

Electrospun Poly(L-lactide-co-ε-caprolactone) Scaffold Potentiates C2C12 Myoblast Bioactivity and Acts as a Stimulus for Cell Commitment in Skeletal Muscle Myogenesis

 ,
,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Scaffold Fabrication
2.3. Scaffold Characterization Methods
2.4. Cell Seeding Conditions
2.5. Scaffold Functionalization
2.6. Cell Culture
2.7. Cell Viability Assay
2.8. Morphological Study
2.9. Quantitative Reverse Transcription Polymerase Chain Reaction (RT-qPCR)
2.10. Western Blotting
2.11. Immunostaining
2.12. Quantification and Statistical Analysis
3. Results
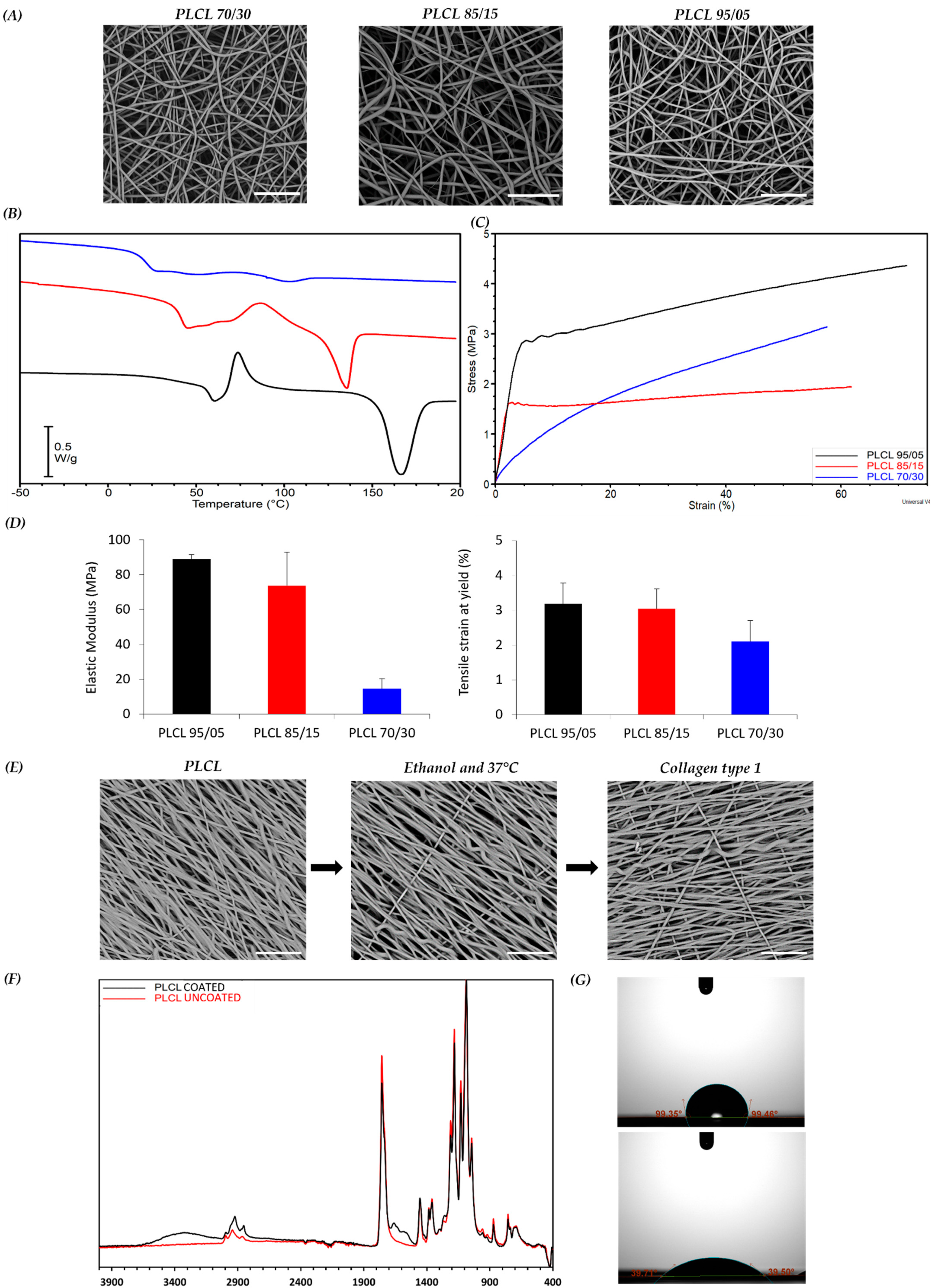
3.1. Fabrication and Characterization of PLCL Scaffold
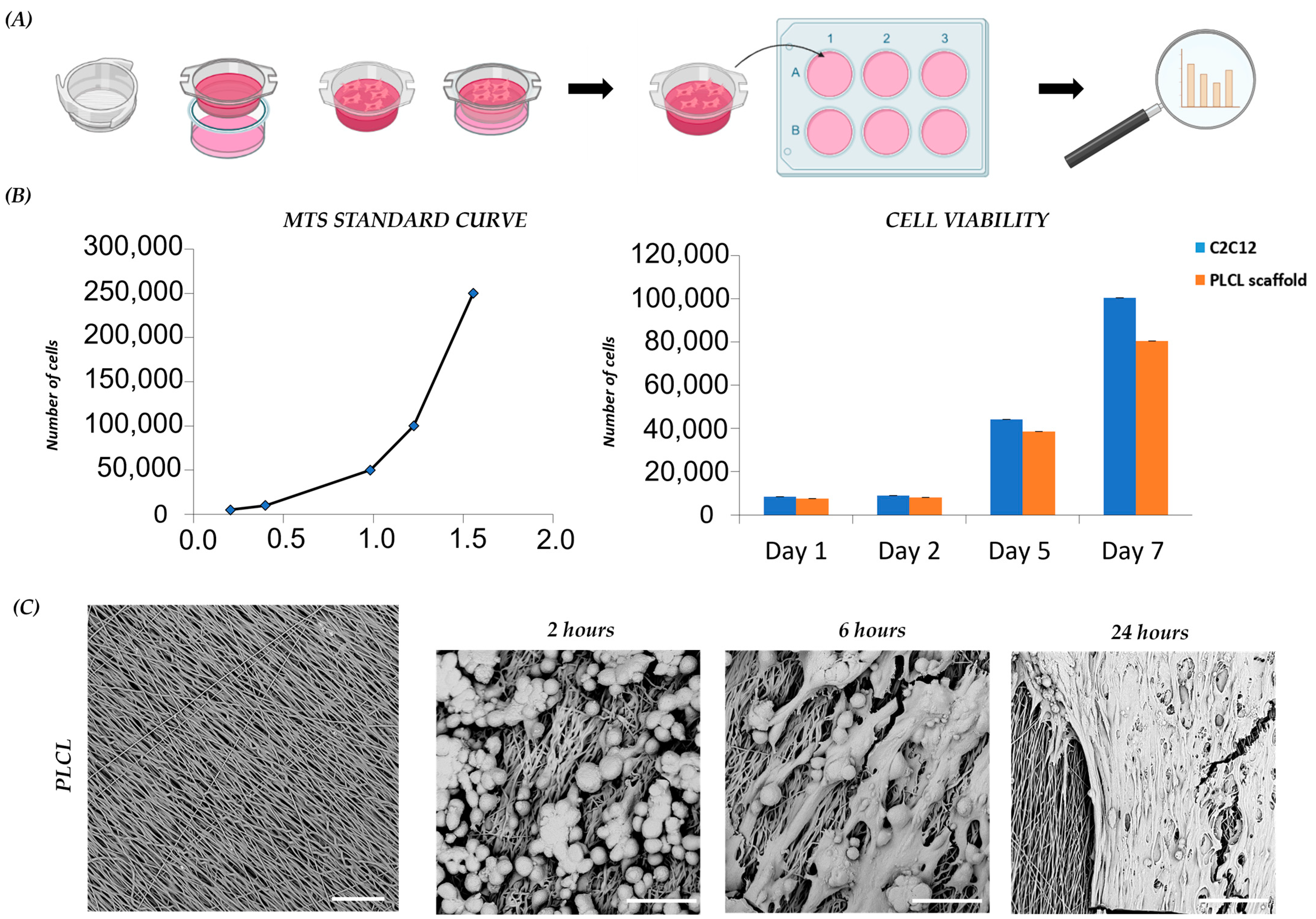
3.2. PLCL Scaffold Influences Proliferation and Morphology of C2C12
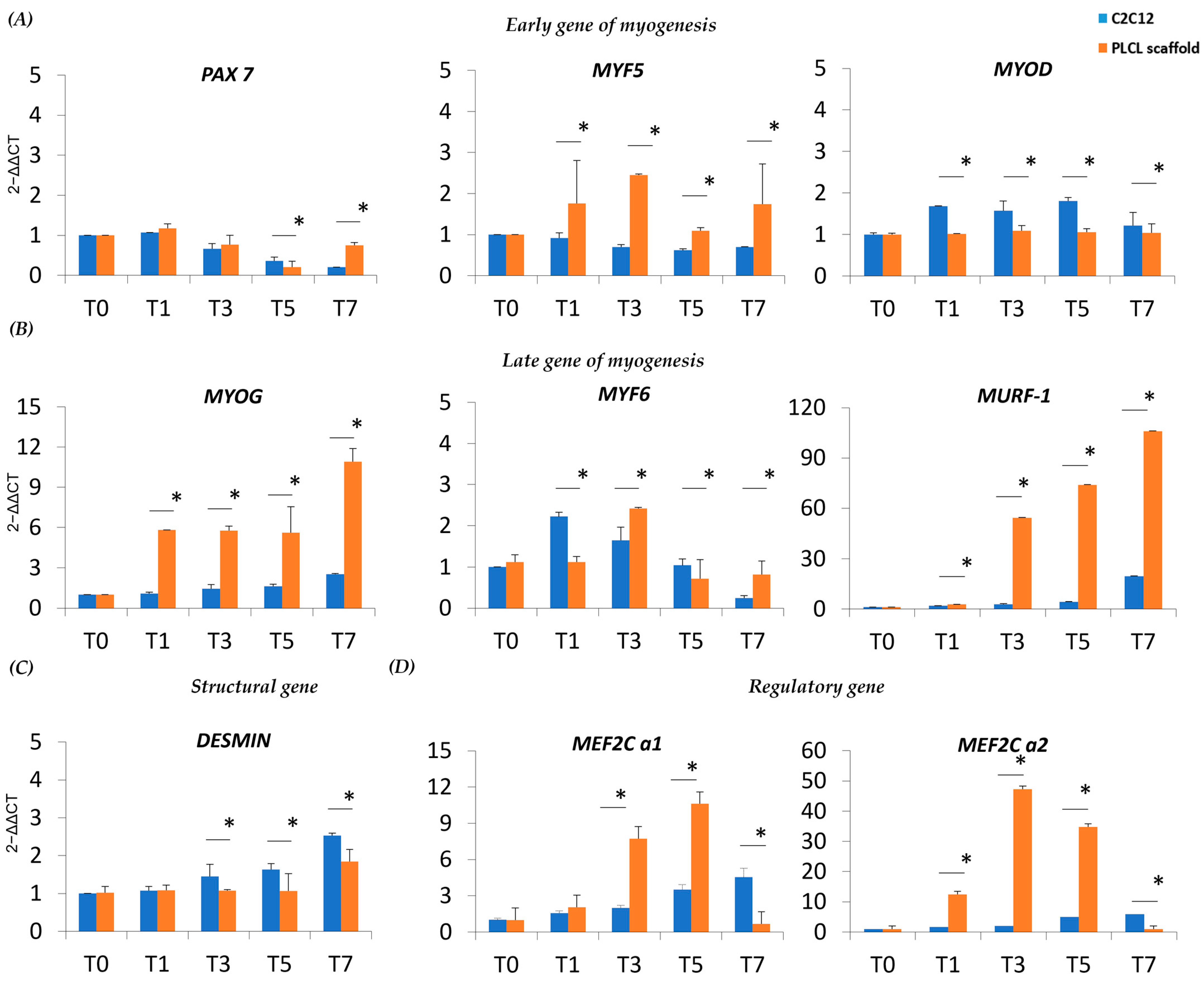
3.3. Myogenesis of C2C12 Cultured on the PLCL Scaffold Differs from the C2C12 Control
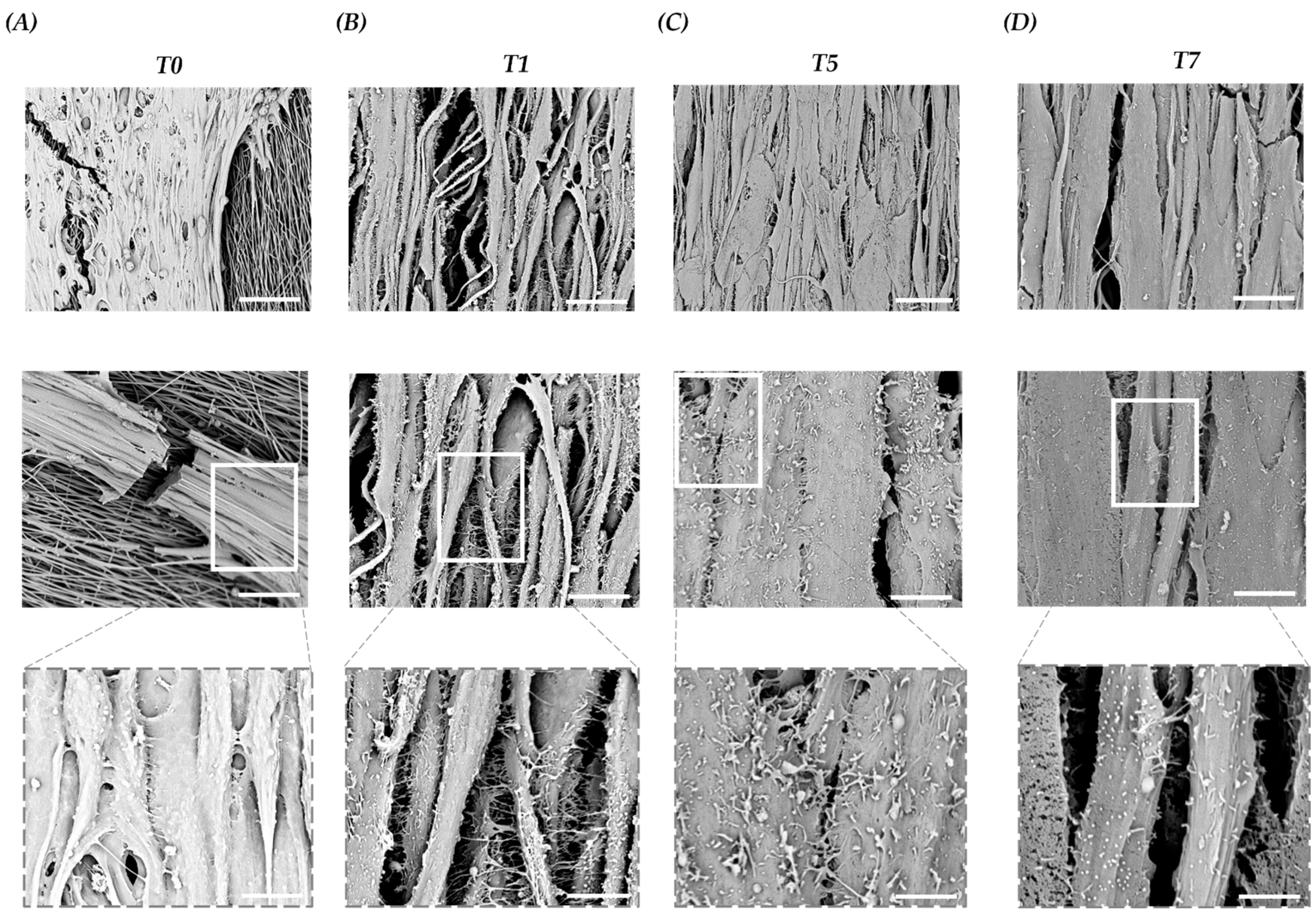
3.4. Scaffold Accelerates Myocyte Fusion and Myotube Formation
3.5. Scaffold Induces Myoblast Interaction and Fusion with The Development of Cell–Cell Adhesions
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mauro, A. Satellite cell of skeletal muscle fibers. J. Cell Biol. 1961, 9, 493–495. [Google Scholar] [CrossRef]
- Corbu, A.; Scaramozza, A.; Badiali-DeGiorgi, L.; Tarantino, L.; Papa, V.; Rinaldi, R.; D’Alessandro, R.; Zavatta, M.; Laus, M.; Lattanzi, G.; et al. Satellite cell characterization from aging human muscle. Neurol. Res. 2010, 32, 63–72. [Google Scholar] [CrossRef]
- Liu, J.; Saul, D.; Böker, K.O.; Ernst, J.; Lehman, W.; Schilling, A.F. Current Methods for Skeletal Muscle Tissue Repair and Regeneration. BioMed Res. Int. 2018, 2018, 1–11. [Google Scholar] [CrossRef]
- Bloise, N.; Berardi, E.; Gualandi, C.; Zaghi, E.; Gigli, M.; Duelen, R.; Ceccarelli, G.; Cortesi, E.E.; Costamagna, D.; Bruni, G.; et al. Ether-oxygen containing electrospun microfibrous and sub-microfibrous scaffolds based on Ppoly(butylene 1,4-cyclohexanedicarboxylate) for skeletal muscle tissue engineering. Int. J. Mol. Sci. 2018, 19, 3212. [Google Scholar] [CrossRef]
