Biotechnological Production of Poly(3-Hydroxybutyrate-co-4-Hydroxybutyrate-co-3-Hydroxyvalerate) Terpolymer by Cupriavidus sp. DSM 19379



Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganisms and Cultivation
2.1.1. Single-Stage Cultivation Mode
2.1.2. Two-Stage Cultivation Mode
2.2. Determination of the CDM and PHA Content
2.3. Polymer Characterization
3. Results and Discussion
3.1. Biosynthesis of P(3HB-co-4HB) Copolymer
3.2. Biosynthesis of P(3HB-co-3HV-co-4HB) Terpolymer through Single-Stage Cultivation
3.3. Biosynthesis of the P(3HB-co-3HV-co-4HB)Tterpolymer through the Two-Stage Cultivation
3.4. Characteristics of Isolated Polymers
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kourmentza, C.; Plácido, J.; Venetsaneas, N.; Burniol-Figols, A.; Varrone, C.; Gavala, H.N.; Reis, M.A. Recent advances and challenges towards sustainable polyhydroxyalkanoate (PHA) production. Bioengineering 2017, 4, 55. [Google Scholar] [CrossRef]
- Obruca, S.; Sedlacek, P.; Koller, M.; Kucera, D.; Pernicova, I. Involvement of polyhydroxyalkanoates in stress resistance of microbial cells: Biotechnological consequences and applications. Biotechnol. Adv. 2018, 36, 856–870. [Google Scholar] [CrossRef] [PubMed]
- Slaninova, E.; Sedlacek, P.; Mravec, F.; Mullerova, L.; Samek, O.; Koller, M.; Hesko, O.; Kucera, D.; Marova, I.; Obruca, S. Light scattering on PHA granules protects bacterial cells against the harmful effects of UV radiation. Appl. Microbiol. Biotechnol. 2018, 102, 1923–1931. [Google Scholar] [CrossRef]
- Haas, C.; Steinwandter, V.; De Apodaca, E.D.; Madurga, B.M.; Smerilli, M.; Dietrich, T.; Neureiter, M. Production of PHB from chicory roots - Comparison of three Cupriavidus necator strains. Chem. Biochem. Eng. Q. 2015, 29, 99–112. [Google Scholar] [CrossRef]
- Verlinden, R.A.J.; Hill, D.J.; Kenward, M.A.; Williams, C.D.; Piotrowska-Seget, Z.; Radecka, I.K. Production of polyhydroxyalkanoates from waste frying oil by Cupriavidus necator. Amb Express 2011, 1, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Ciesielski, S.; Mozejko, J.; Pisutpaisal, N. Plant oils as promising substrates for polyhydroxyalkanoates production. J. Clean. Prod. 2015, 106, 408–421. [Google Scholar] [CrossRef]
- Jiang, G.; Hill, D.J.; Kowalczuk, M.; Johnston, B.; Adamus, G.; Irorere, V.; Radecka, I. Carbon sources for polyhydroxyalkanoates and an integrated biorefinery. Int. J. Mol. Sci. 2016, 17, 1157. [Google Scholar] [CrossRef] [PubMed]
