

Heart Rate Variability Code: Does It Exist and Can We Hack It?


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction

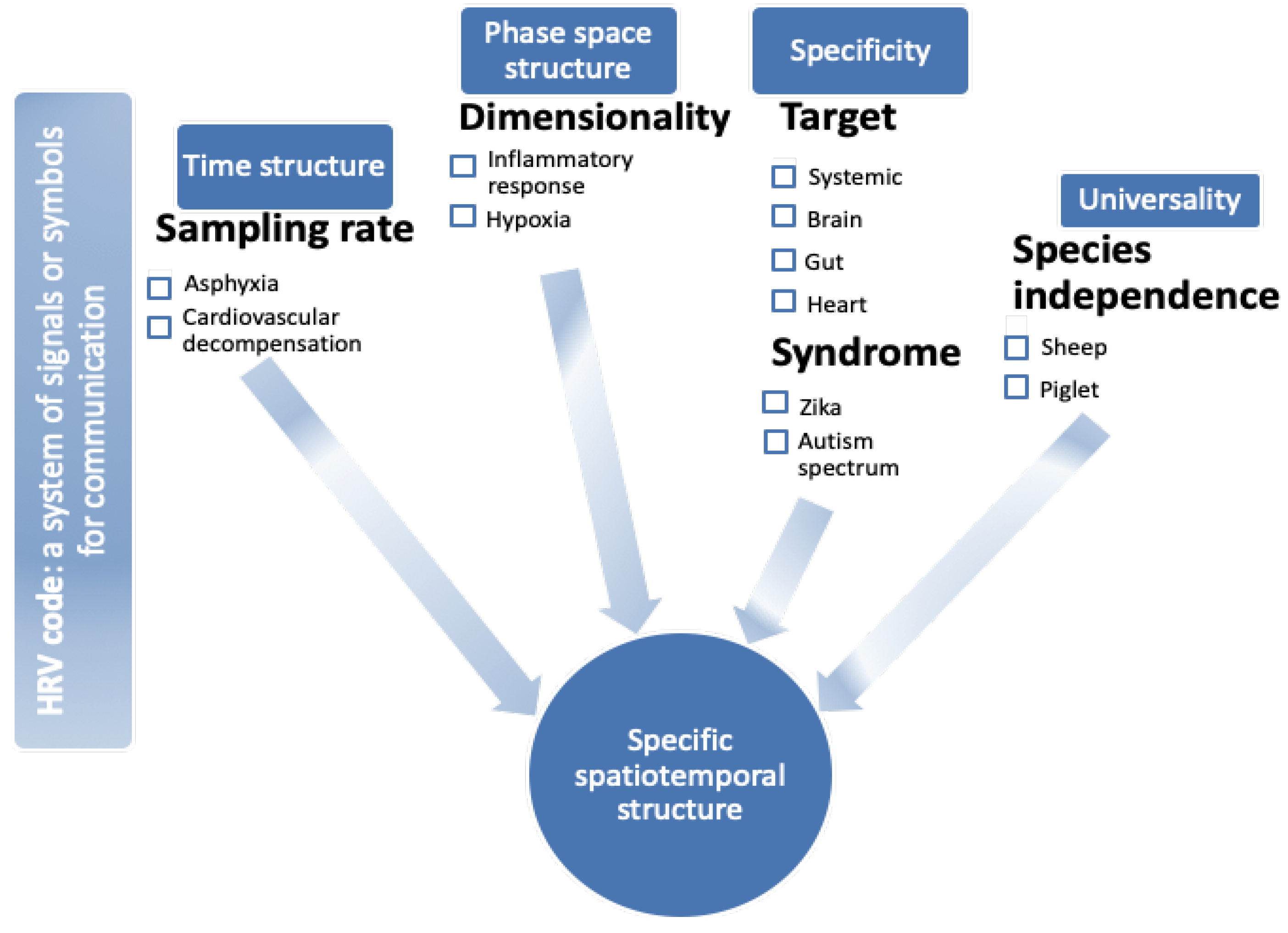
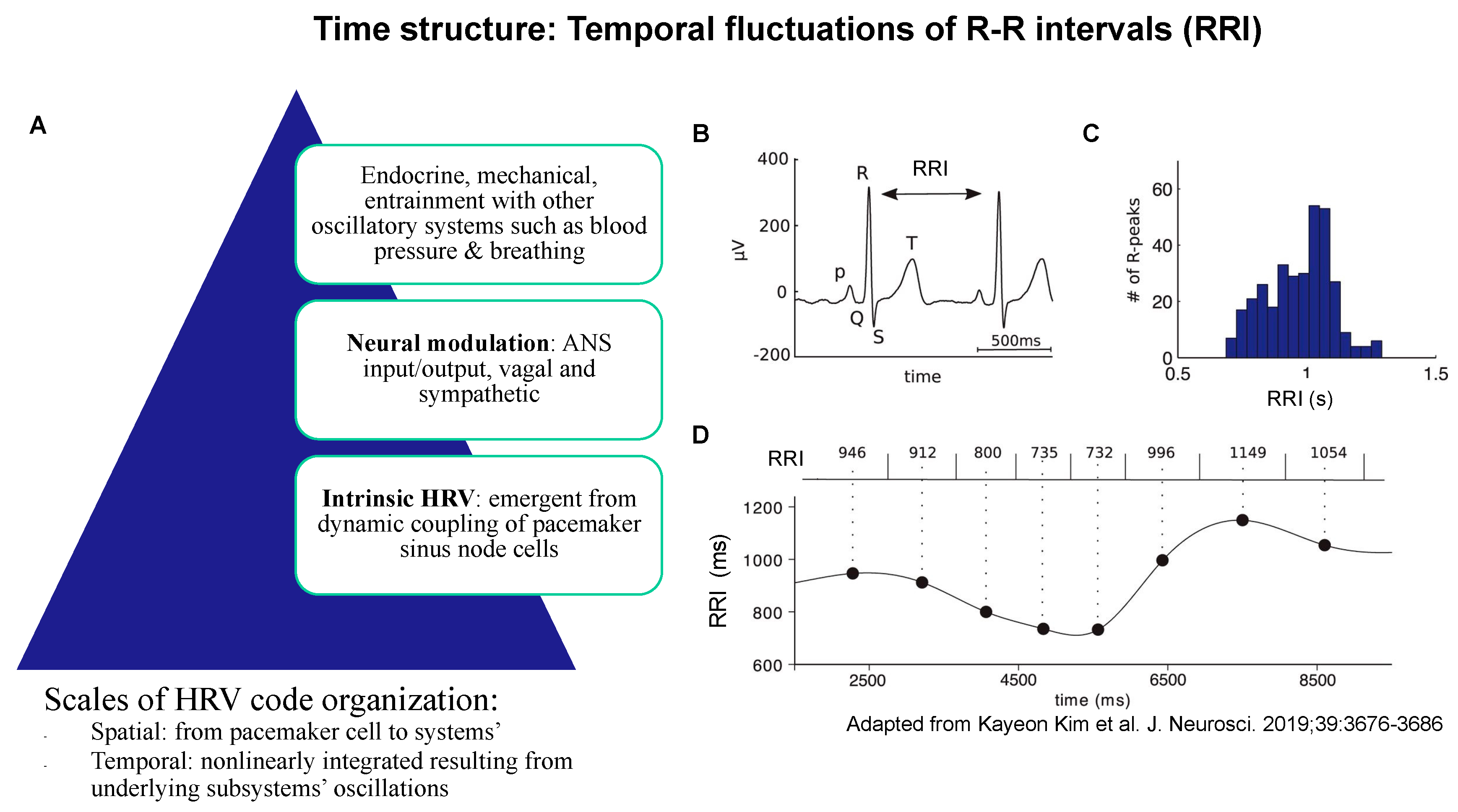
2. Time Structure: Sampling Rate
2.1. Evidence
2.2. Significance
2.3. Future Studies
3. Phase Space Structure: Dimensionality of HRV
3.1. Evidence
3.2. Significance
3.3. Future Studies
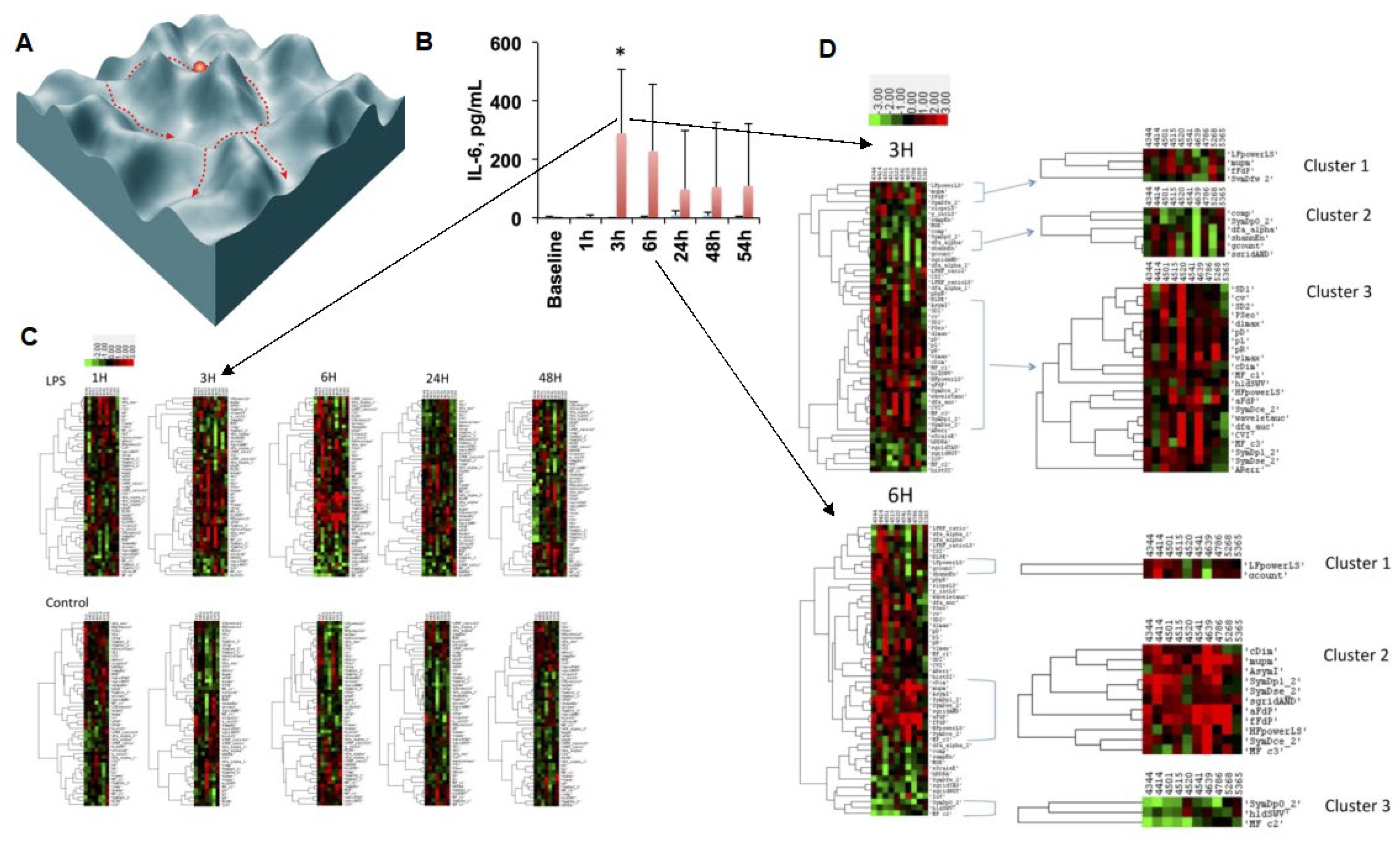
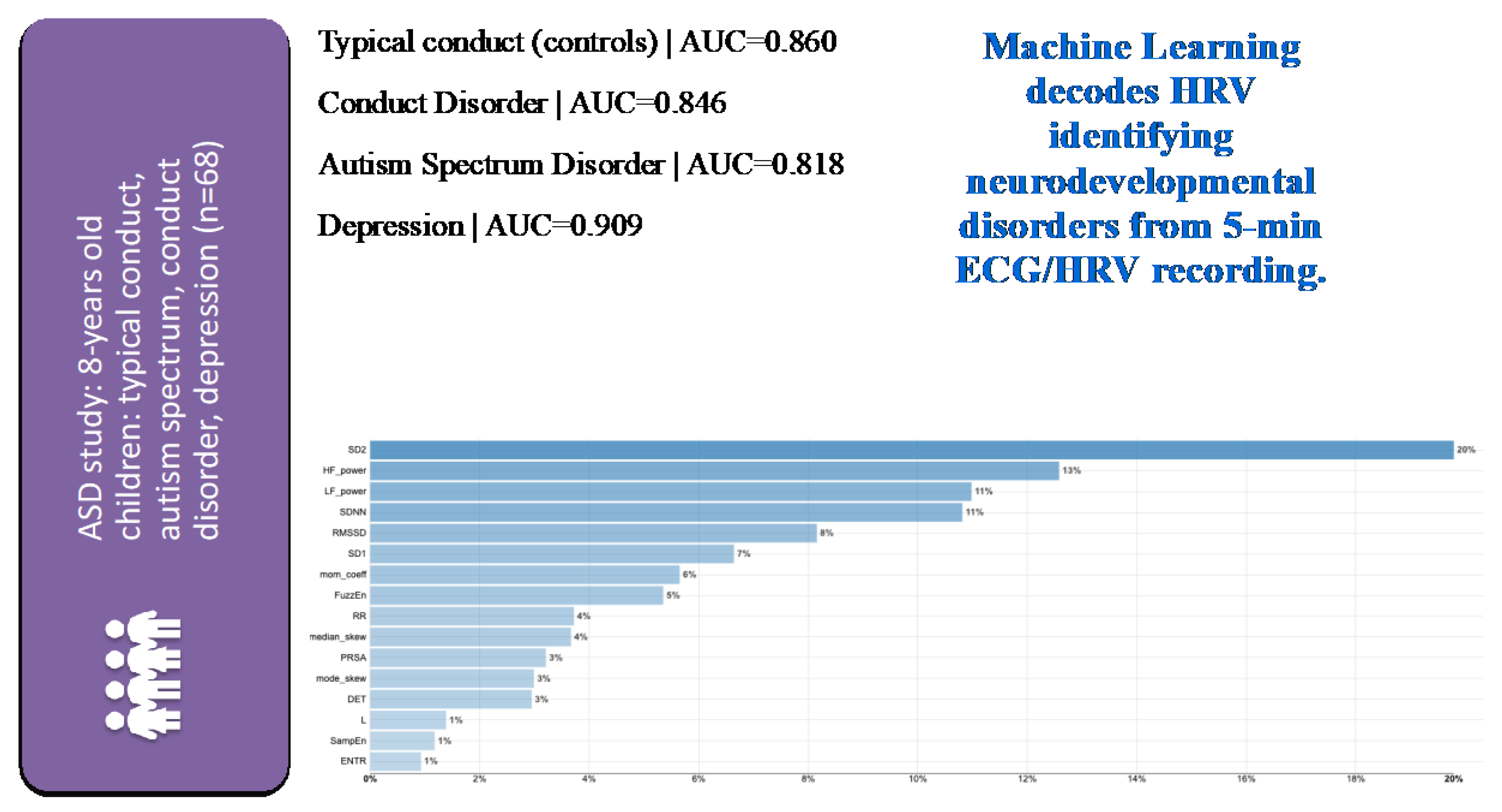
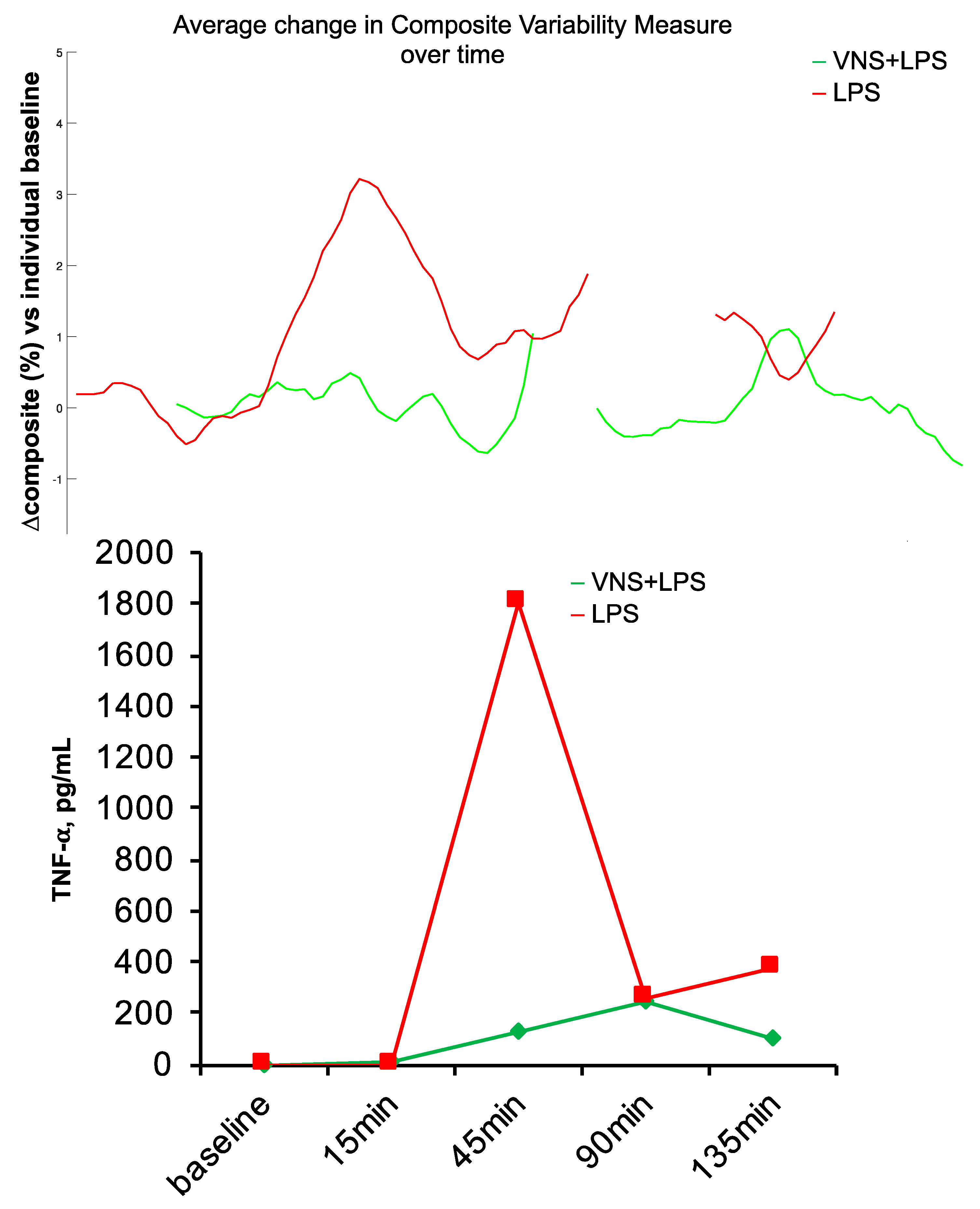
4. Target and Syndrome Specificity
4.1. Evidence
4.2. Significance
4.3. Future Studies
5. Universality
5.1. Evidence
5.2. Significance
5.3. Future Studies
6. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Goldberger, A.L.; Peng, C.-K.; Lipsitz, L.A. What Is Physiologic Complexity and How Does It Change with Aging and Disease? Neurobiol. Aging 2002, 23, 23–26. [Google Scholar] [CrossRef]
- Frasch, M.G. Letter to the Editor: Mind the gap: Epistemology of heart rate variability. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2020, 319, R343–R344. [Google Scholar] [CrossRef] [PubMed]
- Porges, S.W. The polyvagal theory: New insights into adaptive reactions of the autonomic nervous system. Clevel. Clin. J. Med. 2009, 76 (Suppl. S2), S86–S90. [Google Scholar] [CrossRef] [PubMed]
- Thayer, J.F.; Lane, R.D. Claude Bernard and the heart–brain connection: Further elaboration of a model of neurovisceral integration. Neurosci. Biobehav. Rev. 2009, 33, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Bravi, A.; Longtin, A.; Seely, A.J. Review and classification of variability analysis techniques with clinical applications. Biomed. Eng. Online 2011, 10, 90. [Google Scholar] [CrossRef]
- Heart rate variability: Standards of measurement, physiological interpretation and clinical use. Task Force of the European Society of Cardiology and the North American Society of Pacing and Electrophysiology. Circulation 1996, 93, 1043–1065. [CrossRef]
- Herry, C.L.; Burns, P.; Desrochers, A.; Fecteau, G.; Durosier, L.D.; Cao, M.; Seely, A.J.E.; Frasch, M.G. Vagal contributions to fetal heart rate variability: An omics approach. Physiol. Meas. 2019, 40, 065004. [Google Scholar] [CrossRef]
- Herry, C.L.; Cortes, M.; Wu, H.-T.; Durosier, L.D.; Cao, M.; Burns, P.; Desrochers, A.; Fecteau, G.; Seely, A.J.E.; Frasch, M.G. Temporal Patterns in Sheep Fetal Heart Rate Variability Correlate to Systemic Cytokine Inflammatory Response: A Methodological Exploration of Monitoring Potential Using Complex Signals Bioinformatics. PLoS ONE 2016, 11, e0153515. [Google Scholar] [CrossRef]
- Lake, D.E.; Fairchild, K.D.; Moorman, J.R. Complex signals bioinformatics: Evaluation of heart rate characteristics monitoring as a novel risk marker for neonatal sepsis. J. Clin. Monit. Comput. 2014, 28, 329–339. [Google Scholar] [CrossRef]
- Seely, A.J.E.; Green, G.C.; Bravi, A. Continuous Multiorgan Variability monitoring in critically ill patients—Complexity science at the bedside. Conf. Proc. IEEE Eng. Med. Biol. Soc. 2011, 2011, 5503–5506. [Google Scholar] [CrossRef]
