adenocarcinoma. Lab. Chip 2013, 13, 3965–3975. [Google Scholar] [CrossRef]
- Ajeti, V.; Nadiarnykh, O.; Ponik, S.M.; Keely, P.J.; Eliceiri, K.W.; Campagnola, P.J. Structural Changes in Mixed Col I/Col V Collagen Gels Probed by SHG Microscopy: Implications for Probing Stromal Alterations in Human Breast Cancer. Biomed. Opt. Express 2011, 2, 2307–2316. [Google Scholar] [CrossRef]
- Sung, K.E.; Su, G.; Pehlke, C.; Trier, S.M.; Eliceiri, K.W.; Keely, P.J.; Friedl, A.; Beebe, D.J. Control of 3-Dimensional Collagen Matrix Polymerization for Reproducible Human Mammary Fibroblast Cell Culture in Microfluidic Devices. Biomaterials 2009, 30. [Google Scholar] [CrossRef]
- Sung, K.E.; Yang, N.; Pehlke, C.; Keely, P.J.; Eliceiri, K.W.; Friedl, A.; Beebe, D.J. Transition to Invasion in Breast Cancer: A Microfluidic in Vitro Model Enables Examination of Spatial and Temporal Effects. Integr. Biol. 2011, 3. [Google Scholar] [CrossRef]
- Raub, C.B.; Suresh, V.; Krasieva, T.; Lyubovitsky, J.; Mih, J.D.; Putnam, A.J.; Tromberg, B.J.; George, S.C. Noninvasive Assessment of Collagen Gel Microstructure and Mechanics Using Multiphoton Microscopy. Biophys. J. 2007, 92, 2212–2222. [Google Scholar] [CrossRef]
- Brown, E.; McKee, T.; di Tomaso, E.; Pluen, A.; Seed, B.; Boucher, Y.; Jain, R.K. Dynamic Imaging of Collagen and Its Modulation in Tumors in Vivo Using Second-Harmonic Generation. Nat. Med. 2003, 9. [Google Scholar] [CrossRef]
- Walsh, A.J.; Cook, R.S.; Lee, J.H.; Arteaga, C.L.; Skala, M.C. Collagen Density and Alignment in Responsive and Resistant Trastuzumab-Treated Breast Cancer Xenografts. J. Biomed. Opt. 2015, 20, 026004. [Google Scholar] [CrossRef] [PubMed]
- Oldenbourg, R.; Mei, G. New Polarized Light Microscope with Precision Universal Compensator. J. Microsc. 1995, 180, 140–147. [Google Scholar] [CrossRef] [PubMed]
- Oldenbourg, R. Polarization Microscopy with the LC-PolScope. In Live Cell Imaging: A Laboratory Manual; Goldman, R.D., Spector, D.L., Eds.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2005; pp. 205–237. [Google Scholar]
- Keikhosravi, A.; Liu, Y.; Drifka, C.; Woo, K.M.; Verma, A.; Oldenbourg, R.; Eliceiri, K.W. Quantification of Collagen Organization in Histopathology Samples Using Liquid Crystal Based Polarization Microscopy. Biomed. Opt. Express 2017, 8, 4243–4256. [Google Scholar] [CrossRef] [PubMed]
- Provenzano, P.P.; Inman, D.R.; Eliceiri, K.W.; Knittel, J.G.; Yan, L.; Rueden, C.T.; White, J.G.; Keely, P.J. Collagen Density Promotes Mammary Tumor Initiation and Progression. BMC Med. 2008, 6, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Shea, M.P.; O’Leary, K.A.; Wegner, K.A.; Vezina, C.M.; Schuler, L.A. High Collagen Density Augments MTOR-Dependent Cancer Stem Cells in ERα+ Mammary Carcinomas, and Increases MTOR-Independent Lung Metastases. Cancer Lett. 2018, 433, 1–9. [Google Scholar] [CrossRef]
- Drifka, C.R.; Loeffler, A.G.; Mathewson, K.; Keikhosravi, A.; Eickhoff, J.C.; Liu, Y.; Weber, S.M.; Kao, W.J.; Eliceiri, K.W. Highly Aligned Stromal Collagen Is a Negative Prognostic Factor Following Pancreatic Ductal Adenocarcinoma Resection. Oncotarget 2016. [Google Scholar] [CrossRef]
- Chen, G.; Liu, Y.; Zhu, X.; Huang, Z.; Cai, J.; Chen, R.; Xiong, S.; Zeng, H. Phase and Texture Characterizations of Scar Collagen Second-Harmonic Generation Images Varied with Scar Duration. Microsc. Microanal. 2015, 21, 855–862. [Google Scholar] [CrossRef]
