
Unsupervised Multitaper Spectral Method for Identifying REM Sleep in Intracranial EEG Recordings Lacking EOG/EMG Data

Abstract
:1. Introduction
2. Methods
2.1. Data Collection
2.2. Electrode Localization
2.3. Manual Sleep Scoring
2.4. Data Selection
2.5. Filtering and Sectioning
2.6. Multitaper Eigencoefficients
2.7. Multitaper Eigencoefficient Clustering
- Specify the number of clusters.
- Given for each cluster, the feature matrices, , (i.e., one for every 30-s episode), are re-clustered by assigning them the label that maximizes Equation (4).
- Using the new cluster assignments, and are re-computed.
- Steps 3 and 4 are repeated until an iteration occurs without a resulting change in the feature-matrix label. Once this occurs, subsequent iterations result in no change, and the algorithm has converged.
2.8. REM Cluster
2.9. Measures of Label Confidence
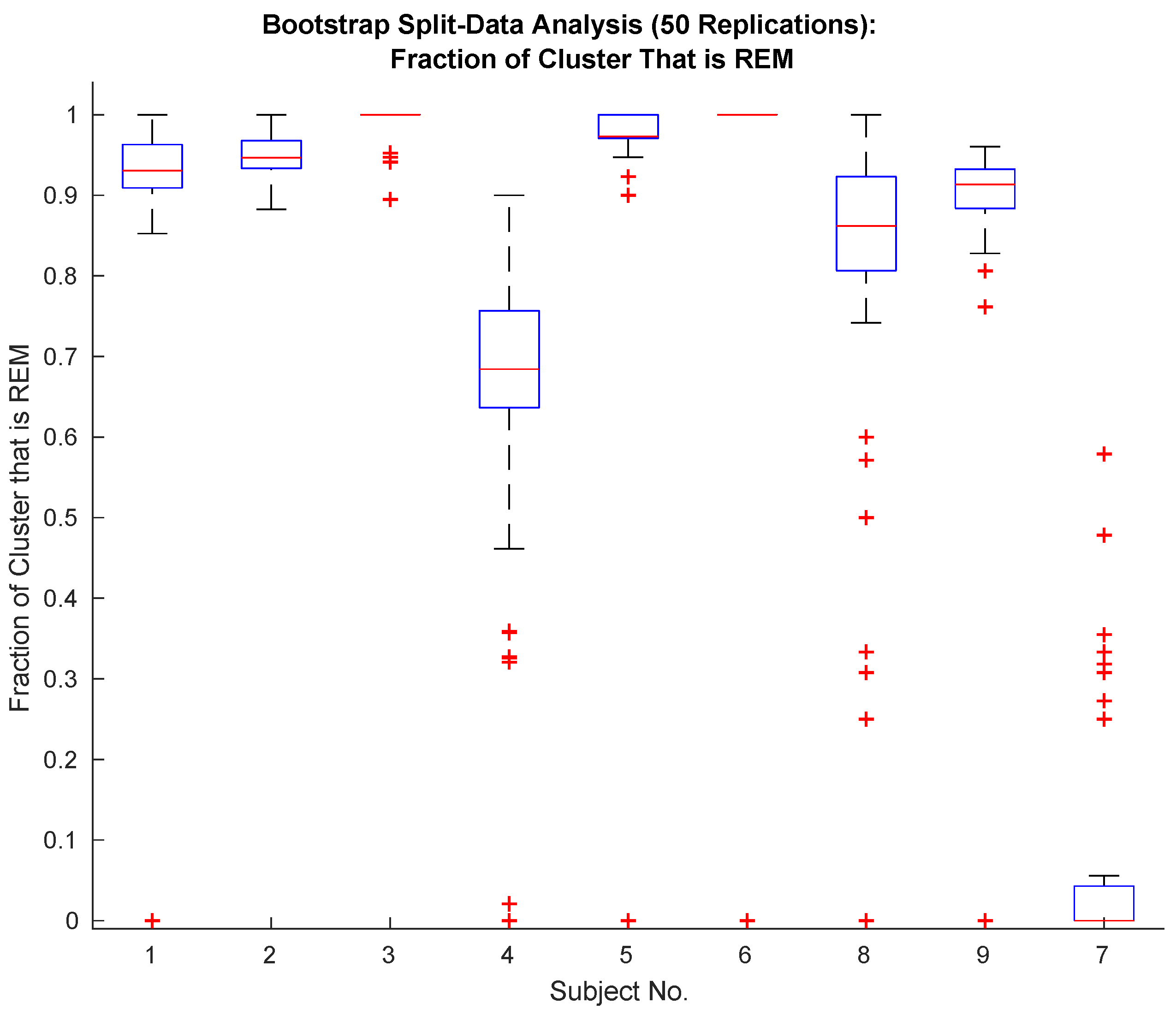
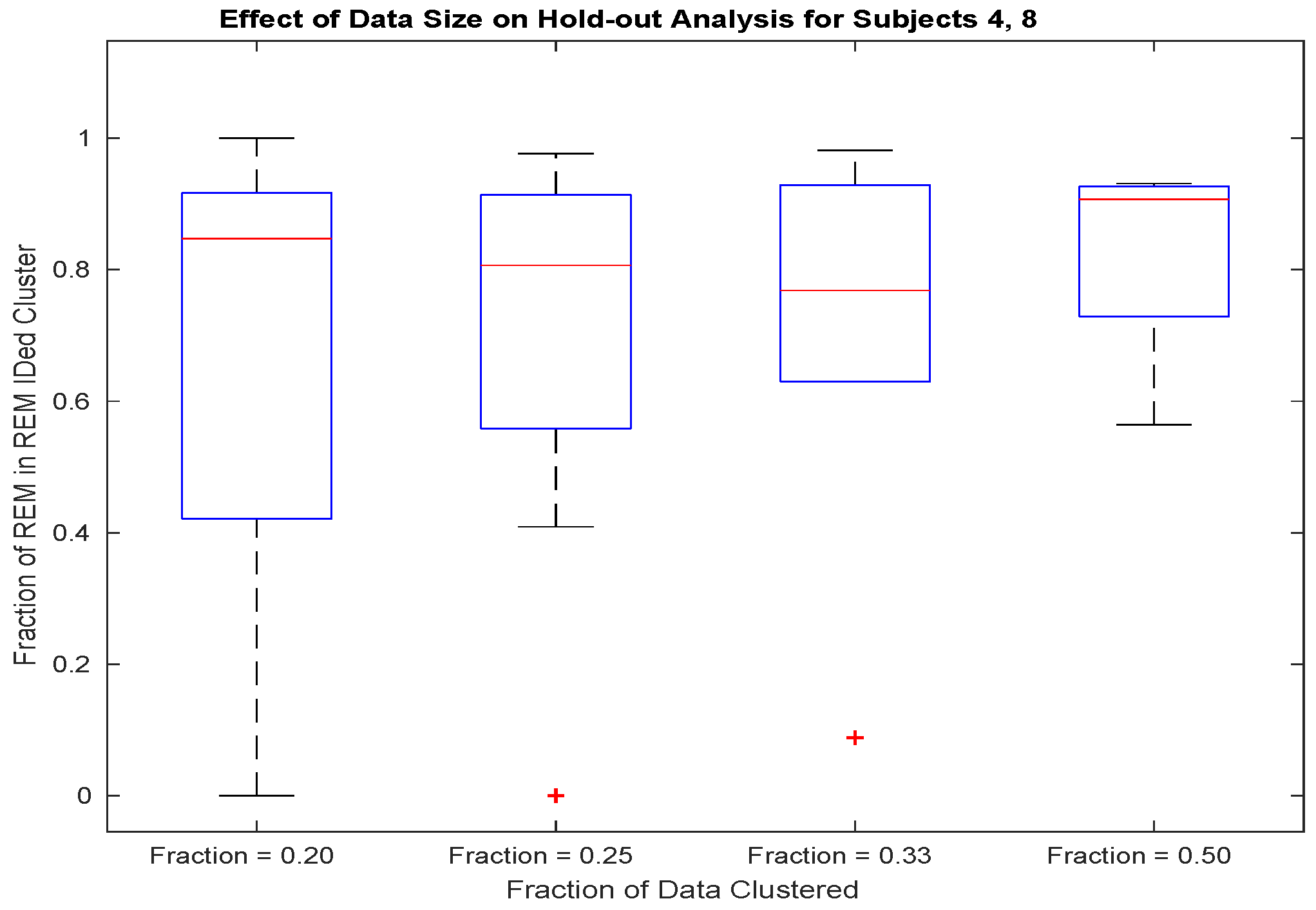
2.10. Hold-Out Analysis
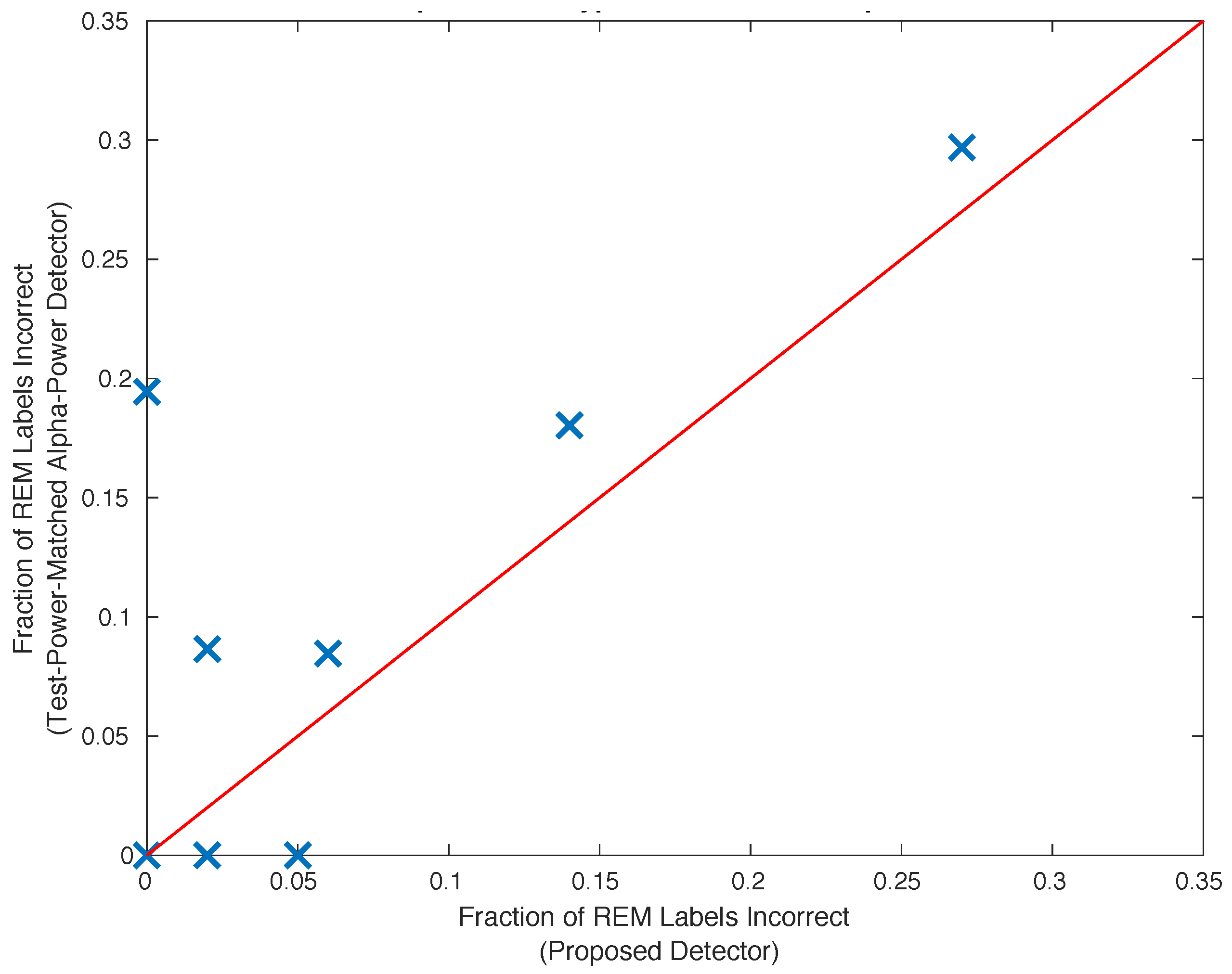
2.11. Comparison with the Alpha Power Detector
2.12. Clustering Pseudo-Code
- Divide the data into 30 s, non-overlapping sections of data.