- Frasch, M.G.; Lobmaier, S.M.; Stampalija, T.; Desplats, P.; Pallarés, M.E.; Pastor, V.; Brocco, M.A.; Wu, H.-T.; Schulkin, J.; Herry, C.L.; et al. Non-invasive biomarkers of fetal brain development reflecting prenatal stress: An integrative multi-scale multi-species perspective on data collection and analysis. Neurosci. Biobehav. Rev. 2020, 117, 165–183. [Google Scholar] [CrossRef] [PubMed]
- Frasch, M.G.; Müller, T.; Wicher, C.; Weiss, C.; Löhle, M.; Schwab, K.; Schubert, H.; Nathanielsz, P.W.; Witte, O.W.; Schwab, M. Fetal body weight and the development of the control of the cardiovascular system in fetal sheep. J. Physiol. 2007, 579, 893–907. [Google Scholar] [CrossRef]
- Morrison, J.L.; Berry, M.J.; Botting, K.J.; Darby, J.R.T.; Frasch, M.G.; Gatford, K.L.; Giussani, D.A.; Gray, C.L.; Harding, R.; Herrera, E.A.; et al. Improving pregnancy outcomes in humans through studies in sheep. Am. J. Physiol. Integr. Comp. Physiol. 2018, 315, R1123–R1153. [Google Scholar] [CrossRef]
- Wikipedia Contributors Code. Available online: https://en.wikipedia.org/w/index.php?title=Code&oldid=934204133 (accessed on 9 January 2020).
- Durosier, L.D.; Egreen, G.; Batkin, I.; Seely, A.J.; Ross, M.G.; Richardson, B.S.; Frasch, M.G. Sampling rate of heart rate variability impacts the ability to detect acidemia in ovine fetuses near-term. Front. Pediatr. 2014, 2, 38. [Google Scholar] [CrossRef]
- Li, X.; Xu, Y.; Herry, C.; Durosier, L.D.; Casati, D.; Stampalija, T.; Maisonneuve, E.; Seely, A.J.E.; Audibert, F.; Alfirevic, Z.; et al. Sampling frequency of fetal heart rate impacts the ability to predict pH and BE at birth: A retrospective multi-cohort study. Physiol. Meas. 2015, 36, L1–L12. [Google Scholar] [CrossRef]
- Gold, N.; Herry, C.L.; Wang, X.; Frasch, M.G. Fetal Cardiovascular Decompensation during Labor Predicted from the Individual Heart Rate: A Prospective Study in Fetal Sheep near Term and the Impact of Low Sampling Rate. arXiv 2019, arXiv:1911.01304. [Google Scholar]
- Gold, N.; Herry, C.L.; Wang, X.; Frasch, M.G. Fetal Cardiovascular Decompensation During Labor Predicted from the Individual Heart Rate Tracing: A Machine Learning Approach in Near-Term Fetal Sheep Model. Front. Pediatr. 2021, 9, 593889. [Google Scholar] [CrossRef]
- Gold, N.; Frasch, M.G.; Herry, C.L.; Richardson, B.S.; Wang, X. A Doubly Stochastic Change Point Detection Algorithm for Noisy Biological Signals. Front. Physiol. 2017, 8, 1112. [Google Scholar] [CrossRef]
- Roux, S.G.; Garnier, N.B.; Abry, P.; Gold, N.; Frasch, M.G. Distance to Healthy Metabolic and Cardiovascular Dynamics From Fetal Heart Rate Scale-Dependent Features in Pregnant Sheep Model of Human Labor Predicts the Evolution of Acidemia and Cardiovascular Decompensation. Front. Pediatr. 2021, 9, 660476. [Google Scholar] [CrossRef]
- Frasch, M.G. Fetal heart rate variability: An ocean of meanings beyond ups and downs. BJOG 2023. [Google Scholar] [CrossRef] [PubMed]
- Masi, E.B.; Levy, T.; Tsaava, T.; Bouton, C.E.; Tracey, K.J.; Chavan, S.S.; Zanos, T.P. Identification of hypoglycemia-specific neural signals by decoding murine vagus nerve activity. Bioelectron. Med. 2019, 5, 9. [Google Scholar] [CrossRef] [PubMed]
- Cotero, V.; Fan, Y.; Tsaava, T.; Kressel, A.M.; Hancu, I.; Fitzgerald, P.; Wallace, K.; Kaanumalle, S.; Graf, J.; Rigby, W.; et al. Noninvasive sub-organ ultrasound stimulation for targeted neuromodulation. Nat. Commun. 2019, 10, 952. [Google Scholar] [CrossRef] [PubMed]