- Hu, W.; Li, H.; Fu, L.; Wang, C.; Gou, S. Characterization of Collagen Fibers by Means of Texture Analysis of Second Harmonic Generation Images Using Orientation-Dependent Gray Level Co-Occurrence Matrix Method. J. Biomed. Opt. 2012, 17, 026007. [Google Scholar] [CrossRef]
- Wen, B.L.; Brewer, M.A.; Nadiarnykh, O.; Hocker, J.D.; Singh, V.; Mackie, T.R.; Campagnola, P.J. Texture Analysis Applied to Second Harmonic Generation Image Data for Ovarian Cancer Classification. J. Biomed. Opt. 2014, 19, 096007. [Google Scholar] [CrossRef]
- Eekhoff, J.D.; Lake, S.P. Three-Dimensional Computation of Fibre Orientation, Diameter and Branching in Segmented Image Stacks of Fibrous Networks. J. R. Soc. Interface 2020, 17, 20200371. [Google Scholar] [CrossRef]
- Brett, E.A.; Sauter, M.A.; Machens, H.-G.; Duscher, D. Tumor-Associated Collagen Signatures: Pushing Tumor Boundaries. Cancer Metab. 2020, 8, 14. [Google Scholar] [CrossRef] [PubMed]
- Quinn, K.P.; Georgakoudi, I. Rapid Quantification of Pixel-Wise Fiber Orientation Data in Micrographs. J. Biomed. Opt. 2013, 18, 046003. [Google Scholar] [CrossRef] [PubMed]
- Rezakhaniha, R.; Agianniotis, A.; Schrauwen, J.T.C.; Griffa, A.; Sage, D.; Bouten, C.V.C.; Van De Vosse, F.; Unser, M.; Stergiopulos, N. Experimental Investigation of Collagen Waviness and Orientation in the Arterial Adventitia Using Confocal Laser Scanning Microscopy. Biomech. Model. Mechanobiol. 2012, 11, 461–473. [Google Scholar] [CrossRef] [PubMed]
- Chaudhuri, S.; Nguyen, H.; Rangayyan, R.M.; Walsh, S.; Frank, C.B. A Fourier Domain Directional Filterng Method for Analysis of Collagen Alignment in Ligaments. IEEE Trans. Biomed. Eng. 1987, BME-34, 509–518. [Google Scholar] [CrossRef] [PubMed]
- Kartasalo, K.; Pölönen, R.-P.; Ojala, M.; Rasku, J.; Lekkala, J.; Aalto-Setälä, K.; Kallio, P. CytoSpectre: A Tool for Spectral Analysis of Oriented Structures on Cellular and Subcellular Levels. BMC Bioinform. 2015, 16, 344. [Google Scholar] [CrossRef] [PubMed]
- Pourdeyhimi, B.; Kim, H. Measuring Fiber Orientation in Nonwovens: The Hough Transform. Text. Res. J. 2002, 72, 803–809. [Google Scholar] [CrossRef]
- Pourdeyhimi, B.; Dent, R.; Davis, H. Measuring Fiber Orientation in Nonwovens Part III: Fourier Transform. Text. Res. J. 1997, 67, 143–151. [Google Scholar] [CrossRef]
- Püspöki, Z.; Storath, M.; Sage, D.; Unser, M. Transforms and Operators for Directional Bioimage Analysis: A Survey. In Focus on Bio-Image Informatics; De Vos, W.H., Munck, S., Timmermans, J.-P., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 69–93. ISBN 978-3-319-28549-8. [Google Scholar]
- Xu, B.; Yu, L. Determining Fiber Orientation Distribution in Nonwovens with Hough Transform Techniques. Text. Res. J. 1997, 67, 563–571. [Google Scholar] [CrossRef]
- Wu, J.; Rajwa, B.; Filmer, D.L.; Hoffmann, C.M.; Yuan, B.; Chiang, C.-S.; Sturgis, J.; Robinson, J.P. Analysis of Orientations of Collagen Fibers by Novel Fiber-Tracking Software. Microsc. Microanal. 2003, 9, 574. [Google Scholar] [CrossRef]
- Stein, A.M.; Vader, D.A.; Jawerth, L.M.; Weitz, D.A.; Sander, L.M. An Algorithm for Extracting the Network Geometry of Three-Dimensional Collagen Gels. J. Microsc. 2008, 232, 463–475. [Google Scholar] [CrossRef]
- Bredfeldt, J.S.; Liu, Y.; Pehlke, C.A.; Conklin, M.W.; Szulczewski, J.M.; Inman, D.R.; Keely, P.J.; Nowak, R.D.; Mackie, T.R.; Eliceiri, K.W. Computational Segmentation of Collagen Fibers from Second-Harmonic Generation Images of Breast Cancer. J. Biomed. Opt. 2014, 19, 16007. [Google Scholar] [CrossRef] [PubMed]
