

The Combination of PPARα Agonist GW7647 and Imeglimin Has Potent Effects on High-Glucose-Induced Cellular Biological Responses in Human Retinal Pigment Epithelium Cells

 , , , ,
, , , , 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture of Human Adult Retinal Pigment Epithelium 19 Cells (ARPE19 Cells)
2.2. Cell Viability Test
2.3. Measurement of Levels of Reactive Oxygen Species (ROS)
2.4. Monolayer Barrier Function of ARPE19 Cells Assessed by Transepithelial Electrical Resistance (TEER)
2.5. Seahorse Cellular Metabolic Function Measurement
2.6. Other Analytical Methods
3. Results
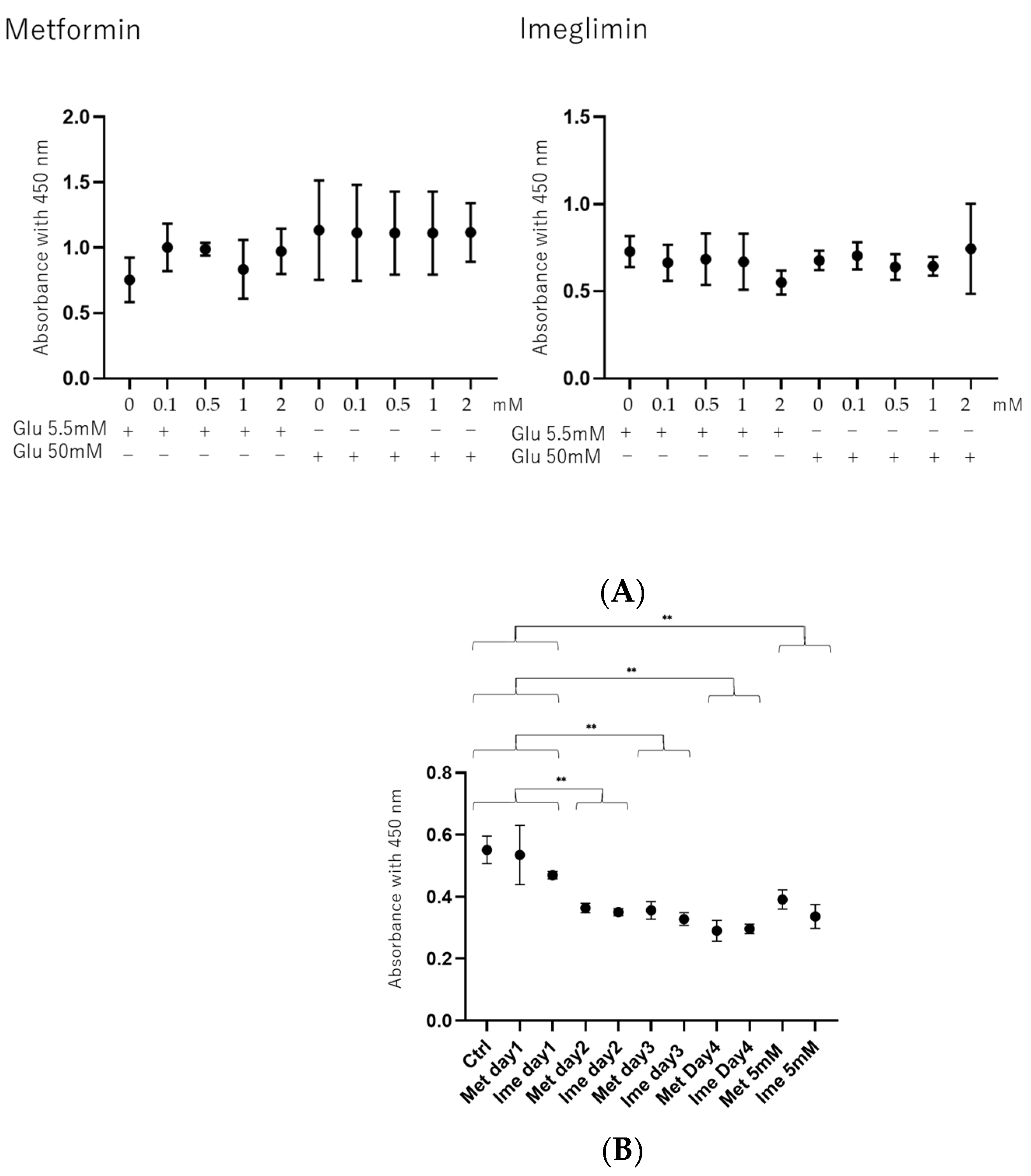
3.1. Effects of Concentrations of Met and Ime on Cell Viability in ARPE19 Cells Under Low- and High-Glucose Conditions
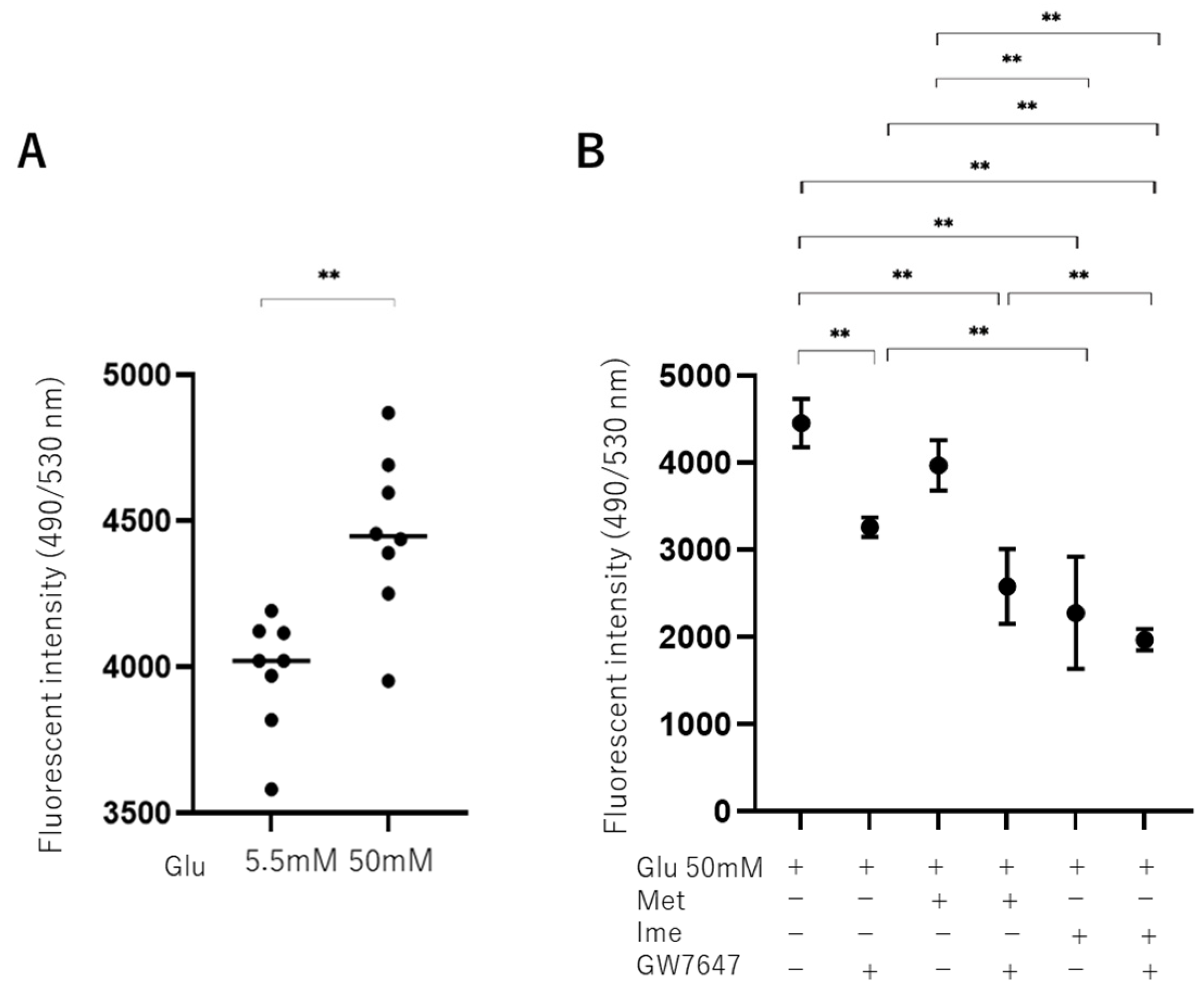
3.2. Effects of Met, Ime, and/or GW7647 on Levels of Reactive Oxygen Species (ROS) in ARPE19 Cells
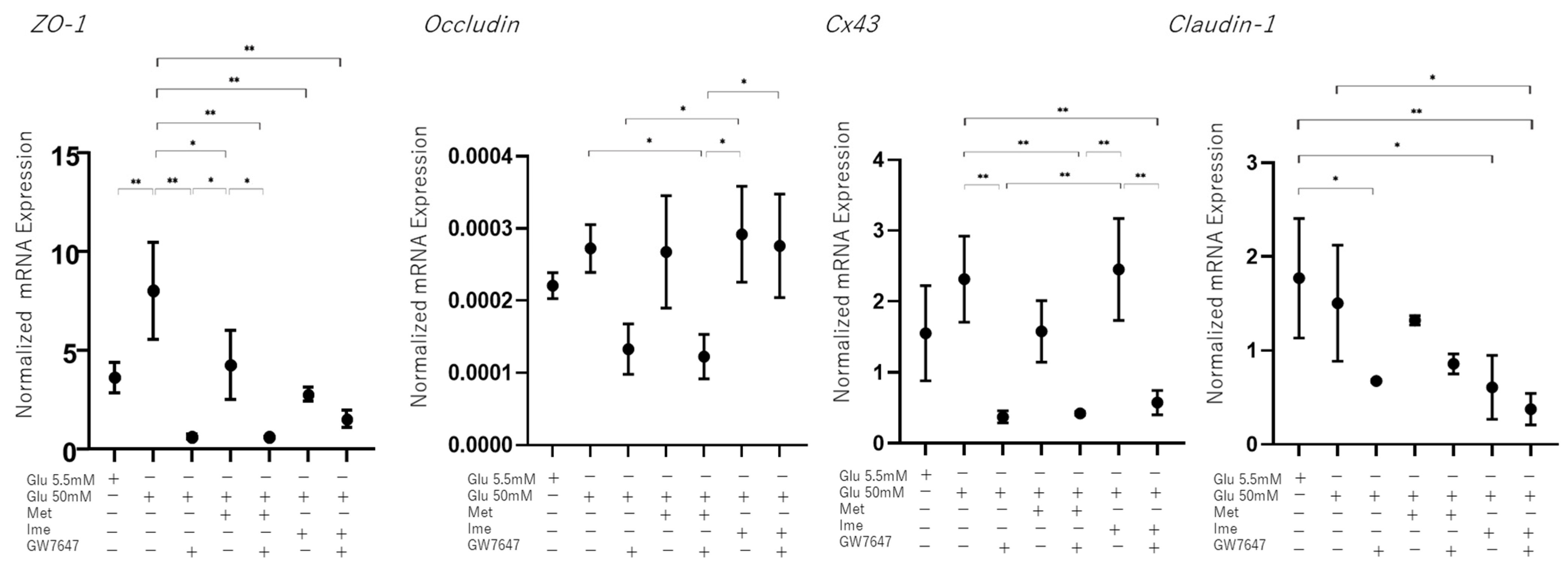
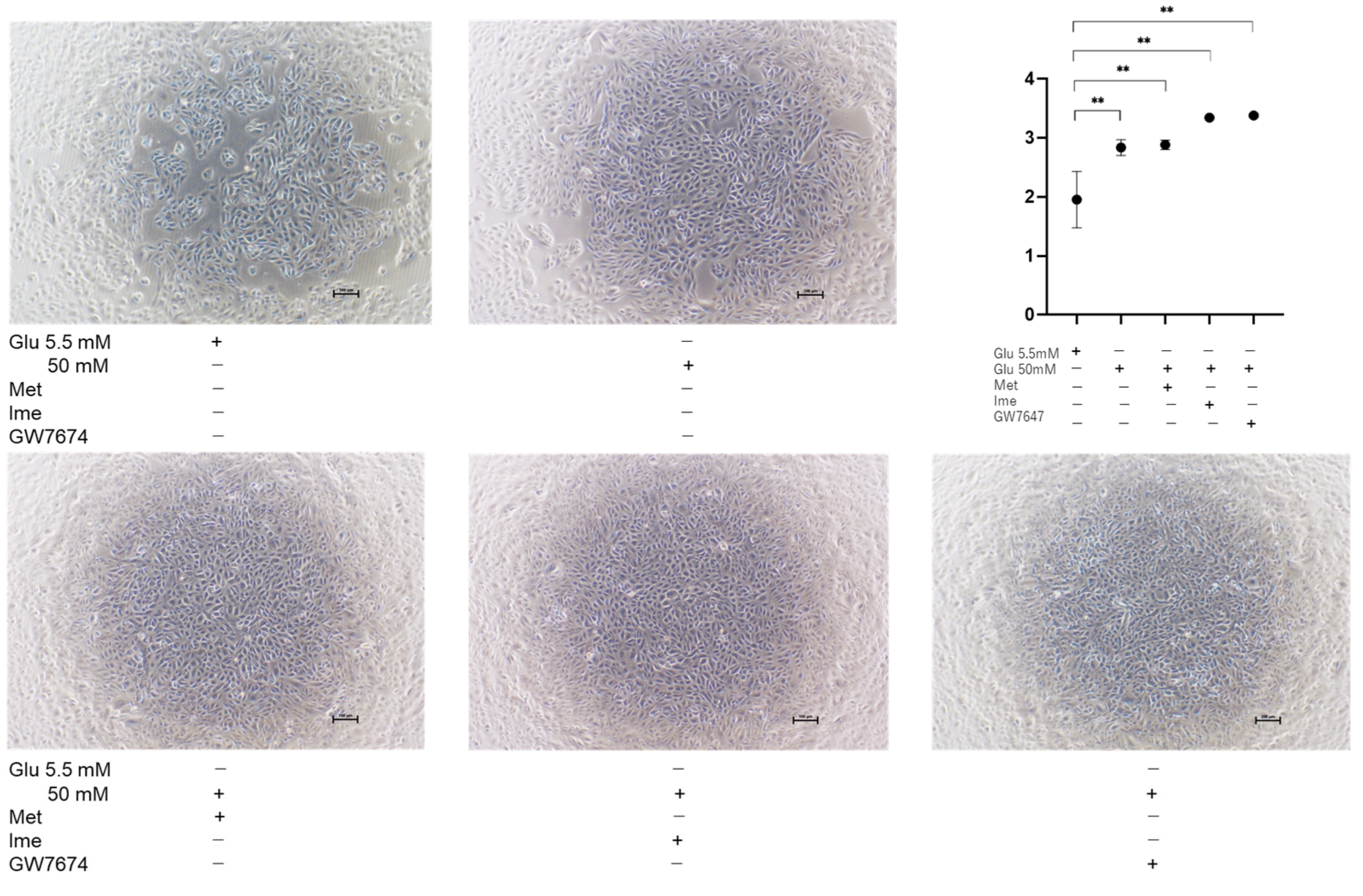
3.3. Effects of Met, Ime, and/or GW7647 on Transendothelial Electrical Resistance (TEER) Values of 2D ARPE19 Cell Monolayers Under Low- and High-Glucose Conditions
3.4. Effects of Metformin, Imeglimin, and/or GW7647 on Cellular Metabolic Functions in ARPE19 Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lightman, S.; Towler, H.M. Diabetic retinopathy. Clin. Cornerstone 2003, 5, 12–21. [Google Scholar] [CrossRef]
- Tong, L.; Vernon, S.A.; Kiel, W.; Sung, V.; Orr, G.M. Association of macular involvement with proliferative retinopathy in Type 2 diabetes. Diabet. Med. A J. Br. Diabet. Assoc. 2001, 18, 388–394. [Google Scholar] [CrossRef] [PubMed]
- Simó, R.; Carrasco, E.; García-Ramírez, M.; Hernández, C. Angiogenic and antiangiogenic factors in proliferative diabetic retinopathy. Curr. Diabetes Rev. 2006, 2, 71–98. [Google Scholar] [CrossRef] [PubMed]