- Cao, M.; Kuthiala, S.; Shafiee, T.; Liu, H.L.; Burns, P.; Desrochers, A.; Fecteau, G.; Frasch, M.G. Role of Vagus Nerve in Fetal Glucosensing and Neuroinflammation. In Proceedings of the Reproductive Sciences; Sage Publications Inc.: Thousand Oaks, CA, USA, 2015; Volume 22, p. 367A. [Google Scholar]
- Cao, M.; Kuthiala, S.; Jean, K.J.; Liu, H.L.; Courchesne, M.; Nygard, K.; Burns, P.; Desrochers, A.; Fecteau, G.; Faure, C.; et al. The Vagus Nerve Regulates Immunometabolic Homeostasis in the Ovine Fetus near Term: Impact on Terminal Ileum. arXiv 2022, arXiv:2203.14967. [Google Scholar]
- Durosier, L.D.; Herry, C.L.; Cortes, M.; Cao, M.; Burns, P.; Desrochers, A.; Fecteau, G.; Seely, A.J.E.; Frasch, M.G. Does heart rate variability reflect the systemic inflammatory response in a fetal sheep model of lipopolysaccharide-induced sepsis? Physiol. Meas. 2015, 36, 2089–2102. [Google Scholar] [CrossRef] [PubMed]
- Berthoud, H.-R.; Neuhuber, W.L. Functional and chemical anatomy of the afferent vagal system. Auton. Neurosci. 2000, 85, 1–17. [Google Scholar] [CrossRef]
- Lanza, G.A.; Sgueglia, G.A.; Cianflone, D.; Rebuzzi, A.G.; Angeloni, G.; Sestito, A.; Infusino, F.; Crea, F.; Maseri, A. Relation of Heart Rate Variability to Serum Levels of C-Reactive Protein in Patients with Unstable Angina Pectoris. Am. J. Cardiol. 2006, 97, 1702–1706. [Google Scholar] [CrossRef]
- Frasch, M.G.; Szynkaruk, M.; Prout, A.P.; Nygard, K.; Cao, M.; Veldhuizen, R.; Hammond, R.; Richardson, B.S. Decreased neuroinflammation correlates to higher vagus nerve activity fluctuations in near-term ovine fetuses: A case for the afferent cholinergic anti-inflammatory pathway? J. Neuroinflamm. 2016, 13, 103. [Google Scholar] [CrossRef]
- Frasch, M.G. Comprehensive HRV estimation pipeline in Python using Neurokit2: Application to sleep physiology. Methodsx 2022, 9, 101782. [Google Scholar] [CrossRef]
- Herry, C.L.; Schüler-Faccini, L.; Silva, H.M.; Barbosa, C.P.; Ecco, G.; Frasch, M.G. #29 Heart rate variability monitoring identifies asymptomatic toddlers exposed to Zika virus during pregnancy. Physiol. Meas. 2021, 17, 42. [Google Scholar] [CrossRef]
- Haensel, A.; Mills, P.J.; Nelesen, R.A.; Ziegler, M.G.; Dimsdale, J.E. The relationship between heart rate variability and inflammatory markers in cardiovascular diseases. Psychoneuroendocrinology 2008, 33, 1305–1312. [Google Scholar] [CrossRef]
- Frasch, M.G.; Shen, C.; Wu, H.-T.; Mueller, A.; Bernier, R.A.; Beauchaine, T.P.; Neuhaus, E. Can a Heart Rate Variability Biomarker Identify the Presence of Autism Spectrum Disorder in Eight Year Old Children? arXiv 2018, arXiv:1808.08306. [Google Scholar]
- Patriquin, M.A.; Hartwig, E.M.; Friedman, B.H.; Porges, S.W.; Scarpa, A. Autonomic response in autism spectrum disorder: Relationship to social and cognitive functioning. Biol. Psychol. 2019, 145, 185–197. [Google Scholar] [CrossRef]
- Frasch, M. Sampling Rate and Heart Rate Variability: On Metrics and Health Outcomes. J. Biomed. Inform. 2022, 129, 104061. [Google Scholar] [CrossRef] [PubMed]
