
Neurovascular Relationships in AGEs-Based Models of Proliferative Diabetic Retinopathy

 ,
, 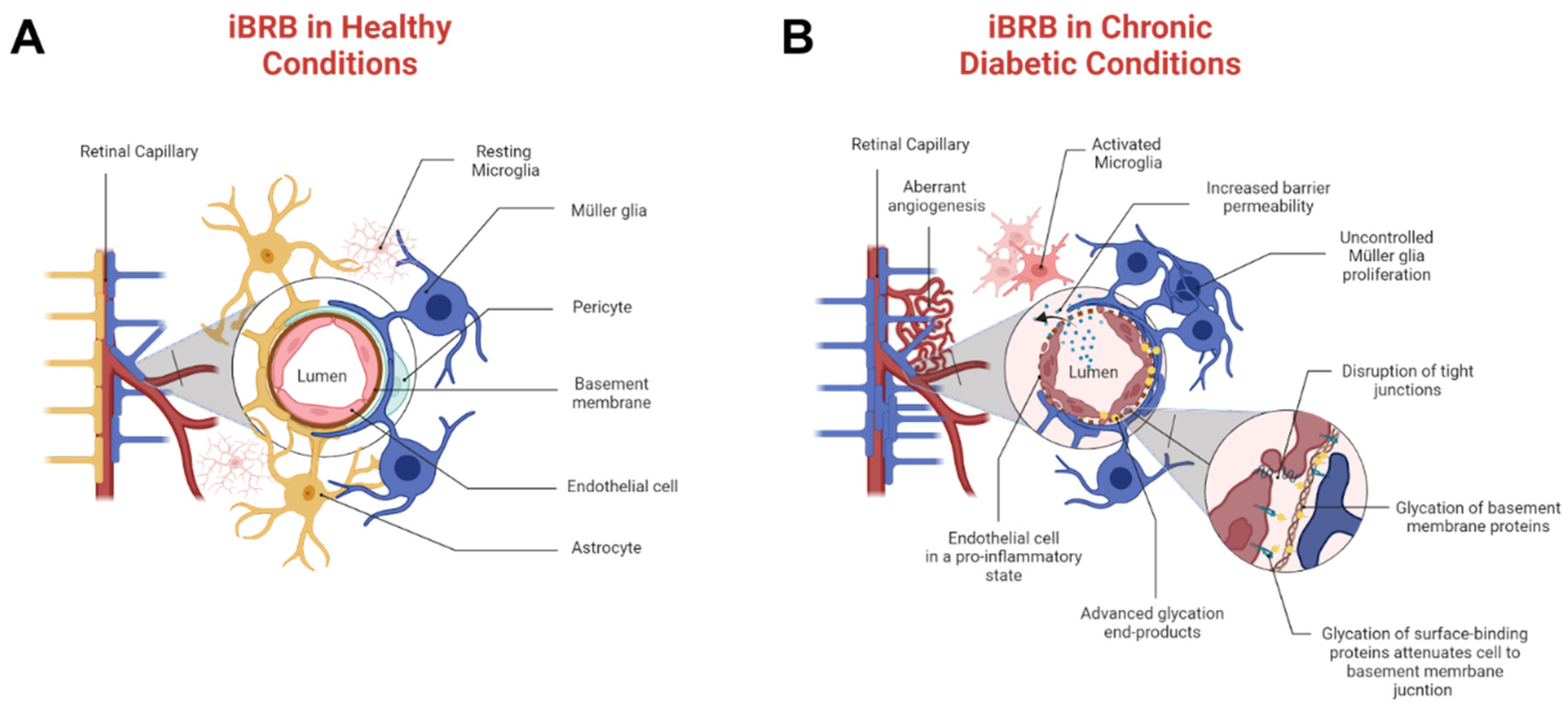{kind=link}
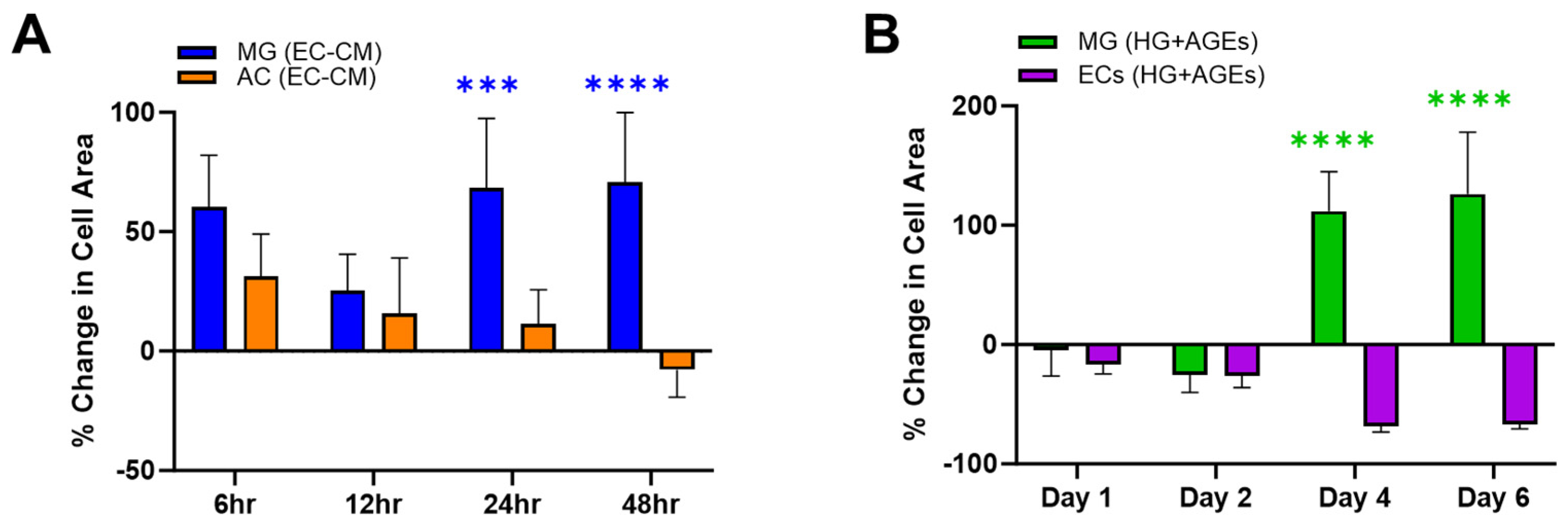{kind=link}
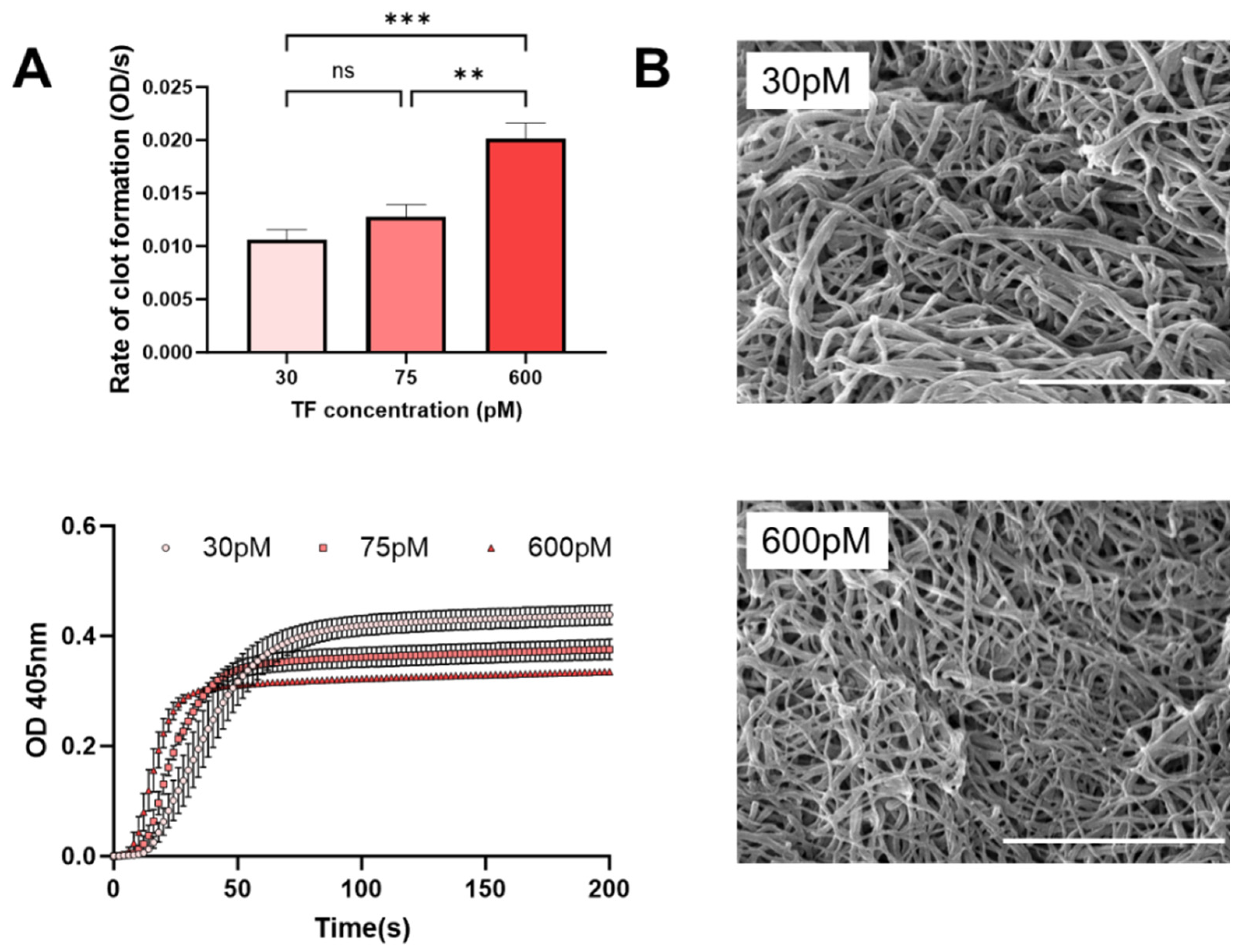{kind=link}
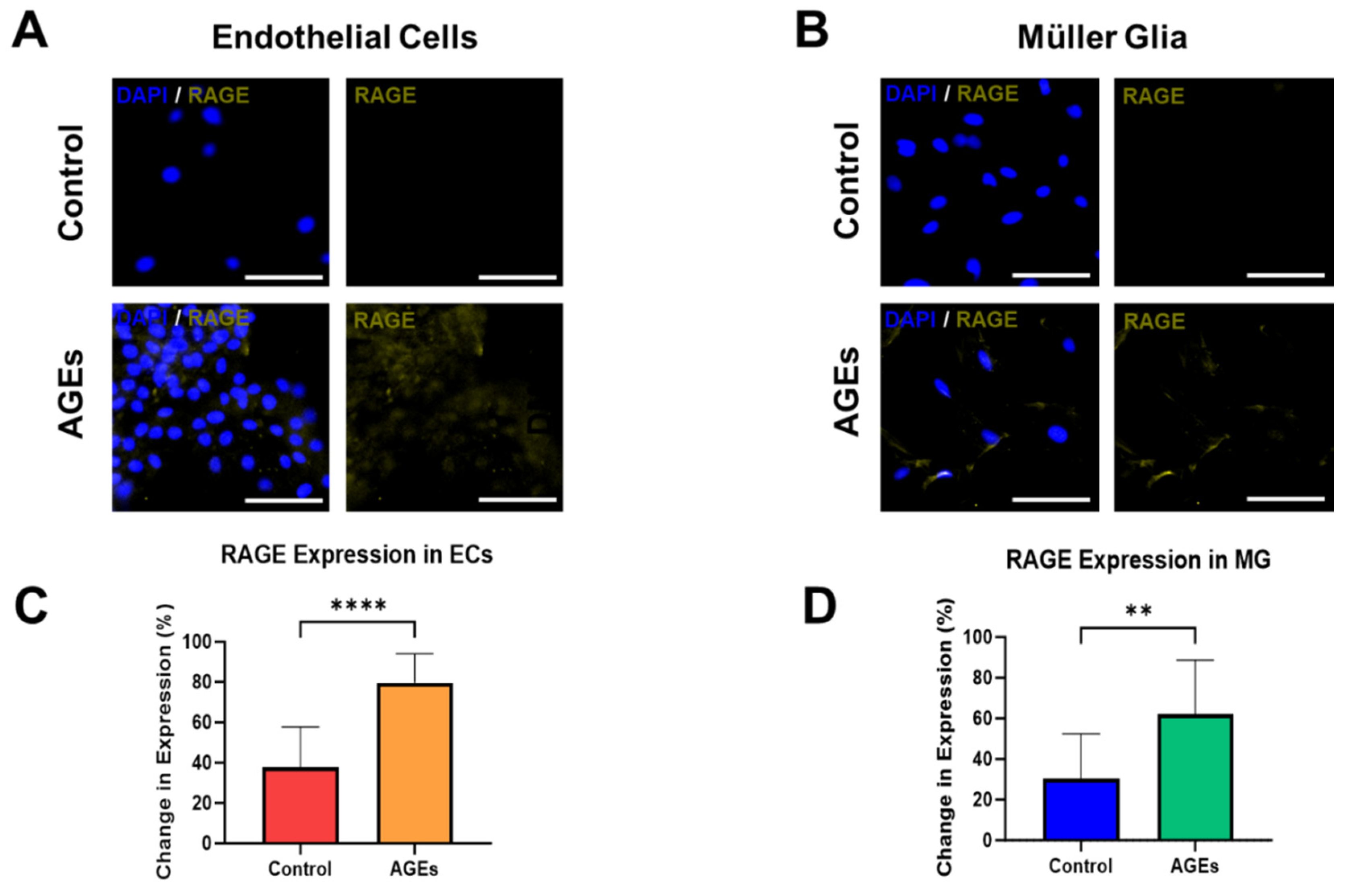{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Conditioned Media
2.3. AGEs/RAGE
2.4. Cell Morphology
2.5. Immunocytochemistry
2.6. Turbidity
2.7. Imaging Analysis
2.8. Statistical Analysis
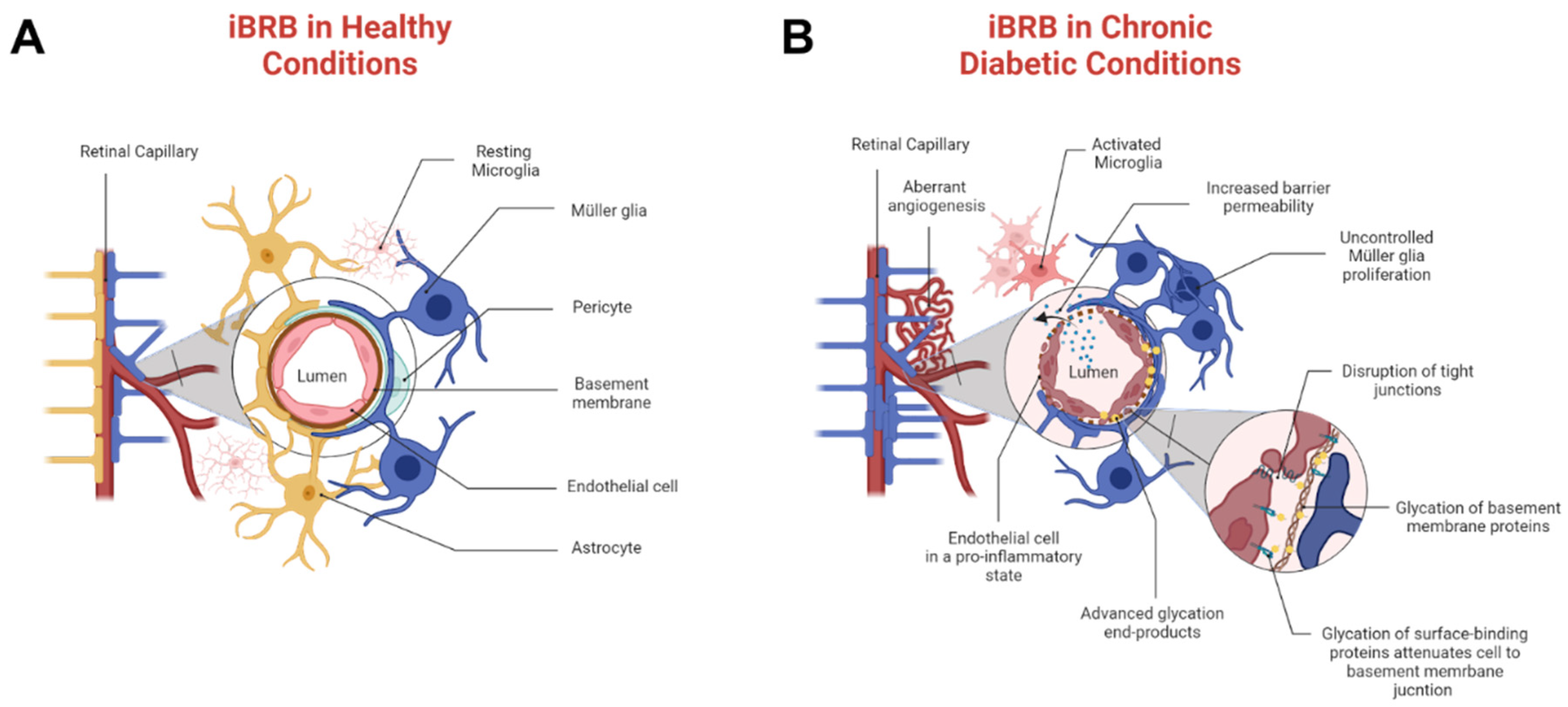
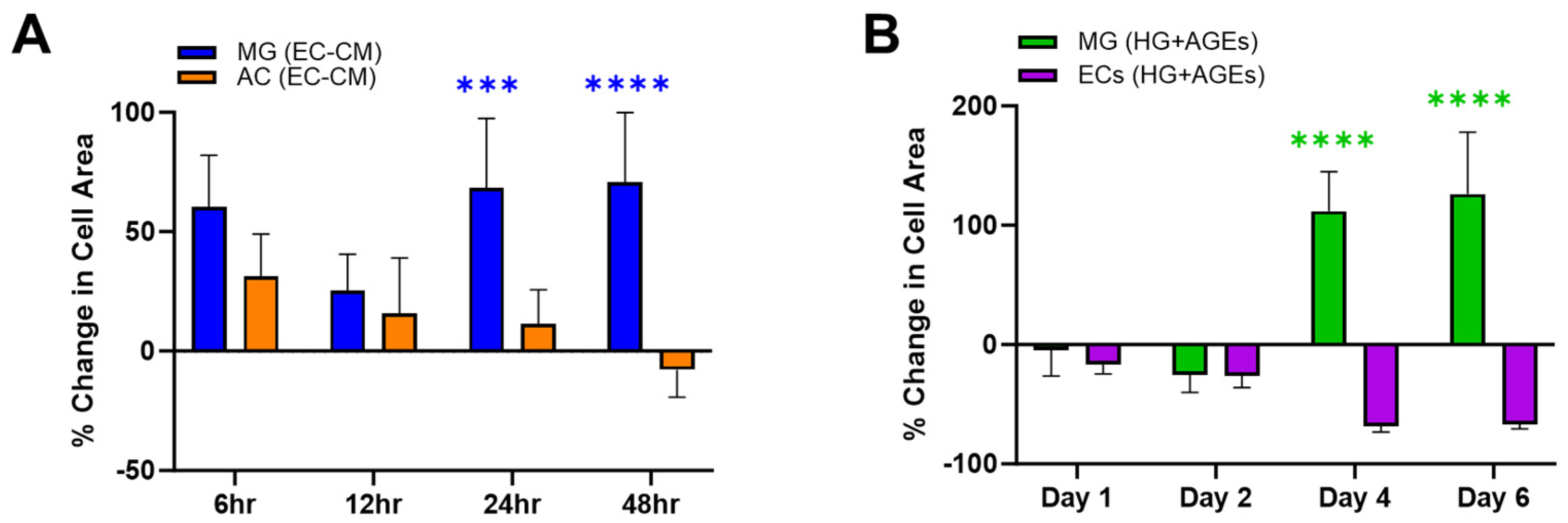
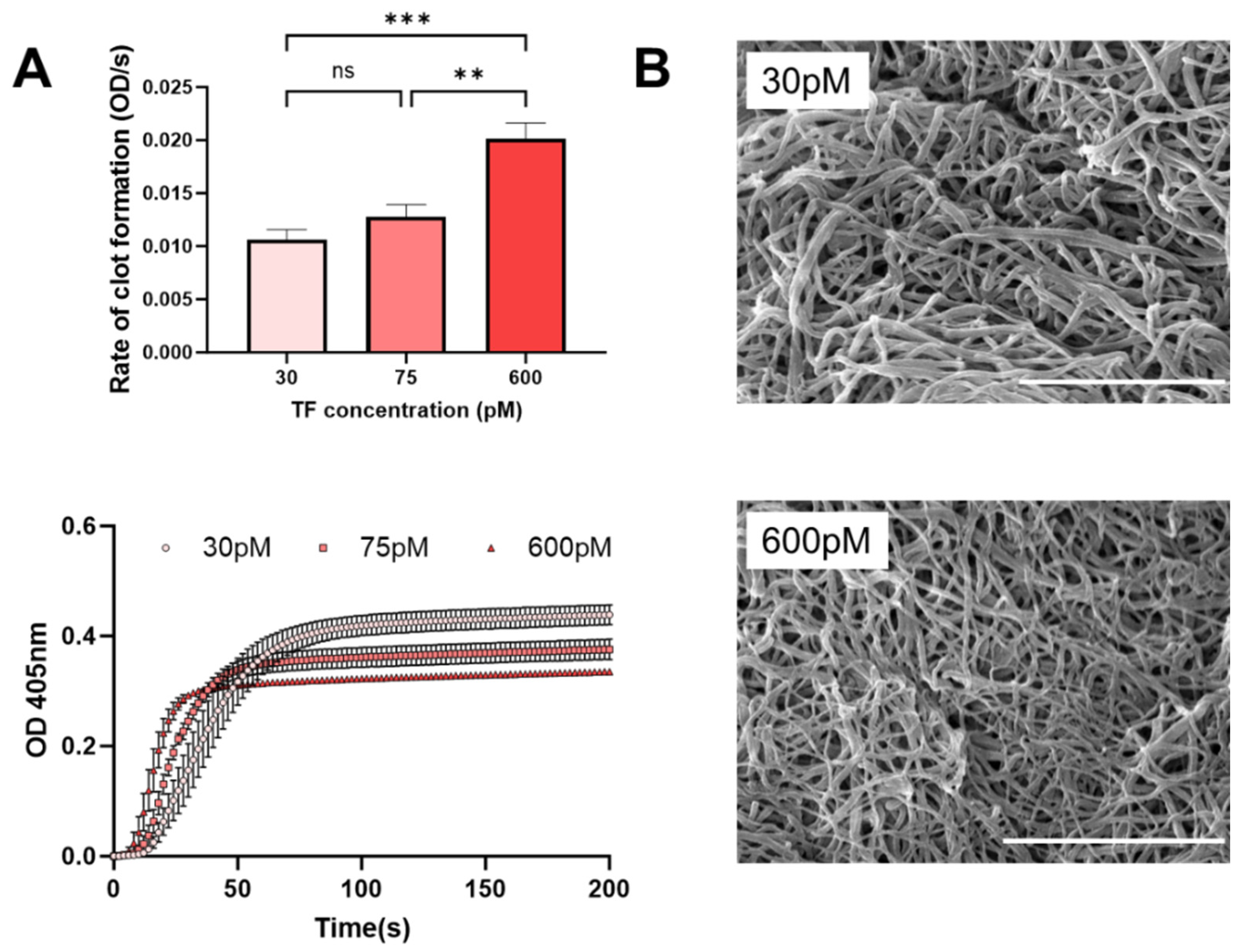
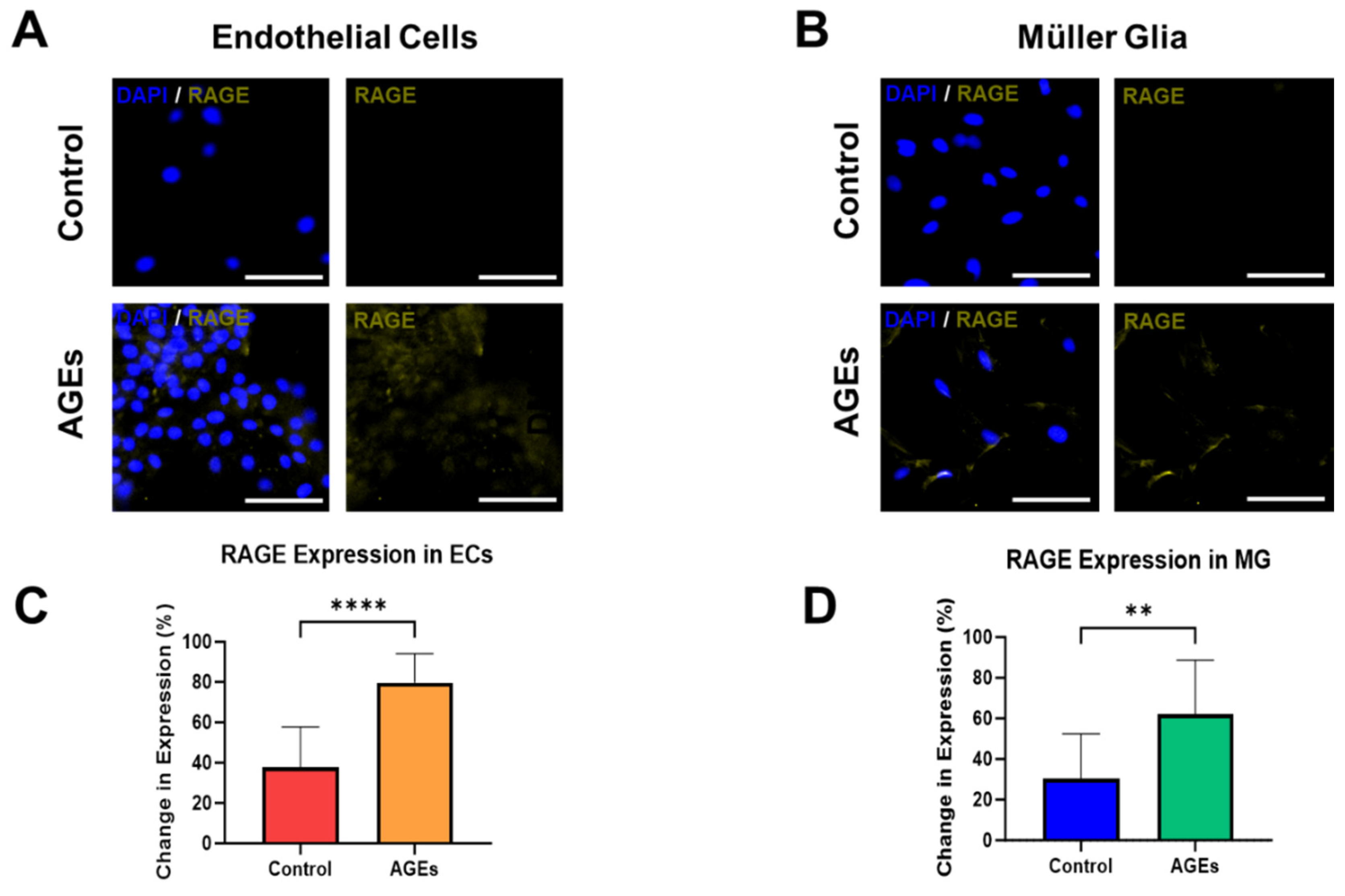
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Teo, Z.L.; Tham, Y.C.; Yu, M.; Chee, M.L.; Rim, T.H.; Cheung, N.; Bikbov, M.M.; Wang, Y.X.; Tang, Y.; Lu, Y.; et al. Global Prevalence of Diabetic Retinopathy and Projection of Burden through 2045: Systematic Review and Meta-analysis. Ophthalmology 2021, 128, 1580–1591. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Chen, L.-J.; Yu, J.; Wang, H.-J.; Zhang, F.; Liu, Q.; Wu, J. Involvement of Advanced Glycation End Products in the Pathogenesis of Diabetic Retinopathy. Cell. Physiol. Biochem. 2018, 48, 705–717. [Google Scholar] [CrossRef] [PubMed]
- Peña, J.S.; Vazquez, M. Reducing health disparities in adult vision loss via interfaces with emerging technology. Eye 2019, 33, 532–533. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H.; Tang, J.; Zhang, F.; Liu, L.; Zhou, J.; Chen, M.; Li, M.; Wu, X.; Nie, Y.; Duan, J. Global trends and performances in diabetic retinopathy studies: A bibliometric analysis. Front. Public Health 2023, 11, 1128008. [Google Scholar] [CrossRef] [PubMed]
- Ferland-McCollough, D.; Slater, S.; Richard, J.; Reni, C.; Mangialardi, G. Pericytes, an overlooked player in vascular pathobiology. Pharmacol. Ther. 2017, 171, 30–42. [Google Scholar] [CrossRef]
- Picard, F.; Adjedj, J.; Varenne, O. Diabetes Mellitus, a prothrombotic disease. Ann. Cardiol. Angeiol. 2017, 66, 385–392. [Google Scholar] [CrossRef] [PubMed]
- Pereiro, X.; Ruzafa, N.; Acera, A.; Urcola, A.; Vecino, E. Optimization of a Method to Isolate and Culture Adult Porcine, Rats and Mice Muller Glia in Order to Study Retinal Diseases. Front. Cell. Neurosci. 2020, 14, 7. [Google Scholar] [CrossRef] [PubMed]
- Dowling, P.; Clynes, M. Conditioned media from cell lines: A complementary model to clinical specimens for the discovery of disease-specific biomarkers. Proteomics 2011, 11, 794–804. [Google Scholar] [CrossRef]
- Cliver, R.N.; Castro, N.; Russomano, T.; Lardieri, G.; Quarrie, L.; van der Merwe, H.; Vazquez, M. Antioxidants derived from natural products reduce radiative damage in cultured retinal glia to prevent oxidative stress. Neuroglia 2022, 3, 84–98. [Google Scholar] [CrossRef]
- Peña, J.S.; Vazquez, M. Harnessing the Neuroprotective Behaviors of Muller Glia for Retinal Repair. Front. Biosci. Landmark Ed. 2022, 27, 169. [Google Scholar] [CrossRef]