- Joussen, A.M.; Smyth, N.; Niessen, C. Pathophysiology of diabetic macular edema. Dev. Ophthalmol. 2007, 39, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Lakkaraju, A.; Umapathy, A.; Tan, L.X.; Daniele, L.; Philp, N.J.; Boesze-Battaglia, K.; Williams, D.S. The cell biology of the retinal pigment epithelium. Prog. Retin. Eye Res. 2020, 78, 100846. [Google Scholar] [CrossRef]
- Philp, N.J.; Ochrietor, J.D.; Rudoy, C.; Muramatsu, T.; Linser, P.J. Loss of MCT1, MCT3, and MCT4 expression in the retinal pigment epithelium and neural retina of the 5A11/basigin-null mouse. Investig. Ophthalmol. Vis. Sci. 2003, 44, 1305–1311. [Google Scholar] [CrossRef]
- Kumagai, A.K.; Glasgow, B.J.; Pardridge, W.M. GLUT1 glucose transporter expression in the diabetic and nondiabetic human eye. Investig. Ophthalmol. Vis. Sci. 1994, 35, 2887–2894. [Google Scholar]
- Saari, J.C. Biochemistry of visual pigment regeneration: The Friedenwald lecture. Investig. Ophthalmol. Vis. Sci. 2000, 41, 337–348. [Google Scholar]
- Xu, H.Z.; Le, Y.Z. Significance of outer blood-retina barrier breakdown in diabetes and ischemia. Investig. Ophthalmol. Vis. Sci. 2011, 52, 2160–2164. [Google Scholar] [CrossRef]
- Damian, I.; Nicoara, S.D. Optical Coherence Tomography Biomarkers of the Outer Blood-Retina Barrier in Patients with Diabetic Macular Oedema. J. Diabetes Res. 2020, 2020, 8880586. [Google Scholar] [CrossRef]
- Zhang, Y.; Xi, X.; Mei, Y.; Zhao, X.; Zhou, L.; Ma, M.; Liu, S.; Zha, X.; Yang, Y. High-glucose induces retinal pigment epithelium mitochondrial pathways of apoptosis and inhibits mitophagy by regulating ROS/PINK1/Parkin signal pathway. Biomed. Pharmacother. 2019, 111, 1315–1325. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Liang, L.; Zhang, S.; Yang, J.; Yue, Y.; Zhang, X. HMGB1 downregulation in retinal pigment epithelial cells protects against diabetic retinopathy through the autophagy-lysosome pathway. Autophagy 2022, 18, 320–339. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Li, X.; Zhang, D.; Han, W. Astragaloside-IV alleviates high glucose-induced ferroptosis in retinal pigment epithelial cells by disrupting the expression of miR-138-5p/Sirt1/Nrf2. Bioengineered 2022, 13, 8240–8254. [Google Scholar] [CrossRef]
- Zhang, C.S.; Jiang, B.; Li, M.; Zhu, M.; Peng, Y.; Zhang, Y.L.; Wu, Y.Q.; Li, T.Y.; Liang, Y.; Lu, Z.; et al. The lysosomal v-ATPase-Ragulator complex is a common activator for AMPK and mTORC1, acting as a switch between catabolism and anabolism. Cell Metab. 2014, 20, 526–540. [Google Scholar] [CrossRef]
- Jaul, E.; Barron, J. Age-Related Diseases and Clinical and Public Health Implications for the 85 Years Old and Over Population. Front. Public Health 2017, 5, 335. [Google Scholar] [CrossRef] [PubMed]
- Foretz, M.; Guigas, B.; Bertrand, L.; Pollak, M.; Viollet, B. Metformin: From mechanisms of action to therapies. Cell Metab. 2014, 20, 953–966. [Google Scholar] [CrossRef]
- Dutta, S.; Shah, R.B.; Singhal, S.; Dutta, S.B.; Bansal, S.; Sinha, S.; Haque, M. Metformin: A Review of Potential Mechanism and Therapeutic Utility Beyond Diabetes. Drug Des. Dev. Ther. 2023, 17, 1907–1932. [Google Scholar] [CrossRef]
- Hsu, S.K.; Cheng, K.C.; Mgbeahuruike, M.O.; Lin, Y.H.; Wu, C.Y.; Wang, H.D.; Yen, C.H.; Chiu, C.C.; Sheu, S.J. New Insight into the Effects of Metformin on Diabetic Retinopathy, Aging and Cancer: Nonapoptotic Cell Death, Immunosuppression, and Effects beyond the AMPK Pathway. Int. J. Mol. Sci. 2021, 22, 9453. [Google Scholar] [CrossRef]
- Xu, L.; Kong, L.; Wang, J.; Ash, J.D. Stimulation of AMPK prevents degeneration of photoreceptors and the retinal pigment epithelium. Proc. Natl. Acad. Sci. USA 2018, 115, 10475–10480. [Google Scholar] [CrossRef]