- Compute the eiegencoefficients characterizing each 30 s section of data.
- Specify the number of clusters to be equal to 2.
- Randomly assign a cluster label to each of the 30 s sections of data.
- Compute each of the cluster statistics conditioned upon the cluster labels.
- For each 30 s section of data, and for each cluster, compute the log-probability of the observed section data conditioned upon each of the cluster labels.
- Assign to each 30 s section of data the cluster label that maximizes the log-probability of the observed 30 s section eigencoefficients.
- Repeat steps 4 through 7 until the cluster labels do not change from one repetition to the next.
- Compute the approximate AIC, , for this number of clusters, using all of the log-probabilities.
- Repeat steps 3 through 9 but increment the number of clusters on each repetition until 14 clusters are used, or until an empty cluster results.
2.13. REM Identification Pseudo-Code
- Average the cluster-specific alpha spectral power across 30 s episodes.
- For each electrode contact, order the average cluster alpha spectral power across clusters.
- For each cluster, compute the median of the ranks across the contacts. This is the spectral rank.
- The REM cluster is the cluster with the smallest spectral rank.
2.14. Software Overview
3. Results

{kind=link}
{kind=link}
{kind=link}
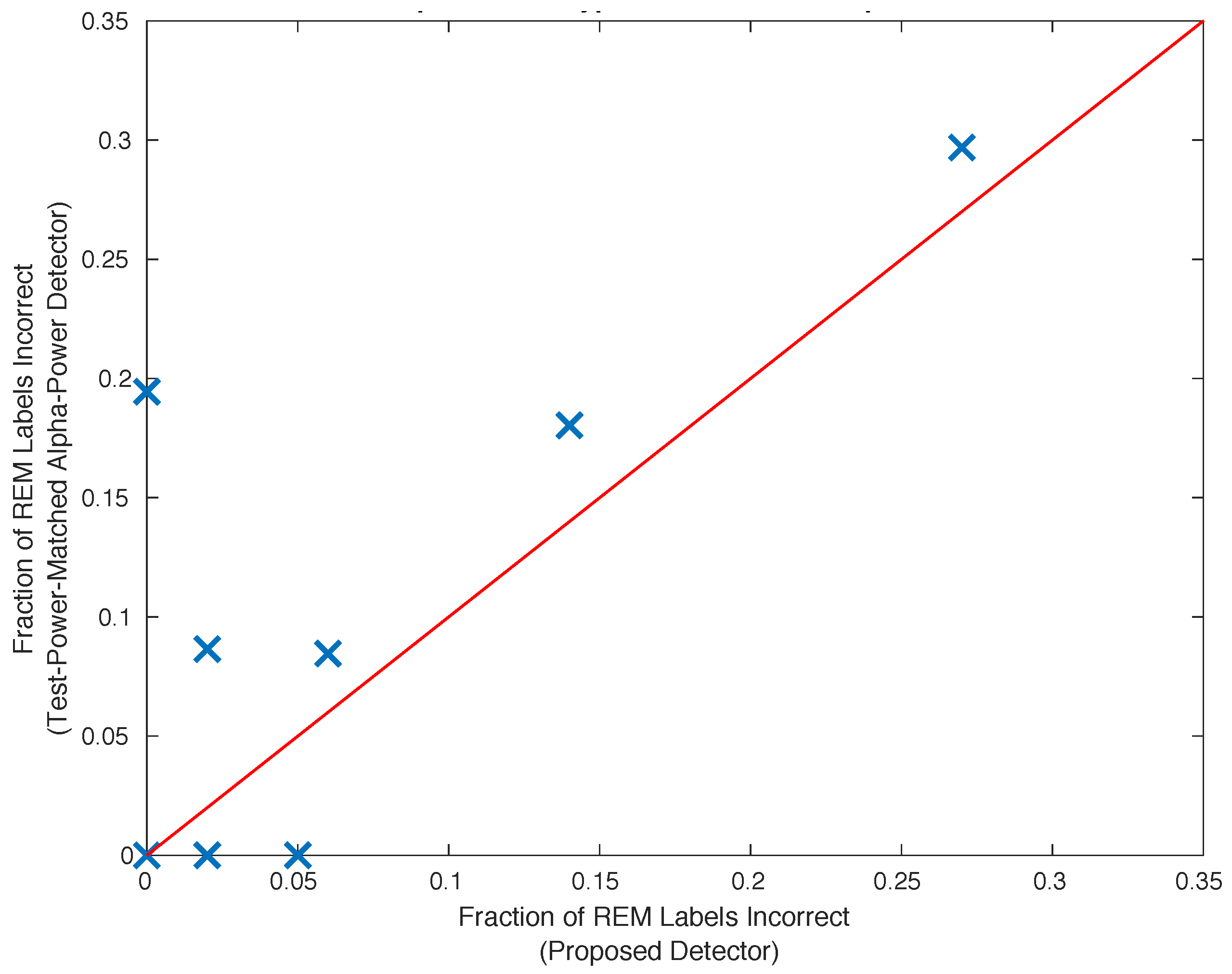{kind=link}
| Subject | 1 | 2 | 3 | 4 | 5 | 6 | 8 | 9 | 7 |
|---|---|---|---|---|---|---|---|---|---|
| Minutes of REM Correctly ID’ed | 47.5 | 26.0 | 14.5 | 22.5 | 31.5 | 62.5 | 25.0 | 60.0 | NA |
| Fraction of REM Labels Correct | 0.98 | 0.95 | 1.00 | 0.73 | 0.98 | 1.00 | 0.86 | 0.94 | 0.00 |
| Fraction of REM Correctly ID’ed | 0.77 | 0.42 | 0.60 | 0.40 | 0.88 | 0.78 | 0.79 | 0.67 | NA |
| Min. Alpha Power Diff. B/W Closest Competing Cluster and REM Cluster | 0.79 | 0.07 | 0.09 | 0.01 | 0.07 | 0.74 | 0.06 | ||
| Fraction of REM Cluster Electrode Alpha Power Not Minimum | 0.00 | 0.10 | 0.20 | 0.08 | 0.09 | 0.00 | 0.00 | 0.18 | 0.00 |
| Number of Clusters | 14 | 12 | 10 | 14 | 8 | 6 | 14 | 14 | 14 |
| Number of Electrode Contacts | 12 | 9 | 10 | 12 | 11 | 12 | 3 | 11 | 2 |
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| EEG | electroencephalography |
| iEEG | intracranial EEG |
| EOG | electrooculography |
| EMG | electromyography |
| REM | rapid eye movement |
| NREM | non rapid eye movement |
| AIC | Akaike information criterion |
References
- Nobili, L.; Frauscher, B.; Eriksson, S.; Gibbs, S.A.; Halasz, P.; Lambert, I.; Manni, R.; Peter-Derex, L.; Proserpio, P.; Provini, F.; et al. Sleep and epilepsy: A snapshot of knowledge and future research lines. J. Sleep Res. 2022, 31, e13622. [Google Scholar] [CrossRef] [PubMed]
- Vakharia, V.N.; Duncan, J.S.; Witt, J.A.; Elger, C.E.; Staba, R.; Engel, J., Jr. Getting the best outcomes from epilepsy surgery. Ann. Neurol. 2018, 83, 676–690. [Google Scholar] [CrossRef] [PubMed]
- Demuru, M.; van Blooijs, D.; Zweiphenning, W.; Hermes, D.; Leijten, F.; Zijlmans, M.; group, R. A practical workflow for organizing clinical intraoperative and long-term iEEG data in BIDS. Neuroinformatics 2022, 20, 727–736. [Google Scholar] [CrossRef] [PubMed]
