
Reversing the Natural Drug Resistance of Gram-Negative Bacteria to Fusidic Acid via Forming Drug–Phospholipid Complex

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
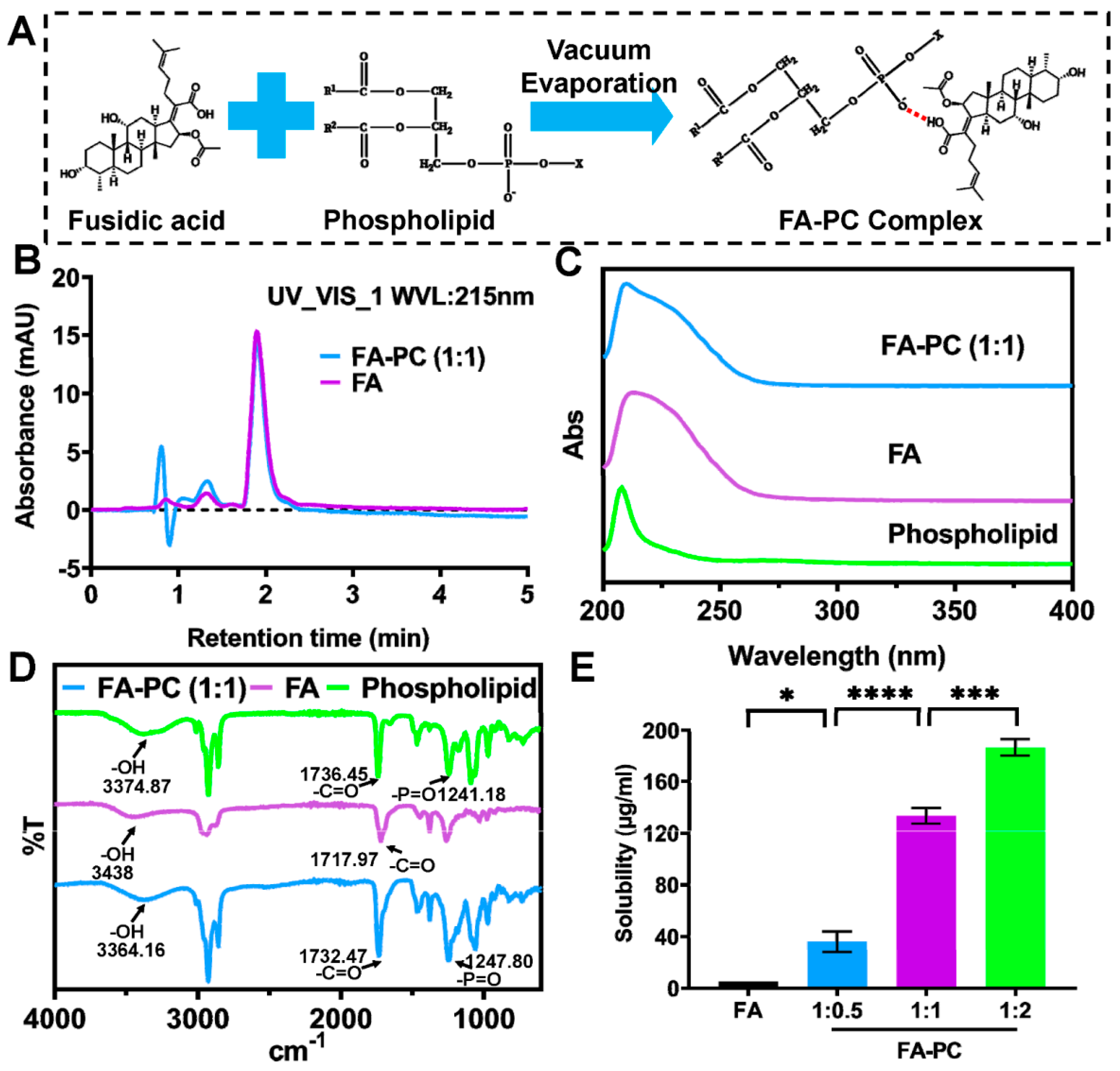
2.2. Preparation of FA-PC
2.3. Characterization of FA-PC
2.3.1. High-Performance Liquid Chromatography (HPLC)
2.3.2. The UV Spectroscopy
2.3.3. The FT-IR Spectroscopy
2.4. Solubility
2.5. Bacterial Culture
2.6. Antibacterial Activity of FA-PC
2.7. Scanning Electron Microscopy (SEM)
2.8. Fluorescence Analysis of Intracellular Drug–Phospholipid Complex
2.9. Competition Inhibition of FA-PC Delivery by Extra Phospholipid
2.10. Statistics
3. Results and Discussion
3.1. Preparation and Characterization of FA-PC
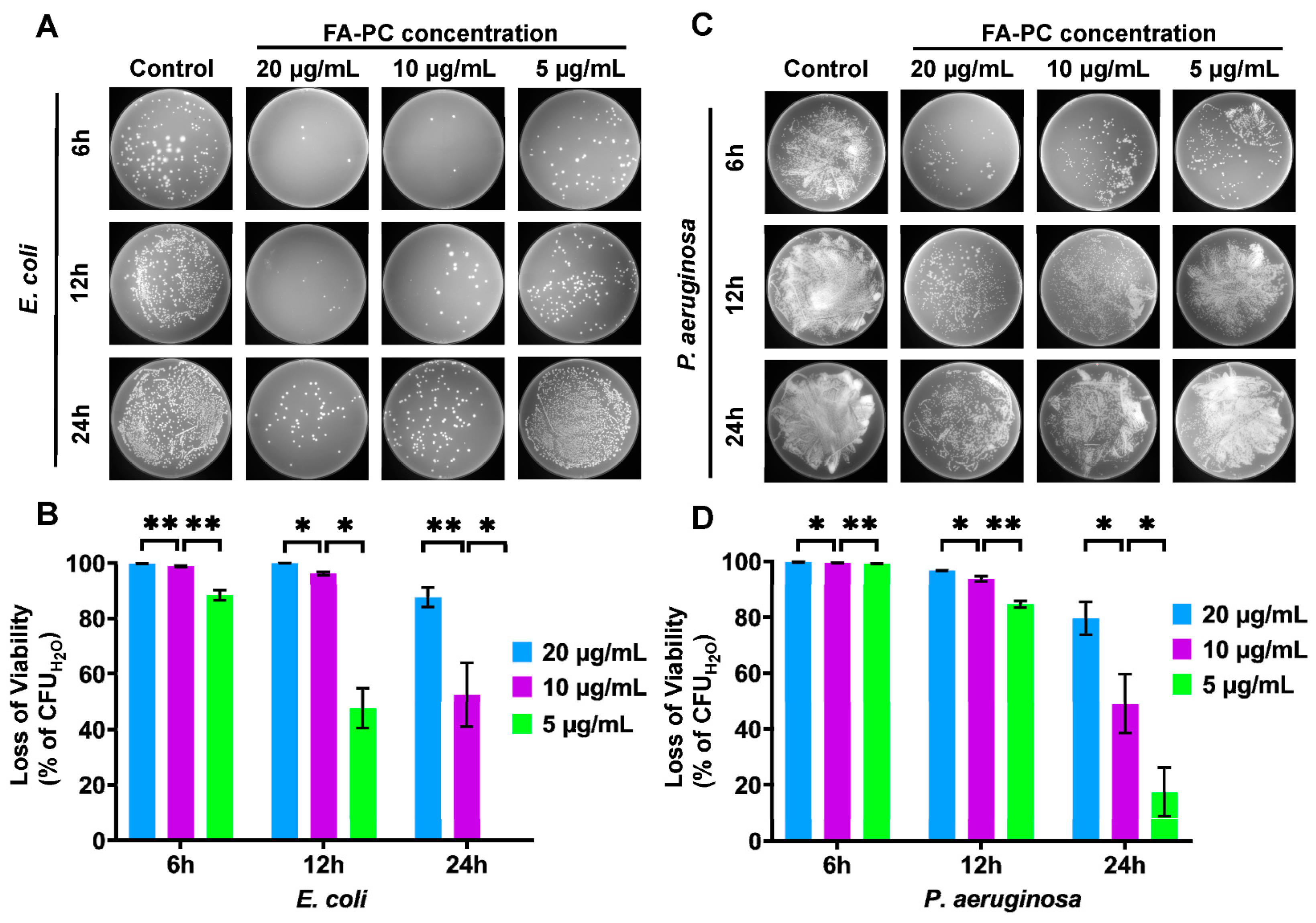
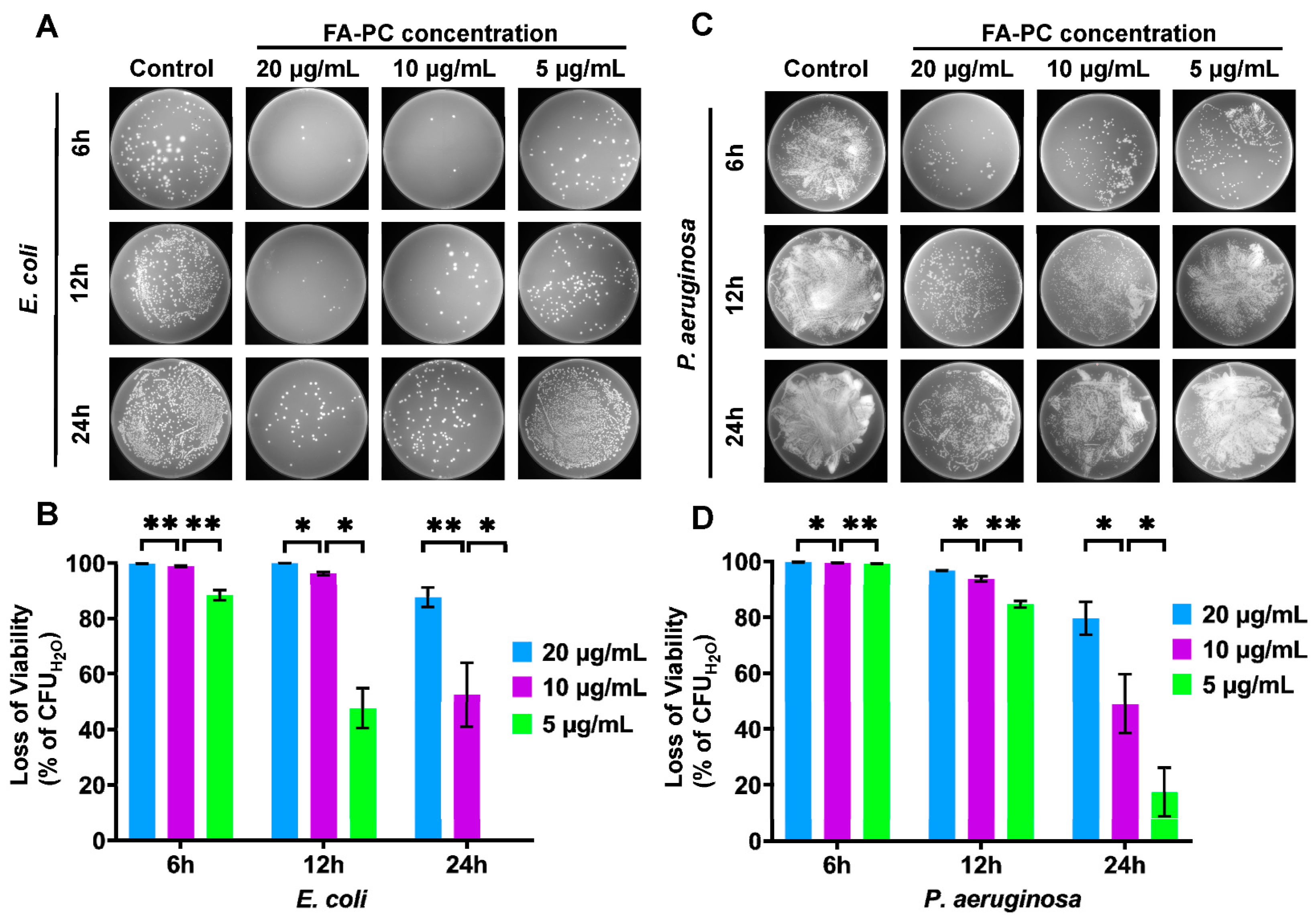
3.2. Antibacterial Activity of FA-PC
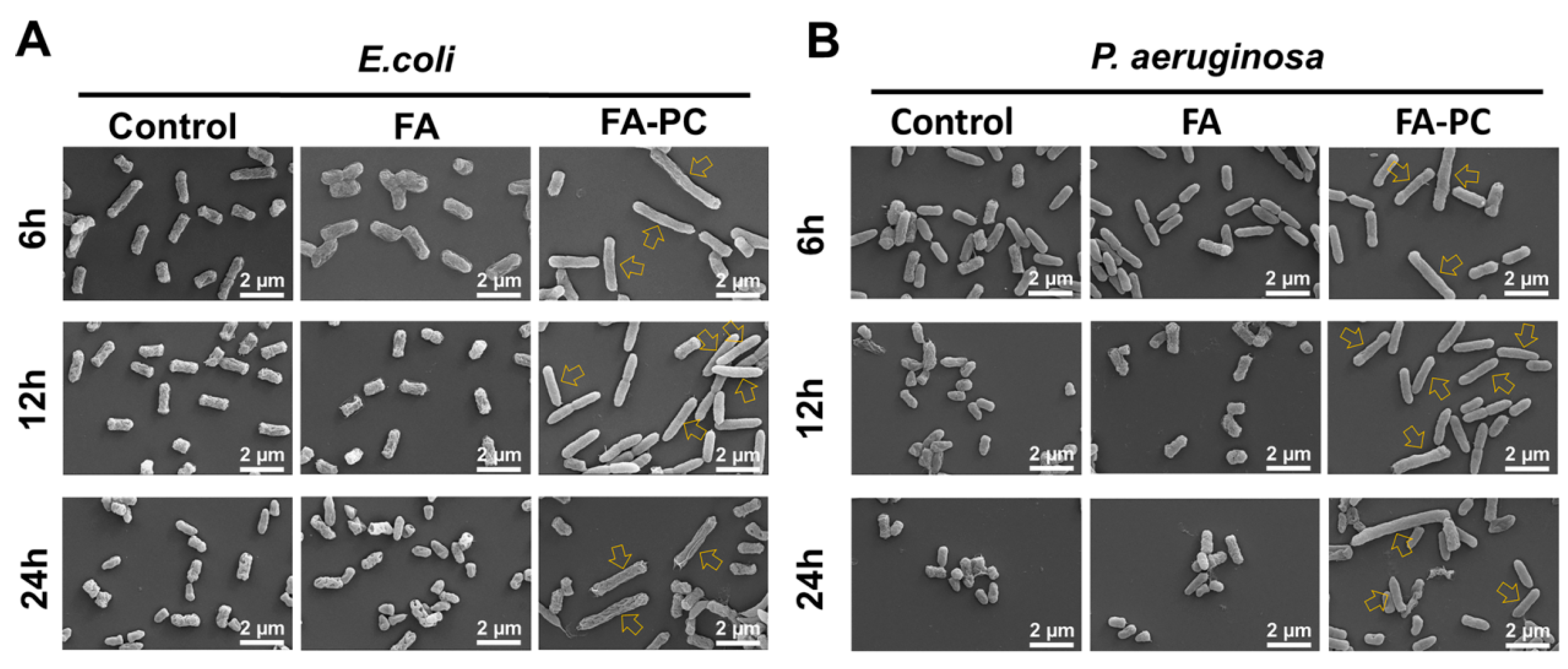
3.3. Morphology Changes in Bacteria Cells
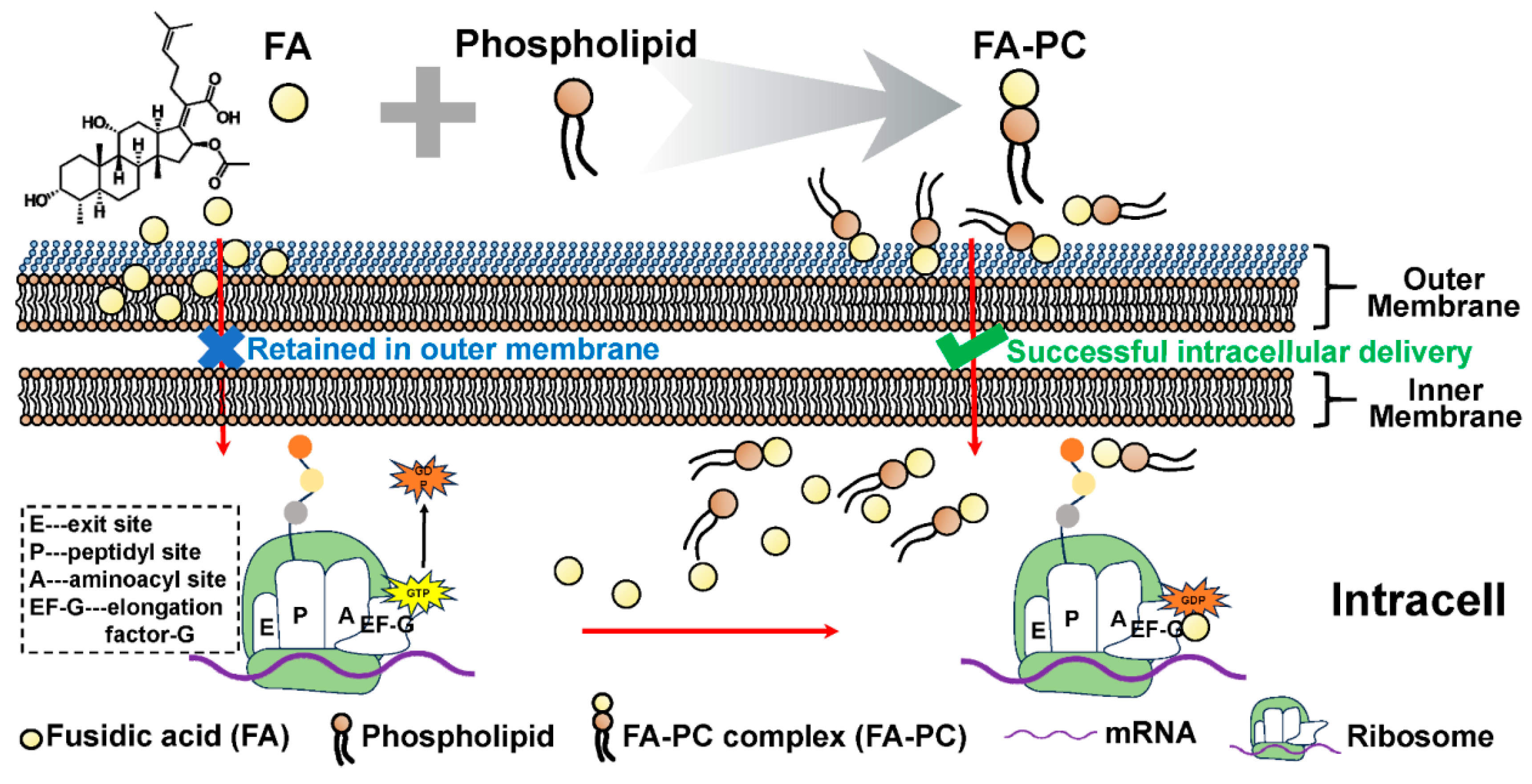
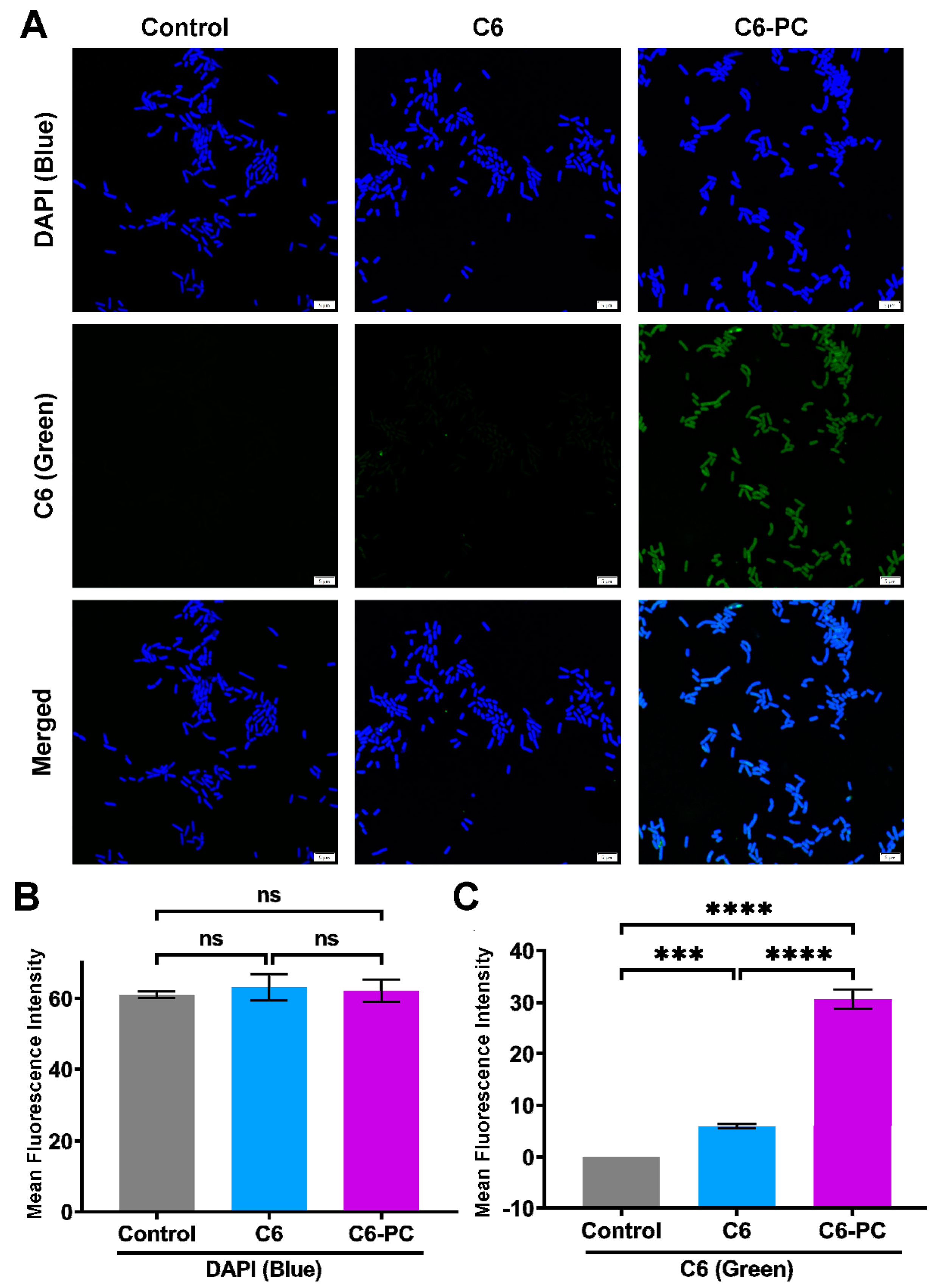
3.4. Intracellular Delivery of Drug–Phospholipid Complex
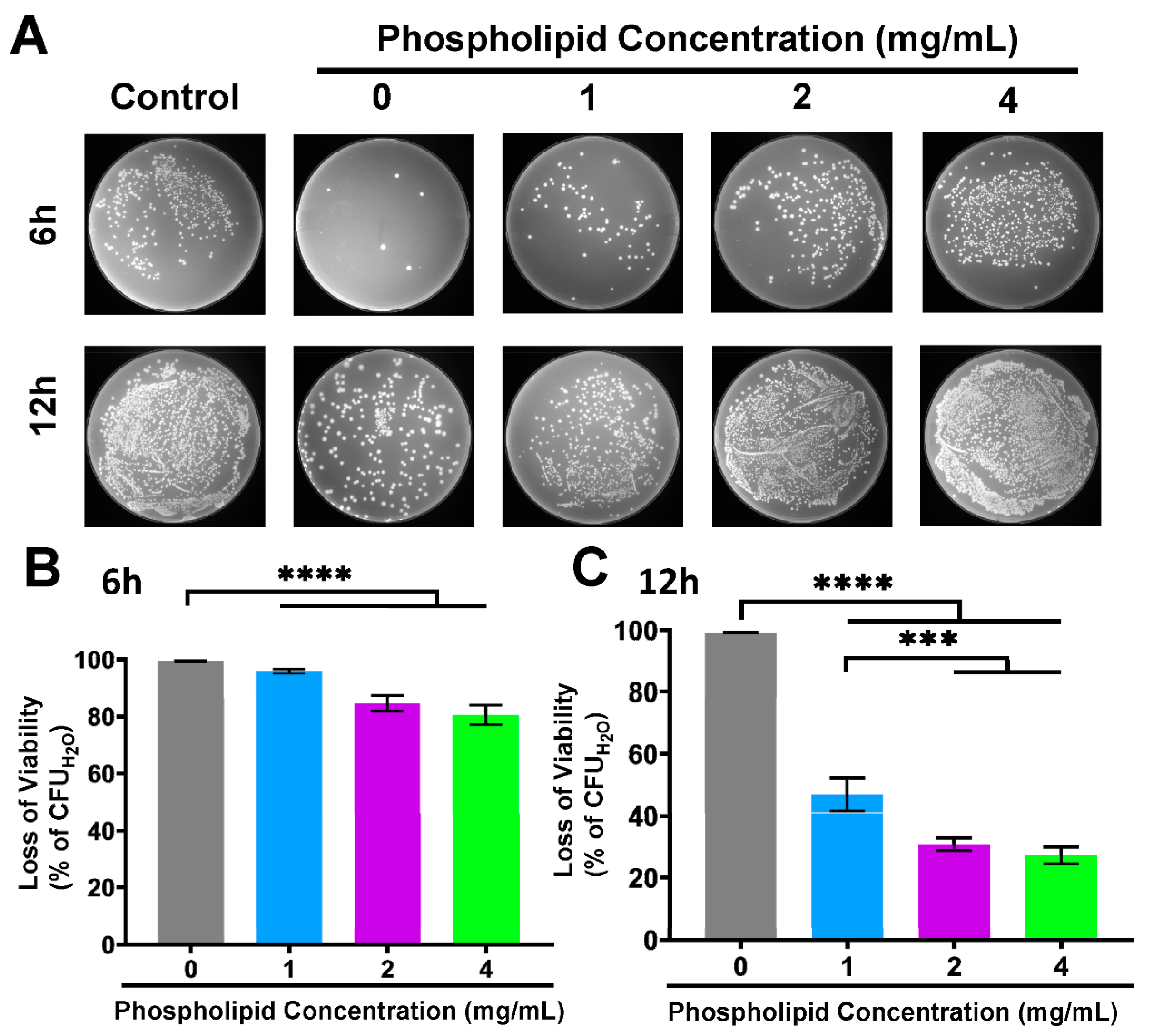
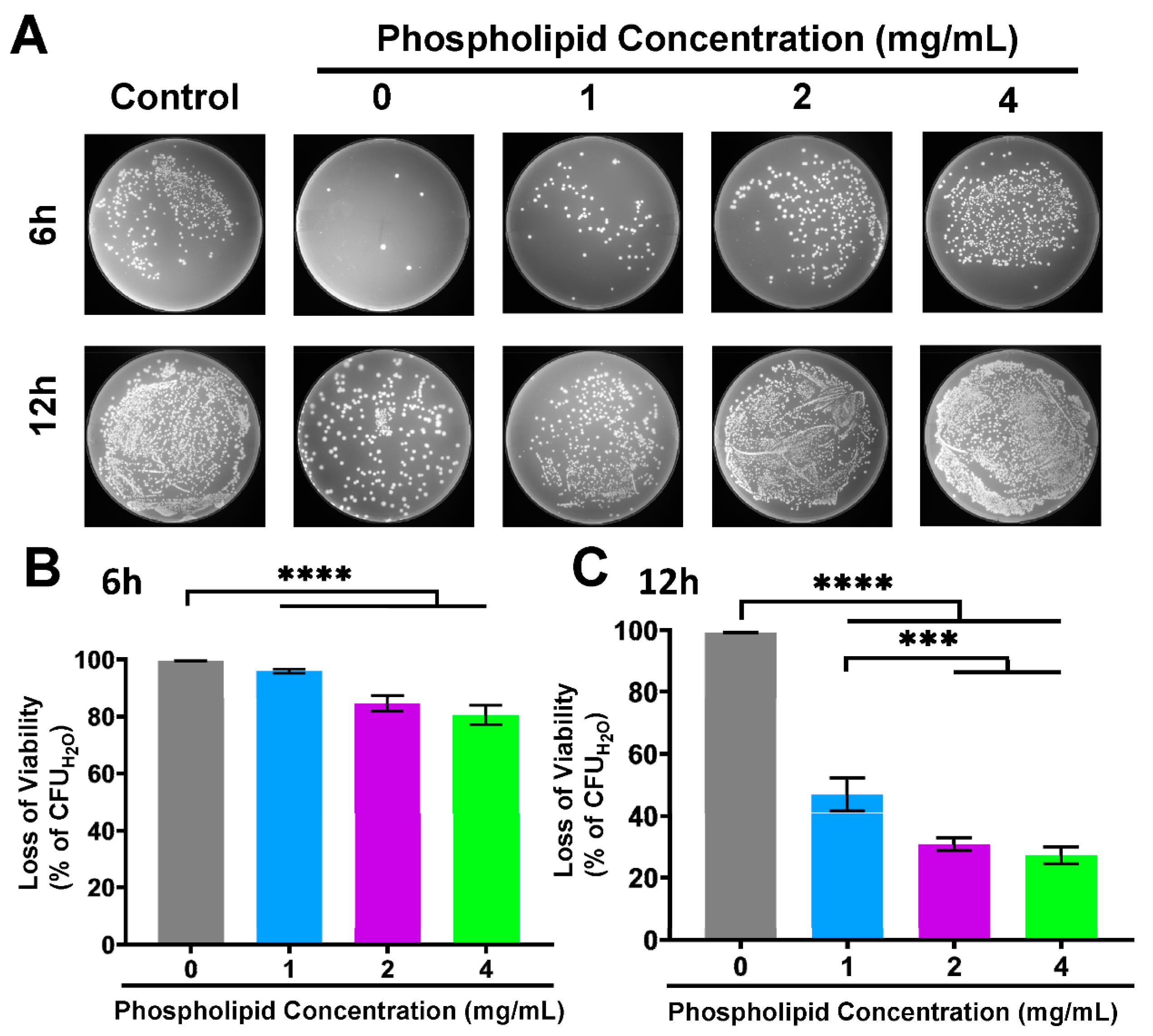
3.5. Compromised Antibacterial Activity of FA-PC by Extra Phospholipid
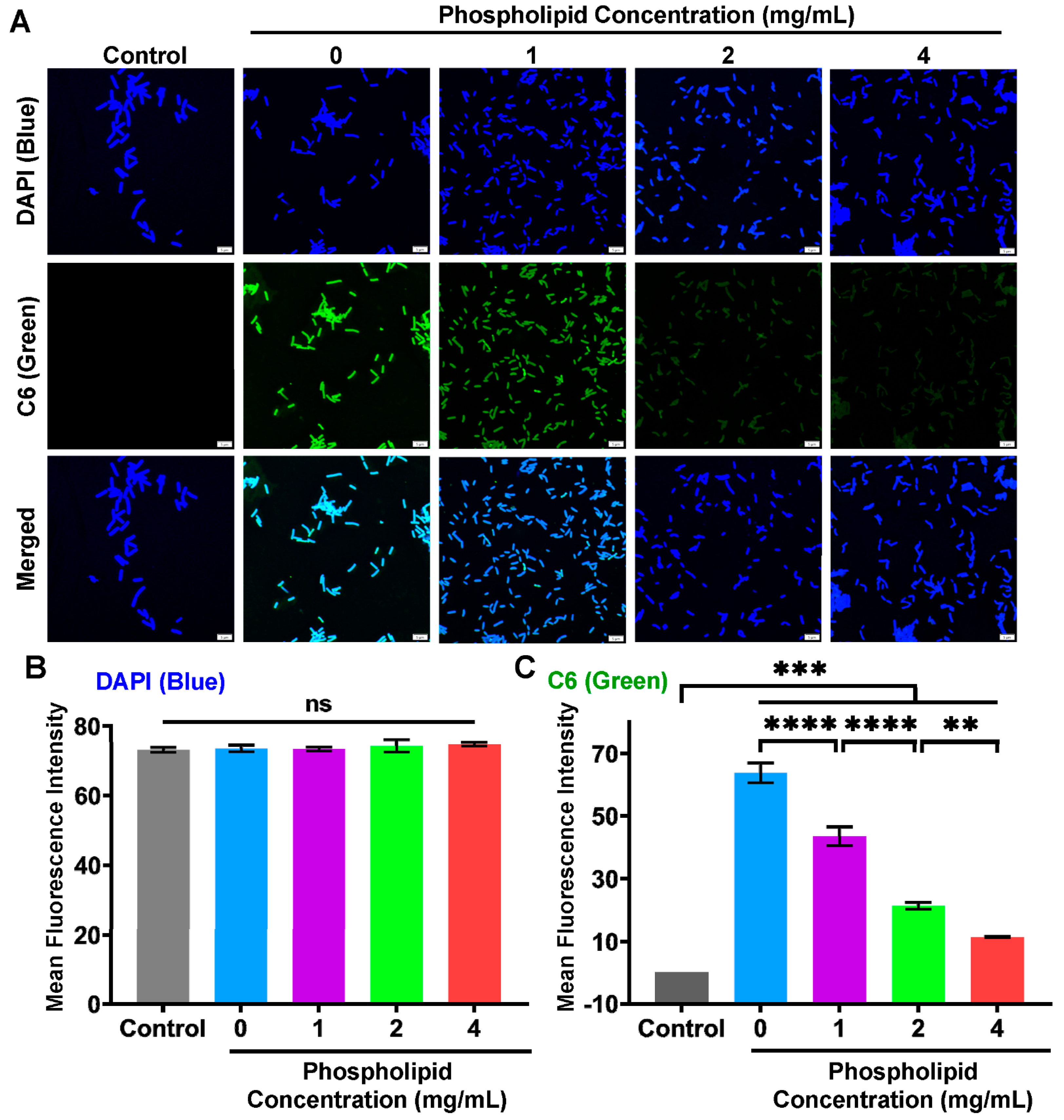
3.6. Inhibited Intracellular Delivery of Drug–Phospholipid Complex by Extra Phospholipid
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gao, Y.; Dong, Y.; Yang, S.; Mo, A.; Zeng, X.; Chen, Q.; Peng, Q. Size-dependent photothermal antibacterial activity of Ti3C2Tx MXene nanosheets against methicillin-resistant Staphylococcus aureus. J. Colloid Interface Sci. 2022, 617, 533–541. [Google Scholar] [CrossRef]