- Moita, R.; Freches, A.; Lemos, P.C. Crude glycerol as feedstock for polyhydroxyalkanoates production by mixed microbial cultures. Water Res. 2014, 58, 9–20. [Google Scholar] [CrossRef]
- Obruca, S.; Benesova, P.; Marsalek, L.; Marova, I. Use of lignocellulosic materials for PHA production. Chem. Biochem. Eng. Q. 2015, 29, 135–144. [Google Scholar] [CrossRef]
- Meixner, K.; Kovalcik, A.; Sykacek, E.; Gruber-Brunhumer, M.; Zeilinger, W.; Markl, K.; Haas, C.; Fritz, I.; Mundigler, N.; Stelzer, F.; et al. Cyanobacteria Biorefinery—Production of poly(3-hydroxybutyrate) with Synechocystis salina and utilisation of residual biomass. J. Biotechnol. 2018, 265, 46–53. [Google Scholar] [CrossRef]
- Troschl, C.; Meixner, K.; Drosg, B. Cyanobacterial PHA Production—Review of Recent Advances and a Summary of Three Years’ Working Experience Running a Pilot Plant. Bioengineering 2017, 4, 26. [Google Scholar] [CrossRef]
- Sedlacek, P.; Slaninova, E.; Enev, V.; Koller, M.; Nebesarova, J.; Marova, I.; Hrubanova, K.; Krzyzanek, V.; Samek, O.; Obruca, S. What keeps polyhydroxyalkanoates in bacterial cells amorphous? A derivation from stress exposure experiments. Appl. Microbiol. Biotechnol. 2019, 103, 1905–1917. [Google Scholar] [CrossRef]
- Koller, M. Chemical and biochemical engineering approaches in manufacturing polyhydroxyalkanoate (PHA) biopolyesters of tailored structure with focus on the diversity of building blocks. Chem. Biochem. Eng. Q. 2018, 32, 413–438. [Google Scholar] [CrossRef]
- Lee, W.H.; Azizan, M.N.M.; Sudesh, K. Effects of culture conditions on the composition of poly(3-hydroxybutyrate-co-4-hydroxybutyrate) synthesized by Comamonas acidovorans. Polym Degrad Stab. 2004, 84, 129–134. [Google Scholar] [CrossRef]
- Rodríguez-Contreras, A.; Calafell-Monfort, M.; Marqués-Calvo, M.S. Enzymatic degradation of poly(3-hydroxybutyrate-co-4-hydroxybutyrate) by commercial lipases. Polym. Degrad. Stabil. 2012, 97, 597–604. [Google Scholar] [CrossRef]
- Saito, Y.; Nakamura, S.; Hiramitsu, M.; Doi, Y. Microbial synthesis and properties of poly(3-hydroxybutyrate-co-4-hydroxybutyrate). Polym. Int. 1996, 39, 167–174. [Google Scholar] [CrossRef]
- Singh, A.K.; Srivastava, J.K.; Chandel, A.K.; Sharma, L.; Mallick, N.; Singh, S.P. Biomedical applications of microbially engineered polyhydroxyalkanoates: an insight into recent advances, bottlenecks, and solutions. Appl. Microbiol. Biotechnol. 2019, 103, 2007–2032. [Google Scholar] [CrossRef]