- Biressi, S.; Filareto, A.; Rando, T.A. Stem cell therapy for muscular dystrophies. J. Clin. Investig. 2020, 130, 5652–5664. [Google Scholar] [CrossRef]
- Judson, R.N.; Rossi, F.M.V. Towards stem cell therapies for skeletal muscle repair. NPJ Regen. Med. 2020, 5, 10. [Google Scholar] [CrossRef] [PubMed]
- Scharner, J.; Zammit, P.S. The muscle satellite cell at 50: The formative years. Skelet. Muscle 2011, 1, 28. [Google Scholar] [CrossRef] [PubMed]
- Harrison, R.G. The outgrowth of the nerve fiber as a mode of protoplasmic movement. J. Exp. Zoöl. 1959, 142, 5–73. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, B.B.; Danda, D.; Gupta, S.; Gehlot, P. Models for prevention and treatment of cancer: Problems vs promises. Biochem. Pharmacol. 2009, 78, 1083–1094. [Google Scholar] [CrossRef] [PubMed]
- Chan, B.P.; Leong, K.W. Scaffolding in tissue engineering: General approaches and tissue-specific considerations. Eur. Spine J. 2008, 17 (Suppl. 4), 467–479. [Google Scholar] [CrossRef] [PubMed]
- Evans, D.J.R.; Britland, S.; Wigmore, P.M. Differential response of fetal and neonatal myoblasts to topographical guidance cues in vitro. Dev. Genes Evol. 1999, 209, 438–442. [Google Scholar] [CrossRef] [PubMed]
- Gualandi, C.; Govoni, M.; Foroni, L.; Valente, S.; Bianchi, M.; Giordano, E.; Pasquinelli, G.; Biscarini, F.; Focarete, M.L. Ethanol disinfection affects physical properties and cell response of electrospun poly(l-lactic acid) scaffolds. Eur. Polym. J. 2012, 48, 2008–2018. [Google Scholar] [CrossRef]
- Pham, Q.P.; Sharma, U.; Mikos, A.G. Electrospun Poly(ε-caprolactone) Microfiber and Multilayer Nanofiber/Microfiber Scaffolds: Characterization of Scaffolds and Measurement of Cellular Infiltration. Biomacromolecules 2006, 7, 2796–2805. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; Gualandi, C.; Focarete, M.L.; Ravichandran, R.; Venugopal, J.R.; Raghunath, M.; Ramakrishna, S. Elastomeric electrospun scaffolds of poly(l-lactide-co-trimethylene carbonate) for myocardial tissue engineering. J. Mater. Sci. Mater. Med. 2011, 22, 1689–1699. [Google Scholar] [CrossRef]