- Iber, C.; Ancoli-Israel, S.; Chesson, A.L.; Quan, S.F. The new sleep scoring manual–the evidence behind the rules. J. Clin. Sleep Med. 2007, 3, 107. [Google Scholar] [CrossRef]
- McLeod, G.A.; Ghassemi, A.; Ng, M.C. Can REM sleep localize the epileptogenic zone? A systematic review and analysis. Front. Neurol. 2020, 11, 584. [Google Scholar] [CrossRef] [PubMed]
- Kang, X.; Boly, M.; Findlay, G.; Jones, B.; Gjini, K.; Maganti, R.; Struck, A.F. Quantitative spatio-temporal characterization of epileptic spikes using high density EEG: Differences between NREM sleep and REM sleep. Sci. Rep. 2020, 10, 1673. [Google Scholar] [CrossRef] [PubMed]
- Noachtar, S.; Borggraefe, I. Epilepsy surgery: A critical review. Epilepsy Behav. 2009, 15, 66–72. [Google Scholar] [CrossRef]
- Englot, D.J.; Han, S.J.; Rolston, J.D.; Ivan, M.E.; Kuperman, R.A.; Chang, E.F.; Gupta, N.; Sullivan, J.E.; Auguste, K.I. Epilepsy surgery failure in children: A quantitative and qualitative analysis. J. Neurosurg. Pediatr. 2014, 14, 386–395. [Google Scholar] [CrossRef]
- Englot, D.J.; Raygor, K.P.; Molinaro, A.M.; Garcia, P.A.; Knowlton, R.C.; Auguste, K.I.; Chang, E.F. Factors associated with failed focal neocortical epilepsy surgery. Neurosurgery 2014, 75, 648. [Google Scholar] [CrossRef]
- Vijayan, S.; Lepage, K.Q.; Kopell, N.J.; Cash, S.S. Frontal beta-theta network during REM sleep. elife 2017, 6, e18894. [Google Scholar] [CrossRef]
- Kubicki, S.; Höller, L.; Berg, I.; Pastelak-Price, C.; Dorow, R. Sleep EEG evaluation: A comparison of results obtained by visual scoring and automatic analysis with the Oxford sleep stager. Sleep 1989, 12, 140–149. [Google Scholar] [CrossRef]
- Flexerand, A.; Dorffner, G.; Sykacekand, P.; Rezek, I. An automatic, continuous and probabilistic sleep stager based on a hidden Markov model. Appl. Artif. Intell. 2002, 16, 199–207. [Google Scholar] [CrossRef]
- Björn, R.; Born, J. About sleep’s role in memory. Physiol. Rev. 2013, 93, 681–766. [Google Scholar]
- Peever, J.; Fuller, P.M. The biology of REM sleep. Curr. Biol. 2017, 27, R1237–R1248. [Google Scholar] [CrossRef] [PubMed]
- Nir, Y.; Tononi, G. Dreaming and the brain: From phenomenology to neurophysiology. Trends Cogn. Sci. 2010, 14, 88–100. [Google Scholar] [CrossRef]
- Blumberg, M.S.; Lesku, J.A.; Libourel, P.A.; Schmidt, M.H.; Rattenborg, N.C. What is REM sleep? Curr. Biol. 2020, 30, R38–R49. [Google Scholar] [CrossRef]
- Siegel, J.M. REM sleep. Princ. Pract. Sleep Med. 2005, 4, 120–135. [Google Scholar]
- Gaillard, J. Principles of Automatic Analysis of sleep records with a hybrid system. Comput. Biomed. Res. 1973, 6, 1–13. [Google Scholar] [CrossRef]
- Lacroix, B.; Hanus, R. Online automatic sleep analysis. JORBEL-Belg. J. Oper. Res. Stat. Comput. Sci. 1984, 24, 39–47. [Google Scholar]
- Gath, I.; Bar-On, E. Computerized method for scoring of polygraphic sleep recordings. Comput. Programs Biomed. 1980, 11, 217–223. [Google Scholar] [CrossRef]
- Lim, A.J.; Winters, W.D. A practical method for automatic real-time EEG sleep state analysis. IEEE Trans. Biomed. Eng. 1980, BME-27, 212–220. [Google Scholar] [CrossRef] [PubMed]