- Chanprateep, S.; Kulpreecha, S. Production and characterization of biodegradable terpolymer poly(3-hydroxybutyrate-co-3-hydroxyvalerate-co-4-hydroxybutyrate) by Alcaligenes sp. A-04. J. Biosci. Bioeng. 2006, 101, 51–56. [Google Scholar] [CrossRef]
- Lee, Y.H.; Kang, M.S.; Jung, Y.M. Regulating the molar fraction of 4-hydroxybutyrate in poly(3-hydroxybutyrate-4-hydroxybutyrate) biosynthesis by Ralstonia eutropha using propionate as a stimulator. J. Biosci. Bioeng. 2000, 89, 380. [Google Scholar] [CrossRef]
- Cavalheiro, J.M.; Raposo, R.S.; de Almeida, M.C.M.; Cesário, M.T.; Sevrin, C.; Grandfils, C.; Da Fonseca, M.M.R. Effect of cultivation parameters on the production of poly(3-hydroxybutyrate-co-4-hydroxybutyrate) and poly(3-hydroxybutyrate-4-hydroxybutyrate-3-hydroxyvalerate) by Cupriavidus necator using waste glycerol. Biores. Technol. 2012, 111, 391. [Google Scholar] [CrossRef]
- Hermann-Krauss, C.; Koller, M.; Muhr, A.; Fasl, H.; Stelzer, F.; Braunegg, G. Archaeal production of polyhydroxyalkanoate (PHA) co-and terpolyesters from biodiesel industry-derived by-products. Archaea 2013, 2013, 129268. [Google Scholar] [CrossRef]
- Ramachandran, H.; Iqbal, N.M.; Sipaut, C.S.; Abdullah, A.A.A. Biosynthesis and characterization of poly (3-hydroxybutyrate-co-3-hydroxyvalerate-co-4-hydroxybutyrate). Terpolymer with various monomer compositions by Cupriavidus sp. USMAA2-4. Appl. Biochem. Biotechnol. 2011, 164, 867–877. [Google Scholar] [CrossRef]
- Obruca, S.; Marova, I.; Melusova, S.; Mravcova, L. Production of polyhydroxyalkanoates from cheese whey employing Bacillus megaterium CCM 2037. Ann. Microbiol. 2011, 61, 947–953. [Google Scholar] [CrossRef]
- Brandl, H.; Gross, R.A.; Lenz, R.W.; Fuller, R.C. Pseudomonas oleovorans as a source of poly(beta-hydroxyalkanoates) for potential application as a biodegradable polyester. Appl. Environ. Microb. 1988, 54, 1977–1982. [Google Scholar]
- Kucera, D.; Pernicová, I.; Kovalcik, A.; Koller, M.; Mullerova, L.; Sedlacek, P.; Mravec, F.; Nebesarova, J.; Kalina, M.; Marova, I.; et al. Characterization of the promising poly(3-hydroxybutyrate) producing halophilic bacterium Halomonas halophila. Biores. Technol. 2018, 256, 552–556. [Google Scholar] [CrossRef]
- Amirul, A.A.; Yahya, A.R.M.; Sudesh, K.; Azizan, M.N.M.; Majid, M.I.A. Biosynthesis of poly(3-hydroxybutyrate-co-4-hydroxybutyrate) copolymer by Cupriavidus sp. USMAA1020 isolated from Lake Kulim, Malaysia. Biores. Technol. 2008, 99, 4903–4909. [Google Scholar]