- Mertens, J.P.; Sugg, K.B.; Lee, J.D.; Larkin, L.M. Engineering muscle constructs for the creation of functional engineered musculoskeletal tissue. Regen. Med. 2014, 9, 89–100. [Google Scholar] [CrossRef] [PubMed]
- Turner, N.J.; Badylak, S.F. Regeneration of skeletal muscle. Cell Tissue Res. 2011, 347, 759–774. [Google Scholar] [CrossRef] [PubMed]
- Chal, J.; Pourquié, O. Making muscle: Skeletal myogenesis in vivo and in vitro. Development 2017, 144, 2104–2122. [Google Scholar] [CrossRef] [PubMed]
- Sanes, J.R. The Basement Membrane/Basal Lamina of Skeletal Muscle. J. Biol. Chem. 2003, 278, 12601–12604. [Google Scholar] [CrossRef]
- Yang, D.; Faraz, F.; Wang, J.; Radacsi, N. Combination of 3D Printing and Electrospinning Techniques for Biofabrication. Adv. Mater. Technol. 2022, 7, 202101309. [Google Scholar] [CrossRef]
- Colapicchioni, V.; Millozzi, F.; Parolini, O.; Palacios, D. Nanomedicine, a valuable tool for skeletal muscle disorders: Challenges, promises, and limitations. WIREs Nanomed. Nanobiotechnol. 2022, 14, e1777. [Google Scholar] [CrossRef]
- Choi, J.S.; Lee, S.J.; Christ, G.J.; Atala, A.; Yoo, J.J. The influence of electrospun aligned poly(ɛ-caprolactone)/collagen nanofiber meshes on the formation of self-aligned skeletal muscle myotubes. Biomaterials 2008, 29, 2899–2906. [Google Scholar] [CrossRef]
- Patel, K.H.; Dunn, A.J.; Talovic, M.; Haas, G.J.; Marcinczyk, M.; Elmashhady, H.; Kalaf, E.G.; Sell, S.A.; Garg, K. Aligned nanofibers of decellularized muscle ECM support myogenic activity in primary satellite cells in vitro. Biomed. Mater. 2019, 14, 035010. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wu, Y.; Guo, B.; Ma, P.X. Nanofiber Yarn/Hydrogel Core–Shell Scaffolds Mimicking Native Skeletal Muscle Tissue for Guiding 3D Myoblast Alignment, Elongation, and Differentiation. ACS Nano 2015, 9, 9167–9179. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Wen, F.; Wong, Y.S.; Boey, F.Y.C.; Subbu, V.S.; Leong, D.T.; Ng, K.W.; Ng, G.K.L.; Tan, L.P. Direct laser machining-induced topographic pattern promotes up-regulation of myogenic markers in human mesenchymal stem cells. Acta Biomater. 2011, 8, 531–539. [Google Scholar] [CrossRef]