- Candès, E.; Demanet, L.; Donoho, D.; Ying, L. Fast Discrete Curvelet Transforms. Multiscale Model. Simul. 2006, 5, 861–899. [Google Scholar] [CrossRef]
- Liu, Y.; Keikhosravi, A.; Pehlke, C.A.; Bredfeldt, J.S.; Dutson, M.; Liu, H.; Mehta, G.S.; Claus, R.; Patel, A.J.; Conklin, M.W.; et al. Fibrillar Collagen Quantification With Curvelet Transform Based Computational Methods. Front. Bioeng. Biotechnol. 2020, 8. [Google Scholar] [CrossRef]
- Bredfeldt, J.S. Collagen Alignment Imaging and Analysis for Breast Cancer Classification. Ph.D. Thesis, The University of Wisconsin, Madison, WI, USA, 2014. [Google Scholar]
- Reis, L.A.; Garcia, A.P.; Gomes, E.F.; Longford, F.G.; Frey, J.G.; Cassali, G.D.; de Paula, A.M. Canine Mammary Cancer Diagnosis from Quantitative Properties of Nonlinear Optical Images. Biomed. Opt. Express 2020, 11, 6413–6427. [Google Scholar] [CrossRef]
- Huttunen, M.J.; Hassan, A.; McCloskey, C.W.; Fasih, S.; Upham, J.; Vanderhyden, B.C.; Boyd, R.W.; Murugkar, S. Automated Classification of Multiphoton Microscopy Images of Ovarian Tissue Using Deep Learning. J. Biomed. Opt. 2018, 23, 066002. [Google Scholar] [CrossRef]
- Wen, B.; Campbell, K.R.; Tilbury, K.; Nadiarnykh, O.; Brewer, M.A.; Patankar, M.; Singh, V.; Eliceiri, K.W.; Campagnola, P.J. 3D Texture Analysis for Classification of Second Harmonic Generation Images of Human Ovarian Cancer. Sci. Rep. 2016, 6, 35734. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Xu, Z.; Liao, C.; Cai, J.; Huang, Y.; Chen, H.; Tao, X.; Huang, Z.; Chen, J.; Dong, J.; et al. Epithelium Segmentation and Automated Gleason Grading of Prostate Cancer via Deep Learning in Label-free Multiphoton Microscopic Images. J. Biophotonics 2020, 13. [Google Scholar] [CrossRef]
- Lindeberg, T. Edge Detection and Ridge Detection with Automatic Scale Selection. Int. J. Comput. Vis. 1998, 30, 117–156. [Google Scholar] [CrossRef]
- Boudaoud, A.; Burian, A.; Borowska-Wykręt, D.; Uyttewaal, M.; Wrzalik, R.; Kwiatkowska, D.; Hamant, O. FibrilTool, an ImageJ Plug-in to Quantify Fibrillar Structures in Raw Microscopy Images. Nat. Protoc. 2014, 9, 457–463. [Google Scholar] [CrossRef]
- Wershof, E.; Barry, D.J.; Jenkins, R.P.; Rullan, A.; Wilkins, A.; Roxanis, I.; Anderson, K.I.; Park, D.; Bates, P.A.; Sahai, E. A FIJI Macro for Quantifying Pattern in Extracellular Matrix. bioRxiv 2019, 867507. [Google Scholar] [CrossRef]
- Bredfeldt, J.S.; Liu, Y.; Conklin, M.W.; Keely, P.J.; Mackie, T.R.; Eliceiri, K.W. Automated Quantification of Aligned Collagen for Human Breast Carcinoma Prognosis. J. Pathol. Inform. 2014, 5. [Google Scholar] [CrossRef]
- Liu, Y.; Keikhosravi, A.; Mehta, G.S.; Drifka, C.R.; Eliceiri, K.W. Methods for Quantifying Fibrillar Collagen Alignment. In Fibrosis: Methods and Protocols; Methods in Molecular Biology; Rittié, L., Ed.; Springer: New York, NY, USA, 2017; pp. 429–451. ISBN 978-1-4939-7113-8. [Google Scholar]
- Zhou, Z.-H.; Ji, C.-D.; Xiao, H.-L.; Zhao, H.-B.; Cui, Y.-H.; Bian, X.-W. Reorganized Collagen in the Tumor Microenvironment of Gastric Cancer and Its Association with Prognosis. J. Cancer 2017, 8, 1466–1476. [Google Scholar] [CrossRef] [PubMed]
- Despotović, S.Z.; Milićević, Đ.N.; Krmpot, A.J.; Pavlović, A.M.; Živanović, V.D.; Krivokapić, Z.; Pavlović, V.B.; Lević, S.; Nikolić, G.; Rabasović, M.D. Altered Organization of Collagen Fibers in the Uninvolved Human Colon Mucosa 10 Cm and 20 Cm Away from the Malignant Tumor. Sci. Rep. 2020, 10, 6359. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Lin, L.; Wang, G.; Zuo, N.; Zhan, Z.; Xie, S.; Chen, G.; Chen, J.; Zhuo, S. Label-Free Classification of Hepatocellular-Carcinoma Grading Using Second Harmonic Generation Microscopy. Biomed. Opt. Express 2018, 9, 3783–3793. [Google Scholar] [CrossRef]
