
Estimating Extrinsic Dyes for Fluorometric Online Monitoring of Antibody Aggregation in CHO Fed-Batch Cultivations

Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Line, Medium, and Culture Conditions
2.2. IC50 Experiments
2.3. Fed-Batch Cultures with Fluorescence Dyes
2.4. Offline Analytics
2.5. Online Data Collection
2.6. Chemometric Modeling and Data Preprocessing
3. Results
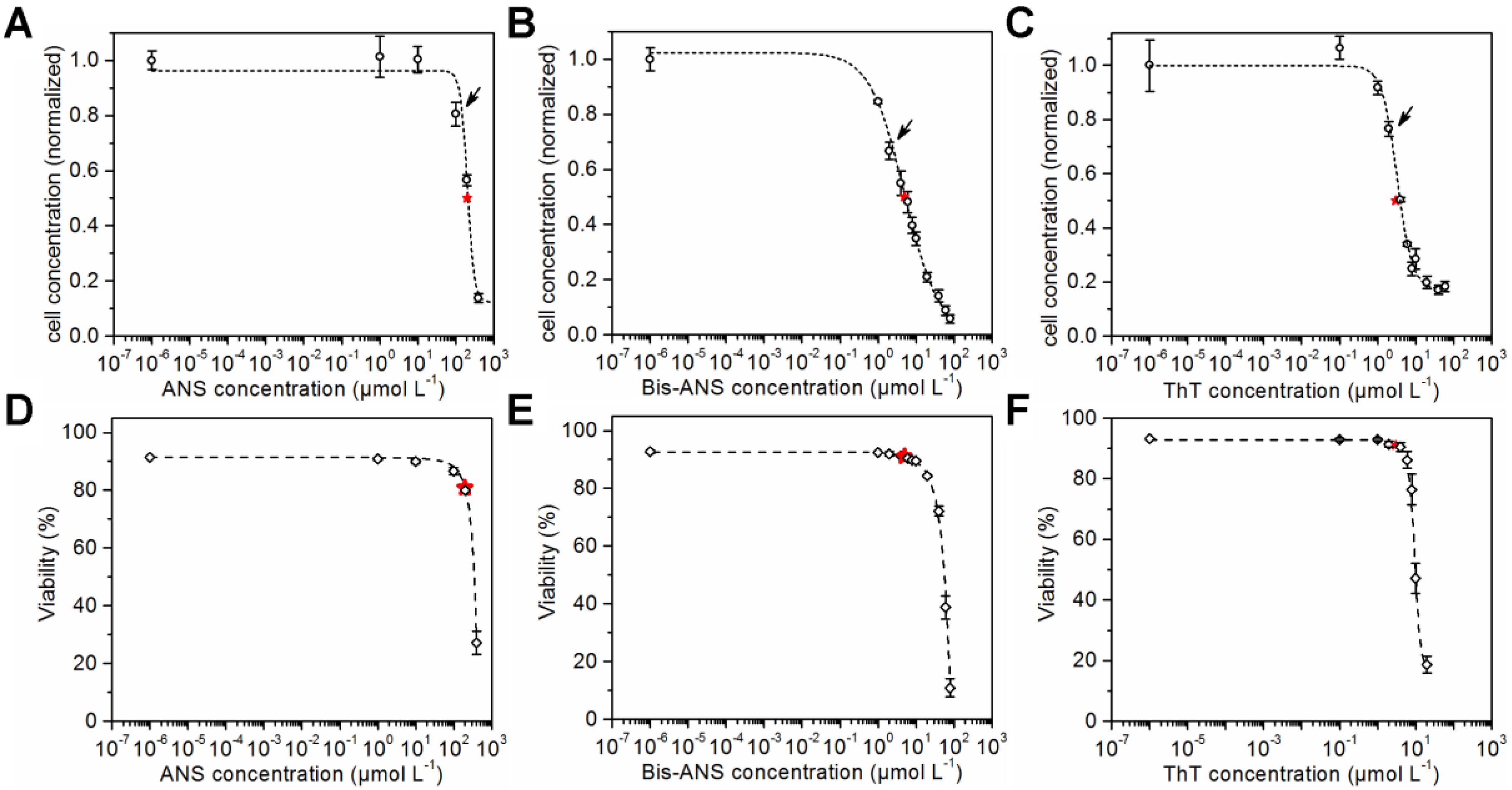
3.1. IC50 Experiments
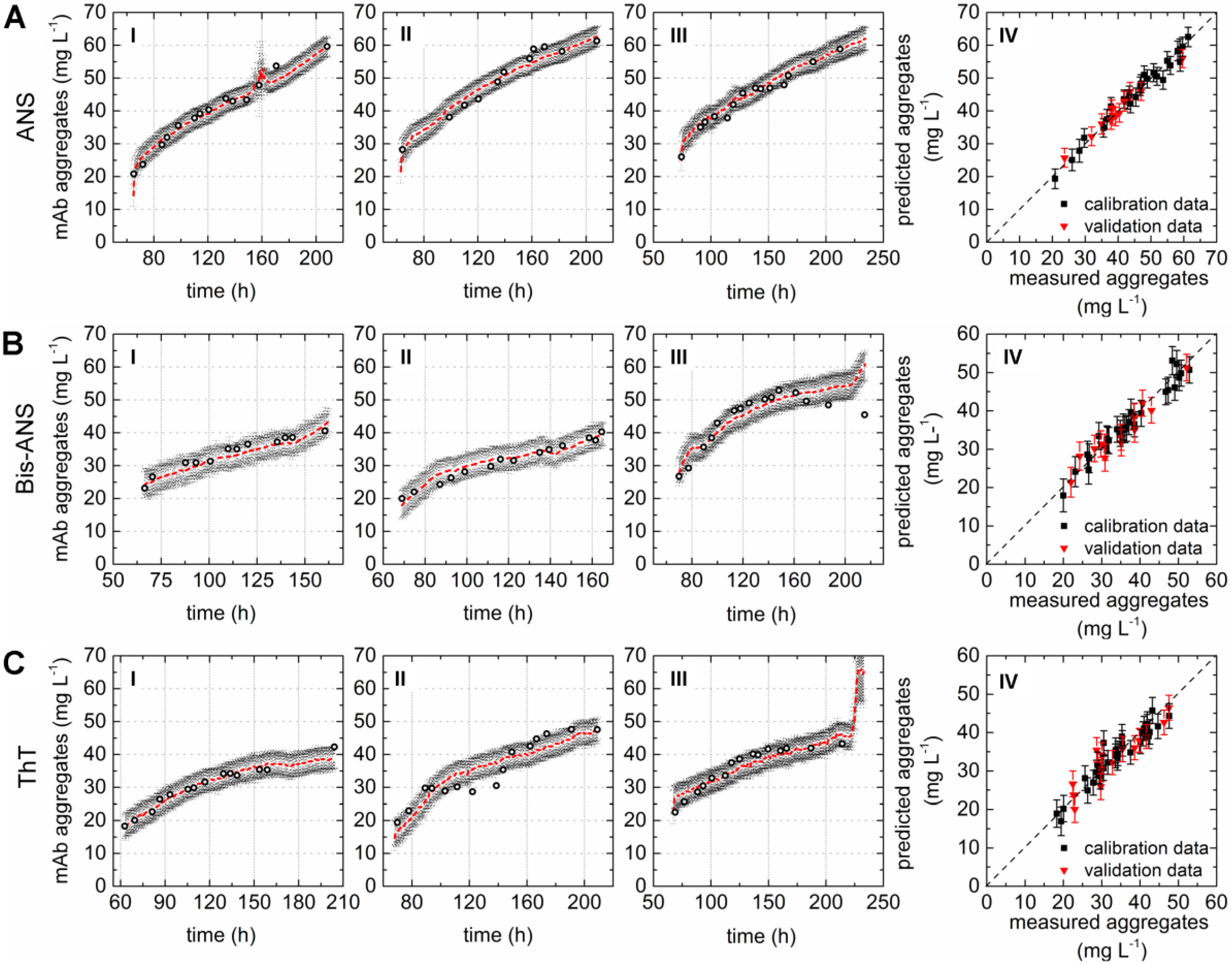
3.2. Online Monitoring Based on Extrinsic Fluorescence
4. Discussion
4.1. Fluorescence Dye Toxicity
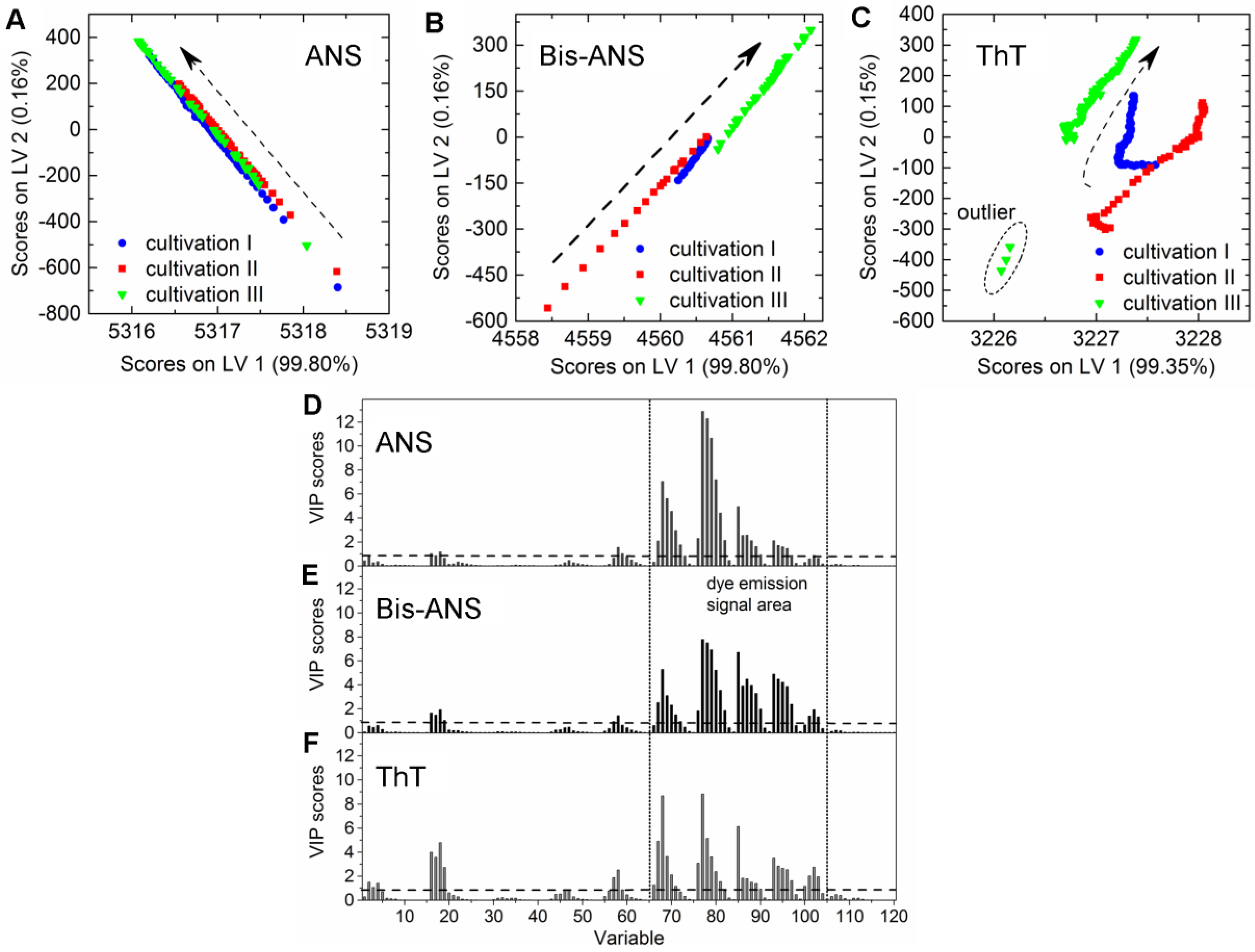
4.2. Extrinsic Dye-Related Emission Signals
4.3. Chemometric Modeling
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Luttmann, R.; Bracewell, D.G.; Cornelissen, G.; Gernaey, K.V.; Glassey, J.; Hass, V.C.; Kaiser, C.; Preusse, C.; Striedner, G.; Mandenius, C.F. Soft sensors in bioprocessing: A status report and recommendations. Biotechnol. J. 2012, 7, 1040–1048. [Google Scholar] [CrossRef] [PubMed]
- Mercier, S.M.; Diepenbroek, B.; Dalm, M.C.; Wijffels, R.H.; Streefland, M. Multivariate data analysis as a PAT tool for early bioprocess development data. J. Biotechnol. 2013, 167, 262–270. [Google Scholar] [CrossRef] [PubMed]
- Glassey, J.; Gernaey, K.; Clemens, C.; Schulz, T.W.; Oliveira, R.; Striedner, G.; Mandenius, C.F. Process analytical technology (PAT) for biopharmaceuticals. Biotechnol. J. 2011, 6, 369–377. [Google Scholar] [CrossRef] [PubMed]
- Mercier, S.M.; Diepenbroek, B.; Wijffels, R.H.; Streefland, M. Multivariate PAT solutions for biopharmaceutical cultivation: Current progress and limitations. Trends Biotechnol. 2014, 32, 329–336. [Google Scholar] [CrossRef] [PubMed]