- Li, S.; Jin, E.; Shi, X.; Cai, Y.; Zhang, H.; Zhao, M. Proteomics of Vitreous Humor Reveals PPARA, RXR, and LXR Are Possible Upstream Regulators of Proliferative Diabetic Retinopathy. Front. Med. 2021, 8, 724695. [Google Scholar] [CrossRef]
- Tajnšek, Š.; Petrovič, D.; Globočnik Petrovič, M.; Kunej, T. Association of Peroxisome Proliferator-Activated Receptors (PPARs) with Diabetic Retinopathy in Human and Animal Models: Analysis of the Literature and Genome Browsers. PPAR Res. 2020, 2020, 1783564. [Google Scholar] [CrossRef] [PubMed]
- Alnuaimi, S.; Reljic, T.; Abdulla, F.S.; Memon, H.; Al-Ali, S.; Smith, T.; Serdarevic, F.; Velija Asimi, Z.; Kumar, A.; Semiz, S. PPAR agonists as add-on treatment with metformin in management of type 2 diabetes: A systematic review and meta-analysis. Sci. Rep. 2024, 14, 8809. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.Y.; Yan, Q.; Wu, W.H.; Zhao, Y.; Zhang, H.; Li, J. PPAR-alpha/gamma agonists, glucagon-like peptide-1 receptor agonists and metformin for non-alcoholic fatty liver disease: A network meta-analysis. J. Int. Med. Res. 2023, 51, 3000605231177191. [Google Scholar] [CrossRef] [PubMed]
- Okishio, S.; Yamaguchi, K.; Ishiba, H.; Tochiki, N.; Yano, K.; Takahashi, A.; Kataoka, S.; Okuda, K.; Seko, Y.; Liu, Y.; et al. PPARα agonist and metformin co-treatment ameliorates NASH in mice induced by a choline-deficient, amino acid-defined diet with 45% fat. Sci. Rep. 2020, 10, 19578. [Google Scholar] [CrossRef]
- Vuylsteke, V.; Chastain, L.M.; Maggu, G.A.; Brown, C. Imeglimin: A Potential New Multi-Target Drug for Type 2 Diabetes. Drugs RD 2015, 15, 227–232. [Google Scholar] [CrossRef]
- Konkwo, C.; Perry, R.J. Imeglimin: Current Development and Future Potential in Type 2 Diabetes. Drugs 2021, 81, 185–190. [Google Scholar] [CrossRef]