- Arun Gopinathan, P.; Kokila, G.; Jyothi, M.; Ananjan, C.; Pradeep, L.; Humaira Nazir, S. Study of Collagen Birefringence in Different Grades of Oral Squamous Cell Carcinoma Using Picrosirius Red and Polarized Light Microscopy. Scientifica 2015, 2015, 802980. [Google Scholar] [CrossRef]
- Gole, L.; Yeong, J.; Lim, J.C.T.; Ong, K.H.; Han, H.; Thike, A.A.; Poh, Y.C.; Yee, S.; Iqbal, J.; Hong, W.; et al. Quantitative Stain-Free Imaging and Digital Profiling of Collagen Structure Reveal Diverse Survival of Triple Negative Breast Cancer Patients. Breast Cancer Res. 2020, 22, 42. [Google Scholar] [CrossRef]
- Garcia, A.M.; Magalhes, F.L.; Soares, J.S.; Junior, E.P.; Lima, M.F.R.d.; Mamede, M.; Paula, A.M.d. Second Harmonic Generation Imaging of the Collagen Architecture in Prostate Cancer Tissue. Biomed. Phys. Eng. Express 2018, 4, 025026. [Google Scholar] [CrossRef]
- Best, S.L.; Liu, Y.; Keikhosravi, A.; Drifka, C.R.; Woo, K.M.; Mehta, G.S.; Altwegg, M.; Thimm, T.N.; Houlihan, M.; Bredfeldt, J.S.; et al. Collagen Organization of Renal Cell Carcinoma Differs between Low and High Grade Tumors. BMC Cancer 2019, 19, 490. [Google Scholar] [CrossRef]
- Pointer, K.B.; Clark, P.A.; Schroeder, A.B.; Salamat, M.S.; Eliceiri, K.W.; Kuo, J.S. Association of Collagen Architecture with Glioblastoma Patient Survival. J. Neurosurg. 2017, 126, 1812–1821. [Google Scholar] [CrossRef]
- Birk, J.W.; Tadros, M.; Moezardalan, K.; Nadyarnykh, O.; Forouhar, F.; Anderson, J.; Campagnola, P. Second Harmonic Generation Imaging Distinguishes Both High-Grade Dysplasia and Cancer from Normal Colonic Mucosa. Dig. Dis. Sci. 2014, 59, 1529–1534. [Google Scholar] [CrossRef]
- Fanous, M.; Keikhosravi, A.; Kajdacsy-Balla, A.; Eliceiri, K.W.; Popescu, G. Quantitative Phase Imaging of Stromal Prognostic Markers in Pancreatic Ductal Adenocarcinoma. Biomed. Opt. Express 2020, 11, 1354–1364. [Google Scholar] [CrossRef] [PubMed]
- Laklai, H.; Miroshnikova, Y.A.; Pickup, M.W.; Collisson, E.A.; Kim, G.E.; Barrett, A.S.; Hill, R.C.; Lakins, J.N.; Schlaepfer, D.D.; Mouw, J.K.; et al. Genotype Tunes Pancreatic Ductal Adenocarcinoma Tissue Tension to Induce Matricellular Fibrosis and Tumor Progression. Nat. Med. 2016, 22, 497–505. [Google Scholar] [CrossRef] [PubMed]
- Esbona, K.; Yi, Y.; Saha, S.; Yu, M.; Van Doorn, R.R.; Conklin, M.W.; Graham, D.S.; Wisinski, K.B.; Ponik, S.M.; Eliceiri, K.W.; et al. The Presence of Cyclooxygenase 2, Tumor-Associated Macrophages, and Collagen Alignment as Prognostic Markers for Invasive Breast Carcinoma Patients. Am. J. Pathol. 2018, 188, 559–573. [Google Scholar] [CrossRef] [PubMed]
- Falzon, G.; Pearson, S.; Murison, R. Analysis of Collagen Fibre Shape Changes in Breast Cancer. Phys. Med. Biol. 2008, 53, 6641–6652. [Google Scholar] [CrossRef] [PubMed]
- Ambekar, R.; Lau, T.-Y.; Walsh, M.; Bhargava, R.; Toussaint, K.C.J. Quantifying Collagen Structure in Breast Biopsies Using Second-Harmonic Generation Imaging. Biomed. Opt. Express 2012, 3, 2021–2035. [Google Scholar] [CrossRef] [PubMed]
- Lukina, M.M.; Dudenkova, V.V.; Shimolina, L.E.; Snopova, L.B.; Zagaynova, E.V.; Shirmanova, M.V. In Vivo Metabolic and SHG Imaging for Monitoring of Tumor Response to Chemotherapy. Cytom. Part A 2019, 95, 47–55. [Google Scholar] [CrossRef]