- Wold, S.; Sjöström, M.; Eriksson, L. PLS-regression: A basic tool of chemometrics. Chemom. Intell. Lab. Syst. 2001, 58, 109–130. [Google Scholar] [CrossRef]
- Cruz, M.V.; Sarraguça, M.C.; Freitas, F.; Lopes, J.A.; Reis, M.A.M. Online monitoring of P(3HB) produced from used cooking oil with near-infrared spectroscopy. J. Biotechnol. 2015, 194, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Jain, G.; Jayaraman, G.; Kökpinar, Ö.; Rinas, U.; Hitzmann, B. On-line monitoring of recombinant bacterial cultures using multi-wavelength fluorescence spectroscopy. Biochem. Eng. J. 2011, 58–59, 133–139. [Google Scholar] [CrossRef]
- Mazarevica, G.; Diewok, J.; Baena, J.R.; Rosenberg, E.; Lendl, B. On-line fermentation monitoring by mid-infrared spectroscopy. Appl. Spectrosc. 2004, 58, 804–810. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, A.P.; Portugal, C.A.M.; Carinhas, N.; Dias, J.M.L.; Crespo, J.P.; Alves, P.M.; Carrondo, M.J.T.; Oliveira, R. In situ 2D fluorometry and chemometric monitoring of mammalian cell cultures. Biotechnol. Bioeng. 2009, 102, 1098–1106. [Google Scholar] [CrossRef] [PubMed]
- Ohadi, K.; Aghamohseni, H.; Legge, R.L.; Budman, H.M. Fluorescence-based soft sensor for at situ monitoring of chinese hamster ovary cell cultures. Biotechnol. Bioeng. 2014, 111, 1577–1586. [Google Scholar] [CrossRef] [PubMed]
- Vázquez-Rey, M.; Lang, D.A. Aggregates in monoclonal antibody manufacturing processes. Biotechnol. Bioeng. 2011, 108, 1494–1508. [Google Scholar] [CrossRef] [PubMed]
- Ring, J.; Seifert, J.; Jesch, F.; Brendel, W. Anaphylactoid reactions due to non-immune complex serum protein aggregates. Monogr. Allergy 1976, 12, 27–35. [Google Scholar]