- Tan, Y.; Su, J.; Fu, M.; Zhang, H.; Zeng, H. Recent Advances in Phage-Based Therapeutics for Multi-Drug Resistant Acinetobacter baumannii. Bioengineering 2023, 10, 35. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Meng, L.; Chen, Y.; Dong, Z.; Peng, Q. Bacterial outer membrane vesicles as potential biological nanomaterials for antibacterial therapy. Acta Biomater. 2022, 140, 102–115. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, D.; Liu, F.; Sheng, S.; Zhang, H.; Li, W.; Li, Y.; Tian, H. Enhancing the drug sensitivity of antibiotics on drug-resistant bacteria via the photothermal effect of FeTGNPs. J. Control Release 2022, 341, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Li, Y.; Liu, Y.; Xu, Z.; Wen, M.; Zhang, L.; Xue, Y.; Shang, L. Highly biocompatible Ag nanocluster-reinforced wound dressing with long-term and synergistic bactericidal activity. J. Colloid Interface Sci. 2023, 633, 851–865. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Sun, X.; Liu, W.; Li, H.; Wang, J.; Wang, D. Amino acid-mediated amorphous copper sulphide with enhanced photothermal conversion efficiency for antibacterial application. J. Colloid Interface Sci. 2024, 657, 142–154. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Sui, S.; Yu, X.; Huang, W.; Wu, Y.; Zeng, X.; Chen, Q.; Wang, J.; Peng, Q. Ti3C2Tx MXene loaded with indocyanine green for synergistic photothermal and photodynamic therapy for drug-resistant bacterium. Colloids Surf. B Biointerfaces 2022, 217, 112663. [Google Scholar] [CrossRef] [PubMed]