- Qian, H.; Bei, J.; Wang, S. Synthesis, characterization and degradation of ABA block copolymer of l-lactide and ε-caprolactone. Polym. Degrad. Stab. 2000, 68, 423–429. [Google Scholar] [CrossRef]
- Penning, J.; Dijkstra, H.; Pennings, A. Preparation and properties of absorbable fibres from l-lactide copolymers. Polymer 1993, 34, 942–951. [Google Scholar] [CrossRef]
- Jeong, S.I.; Kim, B.-S.; Kang, S.W.; Kwon, J.H.; Lee, Y.M.; Kim, S.H.; Kim, Y.H. In vivo biocompatibilty and degradation behavior of elastic poly(l-lactide-co-ε-caprolactone) scaffolds. Biomaterials 2004, 25, 5939–5946. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Zhang, J.; Wang, Y.; Sun, B.; Guo, X.; Morsi, Y.; El-Hamshary, H.; El-Newehy, M.; Mo, X. Development of Dynamic Liquid and Conjugated Electrospun Poly(L-lactide-co-caprolactone)/Collagen Nanoyarns for Regulating Vascular Smooth Muscle Cells Growth. J. Biomed. Nanotechnol. 2017, 13, 303–312. [Google Scholar] [CrossRef]
- Martínez, E.; Lagunas, A.; Mills, C.; Rodríguez-Seguí, S.; Estévez, M.; Oberhansl, S.; Comelles, J.; Samitier, J. Stem cell differentiation by functionalized micro- and nanostructured surfaces. Nanomedicine 2009, 4, 65–82. [Google Scholar] [CrossRef]
- Naidu, P.S.; Ludolph, D.; To, R.; Hinterberger, T.; Konieczny, S.F. Myogenin and MEF2 function syner-gistically to activate the MRF4 promoter during myogenesis. Mol. Cell Biol. 1995, 15, 2707–2718. [Google Scholar] [CrossRef]
- Asfour, H.A.; Allouh, M.Z.; Said, R.S. Myogenic regulatory factors: The orchestrators of myogenesis after 30 years of discovery. Exp. Biol. Med. 2018, 243, 118–128. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef]