- Frasch, M.G.; Mansano, R.; Ross, M.G.; Gagnon, R.; Richardson, B.S. Do Repetitive Umbilical Cord Occlusions (UCO) with Worsening Acidemia Induce the Bezold-Jarisch Reflex (BJR) in the Ovine Fetus near Term? Reprod. Sci. 2008, 15, 129A. [Google Scholar] [CrossRef]
- Frasch, M.G. Saving the brain one heartbeat at a time. J. Physiol. 2018, 596, 5503–5504. [Google Scholar] [CrossRef]
- Garg, V.K.; Wang, Y.-C. 5-Data Communication Concepts. In The Electrical Engineering Handbook; Chen, W.-K., Ed.; Academic Press: Burlington, VT, USA, 2005; pp. 983–988. ISBN 9780121709600. [Google Scholar]
- Kim, K.; Ladenbauer, J.; Babo-Rebelo, M.; Buot, A.; Lehongre, K.; Adam, C.; Hasboun, D.; Lambrecq, V.; Navarro, V.; Ostojic, S.; et al. Resting-State Neural Firing Rate Is Linked to Cardiac-Cycle Duration in the Human Cingulate and Parahippocampal Cortices. J. Neurosci. 2019, 39, 3679–3686. [Google Scholar] [CrossRef]
- Burma, J.S.; Lapointe, A.P.; Soroush, A.; Oni, I.K.; Smirl, J.D.; Dunn, J.F. Insufficient sampling frequencies skew heart rate variability estimates: Implications for extracting heart rate metrics from neuroimaging and physiological data. J. Biomed. Inform. 2021, 123, 103934. [Google Scholar] [CrossRef]
- Lin, Y.-T.; Lo, Y.-L.; Lin, C.-Y.; Frasch, M.G.; Wu, H.-T. Unexpected sawtooth artifact in beat-to-beat pulse transit time measured from patient monitor data. PLoS ONE 2019, 14, e0221319. [Google Scholar] [CrossRef] [PubMed]
- Quer, G.; Gadaleta, M.; Radin, J.M.; Andersen, K.G.; Baca-Motes, K.; Ramos, E.; Topol, E.J.; Steinhubl, S.R. Inter-Individual Variation in Objective Measure of Reactogenicity Following COVID-19 Vaccination via Smartwatches and Fitness Bands. NPJ Digit. Med. 2022, 5, 49. [Google Scholar] [CrossRef]
- Quer, G.; Radin, J.M.; Gadaleta, M.; Baca-Motes, K.; Ariniello, L.; Ramos, E.; Kheterpal, V.; Topol, E.J.; Steinhubl, S.R. Wearable sensor data and self-reported symptoms for COVID-19 detection. Nat. Med. 2020, 27, 73–77. [Google Scholar] [CrossRef]
- Radin, J.M.; Wineinger, N.E.; Topol, E.J.; Steinhubl, S.R. Harnessing wearable device data to improve state-level real-time surveillance of influenza-like illness in the USA: A population-based study. Lancet Digit. Health 2020, 2, e85–e93. [Google Scholar] [CrossRef]
- Nolte, D.D. The tangled tale of phase space. Phys. Today 2010, 63, 33–38. [Google Scholar] [CrossRef]
- MacArthur, B.D.; Lemischka, I.R. Statistical mechanics of pluripotency. Cell 2013, 154, 484–489. [Google Scholar] [CrossRef]
- Luyer, M.D.; Greve, J.W.M.; Hadfoune, M.; Jacobs, J.A.; Dejong, C.H.; Buurman, W.A. Nutritional stimulation of cholecystokinin receptors inhibits inflammation via the vagus nerve. J. Exp. Med. 2005, 202, 1023–1029. [Google Scholar] [CrossRef]
- Brinkman, D.J.; Hove, A.S.T.; Vervoordeldonk, M.J.; Luyer, M.D.; de Jonge, W.J. Neuroimmune Interactions in the Gut and Their Significance for Intestinal Immunity. Cells 2019, 8, 670. [Google Scholar] [CrossRef]
- Liu, H.L.; Garzoni, L.; Herry, C.; Durosier, L.D.; Cao, M.; Burns, P.; Fecteau, G.; Desrochers, A.; Patey, N.; Seely, A.J.E.; et al. Can Monitoring Fetal Intestinal Inflammation Using Heart Rate Variability Analysis Signal Incipient Necrotizing Enterocolitis of the Neonate? Pediatr. Crit. Care Med. 2016, 17, e165–e176. [Google Scholar] [CrossRef]