- Cromwell, M.E.; Hilario, E.; Jacobson, F. Protein aggregation and bioprocessing. AAPS J. 2006, 8, E572–E579. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.B.; Howitt, J.; McCorkle, S.; Lawrence, P.; Springer, K.; Freimuth, P. Protein aggregation during overexpression limited by peptide extensions with large net negative charge. Protein Expr. Purif. 2004, 36, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Gomez, N.; Subramanian, J.; Ouyang, J.; Nguyen, M.D.; Hutchinson, M.; Sharma, V.K.; Lin, A.A.; Yuk, I.H. Culture temperature modulates aggregation of recombinant antibody in CHO cells. Biotechnol. Bioeng. 2012, 109, 125–136. [Google Scholar] [CrossRef] [PubMed]
- Paul, A.J.; Schwab, K.; Prokoph, N.; Haas, E.; Handrick, R.; Hesse, F. Fluorescence dye-based detection of mAb aggregates in CHO culture supernatants. Anal. Bioanal. Chem. 2015, 407, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Renard, D.; Lefebvre, J.; Griffin, M.; Griffin, W. Effects of pH and salt environment on the association of β-lactoglobulin revealed by intrinsic fluorescence studies. Int. J. Biol. Macromol. 1998, 22, 41–49. [Google Scholar] [CrossRef]
- Ohadi, K.; Legge, R.L.; Budman, H.M. Intrinsic fluorescence-based at situ soft sensor for monitoring monoclonal antibody aggregation. Biotechnol. Prog. 2015, 31, 1423–1432. [Google Scholar] [CrossRef] [PubMed]
- Hawe, A.; Sutter, M.; Jiskoot, W. Extrinsic Fluorescent Dyes as Tools for Protein Characterization. Pharm. Res. 2008, 25, 1487–1499. [Google Scholar] [CrossRef] [PubMed]
- Khurana, R.; Coleman, C.; Ionescu-Zanetti, C.; Carter, S.A.; Krishna, V.; Grover, R.K.; Roy, R.; Singh, S. Mechanism of thioflavin T binding to amyloid fibrils. J. Struct. Biol. 2005, 151, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Jayaram, D.T.; Shankar, B.H.; Ramaiah, D. Effective Amyloid Defibrillation by Polyhydroxyl-Substituted Squaraine Dyes. Chem. Asian J. 2015, 10, 2689–2694. [Google Scholar] [CrossRef] [PubMed]