- Sensini, A.; Santare, M.H.; Eichenlaub, E.; Bloom, E.; Gotti, C.; Zucchelli, A.; Cristofolini, L. Tuning the Structure of Nylon 6,6 Electrospun Bundles to Mimic the Mechanical Performance of Tendon Fascicles. Front. Bioeng. Biotechnol. 2021, 9, 626433. [Google Scholar] [CrossRef] [PubMed]
- Alessandri, M.; Lizzo, G.; Gualandi, C.; Mangano, C.; Giuliani, A.; Focarete, M.; Calzà, L. Influence of biological matrix and artificial electrospun scaffolds on proliferation, differentiation and trophic factor synthesis of rat embryonic stem cells. Matrix Biol. 2014, 33, 68–76. [Google Scholar] [CrossRef]
- Costa, R.; Rodia, M.T.; Vianello, S.; Santi, S.; Lattanzi, G.; Angelini, C.; Pegoraro, E.; Cenacchi, G. Transportin 3 (TNPO3) and related proteins in limb girdle muscular dystrophy D2 muscle biopsies: A morphological study and pathogenetic hypothesis. Neuromuscul. Disord. 2020, 30, 685–692. [Google Scholar] [CrossRef]
- Mathur, A.B.; Collinsworth, A.M.; Reichert, W.M.; Kraus, W.E.; Truskey, G.A. Endothelial, cardiac muscle and skeletal muscle exhibit different viscous and elastic properties as determined by atomic force microscopy. J. Biomech. 2001, 34, 1545–1553. [Google Scholar] [CrossRef] [PubMed]
- Narayanan, N.; Jiang, C.; Wang, C.; Uzunalli, G.; Whittern, N.; Chen, D.; Jones, O.G.; Kuang, S.; Deng, M. Harnessing Fiber Diameter-Dependent Effects of Myoblasts Toward Biomimetic Scaffold-Based Skeletal Muscle Regeneration. Front. Bioeng. Biotechnol. 2020, 8, 203. [Google Scholar] [CrossRef] [PubMed]
- Semplicini, C.; Bertolin, C.; Bello, L.; Pantic, B.; Guidolin, F.; Vianello, S.; Catapano, F.; Colombo, I.; Moggio, M.; Gavassini, B.F.; et al. The clinical spectrum of CASQ1-related myopathy. Neurology 2018, 91, e1629–e1641. [Google Scholar] [CrossRef]
- Ulery, B.D.; Nair, L.S.; Laurencin, C.T. Biomedical applications of biodegradable polymers. J. Polym. Sci. Part B Polym. Phys. 2011, 49, 832–864. [Google Scholar] [CrossRef] [PubMed]
- Aviss, K.; Gough, J.; Downes, S. Aligned electrospun polymer fibres for skeletal muscle regeneration. Eur. Cells Mater. 2010, 19, 193–204. [Google Scholar] [CrossRef]