- Frasch, M.G.; Herry, C.L.; Niu, Y.; Giussani, D.A. First evidence that intrinsic fetal heart rate variability exists and is affected by hypoxic pregnancy. J. Physiol. 2020, 598, 249–263. [Google Scholar] [CrossRef]
- Frasch, M.G.; Herry, C.L.; Niu, Y.; Giussani, D.A. First Evidence of Intrinsic Fetal Heart Rate Variability Affected by Chronic Fetal Hypoxia. In Proceedings of the Reproductive Sciences; Sage Publications Inc.: Thousand Oaks, CA, USA, 2017; Volume 24, p. 198A. [Google Scholar]
- Metz, C.N.; Pavlov, V.A. Vagus nerve cholinergic circuitry to the liver and the gastrointestinal tract in the neuroimmune communicatome. Am. J. Physiol. Liver Physiol. 2018, 315, G651–G658. [Google Scholar] [CrossRef]
- Rosas-Ballina, M.; Olofsson, P.S.; Ochani, M.; Valdés-Ferrer, S.I.; Levine, Y.A.; Reardon, C.; Tusche, M.W.; Pavlov, V.A.; Andersson, U.; Chavan, S.; et al. Acetylcholine-Synthesizing T Cells Relay Neural Signals in a Vagus Nerve Circuit. Science 2011, 334, 98–101. [Google Scholar] [CrossRef] [PubMed]
- Diamond, B.; Tracey, K.J. Mapping the immunological homunculus. Proc. Natl. Acad. Sci. USA 2011, 108, 3461–3462. [Google Scholar] [CrossRef] [PubMed]
- Tavakol, S.; Kwan, H.; Jean, K.; Cao, M.; Burns, P.; Desrochers, A.; Fecteau, G.; Frasch, M.G. Fetal Vagal Cholinergic Activity Modulates Inflammatory and Cardiovascular Responses to Low-Dose Endotoxin Exposure. Reprod. Sci. 2016, 23, 228A. [Google Scholar]
- Conway, C.R.; Sheline, Y.I.; Chibnall, J.T.; George, M.S.; Fletcher, J.W.; Mintun, M.A. Cerebral blood flow changes during vagus nerve stimulation for depression. Psychiatry Res. Neuroimaging 2006, 146, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Tracey, K.J. Physiology and immunology of the cholinergic antiinflammatory pathway. J. Clin. Investig. 2007, 117, 289–296. [Google Scholar] [CrossRef]
- Olofsson, P.S.; Rosas-Ballina, M.; Levine, Y.A.; Tracey, K.J. Rethinking inflammation: Neural circuits in the regulation of immunity. Immunol. Rev. 2012, 248, 188–204. [Google Scholar] [CrossRef] [PubMed]
- von Känel, R.; Nelesen, R.A.; Mills, P.J.; Ziegler, M.G.; Dimsdale, J.E. Relationship between heart rate variability, interleukin-6, and soluble tissue factor in healthy subjects. Brain Behav. Immun. 2008, 22, 461–468. [Google Scholar] [CrossRef]
- Hamaad, A.; Sosin, M.; Blann, A.D.; Patel, J.; Lip, G.Y.H.; MacFadyen, R.J. Markers of inflammation in acute coronary syndromes: Association with increased heart rate and reductions in heart rate variability. Clin. Cardiol. 2005, 28, 570–576. [Google Scholar] [CrossRef] [PubMed]
- Castel, A.; Burns, P.; Wakefield, C.; Jean, K.J.; Frank, Y.S.; Cao, M.; Desrochers, A.; Fecteau, G.; Faure, C.; Herry, C.L.; et al. Perinatal Physiology. In Vagus Nerve Stimulation; Frasch, M.G., Porges, E., Eds.; Springer Nature: Berlin/Heidelberg, Germany, 2023. [Google Scholar]
- Magawa, S.; Lear, C.A.; Beacom, M.J.; King, V.J.; Kasai, M.; Galinsky, R.; Ikeda, T.; Gunn, A.J.; Bennet, L. Fetal heart rate variability is a biomarker of rapid but not progressive exacerbation of inflammation in preterm fetal sheep. Sci. Rep. 2022, 12, 1771. [Google Scholar] [CrossRef]