- Loh, H.W.; Ooi, C.P.; Vicnesh, J.; Oh, S.L.; Faust, O.; Gertych, A.; Acharya, U.R. Automated detection of sleep stages using deep learning techniques: A systematic review of the last decade (2010–2020). Appl. Sci. 2020, 10, 8963. [Google Scholar] [CrossRef]
- Michielli, N.; Acharya, U.R.; Molinari, F. Cascaded LSTM recurrent neural network for automated sleep stage classification using single-channel EEG signals. Comput. Biol. Med. 2019, 106, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Khare, S.K.; Bajaj, V.; Taran, S.; Sinha, G. Multiclass sleep stage classification using artificial intelligence based time-frequency distribution and CNN. In Artificial Intelligence-Based Brain-Computer Interface; Elsevier: Amsterdam, The Netherlands, 2022; pp. 1–21. [Google Scholar]
- Eldele, E.; Chen, Z.; Liu, C.; Wu, M.; Kwoh, C.K.; Li, X.; Guan, C. An attention-based deep learning approach for sleep stage classification with single-channel EEG. IEEE Trans. Neural Syst. Rehabil. Eng. 2021, 29, 809–818. [Google Scholar] [CrossRef] [PubMed]
- Sekkal, R.N.; Bereksi-Reguig, F.; Ruiz-Fernandez, D.; Dib, N.; Sekkal, S. Automatic sleep stage classification: From classical machine learning methods to deep learning. Biomed. Signal Process. Control 2022, 77, 103751. [Google Scholar] [CrossRef]
- Van Der Donckt, J.; Van Der Donckt, J.; Deprost, E.; Vandenbussche, N.; Rademaker, M.; Vandewiele, G.; Van Hoecke, S. Do not sleep on traditional machine learning: Simple and interpretable techniques are competitive to deep learning for sleep scoring. Biomed. Signal Process. Control 2023, 81, 104429. [Google Scholar] [CrossRef]
- von Ellenrieder, N.; Peter-Derex, L.; Gotman, J.; Frauscher, B. SleepSEEG: Automatic sleep scoring using intracranial EEG recordings only. J. Neural Eng. 2022, 19, 026057. [Google Scholar] [CrossRef] [PubMed]
- Berger, H. On the electroencephalogram of man. Electroencephalogr. Clin. Neurophysiol. 1969, (Suppl. 28), 37+. [Google Scholar]
- Wright, K.P., Jr.; Badia, P.; Wauquier, A. Topographical and temporal patterns of brain activity during the transition from wakefulness to sleep. Sleep 1995, 18, 880–889. [Google Scholar] [CrossRef]
- Putilov, A.A.; Donskaya, O.G. Alpha attenuation soon after closing the eyes as an objective indicator of sleepiness. Clin. Exp. Pharmacol. Physiol. 2014, 41, 956–964. [Google Scholar] [CrossRef]
- De Gennaro, L.; Ferrara, M.; Bertini, M. The boundary between wakefulness and sleep: Quantitative electroencephalographic changes during the sleep onset period. Neuroscience 2001, 107, 1–11. [Google Scholar] [CrossRef]
- Silber, M.H.; Ancoli-Israel, S.; Bonnet, M.H.; Chokroverty, S.; Grigg-Damberger, M.M.; Hirshkowitz, M.; Kapen, S.; Keenan, S.A.; Kryger, M.H.; Penzel, T.; et al. The visual scoring of sleep in adults. J. Clin. Sleep Med. 2007, 3, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Bokil, H.; Andrews, P.; Kulkarni, J.E.; Mehta, S.; Mitra, P.P. Chronux: A platform for analyzing neural signals. J. Neurosci. Methods 2010, 192, 146–151. [Google Scholar] [CrossRef]
- von Ellenrieder, N. SleepSEEG: Matlab Code for Sleep Scoring Based on Intracranial EEG (v.2). 2022. Available online: https://zenodo.org/record/7410501 (accessed on 5 June 2023).