- Cronin, E.M.; Thurmond, F.A.; Bassel-Duby, R.; Williams, R.S.; Wright, W.E.; Nelson, K.D.; Garner, H.R. Protein-coated poly(L-lactic acid) fibers provide a substrate for differentiation of human skeletal muscle cells. J. Biomed. Mater. Res. 2004, 69A, 373–381. [Google Scholar] [CrossRef] [PubMed]
- Huang, N.F.; Patel, S.; Thakar, R.G.; Wu, J.; Hsiao, B.S.; Chu, B.; Lee, R.J.; Li, S. Myotube Assembly on Nanofibrous and Micropatterned Polymers. Nano Lett. 2006, 6, 537–542. [Google Scholar] [CrossRef]
- Engler, A.J.; Sen, S.; Sweeney, H.L.; Discher, D.E. Matrix Elasticity Directs Stem Cell Lineage Specification. Cell 2006, 126, 677–689. [Google Scholar] [CrossRef] [PubMed]
- Bacáková, L.; Filová, E.; Rypácek, F.; Svorcík, V.; Starý, V. Cell adhesion on artificial materials for tissue engineering. Physiol. Res. 2004, 53 (Suppl. 1), S35–S45. [Google Scholar]
- Collinsworth, A.M.; Zhang, S.; Kraus, W.E.; Truskey, G.A. Apparent elastic modulus and hysteresis of skeletal muscle cells throughout differentiation. Am. J. Physiol. Physiol. 2002, 283, C1219–C1227. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zhu, B.; Davie, J. Alternative Splicing of MEF2C pre-mRNA Controls Its Activity in Normal Myogenesis and Promotes Tumorigenicity in Rhabdomyosarcoma Cells. J. Biol. Chem. 2015, 290, 310–324. [Google Scholar] [CrossRef] [PubMed]
- Badodi, S.; Baruffaldi, F.; Ganassi, M.; Battini, R.; Molinari, S. Phosphorylation-dependent degradation of MEF2C contributes to regulate G2/M transition. Cell Cycle 2015, 14, 1517–1528. [Google Scholar] [CrossRef]
- Witt, S.H.; Granzier, H.; Witt, C.C.; Labeit, S. MURF-1 and MURF-2 Target a Specific Subset of Myofibrillar Proteins Redundantly: Towards Understanding MURF-dependent Muscle Ubiquitination. J. Mol. Biol. 2005, 350, 713–722. [Google Scholar] [CrossRef]
- McClure, M.J.; Ramey, A.N.; Rashid, M.; Boyan, B.D.; Schwartz, Z. Integrin-α7 signaling regulates connexin 43, M-cadherin, and myoblast fusion. Am. J. Physiol. Physiol. 2019, 316, C876–C887. [Google Scholar] [CrossRef]