- Pirags, V.; Lebovitz, H.; Fouqueray, P. Imeglimin, a novel glimin oral antidiabetic, exhibits a good efficacy and safety profile in type 2 diabetic patients. Diabetes Obes. Metab. 2012, 14, 852–858. [Google Scholar] [CrossRef]
- Fouqueray, P.; Pirags, V.; Diamant, M.; Schernthaner, G.; Lebovitz, H.E.; Inzucchi, S.E.; Bailey, C.J. The efficacy and safety of imeglimin as add-on therapy in patients with type 2 diabetes inadequately controlled with sitagliptin monotherapy. Diabetes Care 2014, 37, 1924–1930. [Google Scholar] [CrossRef]
- Fouqueray, P.; Pirags, V.; Inzucchi, S.E.; Bailey, C.J.; Schernthaner, G.; Diamant, M.; Lebovitz, H.E. The efficacy and safety of imeglimin as add-on therapy in patients with type 2 diabetes inadequately controlled with metformin monotherapy. Diabetes Care 2013, 36, 565–568. [Google Scholar] [CrossRef]
- Lachaux, M.; Soulié, M.; Hamzaoui, M.; Bailly, A.; Nicol, L.; Rémy-Jouet, I.; Renet, S.; Vendeville, C.; Gluais-Dagorn, P.; Hallakou-Bozec, S.; et al. Short-and long-term administration of imeglimin counters cardiorenal dysfunction in a rat model of metabolic syndrome. Endocrinol. Diabetes Metab. 2020, 3, e00128. [Google Scholar] [CrossRef]
- Aoyagi, K.; Nishiwaki, C.; Nakamichi, Y.; Yamashita, S.I.; Kanki, T.; Ohara-Imaizumi, M. Imeglimin mitigates the accumulation of dysfunctional mitochondria to restore insulin secretion and suppress apoptosis of pancreatic β-cells from db/db mice. Sci. Rep. 2024, 14, 6178. [Google Scholar] [CrossRef] [PubMed]
- Nishikiori, N.; Watanabe, M.; Sato, T.; Umetsu, A.; Higashide, M.; Furuhashi, M.; Ohguro, H. Intraocular fatty acids induce reinforcement of barrier functions on the outer blood-retinal barrier. Prostaglandins Leukot. Essent. Fat. Acids 2024, 202, 102637. [Google Scholar] [CrossRef]