- Capelle, M.A.; Gurny, R.; Arvinte, T. High throughput screening of protein formulation stability: Practical considerations. Eur. J. Pharm. Biopharm. 2007, 65, 131–148. [Google Scholar] [CrossRef] [PubMed]
- Hawe, A.; Friess, W.; Sutter, M.; Jiskoot, W. Online fluorescent dye detection method for the characterization of immunoglobulin G aggregation by size exclusion chromatography and asymmetrical flow field flow fractionation. Anal. Biochem. 2008, 378, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Cummings, B.S.; Schnellmann, R.G. Measurement of cell death in mammalian cells. Curr. Protoc. Pharmacol. 2004. [Google Scholar] [CrossRef]
- Paul, A.J.; Schwab, K.; Hesse, F. Direct analysis of mAb aggregates in mammalian cell culture supernatant. BMC Biotechnol. 2014, 14, 99. [Google Scholar] [CrossRef] [PubMed]
- Marose, S.; Lindemann, C.; Scheper, T. Two-Dimensional Fluorescence Spectroscopy: A New Tool for On-Line Bioprocess Monitoring. Biotechnol. Prog. 1998, 14, 63–74. [Google Scholar] [CrossRef] [PubMed]
- Schwab, K.; Amann, T.; Schmid, J.; Handrick, R.; Hesse, F. Exploring the Capabilities of Fluorometric Online Monitoring on CHO Cell Cultivations Producing a Monoclonal Antibody. Biotechnol. Prog. 2016. [Google Scholar] [CrossRef] [PubMed]
- Schwab, K.; Lauber, J.; Hesse, F. Fluorometric In Situ Monitoring of an Escherichia coli Cell Factory with Cytosolic Expression of Human Glycosyltransferase GalNAcT2: Prospects and Limitations. Bioengineering 2016, 3, 32. [Google Scholar] [CrossRef]
- Wise, B.M.; Gallagher, N.B.; Bro, R.; Shaver, J.M. PLS Toolbox 3.0; Eigenvector Research Inc.: Manson, WA, USA, 2003. [Google Scholar]
- Roger, J.M.; Chauchard, F.; Bellon-Maurel, V. EPO-PLS external parameter orthogonalisation of PLS application to temperature-independent measurement of sugar content of intact fruits. Chemom. Intell. Lab. Syst. 2003, 66, 191–204. [Google Scholar] [CrossRef]