- Li, L.; Kang, D.; Huang, Z.; Zhan, Z.; Feng, C.; Zhou, Y.; Tu, H.; Zhuo, S.; Chen, J. Multimodal Multiphoton Imaging for Label-Free Monitoring of Early Gastric Cancer. BMC Cancer 2019, 19, 295. [Google Scholar] [CrossRef]
- Golaraei, A.; Cisek, R.; Krouglov, S.; Navab, R.; Niu, C.; Sakashita, S.; Yasufuku, K.; Tsao, M.-S.; Wilson, B.C.; Barzda, V. Characterization of Collagen in Non-Small Cell Lung Carcinoma with Second Harmonic Polarization Microscopy. Biomed. Opt. Express 2014, 5, 3562–3567. [Google Scholar] [CrossRef]
- Manjunatha, B.S.; Agrawal, A.; Shah, V. Histopathological Evaluation of Collagen Fibers Using Picrosirius Red Stain and Polarizing Microscopy in Oral Squamous Cell Carcinoma. J. Cancer Res. Ther. 2015, 11, 272–276. [Google Scholar] [CrossRef]
- Hompland, T.; Erikson, A.; Lindgren, M.; Lindmo, T.; de Lange Davies, C. Second-Harmonic Generation in Collagen as a Potential Cancer Diagnostic Parameter. J. Biomed. Opt. 2008, 13, 054050. [Google Scholar] [CrossRef]
- Kirkpatrick, N.D.; Brewer, M.A.; Utzinger, U. Endogenous Optical Biomarkers of Ovarian Cancer Evaluated with Multiphoton Microscopy. Cancer Epidemiol. Biomark. Prev. 2007, 16, 2048–2057. [Google Scholar] [CrossRef] [PubMed]
- Adur, J.; Pelegati, V.B.; de Thomaz, A.A.; Baratti, M.O.; Andrade, L.A.L.A.; Carvalho, H.F.; Bottcher-Luiz, F.; Cesar, C.L. Second Harmonic Generation Microscopy as a Powerful Diagnostic Imaging Modality for Human Ovarian Cancer. J. Biophotonics 2014, 7, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Tilbury, K.B.; Campbell, K.R.; Eliceiri, K.W.; Salih, S.M.; Patankar, M.; Campagnola, P.J. Stromal Alterations in Ovarian Cancers via Wavelength Dependent Second Harmonic Generation Microscopy and Optical Scattering. BMC Cancer 2017, 17, 102. [Google Scholar] [CrossRef] [PubMed]
- Vennin, C.; Chin, V.T.; Warren, S.C.; Lucas, M.C.; Herrmann, D.; Magenau, A.; Melenec, P.; Walters, S.N.; Monte-Nieto, G.d.; Conway, J.R.W.; et al. Transient Tissue Priming via ROCK Inhibition Uncouples Pancreatic Cancer Progression, Sensitivity to Chemotherapy, and Metastasis. Sci. Transl. Med. 2017, 9. [Google Scholar] [CrossRef] [PubMed]
- Ling, Y.; Li, C.; Feng, K.; Palmer, S.; Appleton, P.L.; Lang, S.; McGloin, D.; Huang, Z.; Nabi, G. Second Harmonic Generation (SHG) Imaging of Cancer Heterogeneity in Ultrasound Guided Biopsies of Prostate in Men Suspected with Prostate Cancer. J. Biophotonics 2017, 10, 911–918. [Google Scholar] [CrossRef]
- Galli, R.; Sablinskas, V.; Dasevicius, D.; Laurinavicius, A.; Jankevicius, F.; Koch, E.; Steiner, G. Non-Linear Optical Microscopy of Kidney Tumours. J. Biophotonics 2014, 7, 23–27. [Google Scholar] [CrossRef]
- Burchardt, M.; Engers, R.; Müller, M.; Burchardt, T.; Willers, R.; Epstein, J.I.; Ackermann, R.; Gabbert, H.; De La Taille, A.; Rubin, M. Interobserver Reproducibility of Gleason Grading: Evaluation Using Prostate Cancer Tissue Microarrays. J. Cancer Res. Clin. Oncol. 2008, 134, 1071–1078. [Google Scholar] [CrossRef]
- Ogawa, K.; Oguchi, M.; Nakashima, Y.; Yamabe, H. Distribution of Collagen Type IV in Brain Tumors: An Immunohistochemical Study. J. Neurooncol. 1989, 7, 357–366. [Google Scholar] [CrossRef]
- Esquibel, C.R.; Wendt, K.D.; Lee, H.C.; Gaire, J.; Shoffstall, A.; Urdaneta, M.E.; Chacko, J.V.; Brodnick, S.K.; Otto, K.J.; Capadona, J.R.; et al. Second Harmonic Generation Imaging of Collagen in Chronically Implantable Electrodes in Brain Tissue. Front. Neurosci. 2020, 14, 95. [Google Scholar] [CrossRef]