- Higashide, M.; Watanabe, M.; Sato, T.; Ogawa, T.; Umetsu, A.; Suzuki, S.; Furuhashi, M.; Ohguro, H.; Nishikiori, N. Unexpected and Synergistical Effects of All-Trans Retinoic Acid and TGF-β2 on Biological Aspects of 2D and 3D Cultured ARPE19 Cells. Biomedicines 2024, 12, 2228. [Google Scholar] [CrossRef] [PubMed]
- Oouchi, Y.; Watanabe, M.; Ida, Y.; Ohguro, H.; Hikage, F. Rosiglitasone and ROCK Inhibitors Modulate Fibrogenetic Changes in TGF-β2 Treated Human Conjunctival Fibroblasts (HconF) in Different Manners. Int. J. Mol. Sci. 2021, 22, 7335. [Google Scholar] [CrossRef]
- Kaneko, Y.; Ohta, M.; Inoue, T.; Mizuno, K.; Isobe, T.; Tanabe, S.; Tanihara, H. Effects of K-115 (Ripasudil), a novel ROCK inhibitor, on trabecular meshwork and Schlemm’s canal endothelial cells. Sci. Rep. 2016, 6, 19640. [Google Scholar] [CrossRef] [PubMed]
- Ida, Y.; Hikage, F.; Itoh, K.; Ida, H.; Ohguro, H. Prostaglandin F2α agonist-induced suppression of 3T3-L1 cell adipogenesis affects spatial formation of extra-cellular matrix. Sci. Rep. 2020, 10, 7958. [Google Scholar] [CrossRef]
- Itoh, K.; Hikage, F.; Ida, Y.; Ohguro, H. Prostaglandin F2α Agonists Negatively Modulate the Size of 3D Organoids from Primary Human Orbital Fibroblasts. Investig. Ophthalmol. Vis. Sci. 2020, 61, 13. [Google Scholar] [CrossRef]
- Lee, H.Y.; Gao, X.; Barrasa, M.I.; Li, H.; Elmes, R.R.; Peters, L.L.; Lodish, H.F. PPAR-α and glucocorticoid receptor synergize to promote erythroid progenitor self-renewal. Nature 2015, 522, 474–477. [Google Scholar] [CrossRef]
- Lin, Y.; Berg, A.H.; Iyengar, P.; Lam, T.K.; Giacca, A.; Combs, T.P.; Rajala, M.W.; Du, X.; Rollman, B.; Li, W.; et al. The hyperglycemia-induced inflammatory response in adipocytes: The role of reactive oxygen species. J. Biol. Chem. 2005, 280, 4617–4626. [Google Scholar] [CrossRef]
- Swanson, M. Retinopathy screening in individuals with type 2 diabetes: Who, how, how often, and at what cost--an epidemiologic review. Optometry 2005, 76, 636–646. [Google Scholar] [CrossRef]