- Frasch, M.G. Heart Rate as a Non-Invasive Biomarker of Inflammation: Implications for Digital Health. Front. Immunol. 2022, 13, 930445. [Google Scholar] [CrossRef] [PubMed]
- Huysmans, D.; Borzée, P.; Testelmans, D.; Buyse, B.; Willemen, T.; Van Huffel, S.; Varon, C. Evaluation of a Commercial Ballistocardiography Sensor for Sleep Apnea Screening and Sleep Monitoring. Sensors 2019, 19, 2133. [Google Scholar] [CrossRef]
- Vesterinen, V.; Rinkinen, N.; Nummela, A. A Contact-Free, Ballistocardiography-Based Monitoring System (Emfit QS) for Measuring Nocturnal Heart Rate and Heart Rate Variability: Validation Study. JMIR Biomed. Eng. 2020, 5, e16620. [Google Scholar] [CrossRef]
- Diraco, G.; Leone, A.; Siciliano, P. A Radar-Based Smart Sensor for Unobtrusive Elderly Monitoring in Ambient Assisted Living Applications. Biosensors 2017, 7, 55. [Google Scholar] [CrossRef]
- Islam, S.M.M. Radar-Based Remote Physiological Sensing: Progress, Challenges, and Opportunities. Front. Physiol. 2022, 13, 955208. [Google Scholar] [CrossRef]
- Khan, F.; Azou, S.; Youssef, R.; Morel, P.; Radoi, E. IR-UWB Radar-Based Robust Heart Rate Detection Using a Deep Learning Technique Intended for Vehicular Applications. Electronics 2022, 11, 2505. [Google Scholar] [CrossRef]
- Nosrati, M.; Tavassolian, N. High-Accuracy Heart Rate Variability Monitoring Using Doppler Radar Based on Gaussian Pulse Train Modeling and FTPR Algorithm. IEEE Trans. Microw. Theory Tech. 2018, 66, 556–567. [Google Scholar] [CrossRef]
- Will, C.; Shi, K.; Schellenberger, S.; Steigleder, T.; Michler, F.; Fuchs, J.; Weigel, R.; Ostgathe, C.; Koelpin, A. Radar-Based Heart Sound Detection. Sci. Rep. 2018, 8, 11551. [Google Scholar] [CrossRef] [PubMed]
- Tsaava, T.; Datta-Chaudhuri, T.; Addorisio, M.E.; Masi, E.B.; Silverman, H.A.; Newman, J.E.; Imperato, G.H.; Bouton, C.; Tracey, K.J.; Chavan, S.S.; et al. Serum Cytokine Levels Are Modulated by Specific Frequencies, Amplitudes, and Pulse Widths of Vagus Nerve Stimulation. bioRxiv 2020. [Google Scholar] [CrossRef]
- Zanos, T.P.; Silverman, H.A.; Levy, T.; Tsaava, T.; Battinelli, E.; Lorraine, P.W.; Ashe, J.M.; Chavan, S.S.; Tracey, K.J.; Bouton, C.E. Identification of cytokine-specific sensory neural signals by decoding murine vagus nerve activity. Proc. Natl. Acad. Sci. USA 2018, 115, E4843–E4852. [Google Scholar] [CrossRef] [PubMed]
- Hirsch, A.J.; Roberts, V.H.J.; Grigsby, P.L.; Haese, N.; Schabel, M.C.; Wang, X.; Lo, J.O.; Liu, Z.; Kroenke, C.D.; Smith, J.L.; et al. Zika virus infection in pregnant rhesus macaques causes placental dysfunction and immunopathology. Nat. Commun. 2018, 9, 263. [Google Scholar] [CrossRef]
- Beauchaine, T. Vagal Tone, Development, and Gray’s Motivational Theory: Toward an Integrated Model of Autonomic Nervous System Functioning in Psychopathology. Dev. Psychopathol. 2001, 13, 183–214. [Google Scholar] [CrossRef]
- Frasch, M.G.; Yoon, B.-J.; Helbing, D.L.; Snir, G.; Antonelli, M.C.; Bauer, R. Autism Spectrum Disorder: A Neuro-Immunometabolic Hypothesis of the Developmental Origins. Biology 2023, 12, 914. [Google Scholar] [CrossRef]