- Han, W.; Chen, S.; Yuan, W.; Fan, Q.; Tian, J.; Wang, X.; Chen, L.; Zhang, X.; Wei, W.; Liu, R.; et al. Oriented Collagen Fibers Direct Tumor Cell Intravasation. Proc. Natl. Acad. Sci. USA 2016, 113, 11208–11213. [Google Scholar] [CrossRef]
- Suzuki, K.; Gerelchuluun, A.; Hong, Z.; Sun, L.; Zenkoh, J.; Moritake, T.; Tsuboi, K. Celecoxib Enhances Radiosensitivity of Hypoxic Glioblastoma Cells through Endoplasmic Reticulum Stress. Neuro-Oncology 2013, 15, 1186–1199. [Google Scholar] [CrossRef] [PubMed]
- Tomko, L.A.; Hill, R.C.; Barrett, A.; Szulczewski, J.M.; Conklin, M.W.; Eliceiri, K.W.; Keely, P.J.; Hansen, K.C.; Ponik, S.M. Targeted Matrisome Analysis Identifies Thrombospondin-2 and Tenascin-C in Aligned Collagen Stroma from Invasive Breast Carcinoma. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Sawyer, A.J.; Kyriakides, T.R. Matricellular Proteins in Drug Delivery: Therapeutic Targets, Active Agents, and Therapeutic Localization. Adv. Drug Deliv. Rev. 2016, 97, 56–68. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Torphy, R.J.; Steiger, K.; Hongo, H.; Ritchie, A.J.; Kriegsmann, M.; Horst, D.; Umetsu, S.E.; Joseph, N.M.; McGregor, K.; et al. Pancreatic Ductal Adenocarcinoma Progression Is Restrained by Stromal Matrix. J. Clin. Investig. 2020, 130, 4704–4709. [Google Scholar] [CrossRef]
- Lee, J.J.; Perera, R.M.; Wang, H.; Wu, D.-C.; Liu, X.S.; Han, S.; Fitamant, J.; Jones, P.D.; Ghanta, K.S.; Kawano, S.; et al. Stromal Response to Hedgehog Signaling Restrains Pancreatic Cancer Progression. Proc. Natl. Acad. Sci. USA 2014, 111, E3091–E3100. [Google Scholar] [CrossRef]
- Moore, M.J.; Hamm, J.; Dancey, J.; Eisenberg, P.; Dagenais, M.; Fields, A.; Hagan, K.; Greenberg, B.; Colwell, B.; Zee, B.; et al. Comparison of Gemcitabine versus the Matrix Metalloproteinase Inhibitor BAY 12-9566 in Patients with Advanced or Metastatic Adenocarcinoma of the Pancreas: A Phase III Trial of the National Cancer Institute of Canada Clinical Trials Group. J. Clin. Oncol. 2003, 21, 3296–3302. [Google Scholar] [CrossRef]
- Nguyen, E.V.; Pereira, B.A.; Lawrence, M.G.; Ma, X.; Rebello, R.J.; Chan, H.; Niranjan, B.; Wu, Y.; Ellem, S.; Guan, X.; et al. Proteomic Profiling of Human Prostate Cancer-Associated Fibroblasts (CAF) Reveals LOXL2-Dependent Regulation of the Tumor Microenvironment. Mol. Cell. Proteomics 2019, 18, 1410–1427. [Google Scholar] [CrossRef]
- Kocher, H.M.; Basu, B.; Froeling, F.E.; Sarker, D.; Slater, S.; Carlin, D.; de Souza, N.M.; De Paepe, K.N.; Goulart, M.R.; Hughes, C.; et al. Phase I Clinical Trial Repurposing All-Trans Retinoic Acid as a Stromal Targeting Agent for Pancreatic Cancer. Nat. Commun. 2020, 11, 1–9. [Google Scholar] [CrossRef]
- Sheridan, C. Pancreatic Cancer Provides Testbed for First Mechanotherapeutics. Nat. Biotechnol. 2019, 37, 829–831. [Google Scholar] [CrossRef]
- Jiang, H.; Hegde, S.; Knolhoff, B.L.; Zhu, Y.; Herndon, J.M.; Meyer, M.A.; Nywening, T.M.; Hawkins, W.G.; Shapiro, I.M.; Weaver, D.T.; et al. Targeting Focal Adhesion Kinase Renders Pancreatic Cancers Responsive to Checkpoint Immunotherapy. Nat. Med. 2016, 22, 851–860. [Google Scholar] [CrossRef]
- Lu, P.; Weaver, V.M.; Werb, Z. The Extracellular Matrix: A Dynamic Niche in Cancer Progression. J. Cell Biol. 2012, 196, 395–406. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Xu, H.; Wang, W.; Li, S.; Li, H.; Li, T.; Zhang, W.; Yu, X.; Liu, L. The Role of Collagen in Cancer: From Bench to Bedside. J. Transl. Med. 2019, 17, 309. [Google Scholar] [CrossRef] [PubMed]