- Madsen-Bouterse, S.A.; Mohammad, G.; Kanwar, M.; Kowluru, R.A. Role of mitochondrial DNA damage in the development of diabetic retinopathy, and the metabolic memory phenomenon associated with its progression. Antioxid. Redox Signal. 2010, 13, 797–805. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Chen, H.; Wang, H.; Ke, B.; Zheng, B.; Li, Q.; Li, P.; Su, L.; Gu, Q.; Xu, X. Improvement of retinal vascular injury in diabetic rats by statins is associated with the inhibition of mitochondrial reactive oxygen species pathway mediated by peroxisome proliferator-activated receptor gamma coactivator 1alpha. Diabetes 2010, 59, 2315–2325. [Google Scholar] [CrossRef] [PubMed]
- Hozumi, K.; Sugawara, K.; Ishihara, T.; Ishihara, N.; Ogawa, W. Effects of imeglimin on mitochondrial function, AMPK activity, and gene expression in hepatocytes. Sci. Rep. 2023, 13, 746. [Google Scholar] [CrossRef] [PubMed]
- Vial, G.; Lamarche, F.; Cottet-Rousselle, C.; Hallakou-Bozec, S.; Borel, A.L.; Fontaine, E. The mechanism by which imeglimin inhibits gluconeogenesis in rat liver cells. Endocrinol. Diabetes Metab. 2021, 4, e00211. [Google Scholar] [CrossRef]
- Clémence, C.; Fouqueray, P.; Sébastien, B. In Vitro Investigation, Pharmacokinetics, and Disposition of Imeglimin, a Novel Oral Antidiabetic Drug, in Preclinical Species and Humans. Drug Metab. Dispos. Biol. Fate Chem. 2020, 48, 1330–1346. [Google Scholar] [CrossRef]
- Bustamante-Barrientos, F.A.; Luque-Campos, N.; Araya, M.J.; Lara-Barba, E.; de Solminihac, J.; Pradenas, C.; Molina, L.; Herrera-Luna, Y.; Utreras-Mendoza, Y.; Elizondo-Vega, R.; et al. Mitochondrial dysfunction in neurodegenerative disorders: Potential therapeutic application of mitochondrial transfer to central nervous system-residing cells. J. Transl. Med. 2023, 21, 613. [Google Scholar] [CrossRef]