- Ni, J.; Guo, M.; Cao, Y.; Lei, L.; Liu, K.; Wang, B.; Lu, F.; Zhai, R.; Gao, X.; Yan, C.; et al. Discovery, synthesis of novel fusidic acid derivatives possessed amino-terminal groups at the 3-hydroxyl position with anticancer activity. Eur. J. Med. Chem. 2019, 162, 122–131. [Google Scholar] [CrossRef] [PubMed]
- Long, J.; Ying, T.; Zhang, L.; Yu, T.; Wu, J.; Liu, Y.; Li, X.; You, G.; Zhang, L.; Bi, Y. Discovery of fusidic acid derivatives as novel STING inhibitors for treatment of sepsis. Eur. J. Med. Chem. 2022, 244, 114814. [Google Scholar] [CrossRef]
- Marian, E.; Tita, B.; Duteanu, N.; Vicas, L.; Ciocan, S.; Jurca, T.; Antal, L.; Tica, O.; Mureşan, M.; Pallag, A.; et al. Antimicrobial activity of fusidic acid inclusion complexes. Int. J. Infect. Dis. 2020, 101, 65–73. [Google Scholar] [CrossRef]
- Chavez, M.G.; Garcia, A.; Lee, H.Y.; Lau, G.W.; Parker, E.N.; Komnick, K.E.; Hergenrother, P.J. Synthesis of Fusidic Acid Derivatives Yields a Potent Antibiotic with an Improved Resistance Profile. ACS Infect. Dis. 2021, 7, 493–505. [Google Scholar] [CrossRef]
- Long, J.; Ji, W.; Zhang, D.; Zhu, Y.; Bi, Y. Bioactivities and Structure–Activity Relationships of Fusidic Acid Derivatives: A Review. Front. Pharm. 2021, 12, 759220. [Google Scholar] [CrossRef] [PubMed]
- Romaru, J.; Limelette, A.; Lebrun, D.; Bonnet, M.; Garnier, V.V.; N’Guyen, Y. Fusidic acid in a tertiary hospital: An observational study focusing on prescriptions, tolerance and susceptibility of Staphylococcus and Cutibacterium spp. strains from bone samples. Eur. J. Clin. Microbiol. Infect. Dis. 2022, 41, 1107–1113. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.; Dziwornu, G.A.; Mabhula, A.; Chibale, K. Rv0684/fusA1, an Essential Gene, Is the Target of Fusidic Acid and Its Derivatives in Mycobacterium tuberculosis. ACS Infect. Dis. 2021, 7, 2437–2444. [Google Scholar] [CrossRef] [PubMed]