- Gorbe, A.; Becker, D.L.; Dux, L.; Krenacs, L.; Krenacs, T. In differentiating prefusion myoblasts connexin43 gap junction coupling is upregulated before myoblast alignment then reduced in post-mitotic cells. Histochem. 2005, 125, 705–716. [Google Scholar] [CrossRef]
- Gorbe, A.; Becker, D.L.; Dux, L.; Stelkovics, E.; Krenács, L.; Bagdi, E.; Krenacs, T. Transient upregulation of connexin43 gap junctions and synchronized cell cycle control precede myoblast fusion in regenerating skeletal muscle in vivo. Histochem. 2005, 123, 573–583. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, S.P.; Neef, K.; Treskes, P.; Liakopoulos, O.J.; Stamm, C.; Cowan, D.B.; Madershahian, N.; Kuhn, E.; Slottosch, I.; Wittwer, T.; et al. Enhanced gap junction expression in myoblast-containing engineered tissue. Biochem. Biophys. Res. Commun. 2012, 422, 462–468. [Google Scholar] [CrossRef]
- Cosgrove, B.D.; Mui, K.L.; Driscoll, T.P.; Caliari, S.R.; Mehta, K.D.; Assoian, R.K.; Burdick, J.A.; Mauck, R.L. N-cadherin adhesive interactions modulate matrix mechanosensing and fate commitment of mesenchymal stem cells. Nat. Mater. 2016, 15, 1297–1306. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Brand, N.J.; Allen, S.; Khan, M.A.; Farrell, A.O.; Murtuza, B.; El Oakley, R.; Yacoub, M.H. Overexpression of connexin 43 in skeletal myoblasts: Relevance to cell transplantation to the heart. J. Thorac. Cardiovasc. Surg. 2001, 122, 759–766. [Google Scholar] [CrossRef] [PubMed]
- Charrasse, S.; Causeret, M.; Comunale, F.; Bonet-Kerrache, A.; Gauthier-Rouvière, C. Rho GTPases and cadher-in-based cell adhesion in skeletal muscle development. J. Muscle Res. Cell Motil. 2003, 24, 309–313. [Google Scholar] [CrossRef] [PubMed]
- Pagnotta, G.; Graziani, G.; Baldini, N.; Maso, A.; Focarete, M.L.; Berni, M.; Biscarini, F.; Bianchi, M.; Gualandi, C. Nanodecoration of electrospun polymeric fibers with nanostructured silver coatings by ionized jet deposition for antibacterial tissues. Mater. Sci. Eng. C 2020, 113, 110998. [Google Scholar] [CrossRef] [PubMed]