- Chong, I.G.; Jun, C.H. Performance of some variable selection methods when multicollinearity is present. Chemom. Intell. Lab. Syst. 2005, 78, 103–112. [Google Scholar] [CrossRef]
- He, F.; Phan, D.H.; Hogan, S.; Bailey, R.; Becker, G.W.; Narhi, L.O.; Razinkov, V.I. Detection of IgG aggregation by a high throughput method based on extrinsic fluorescence. J. Pharm. Sci. 2010, 99, 2598–2608. [Google Scholar] [CrossRef] [PubMed]
- Amani, S.; Nasim, F.; Khan, T.A.; Fazili, N.A.; Furkan, M.; Bhat, I.A.; Khan, J.M.; Khan, R.H.; Naeem, A. Detergent induces the formation of IgG aggregates: A multi-methodological approach. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2014, 120, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Franey, H.; Brych, S.R.; Kolvenbach, C.G.; Rajan, R.S. Increased aggregation propensity of IgG2 subclass over IgG1: Role of conformational changes and covalent character in isolated aggregates. Protein Sci. 2010, 19, 1601–1615. [Google Scholar] [CrossRef] [PubMed]
- Hawe, A.; Filipe, V.; Jiskoot, W. Fluorescent molecular rotors as dyes to characterize polysorbate-containing IgG formulations. Pharm. Res. 2010, 27, 314–326. [Google Scholar] [CrossRef] [PubMed]
- Bahram, M.; Bro, R.; Stedmon, C.; Afkhami, A. Handling of Rayleigh and Raman scatter for PARAFAC modeling of fluorescence data using interpolation. J. Chemom. 2006, 20, 99–105. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Start Concentration | |||||||
|---|---|---|---|---|---|---|---|
| Cultivation | Glucose (g·L−1) | Glutamine (mmol·L−1) | Cultivation Time (h) | Time of Dye Addition (h) | XV max (×106 mL−1) | mAb (mg·L−1) | Aggregated mAb (mg·L−1) |
| ANS I | 1.63 | 1.26 | 208 | 64 | 5.06 | 95 | 60 |