- Borg, A.; Pavlov, M.; Ehrenberg, M. Mechanism of fusidic acid inhibition of RRF- and EF-G-dependent splitting of the bacterial post-termination ribosome. Nucleic Acids Res. 2016, 44, 3264–3275. [Google Scholar] [CrossRef] [PubMed]
- Ayyub, S.A.; Lahry, K.; Dobriyal, D.; Mondal, S.; Varshney, U. Antimicrobial activity of fusidic acid in Escherichia coli is dependent on the relative levels of ribosome recycling factor and elongation factor G. FEMS Microbiol. Lett. 2018, 365, fny133. [Google Scholar] [CrossRef] [PubMed]
- Borg, A.; Holm, M.; Shiroyama, I.; Hauryliuk, V.; Pavlov, M.; Sanyal, S.; Ehrenberg, M. Fusidic acid targets elongation factor G in several stages of translocation on the bacterial ribosome. J. Biol. Chem. 2015, 290, 3440–3454. [Google Scholar] [CrossRef] [PubMed]
- Belardinelli, R.; Rodnina, M.V. Effect of Fusidic Acid on the Kinetics of Molecular Motions During EF-G-Induced Translocation on the Ribosome. Sci. Rep. 2017, 7, 10536. [Google Scholar] [CrossRef]
- Biedenbach, D.J.; Rhomberg, P.R.; Mendes, R.E.; Jones, R.N. Spectrum of activity, mutation rates, synergistic interactions, and the effects of pH and serum proteins for fusidic acid (CEM-102). Diagn. Microbiol. Infect. Dis. 2010, 66, 301–307. [Google Scholar] [CrossRef]