- Janca, R.; Jezdik, P.; Cmejla, R.; Tomasek, M.; Worrell, G.A.; Stead, M.; Wagenaar, J.; Jefferys, J.G.; Krsek, P.; Komarek, V.; et al. Detection of interictal epileptiform discharges using signal envelope distribution modelling: Application to epileptic and non-epileptic intracranial recordings. Brain Topogr. 2015, 28, 172–183. [Google Scholar] [CrossRef] [PubMed]
- Gath, I.; Feuerstein, C.; Geva, A. Unsupervised classification and adaptive definition of sleep patterns. Pattern Recognit. Lett. 1994, 15, 977–984. [Google Scholar] [CrossRef]
- Aboalayon, K.A.I.; Faezipour, M.; Almuhammadi, W.S.; Moslehpour, S. Sleep Stage Classification Using EEG Signal Analysis: A Comprehensive Survey and New Investigation. Entropy 2016, 18, 272. [Google Scholar] [CrossRef]
- Güneş, S.; Polat, K.; Yosunkaya, S.; Dursun, M. A novel data pre-processing method on automatic determining of sleep stages: K-means clustering based feature weighting. Complex Syst. Appl. ICCSA 2009, 6, 112–117. [Google Scholar]
- Güneş, S.; Polat, K.; Yosunkaya, Ş. Efficient sleep stage recognition system based on EEG signal using k-means clustering based feature weighting. Expert Syst. Appl. 2010, 37, 7922–7928. [Google Scholar] [CrossRef]
- Buckelmüller, J.; Landolt, H.P.; Stassen, H.; Achermann, P. Trait-like individual differences in the human sleep electroencephalogram. Neuroscience 2006, 138, 351–356. [Google Scholar] [CrossRef]
- Koupparis, A.M.; Kokkinos, V.; Kostopoulos, G.K. Semi-automatic sleep EEG scoring based on the hypnospectrogram. J. Neurosci. Methods 2014, 221, 189–195. [Google Scholar] [CrossRef]
- Dale, A.M.; Fischl, B.; Sereno, M.I. Cortical surface-based analysis: I. Segmentation and surface reconstruction. Neuroimage 1999, 9, 179–194. [Google Scholar] [CrossRef] [PubMed]
- Fischl, B.; Salat, D.H.; Busa, E.; Albert, M.; Dieterich, M.; Haselgrove, C.; Van Der Kouwe, A.; Killiany, R.; Kennedy, D.; Klaveness, S.; et al. Whole brain segmentation: Automated labeling of neuroanatomical structures in the human brain. Neuron 2002, 33, 341–355. [Google Scholar] [CrossRef] [PubMed]
- Desikan, R.S.; Ségonne, F.; Fischl, B.; Quinn, B.T.; Dickerson, B.C.; Blacker, D.; Buckner, R.L.; Dale, A.M.; Maguire, R.P.; Hyman, B.T.; et al. An automated labeling system for subdividing the human cerebral cortex on MRI scans into gyral based regions of interest. Neuroimage 2006, 31, 968–980. [Google Scholar] [CrossRef]
- LaPlante, R.A.; Tang, W.; Peled, N.; Vallejo, D.I.; Borzello, M.; Dougherty, D.D.; Eskandar, E.N.; Widge, A.S.; Cash, S.S.; Stufflebeam, S.M. The interactive electrode localization utility: Software for automatic sorting and labeling of intracranial subdural electrodes. Int. J. Comput. Assist. Radiol. Surg. 2017, 12, 1829–1837. [Google Scholar] [CrossRef] [PubMed]
- Jain, S. Investigation of Sleep Neural Dynamics in Intracranial EEG Patients. Master’s Thesis, Virginia Tech, Blacksburg, VA, USA, 2021. [Google Scholar]
- Slepian, D. Prolate spheroidal wave functions, Fourier analysis, and uncertainty V: The discrete case. Bell Syst. Tech. J. 1978, 57, 1371–1430. [Google Scholar] [CrossRef]
- Thomson, D.J. Spectrum estimation and harmonic analysis. Proc. IEEE 1982, 70, 1055–1096. [Google Scholar] [CrossRef]