- Reichenbach, A.; Bringmann, A. Glia of the human retina. Glia 2020, 68, 768–796. [Google Scholar] [CrossRef] [PubMed]
- Albert-Garay, J.S.; Riesgo-Escovar, J.R.; Salceda, R. High glucose concentrations induce oxidative stress by inhibiting Nrf2 expression in rat Muller retinal cells in vitro. Sci. Rep. 2022, 12, 1261. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Castro, B.; Robel, S.; Mishra, A. Astrocyte Endfeet in Brain Function and Pathology: Open Questions. Annu. Rev. Neurosci. 2023, 46, 101–121. [Google Scholar] [CrossRef] [PubMed]
- Bora, K.; Kushwah, N.; Maurya, M.; Pavlovich, M.C.; Wang, Z.; Chen, J. Assessment of Inner Blood-Retinal Barrier: Animal Models and Methods. Cells 2023, 12, 2443. [Google Scholar] [CrossRef] [PubMed]
- Maurissen, T.L.; Pavlou, G.; Bichsel, C.; Villaseñor, R.; Kamm, R.D.; Ragelle, H. Microphysiological Neurovascular Barriers to Model the Inner Retinal Microvasculature. J. Pers. Med. 2022, 12, 148. [Google Scholar] [CrossRef]
- Chang, W.C.; Lin, C.; Lee, C.H.; Sung, T.L.; Tung, T.H.; Liu, J.H. Vitrectomy with or without internal limiting membrane peeling for idiopathic epiretinal membrane: A meta-analysis. PLoS ONE 2017, 12, e0179105. [Google Scholar] [CrossRef]
- Hosoya, K.; Tachikawa, M. The inner blood-retinal barrier: Molecular structure and transport biology. Adv. Exp. Med. Biol. 2012, 763, 85–104. [Google Scholar]
- Robles-Osorio, M.L.; Sabath, E. Tight junction disruption and the pathogenesis of the chronic complications of diabetes mellitus: A narrative review. World J. Diabetes 2023, 14, 1013–1026. [Google Scholar] [CrossRef]
- Xiong, F.; Leonov, S.; Howard, A.C.; Xiong, S.; Zhang, B.; Mei, L.; McNeil, P.; Simon, S.; Xiong, W.C. Receptor for advanced glycation end products (RAGE) prevents endothelial cell membrane resealing and regulates F-actin remodeling in a beta-catenin-dependent manner. J. Biol. Chem. 2011, 286, 35061–35070. [Google Scholar] [CrossRef]