- Nicolosi, D.; Cupri, S.; Genovese, C.; Tempera, G.; Mattina, R.; Pignatello, R. Nanotechnology approaches for antibacterial drug delivery: Preparation and microbiological evaluation of fusogenic liposomes carrying fusidic acid. Int. J. Antimicrob. Agents 2015, 45, 622–626. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, P.; Wu, C.; Wang, J.; Zhou, Y.; Zhang, J.; Wang, B.; Zhao, H.; Rao, L.; Li, M.; et al. Polymyxin B and fusidic acid, a novel potent synergistic combination against Klebsiella pneumoniae and Escherichia coli isolates with polymyxin B resistance. Front. Microbiol. 2023, 14, 1220683. [Google Scholar] [CrossRef]
- Helle, A.; Makitalo, J.; Huhtanen, J.; Holopainen, J.M.; Wiedmer, S.K. Antibiotic fusidic acid has strong interactions with negatively charged lipid membranes: An electrokinetic capillary chromatographic study. Biochim. Biophys. Acta 2008, 1778, 2640–2647. [Google Scholar] [CrossRef]
- Falck, E.; Hautala, J.T.; Karttunen, M.; Kinnunen, P.K.; Patra, M.; Saaren-Seppala, H.; Vattulainen, I.; Wiedmer, S.K.; Holopainen, J.M. Interaction of fusidic acid with lipid membranes: Implications to the mechanism of antibiotic activity. Biophys. J. 2006, 91, 1787–1799. [Google Scholar] [CrossRef]
- Gilchrist, S.E.; Rickard, D.L.; Letchford, K.; Needham, D.; Burt, H.M. Phase separation behavior of fusidic acid and rifampicin in PLGA microspheres. Mol. Pharm. 2012, 9, 1489–1501. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Chen, D.; Li, Y.; Chen, Y.; Liu, Q.; Zhou, X.; Qian, K.; Li, Z.; Ruan, H.; Hou, Z.; et al. Zinc phthalocyanine-soybean phospholipid complex based drug carrier for switchable photoacoustic/fluorescence image, multiphase photothermal/photodynamic treatment and synergetic therapy. J. Control Release 2018, 284, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Ebada, H.M.K.; Nasra, M.M.A.; Elnaggar, Y.S.R.; Abdallah, O.Y. Novel rhein-phospholipid complex targeting skin diseases: Development, in vitro, ex vivo, and in vivo studies. Drug Deliv. Transl. Res. 2021, 11, 1107–1118. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Shi, C.; Zhou, X.; Lin, T.; Gong, Y.; Yin, M.; Fan, L.; Wang, W.; Fang, J. Preparation of a nanoscale dihydromyricetin-phospholipid complex to improve the bioavailability: In vitro and in vivo evaluations. Eur. J. Pharm. Sci. 2019, 138, 104994. [Google Scholar] [CrossRef] [PubMed]
- Peng, Q.; Gong, T.; Zuo, J.; Liu, J.; Zhao, D.; Zhang, Z. Enhanced the oral bioavailability of salvianolic acid B by phospholipid complex loaded nanoparticles. Pharmazie 2008, 63, 661–666. [Google Scholar] [PubMed]
- Peng, Q.; Zhang, Z.-R.; Sun, X.; Zuo, J.; Zhao, D.; Gong, T. Mechanisms of Phospholipid Complex Loaded Nanoparticles Enhancing the Oral Bioavailability. Mol. Pharm. 2010, 7, 565–575. [Google Scholar] [CrossRef] [PubMed]
- Peng, Q.; Yang, Y.J.; Zhang, T.; Wu, C.Y.; Yang, Q.; Sun, X.; Gong, T.; Zhang, L.; Zhang, Z.R. The implantable and biodegradable PHBHHx 3D scaffolds loaded with protein-phospholipid complex for sustained delivery of proteins. Pharm. Res. 2013, 30, 1077–1085. [Google Scholar] [CrossRef]
- Peng, Q.; Zhang, Z.R.; Gong, T.; Chen, G.Q.; Sun, X. A rapid-acting, long-acting insulin formulation based on a phospholipid complex loaded PHBHHx nanoparticles. Biomaterials 2012, 33, 1583–1588. [Google Scholar] [CrossRef]
- Li, Y.; Yin, H.; Wu, C.; He, J.; Wang, C.; Ren, B.; Wang, H.; Geng, D.; Zhang, Y.; Zhao, L. Preparation and in vivo evaluation of an intravenous emulsion loaded with an aprepitant-phospholipid complex. Drug Deliv. 2023, 30, 2183834. [Google Scholar] [CrossRef]