- Rahayu, A.; Zaleha, Z.; Yahya, A.R.M.; Majid, M.I.A.; Amirul, A. A Production of copolymer poly(3-hydroxybutyrate-co-4-hydroxybutyrate) through a one-step cultivation process. World J. Microbiol. Biotechnol. 2008, 24, 2403–2409. [Google Scholar] [CrossRef]
- Lopar, M.; Špoljarić, I.V.; Cepanec, N.; Koller, M.; Braunegg, G.; Horvat, P. Study of metabolic network of Cupriavidus necator DSM 545 growing on glycerol by applying elementary flux modes and yield space analysis. J. Ind. Microbiol. Biotechnol. 2014, 41, 913–930. [Google Scholar] [CrossRef]
- Lindenkamp, N.; Peplinski, K.; Volodina, E.; Ehrenreich, A.; Steinbuchel, A. Impact of multiple beta-ketothiolase deletion mutations in Ralstonia eutropha H16 on the composition of 3-mercaptopropionic acid-containing copolymers. Appl. Environ. Microbiol. 2010, 76, 5373–5382. [Google Scholar] [CrossRef]
- Fahima Azira, T.M.; Nursolehah, A.A.; Norhayati, Y.; Majid, M.I.A.; Amirul, A.A. Biosynthesis of Poly(3-hydroxybutyrate-co-3-hydroxyvalerate-co-4-hydroxybutyrate) terpolymer by Cupriavidus sp. USMAA2-4 through two-step cultivation process. World J. Microbiol. Biotechnol. 2011, 27, 2287–2295. [Google Scholar] [CrossRef]


{kind=link}
| Substrate | CDM (g/L) | PHA (wt%) | PHA (g/L) | 4HB (mol%) | 3HB (mol%) |
|---|---|---|---|---|---|
| fructose | 10.78 ± 0.06 | 69.95 ± 0.42 | 7.54 ± 0.10 | 0 | 100 |
| glucose | 2.29 ± 0.06 | 10.03 ± 0.06 | 0.23 ± 0.02 | 0 | 100 |
| glycerol | 4.60 ± 0.04 | 5.30 ± 0.05 | 0.24 ± 0.02 | 0 | 100 |
| sunflower oil | 1.33 ± 0.05 | 0 | 0 | 0 | 0 |
| GBL | 4.50 ± 0.02 | 35.84 ± 0.92 | 1.61 ± 0.12 | 22.18 ± 1.06 | 77.82 ± 1.06 |
| 1,4-butanediol | 4.01 ± 0.02 | 11.67 ± 0.06 | 0.47 ± 0.03 | 23.12 ± 1.61 | 76.88 ± 1.61 |
| ε-caprolactone | 0.22 ± 0.04 | 42.80 ± 0.61 | 0.10 ± 0.04 | 68.89 ± 1.12 | 31.11 ± 1.12 |
| 1,6-hexanediol | 2.64 ± 0.01 | 39.83 ± 0.95 | 1.05 ± 0.07 | 34.35 ± 0.96 | 65.65 ± 0.96 |
| Primary Substrate | 3HV Precursor | CDM (g/L) | PHA (g/L) | PHA (wt%) | 3HB (mol%) | 4HB (mol%) | 3HV (mol%) |
|---|---|---|---|---|---|---|---|
| GBL | none | 3.64 ± 0.03 | 0.81 ± 0.05 | 22.14 ± 0.01 | 68.40 ± 0.23 | 31.60 ± 0.23 | 0 |
| propionic acid | 5.06 ± 0.37 | 0.62 ± 0.06 | 12.16 ± 0.00 | 69.18 ± 0.22 | 23.41 ± 0.05 | 7.41 ± 0.16 | |
| valeric acid | 7.97 ± 1.85 | 0.82 ± 0.09 | 10.41 ± 0.01 | 56.22 ± 0.32 | 25.85 ± 0.40 | 17.92 ± 0.07 | |
| 1,4-butanediol | none | 7.41 ± 0.51 | 1.05 ± 0.19 | 14.44 ± 0.02 | 68.97 ± 2.26 | 31.03 ± 2.26 | 0 |
| propionic acid | 8.19 ± 0.35 | 1.65 ± 0.43 | 20.01 ± 0.04 | 63.81 ± 1.71 | 27.87 ± 0.10 | 8.32 ± 1.80 | |
| valeric acid | 8.68 ± 0.14 | 1.79 ± 0.88 | 20.52 ± 0.10 | 60.63 ± 2.90 | 24.72 ± 7.42 | 14.65 ± 4.53 |