- Weitz, G.; Bonnemeier, H.; Süfke, S.; Wellhöner, P.; Lehnert, H.; Dodt, C. Heart rate variability and metabolic rate in healthy young adults with low birth weight. Am. J. Cardiovasc. Dis. 2013, 3, 239–246. [Google Scholar]
- Woods, A.L.; Rice, A.J.; Garvican-Lewis, L.A.; Wallett, A.M.; Lundy, B.; Rogers, M.A.; Welvaert, M.; Halson, S.; McKune, A.; Thompson, K.G. The effects of intensified training on resting metabolic rate (RMR), body composition and performance in trained cyclists. PLoS ONE 2018, 13, e0191644. [Google Scholar] [CrossRef]
- Carrillo, A.E.; Flouris, A.D.; Herry, C.L.; Notley, S.R.; Macartney, M.J.; Seely, A.J.; Beatty, H.E.W.; Kenny, G.P. Age-related reductions in heart rate variability do not worsen during exposure to humid compared to dry heat: A secondary analysis. Temperature 2019, 6, 341–345. [Google Scholar] [CrossRef]
- Kaltsatou, A.; Flouris, A.D.; Herry, C.L.; Notley, S.R.; Seely, A.J.E.; Beatty, H.W.; Kenny, G.P. Age differences in cardiac autonomic regulation during intermittent exercise in the heat. Eur. J. Appl. Physiol. 2020, 120, 453–465. [Google Scholar] [CrossRef]
- Flouris, A.D.; Bravi, A.; Wright-Beatty, H.E.; Green, G.; Seely, A.J.; Kenny, G.P. Heart rate variability during exertional heat stress: Effects of heat production and treatment. Eur. J. Appl. Physiol. 2014, 114, 785–792. [Google Scholar] [CrossRef]
- Park, H.; Dong, S.-Y.; Lee, M.; Youn, I. The Role of Heart-Rate Variability Parameters in Activity Recognition and Energy-Expenditure Estimation Using Wearable Sensors. Sensors 2017, 17, 1698. [Google Scholar] [CrossRef] [PubMed]
- Robertson, A.H.; King, K.; Ritchie, S.D.; Gauthier, A.P.; Laurence, M.; Dorman, S.C. Validating the Use of Heart Rate Variability for Estimating Energy Expenditure. Int. J. Hum. Mov. Sports Sci. 2015, 3, 19–26. [Google Scholar] [CrossRef]
- Oishi, K.; Himeno, Y.; Miwa, M.; Anzai, H.; Kitajima, K.; Yasunaka, Y.; Kumagai, H.; Ieiri, S.; Hirooka, H. Correcting the Activity-Specific Component of Heart Rate Variability Using Dynamic Body Acceleration Under Free-Moving Conditions. Front. Physiol. 2018, 9, 1063. [Google Scholar] [CrossRef] [PubMed]
- Kwan, H.; Garzoni, L.; Liu, H.L.; Cao, M.; Desrochers, A.; Fecteau, G.; Burns, P.; Frasch, M.G. Vagus Nerve Stimulation for Treatment of Inflammation: Systematic Review of Animal Models and Clinical Studies. Bioelectron. Med. 2016, 3, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Frank, Y.; Last, M.; Frasch, M.G. Detection and Treatment of Fetal and Newborn Infections via Heart Rate Monitoring and Vagus Nerve Stimulation. In Proceedings of the Reproductive Sciences; Sage Publications Inc.: Thousand Oaks, CA, USA, 2018; Volume 25, pp. 213A–214A. [Google Scholar]
- Burns, P.; Herry, C.L.; Jean, K.J.; Frank, Y.; Wakefield, C.; Cao, M.; Desrochers, A.; Fecteau, G.; Last, M.; Faure, C.; et al. The Neonatal Sepsis Is Diminished by Cervical Vagus Nerve Stimulation and Tracked Non-Invasively by ECG: A Preliminary Report in the Piglet Model. arXiv 2020, arXiv:2002.04136. [Google Scholar]
- Laborde, S.; Mosley, E.; Thayer, J.F. Heart Rate Variability and Cardiac Vagal Tone in Psychophysiological Research—Recommendations for Experiment Planning, Data Analysis, and Data Reporting. Front. Psychol. 2017, 8, 213. [Google Scholar] [CrossRef]