- Telange, D.R.; Sohail, N.K.; Hemke, A.T.; Kharkar, P.S.; Pethe, A.M. Phospholipid complex-loaded self-assembled phytosomal soft nanoparticles: Evidence of enhanced solubility, dissolution rate, ex vivo permeability, oral bioavailability, and antioxidant potential of mangiferin. Drug Deliv. Transl. Res. 2021, 11, 1056–1083. [Google Scholar] [CrossRef]
- Dong, Y.; Liu, J.; Chen, Y.; Zhu, T.; Li, Y.; Zhang, C.; Zeng, X.; Chen, Q.; Peng, Q. Photothermal and natural activity-based synergistic antibacterial effects of Ti3C2Tx MXene-loaded chitosan hydrogel against methicillin-resistant Staphylococcus aureus. Int. J. Biol. Macromol. 2023, 240, 124482. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xu, J.; Yu, C.; Zhou, X.; Chang, L.; Liu, J.; Peng, Q. Prevention of bacterial biofilm formation on orthodontic brackets by non-crosslinked chitosan coating. Int. J. Biol. Macromol. 2023, 251, 126283. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Gao, H.; Chang, L.; Xu, J.; Zhou, X.; Zhang, C.; Peng, Q. Efficient Removal of Dental Plaque Biofilm from Training Typodont Teeth via Water Flosser. Bioengineering 2023, 10, 1061. [Google Scholar] [CrossRef] [PubMed]
- Chi, C.; Zhang, C.; Liu, Y.; Nie, H.; Zhou, J.; Ding, Y. Phytosome-nanosuspensions for silybin-phospholipid complex with increased bioavailability and hepatoprotection efficacy. Eur. J. Pharm. Sci. 2020, 144, 105212. [Google Scholar] [CrossRef]