| ANS II | 1.90 | 1.75 | 208 | 63 | 4.33 | 89 | 61 |
| ANS III | 2.84 | 2.03 | 212 | 74 | 3.38 | 88 | 59 |
| Bis-ANS I | 1.92 | 2.14 | 161 | 66 | 4.86 | 61 | 41 |
| Bis-ANS II | 2.37 | 1.93 | 165 | 69 | 4.55 | 57 | 40 |
| Bis-ANS III | 2.40 | 2.02 | 215 | 70 | 3.00 | 71 | 45 |
| Th T I | 1.99 | 2.08 | 204 | 63 | 4.86 | 58 | 42 |
| Th T II | 1.93 | 1.91 | 209 | 68 | 4.06 | 65 | 48 |
| Th T III | 2.84 | 2.03 | 214 | 67 | 3.67 | 59 | 43 |
| Calibration | Cross-Validation | Prediction | ||||||
|---|---|---|---|---|---|---|---|---|
| Extrinsic Dye | Preprocessing | LV | R2cal | RMSEC [mg·mL−1] | R2CV | RMSECV [mg·mL−1] | R2P | RMSEP [mg·mL−1] |
| ANS | MSC (mean) EPO (1 PC) | 5 | 0.97 | 1.1 | 0.94 | 2.6 | 0.97 | 1.7 |
| Bis-ANS | MSC (mean) EPO (1 PC) | 5 | 0.96 | 1.9 | 0.89 | 3.2 | 0.92 | 2.3 |
| ThT | MSC (mean) | 6 | 0.92 | 2.2 | 0.85 | 3.0 | 0.85 | 3.1 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schwab, K.; Hesse, F. Estimating Extrinsic Dyes for Fluorometric Online Monitoring of Antibody Aggregation in CHO Fed-Batch Cultivations. Bioengineering 2017, 4, 65. https://doi.org/10.3390/bioengineering4030065
Schwab K, Hesse F. Estimating Extrinsic Dyes for Fluorometric Online Monitoring of Antibody Aggregation in CHO Fed-Batch Cultivations. Bioengineering. 2017; 4(3):65. https://doi.org/10.3390/bioengineering4030065
Chicago/Turabian StyleSchwab, Karen, and Friedemann Hesse. 2017. "Estimating Extrinsic Dyes for Fluorometric Online Monitoring of Antibody Aggregation in CHO Fed-Batch Cultivations" Bioengineering 4, no. 3: 65. https://doi.org/10.3390/bioengineering4030065
APA StyleSchwab, K., & Hesse, F. (2017). Estimating Extrinsic Dyes for Fluorometric Online Monitoring of Antibody Aggregation in CHO Fed-Batch Cultivations. Bioengineering, 4(3), 65. https://doi.org/10.3390/bioengineering4030065