- Lepage, K.Q.; Thomson, D.J. Reduced mean-square error quadratic inverse spectrum estimator. IEEE Trans. Signal Process. 2014, 62, 2958–2972. [Google Scholar]
- Lepage, K.Q.; Fleming, C.N.; Witcher, M.; Vijayan, S. Multitaper estimates of phase-amplitude coupling. J. Neural Eng. 2021, 18, 056028. [Google Scholar] [CrossRef]
- Cerioli, A. K-means cluster analysis and mahalanobis metrics: A problematic match or an overlooked opportunity. Stat. Appl. 2005, 17, 61–73. [Google Scholar]
- Martino, A.; Ghiglietti, A.; Ieva, F.; Paganoni, A.M. A K-means procedure based on a Mahalanobis type distance for clustering multivariate functional data. Stat. Methods Appl. 2019, 28, 301–322. [Google Scholar] [CrossRef]
- Brillinger, D.R. Time Series: Data Analysis and Theory; Society for Industrial and Applied Mathematics: Philadelphia, PA, USA, 2001. [Google Scholar]
- Faber, V. Clustering and the continuous k-means algorithm. Los Alamos Sci. 1994, 22, 67. [Google Scholar]
- Akaike, H. A new look at the statistical model identification. IEEE Trans. Autom. Control 1974, 19, 716–723. [Google Scholar] [CrossRef]
- deLeeuw, J. Introduction to Akaike (1973) information theory and an extension of the maximum likelihood principle. In Breakthroughs in Statistics; Springer: New York, NY, USA, 1992; pp. 599–609. [Google Scholar]
- Prieto, G.A.; Parker, R.; Vernon Iii, F. A Fortran 90 library for multitaper spectrum analysis. Comput. Geosci. 2009, 35, 1701–1710. [Google Scholar] [CrossRef]
- Lees, J.M.; Park, J. Multiple-taper spectral analysis: A stand-alone C-subroutine. Comput. Geosci. 1995, 21, 199–236. [Google Scholar] [CrossRef]
- Denovellis, E.L.; Myroshnychenko, M.; Sarmashghi, M.; Stephen, E.P. Spectral Connectivity: A python package for computing multitaper spectral estimates and frequency-domain brain connectivity measures on the CPU and GPU. J. Open Source Softw. 2022, 7, 4840. [Google Scholar] [CrossRef]
- O’Brien, C.B.; Baghdoyan, H.A.; Lydic, R. Computer-based multitaper spectrogram program for electroencephalographic data. JoVE J. Vis. Exp. 2019, 153, e60333. [Google Scholar]
- Rahim, K.; Burr, W. Multitaper: Spectral Analysis Tools Using the Multitaper Method. 2020. Available online: https://cran.r-project.org/web/packages/multitaper/index.html (accessed on 14 August 2023).
- Danker-hopfe, H.; Anderer, P.; Zeitlhofer, J.; Boeck, M.; Dorn, H.; Gruber, G.; Heller, E.; Loretz, E.; Moser, D.; Parapatics, S.; et al. Interrater reliability for sleep scoring according to the Rechtschaffen & Kales and the new AASM standard. J. Sleep Res. 2009, 18, 74–84. [Google Scholar]
- Percival, D.; Walden, A. Spectral Analysis For Physical Applications; Cambridge University Press: Cambridge, UK, 1993. [Google Scholar]
- Scharf, L.L. Statistical Signal Processing: Detection, Estimation, and Time Series Analysis; Addison-Wesley Publishing Company, Inc.: Boston, MA, USA, 1991. [Google Scholar]
- Hastie, T.; Tibshirani, R.; Friedman, J.H.; Friedman, J.H. The Elements of Statistical Learning: Data Mining, Inference, and Prediction; Springer: New York, NY, USA, 2009; Volume 2. [Google Scholar]