- Park, D.; Wershof, E.; Boeing, S.; Labernadie, A.; Jenkins, R.P.; George, S.; Trepat, X.; Bates, P.A.; Sahai, E. Extracellular Matrix Anisotropy Is Determined by TFAP2C-Dependent Regulation of Cell Collisions. Nat. Mater. 2020, 19, 227–238. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Disease | Metric | Visualization | Findings | References |
|---|---|---|---|---|
| Breast cancer (p) | Intensity, area, density, collagen reticular index (junctions:length), fiber length, fiber thickness | SHG | Increased amount of aggregated collagen in the tissue, brighter collagen fibers, are associated with increased survival. | [92] |
| Breast cancer (p) | COX2, TAMs, collagen alignment | SHG, Masson’s Trichrome, Immunofluorescence | Collagen orientation perpendicular to the tumor boundary is associated with poor overall survival. | [86,99] |
| Breast cancer (d) | Fiber shape | SHG | Shape different between normal, benign, and malignant breast. | [100] |
| Breast cancer (d) | Relative angle | SHG | TACS as a function of disease progression: More aligned collagen associated with invasion. | [8] |
| Breast cancer (d) | d15/d31 tensor element ratio of the second order susceptibility χ2 | SHG | Malignant tissue contains locally aligned fibers compared to normal, hyperplastic, and dysplastic tissues. | [101] |
| Breast cancer (p) | Relative angle | SHG | Radially aligned fibrillar collagen is a poor prognostic factor. | [46] |
| Breast cancer (xenograft) | Alignment | SHG | Significantly less aligned collagen after trastuzumab treatment. | [53] |
| Colon Cancer (d) | Fiber width, straightness, alignment | SHG, EM | Changes in collagen alignment are apparent 10-20cm from the tumor. Increase in collagen width, straightness, and alignment further from the tumor. | [89] |
| Colorectal cancer (d) | Average intensity per pixel (AIPP) and fiber alignment | SHG | SHG signal intensity can differentiate malignant from non-malignant colonic polyp tissue with high sensitivity and specificity. Anisotropic polarization can discern high-grade dysplasia from normal colonic polyp tissue. | [96,102] |
| Esophageal Cancer (p) | Length | SHG | Elongated collagen fibers are associated with poorer prognosis, alignment did not predict survival. | [10] |
| Gastric cancer (d) | Average intensity per pixel | MPM, SHG | Increase in overall collagen intensity in early gastric cancer vs normal | [103] |
| Gastric cancer (d) | CT-FIRE (alignment, length, straightness, width), | SHG, IHC (ColI, PICP, PINP, LOX, LOXL2) | Increased stromal collagen alignment, length, straightness, and width in gastric cancer. | [88] |
| Glioblastoma (p) | Alignment | SHG | More organized collagen is associated with better prognosis. | [95] |
| Head and neck, (p) | Length | SHG | Elongated collagen fibers are associated with poorer prognosis, alignment did not predict survival. | [10] |
| Hepatocellular carcinoma (d) | Aggregated and distributed collagen fiber ratio, individual percentage, number, length, width, and cross-link density | SHG | Collagen architecture varies with different grades of HCC; can be used to accurately predict HCC grading. | [90] |
| Lung cancer | Alignment | SHG | Aligned collagen prevents T cells from migrating to tumor islets. | [13] |
| Non-small cell lung carcinoma (d) | second-order susceptibility component ratio | Polarization-in, polarization-out SHG | Collagen in less compact and has larger disorder in tumor tissue. | [104] |
| Oral squamous cell carcinoma (d) | Fiber thickness | Picrosirius Red | Thin fibers increased and thick fibers decreased with increasing grade. | [91] |