- Soma, P.; Swanepoel, A.C.; Bester, J.; Pretorius, E. Tissue factor levels in type 2 diabetes mellitus. Inflamm. Res. 2017, 66, 365–368. [Google Scholar] [CrossRef]
- Ichikawa, K.; Yoshinari, M.; Iwase, M.; Wakisaka, M.; Doi, Y.; Iino, K.; Yamamoto, M.; Fujishima, M. Advanced glycosylation end products induced tissue factor expression in human monocyte-like U937 cells and increased tissue factor expression in monocytes from diabetic patients. Atherosclerosis 1998, 136, 281–287. [Google Scholar] [CrossRef] [PubMed]
- Stirban, A.; Gawlowski, T.; Roden, M. Vascular effects of advanced glycation endproducts: Clinical effects and molecular mechanisms. Mol. Metab. 2014, 3, 94–108. [Google Scholar] [CrossRef] [PubMed]
- Kaur, G.; Singh, N.K. The Role of Inflammation in Retinal Neurodegeneration and Degenerative Diseases. Int. J. Mol. Sci. 2021, 23, 386. [Google Scholar] [CrossRef] [PubMed]
- Olekson, M.P.; Faulknor, R.A.; Hsia, H.C.; Schmidt, A.M.; Berthiaume, F. Soluble Receptor for Advanced Glycation End Products Improves Stromal Cell-Derived Factor-1 Activity in Model Diabetic Environments. Adv. Wound Care 2016, 5, 527–538. [Google Scholar] [CrossRef]
- Nonaka, K.; Kajiura, Y.; Bando, M.; Sakamoto, E.; Inagaki, Y.; Lew, J.H.; Naruishi, K.; Ikuta, T.; Yoshida, K.; Kobayashi, T.; et al. Advanced glycation end-products increase IL-6 and ICAM-1 expression via RAGE, MAPK and NF-kappaB pathways in human gingival fibroblasts. J. Periodontal Res. 2018, 53, 334–344. [Google Scholar] [CrossRef]
- Zong, H.; Ward, M.; Madden, A.; Yong, P.H.; Limb, G.A.; Curtis, T.M.; Stitt, A.W. Hyperglycaemia-induced pro-inflammatory responses by retinal Muller glia are regulated by the receptor for advanced glycation end-products (RAGE). Diabetologia 2010, 53, 2656–2666. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peña, J.S.; Ramanujam, R.K.; Risman, R.A.; Tutwiler, V.; Berthiaume, F.; Vazquez, M. Neurovascular Relationships in AGEs-Based Models of Proliferative Diabetic Retinopathy. Bioengineering 2024, 11, 63. https://doi.org/10.3390/bioengineering11010063
Peña JS, Ramanujam RK, Risman RA, Tutwiler V, Berthiaume F, Vazquez M. Neurovascular Relationships in AGEs-Based Models of Proliferative Diabetic Retinopathy. Bioengineering. 2024; 11(1):63. https://doi.org/10.3390/bioengineering11010063
Chicago/Turabian StylePeña, Juan S., Ranjini K. Ramanujam, Rebecca A. Risman, Valerie Tutwiler, Francois Berthiaume, and Maribel Vazquez. 2024. "Neurovascular Relationships in AGEs-Based Models of Proliferative Diabetic Retinopathy" Bioengineering 11, no. 1: 63. https://doi.org/10.3390/bioengineering11010063
APA StylePeña, J. S., Ramanujam, R. K., Risman, R. A., Tutwiler, V., Berthiaume, F., & Vazquez, M. (2024). Neurovascular Relationships in AGEs-Based Models of Proliferative Diabetic Retinopathy. Bioengineering, 11(1), 63. https://doi.org/10.3390/bioengineering11010063