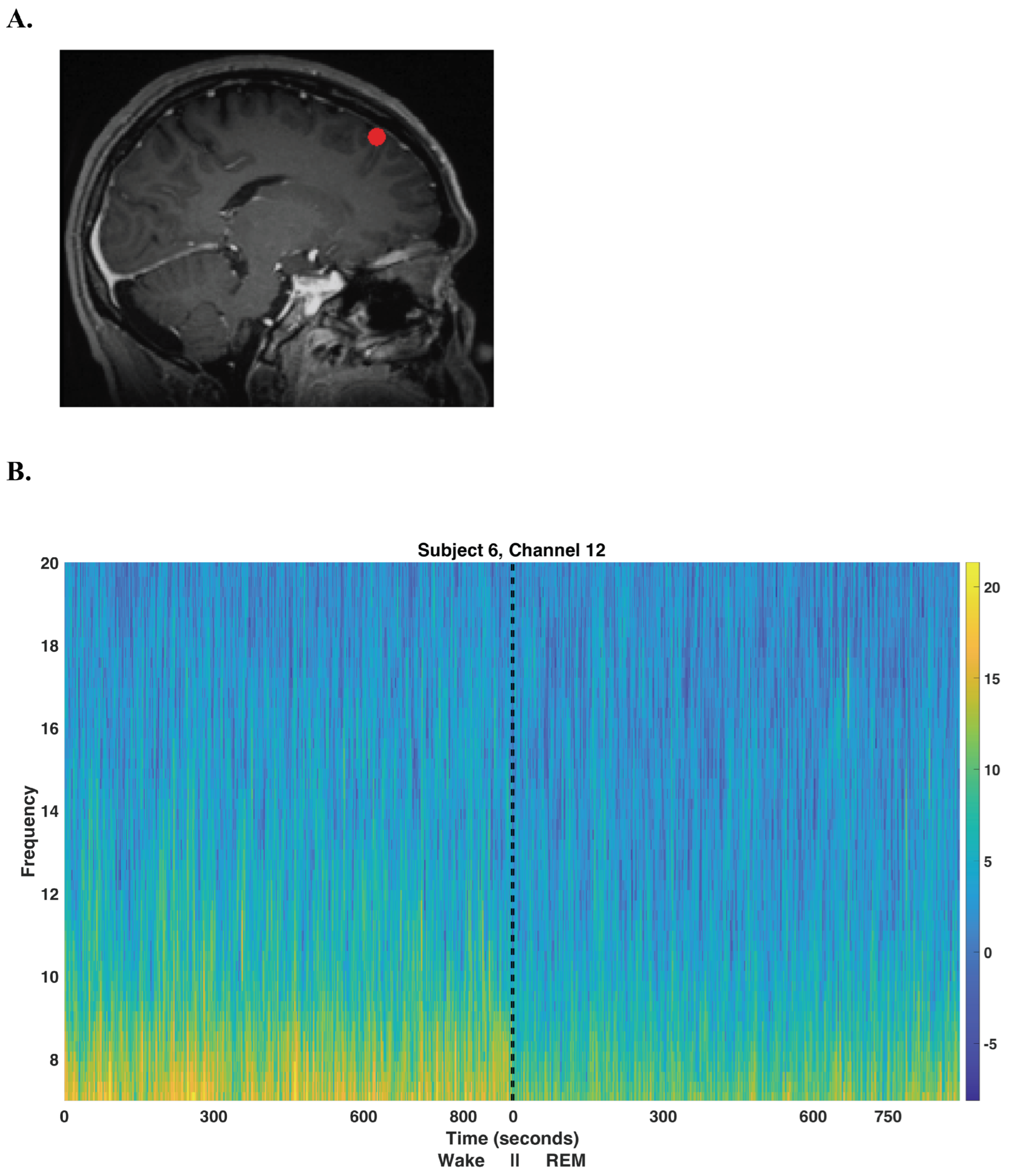
| Subject | Gender | Age | Handedness | Diagnosis | Imaging |
|---|---|---|---|---|---|
| 1 | F | 24 | R | Focal epilepsy, left and right temporal lobe | PET: Bilateral medial temporal hypometabolism. MRI: Normal |
| 2 | F | 41 | R | Focal epilepsy, left temporal lobe | PET: Borderline symmetric hypometabolism involving mesial temporal lobes. MRI: Normal |
| 3 | F | 55 | R | Medial refractory epilepsy with bilateral hippocampal foci | PET: Minimal hypometabolism of left medial and inferior temporal lobe. MRI: Normal |
| 4 | F | 52 | R | Intractable epilepsy, bilateral hippocampal | PET: Hypometabolism left para-hippocampal gyrus. MRI: Bilateral Hippocampal Cyst, 1-2 mm. Infarct, superior right cerebellum |
| 5 | M | 29 | R | Epilepsy of bilateral temporal origin. | PET: Hypometabolism bilaterally in the medial temporal lobes. MRI: Normal |
| 6 | M | 19 | L | Seizures of right temporal origin | PET: Hypometabolism in the right temporal lobe relative to left temporal lobe. MRI: Normal |
| 7 | F | 20 | R | Refractory epilepsy, left frontotemporal | PET: Small area of non-significant hypometabolism in left mesiotemporal region. MRI: Normal |
| 8 | M | 38 | R | Medically intractable epilepsy, left hemisphere | PET: Subtle hypometabolism predominantly in right parietal lobe. MRI: Normal |
| 9 | F | 20 | R | Medical refractory epilepsy with bilateral hippocampal foci | PET: Minimal hypometabolism of right lateral temporal lobe.MRI: Unremarkable (incidental finding of bilateral hippocampal cysts) |
| Subject Number | No. of Electrodes Implanted | No. of Electrodes Implanted, by Location | |||
|---|---|---|---|---|---|
| Left Hemisphere | Right Hemisphere | ||||
| Frontal | Temporal | Frontal | Temporal | ||
| 1 | 16 | 4 | 4 | 4 | 4 |
| 2 | 16 | 4 | 4 | 4 | 4 |
| 3 | 16 | 4 | 4 | 4 | 4 |
| 4 | 12 | 3 | 3 | 3 | 3 |
| 5 | 14 | 3 | 4 | 3 | 4 |
| 6 | 11 | 3 | 2 | 3 | 3 |
| 7 | 2 | 0 | 2 | 0 | 0 |
| 8 | 4 | 0 | 2 | 0 | 2 |
| 9 | 12 | 3 | 3 | 3 | 3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lepage, K.Q.; Jain, S.; Kvavilashvili, A.; Witcher, M.; Vijayan, S. Unsupervised Multitaper Spectral Method for Identifying REM Sleep in Intracranial EEG Recordings Lacking EOG/EMG Data. Bioengineering 2023, 10, 1009. https://doi.org/10.3390/bioengineering10091009
Lepage KQ, Jain S, Kvavilashvili A, Witcher M, Vijayan S. Unsupervised Multitaper Spectral Method for Identifying REM Sleep in Intracranial EEG Recordings Lacking EOG/EMG Data. Bioengineering. 2023; 10(9):1009. https://doi.org/10.3390/bioengineering10091009
Chicago/Turabian StyleLepage, Kyle Q., Sparsh Jain, Andrew Kvavilashvili, Mark Witcher, and Sujith Vijayan. 2023. "Unsupervised Multitaper Spectral Method for Identifying REM Sleep in Intracranial EEG Recordings Lacking EOG/EMG Data" Bioengineering 10, no. 9: 1009. https://doi.org/10.3390/bioengineering10091009
APA StyleLepage, K. Q., Jain, S., Kvavilashvili, A., Witcher, M., & Vijayan, S. (2023). Unsupervised Multitaper Spectral Method for Identifying REM Sleep in Intracranial EEG Recordings Lacking EOG/EMG Data. Bioengineering, 10(9), 1009. https://doi.org/10.3390/bioengineering10091009