| Oral squamous cell carcinoma (d) | Alignment | Picrosirius Red | Parallel fibers in neoplastic regions. | [105] |
| Osteosarcoma, breast cancer, melanoma (d) | d22 coefficient and anisotropy | SHG | Structure different in tumor tissue, low collagen density correlates with cancer. | [106] |
| Ovarian cancer (d) | Gray-level co-occurrence matrix to measure fibril size and separation | SHG, TPEF | Normal tissue is more highly structured. Loss of fine structure and structural organization with wavy collagen fibers in ovarian cancer. | [107] |
| Ovarian cancer (d) | SHG emission attributes (directionality and relative intensity) and bulk optical parameters | SHG | Malignant tissue has higher fiber regularity. | [9] |
| Ovarian cancer (d) | Alignment of collagen fibers, anisotropy, and correlation | SHG | Significant differences in the distribution and organization of collagen fibers in the stroma component of serous, mucinous, endometrioid, and mixed ovarian tumors as compared with normal ovary tissue. | [108] |
| Ovarian cancer (d) | Intensity, scattering, anisotropy | SHG, Optical Scattering | Collagen fibers are more organized in high-grade serous compared to normal tissue. | [109] |
| Pancreatic ductal adenocarcinoma (p) | Fiber alignment | SLIM | Inverse relationship between survival data and fiber width and length. | [97] |
| Pancreatic ductal adenocarcinoma (p) | Alignment | SHG | High alignment is a poor prognostic factor. | [59,110] |
| Pancreatic ductal adenocarcinoma (d) | Alignment | SHG | Increased alignment, length, and width in PDAC vs. normal and chronic pancreatitis. | [45] |
| Prostate cancer (d) | Fiber orientation, anisotropy | SHG, US | Increased fiber alignment associated with higher Gleason score. | [111] |
| Prostate cancer (d) | Trained CNN using SHG | SHG, CNN | TPEF and SHG can be used in combination with deep learning for accurate and automated Gleason grading of unstained prostate tissues. | [82] |
| Prostate cancer (d) | Anisotropy vs Gleason (ImageJ and FibrilTool) | SHG | Increased fiber alignment associated with higher cancer grading. | [93] |
| Renal cell carcinoma (d) | Alignment and density | SHG | Increased fiber density and alignment in RCC grade 4 compared to grade 1. | [94] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ouellette, J.N.; Drifka, C.R.; Pointer, K.B.; Liu, Y.; Lieberthal, T.J.; Kao, W.J.; Kuo, J.S.; Loeffler, A.G.; Eliceiri, K.W. Navigating the Collagen Jungle: The Biomedical Potential of Fiber Organization in Cancer. Bioengineering 2021, 8, 17. https://doi.org/10.3390/bioengineering8020017
Ouellette JN, Drifka CR, Pointer KB, Liu Y, Lieberthal TJ, Kao WJ, Kuo JS, Loeffler AG, Eliceiri KW. Navigating the Collagen Jungle: The Biomedical Potential of Fiber Organization in Cancer. Bioengineering. 2021; 8(2):17. https://doi.org/10.3390/bioengineering8020017
Chicago/Turabian StyleOuellette, Jonathan N., Cole R. Drifka, Kelli B. Pointer, Yuming Liu, Tyler J Lieberthal, W John Kao, John S. Kuo, Agnes G. Loeffler, and Kevin W. Eliceiri. 2021. "Navigating the Collagen Jungle: The Biomedical Potential of Fiber Organization in Cancer" Bioengineering 8, no. 2: 17. https://doi.org/10.3390/bioengineering8020017
APA StyleOuellette, J. N., Drifka, C. R., Pointer, K. B., Liu, Y., Lieberthal, T. J., Kao, W. J., Kuo, J. S., Loeffler, A. G., & Eliceiri, K. W. (2021). Navigating the Collagen Jungle: The Biomedical Potential of Fiber Organization in Cancer. Bioengineering, 8(2), 17. https://doi.org/10.3390/bioengineering8020017