- Frasch, M.G.; Giussani, D.A. Heart during Acidosis: Etiology and Early Detection of Cardiac Dysfunction. EClinicalMedicine 2021, 37, 100994. [Google Scholar] [CrossRef]
- Clifford, G.D. PhysioNet Cardiovascular Signal Toolbox. 2018. Available online: https://physionet.org/content/pcst/1.0.0/ (accessed on 1 January 2020).
- Goldberger, A.L.; Amaral, L.A.; Glass, L.; Hausdorff, J.M.; Ivanov, P.C.; Mark, R.G.; Mietus, J.E.; Moody, G.B.; Peng, C.-K.; Stanley, H.E. PhysioBank, PhysioToolkit, and PhysioNet: Components of a New Research Resource for Complex Physiologic Signals. Circulation 2000, 101, E215–E220. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-C.; Park, Y.; Bin Yoon, S.; Yang, S.M.; Park, D.; Jung, C.-W. VitalDB, a high-fidelity multi-parameter vital signs database in surgical patients. Sci. Data 2022, 9, 279. [Google Scholar] [CrossRef]
- Delude, C.M. Deep phenotyping: The details of disease. Nature 2015, 527, S14–S15. [Google Scholar] [CrossRef] [PubMed]
- McDonnell, M.D.; Abbott, D. What Is Stochastic Resonance? Definitions, Misconceptions, Debates, and Its Relevance to Biology. PLoS Comput. Biol. 2009, 5, e1000348. [Google Scholar] [CrossRef]
- Jerbi, J.K.; Vidal, J.R.; Ossandon, T.; Dalal, S.S.; Jung, J.; Hoffmann, D.; Minotti, L.; Bertrand, O.; Kahane, P.; Lachaux, J.P. Exploring the electrophysiological correlates of the default-mode network with intracerebral EEG. Front. Syst. Neurosci. 2010, 4, 27. [Google Scholar] [CrossRef]
- Al, E.; Iliopoulos, F.; Forschack, N.; Nierhaus, T.; Grund, M.; Motyka, P.; Gaebler, M.; Nikulin, V.V.; Villringer, A. Heart-Brain Interactions Shape Somatosensory Perception and Evoked Potentials. Proc. Natl. Acad. Sci. USA 2020, 117, 10575–10584. [Google Scholar] [CrossRef]
- Galvez-Pol, A.; Virdee, P.; Villacampa, J.; Kilner, J. Active tactile discrimination is coupled with and modulated by the cardiac cycle. eLife 2022, 11, 78126. [Google Scholar] [CrossRef]
- Herman, A.M. In Sync with the Heart. eLife 2022, 11, e84298. [Google Scholar] [CrossRef]
- Tallon-Baudry, C. Interoception: Probing internal state is inherent to perception and cognition. Neuron 2023, 111, 1854–1857. [Google Scholar] [CrossRef]
- Frasch, M.G.; Giussani, D.A. Impact of Chronic Fetal Hypoxia and Inflammation on Cardiac Pacemaker Cell Development. Cells 2020, 9, 733. [Google Scholar] [CrossRef] [PubMed]
- Frasch, M.G. Heart Rate Variability Code: Does It Exist and Can We Hack It? arXiv 2020, arXiv:2001.08264. [Google Scholar]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Frasch, M.G. Heart Rate Variability Code: Does It Exist and Can We Hack It? Bioengineering 2023, 10, 822. https://doi.org/10.3390/bioengineering10070822
Frasch MG. Heart Rate Variability Code: Does It Exist and Can We Hack It? Bioengineering. 2023; 10(7):822. https://doi.org/10.3390/bioengineering10070822
Chicago/Turabian StyleFrasch, Martin Gerbert. 2023. "Heart Rate Variability Code: Does It Exist and Can We Hack It?" Bioengineering 10, no. 7: 822. https://doi.org/10.3390/bioengineering10070822
APA StyleFrasch, M. G. (2023). Heart Rate Variability Code: Does It Exist and Can We Hack It? Bioengineering, 10(7), 822. https://doi.org/10.3390/bioengineering10070822