- Chakravarti, R.K.; Kaur, S.; Samal, S.K.; Kashyap, M.C.; Sangamwar, A.T. Combination of Phospholipid Complex and Matrix Dispersion. AAPS PharmSciTech 2021, 22, 189. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Xu, J.; Dong, Y.; Wang, Y.; Yu, C.; Li, Y.; Zhang, C.; Chen, Q.; Chen, S.; Peng, Q. Drug-free and non-crosslinked chitosan/hyaluronic acid hybrid hydrogel for synergistic healing of infected diabetic wounds. Carbohydr. Polym. 2023, 314, 120962. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Fruhauf, A.; Fan, C.; Ponce, J.; Ueberheide, B.; Bhabha, G.; Ekiert, D.C. Structure of an endogenous mycobacterial MCE lipid transporter. Nature 2023, 620, 445–452. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, J.; Lai, X.; Li, Y.; Yu, Z.; Wang, X.; Zhang, C.; Peng, Q. Reversing the Natural Drug Resistance of Gram-Negative Bacteria to Fusidic Acid via Forming Drug–Phospholipid Complex. Bioengineering 2024, 11, 177. https://doi.org/10.3390/bioengineering11020177
Liu J, Lai X, Li Y, Yu Z, Wang X, Zhang C, Peng Q. Reversing the Natural Drug Resistance of Gram-Negative Bacteria to Fusidic Acid via Forming Drug–Phospholipid Complex. Bioengineering. 2024; 11(2):177. https://doi.org/10.3390/bioengineering11020177
Chicago/Turabian StyleLiu, Jianhong, Xuyang Lai, Yuanhong Li, Zhuohang Yu, Xuan Wang, Chaoliang Zhang, and Qiang Peng. 2024. "Reversing the Natural Drug Resistance of Gram-Negative Bacteria to Fusidic Acid via Forming Drug–Phospholipid Complex" Bioengineering 11, no. 2: 177. https://doi.org/10.3390/bioengineering11020177
APA StyleLiu, J., Lai, X., Li, Y., Yu, Z., Wang, X., Zhang, C., & Peng, Q. (2024). Reversing the Natural Drug Resistance of Gram-Negative Bacteria to Fusidic Acid via Forming Drug–Phospholipid Complex. Bioengineering, 11(2), 177. https://doi.org/10.3390/bioengineering11020177