| Primary Substrate | Secondary Precursor | CDM (g/L) | PHA (g/L) | PHA (wt%) | 3HB (mol%) | 4HB (mol%) | 3HV (mol%) |
|---|---|---|---|---|---|---|---|
| Glycerol | 1,4-butanediol | 1.60 ± 0.03 | 0.84 ± 0.02 | 52.25 ± 0.12 | 80.85 ± 0.68 | 18.09 ± 0.26 | 1.06 ± 0.43 |
| 1,4-butanediol + valeric acid | 2.73 ± 0.58 | 1.42 ± 0.25 | 52.12 ± 1.76 | 53.78 ± 0.61 | 16.76 ± 0.87 | 29.46 ± 0.26 | |
| Glycerol + 1,4-butanediol | 1,4-butanediol | 3.26 ± 0.11 | 2.09 ± 0.01 | 64.14 ± 2.38 | 77.89 ± 0.53 | 21.60 ± 0.54 | 0.51 ± 0.01 |
| 1,4-butanediol + valeric acid | 5.94 ± 0.14 | 4.14 ± 0.05 | 69.64 ± 0.73 | 65.68 ± 1.02 | 16.46 ± 1.28 | 17.86 ± 0.26 |
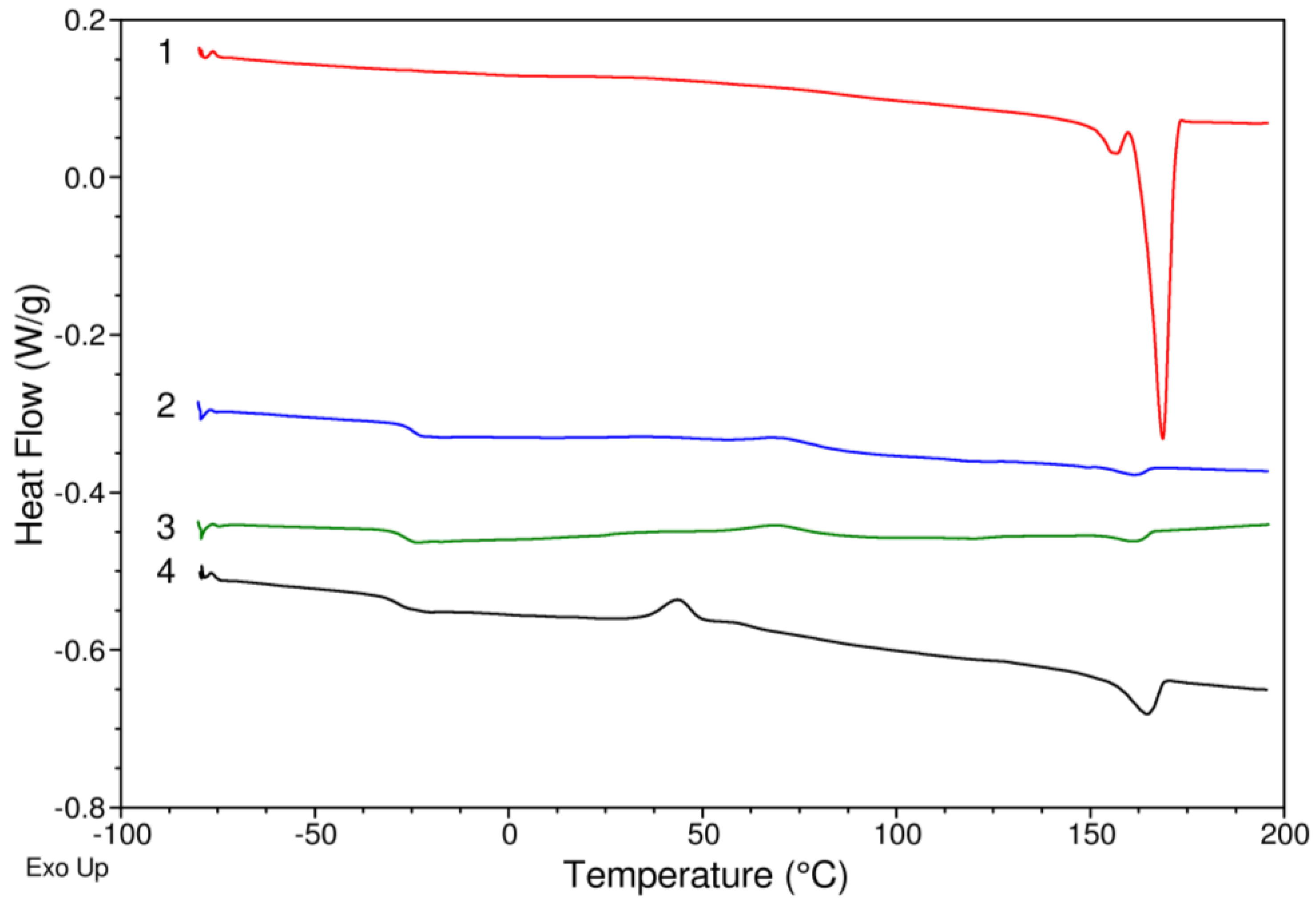
| Sample | 3HB (mol%) | 4HB (mol%) | 3HV (mol%) | Mw (kDa) | Đ (-) | Tg (°C) | Tm (°C) | ΔH (J/g) | Tdonset (°C) | Tdmax (°C) |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 99.33 | 0.67 | 0 | 155.97 | 1.04 | - | 155.79 | 4.70 | 271.88 | 287.94 |
| 168.70 | 64.89 | |||||||||
| 2 | 60.63 | 24.72 | 14.65 | 258.66 | 1.02 | 24.78 | 161.34 | 2.80 | 271.48 | 293.49 |
| 3 | 63.81 | 27.87 | 8.32 | 314.60 | 1.01 | 26.19 | 161.67 | 3.04 | 275.24 | 300.83 |
| 4 | 53.78 | 16.76 | 29.46 | 137.89 | 1.17 | 29.00 | 164.63 | 12.69 | 271.36 | 295.53 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kucera, D.; Novackova, I.; Pernicova, I.; Sedlacek, P.; Obruca, S. Biotechnological Production of Poly(3-Hydroxybutyrate-co-4-Hydroxybutyrate-co-3-Hydroxyvalerate) Terpolymer by Cupriavidus sp. DSM 19379. Bioengineering 2019, 6, 74. https://doi.org/10.3390/bioengineering6030074
Kucera D, Novackova I, Pernicova I, Sedlacek P, Obruca S. Biotechnological Production of Poly(3-Hydroxybutyrate-co-4-Hydroxybutyrate-co-3-Hydroxyvalerate) Terpolymer by Cupriavidus sp. DSM 19379. Bioengineering. 2019; 6(3):74. https://doi.org/10.3390/bioengineering6030074
Chicago/Turabian StyleKucera, Dan, Ivana Novackova, Iva Pernicova, Petr Sedlacek, and Stanislav Obruca. 2019. "Biotechnological Production of Poly(3-Hydroxybutyrate-co-4-Hydroxybutyrate-co-3-Hydroxyvalerate) Terpolymer by Cupriavidus sp. DSM 19379" Bioengineering 6, no. 3: 74. https://doi.org/10.3390/bioengineering6030074
APA StyleKucera, D., Novackova, I., Pernicova, I., Sedlacek, P., & Obruca, S. (2019). Biotechnological Production of Poly(3-Hydroxybutyrate-co-4-Hydroxybutyrate-co-3-Hydroxyvalerate) Terpolymer by Cupriavidus sp. DSM 19379. Bioengineering, 6(3), 74. https://doi.org/10.3390/bioengineering6030074