- Detaille, D.; Vial, G.; Borel, A.L.; Cottet-Rouselle, C.; Hallakou-Bozec, S.; Bolze, S.; Fouqueray, P.; Fontaine, E. Imeglimin prevents human endothelial cell death by inhibiting mitochondrial permeability transition without inhibiting mitochondrial respiration. Cell Death Discov. 2016, 2, 15072. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nishikiori, N.; Watanabe, M.; Higashide, M.; Umetsu, A.; Ogawa, T.; Furuhashi, M.; Ohguro, H.; Sato, T. The Combination of PPARα Agonist GW7647 and Imeglimin Has Potent Effects on High-Glucose-Induced Cellular Biological Responses in Human Retinal Pigment Epithelium Cells. Bioengineering 2025, 12, 265. https://doi.org/10.3390/bioengineering12030265
Nishikiori N, Watanabe M, Higashide M, Umetsu A, Ogawa T, Furuhashi M, Ohguro H, Sato T. The Combination of PPARα Agonist GW7647 and Imeglimin Has Potent Effects on High-Glucose-Induced Cellular Biological Responses in Human Retinal Pigment Epithelium Cells. Bioengineering. 2025; 12(3):265. https://doi.org/10.3390/bioengineering12030265
Chicago/Turabian StyleNishikiori, Nami, Megumi Watanabe, Megumi Higashide, Araya Umetsu, Toshifumi Ogawa, Masato Furuhashi, Hiroshi Ohguro, and Tatsuya Sato. 2025. "The Combination of PPARα Agonist GW7647 and Imeglimin Has Potent Effects on High-Glucose-Induced Cellular Biological Responses in Human Retinal Pigment Epithelium Cells" Bioengineering 12, no. 3: 265. https://doi.org/10.3390/bioengineering12030265
APA StyleNishikiori, N., Watanabe, M., Higashide, M., Umetsu, A., Ogawa, T., Furuhashi, M., Ohguro, H., & Sato, T. (2025). The Combination of PPARα Agonist GW7647 and Imeglimin Has Potent Effects on High-Glucose-Induced Cellular Biological Responses in Human Retinal Pigment Epithelium Cells. Bioengineering, 12(3), 265. https://doi.org/10.3390/bioengineering12030265