- Gotti, C.; Sensini, A.; Fornaia, G.; Gualandi, C.; Zucchelli, A.; Focarete, M.L. Biomimetic Hierarchically Arranged Nanofibrous Structures Resembling the Architecture and the Passive Mechanical Properties of Skeletal Muscles: A Step Forward Toward Artificial Muscle. Front. Bioeng. Biotechnol. 2020, 8, 767. [Google Scholar] [CrossRef] [PubMed]
- Guex, A.; Kocher, F.; Fortunato, G.; Körner, E.; Hegemann, D.; Carrel, T.; Tevaearai, H.; Giraud, M. Fine-tuning of substrate architecture and surface chemistry promotes muscle tissue development. Acta Biomater. 2012, 8, 1481–1489. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene (GenBank Accession Number) (Mus Musculus) | Primers |
|---|---|
| Myf5 Myogenic Factor 5 (NM_008656.5) | Fw: 5’-AGGTGGAGAACTATTACAGC |
| Rev: 5’-TGATACATCAGGACAGTAGATG | |
| Myog Myogenin (NM_031189.2) | Fw: 5’-AGTACATTGAGCGCCTAC |
| Rev: 5’-CAAATGATCTCCTGGGTTG | |
| Des Desmin (NM_010043.2) | Fw: 5’-ACACCTAAAGGATGAGATGG |
| Rev: 5’-GAGAAGGTCTGGATAGGAAG | |
| Murf-1(TRIM63) Muscle-specific RING finger protein 1 (NM_001039048.2) | Fw: 5’-GACTTAGAACACATAGCAGAG |
| Rev: 5’-CTCTTCTGTAAACTCCTCCTC | |
| Myf6 Myogenic Factor 6 (NM_001003982.1) | Fw: 5’-ATAGAGAAGGAGCCGTGTTGG |
| Rev: 5’-TTCTCTGAGATCTGGCTGGGA | |
| Mef2c isoforma α1 Myocyte Enhancer Factor 2 C (NM_001170537.1) | Fw: 5’-CTCAGACATTGTGGAGACATT |
| Rev: 5’-TCAGGGCTGTGACCTACTG | |
| Mef2c isoforma α2 Myocyte Enhancer Factor 2 C (NM_001347568.1) | Fw: 5’-CTCAGACATTGTGGAGGCAT |
| Rev: 5’-TTCTTCAGTGCGTGGGGT | |
| GAPDH Glyceraldehyde-3-Phosphate Dehydrogenase (NM_001256799.2) | Fw: 5’-CTCTGATTTGGTCGTATTGG |
| Rev: 5’-GTAAACCATGTAGTTGAGGTC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pacilio, S.; Costa, R.; Papa, V.; Rodia, M.T.; Gotti, C.; Pagnotta, G.; Cenacchi, G.; Focarete, M.L. Electrospun Poly(L-lactide-co-ε-caprolactone) Scaffold Potentiates C2C12 Myoblast Bioactivity and Acts as a Stimulus for Cell Commitment in Skeletal Muscle Myogenesis. Bioengineering 2023, 10, 239. https://doi.org/10.3390/bioengineering10020239
Pacilio S, Costa R, Papa V, Rodia MT, Gotti C, Pagnotta G, Cenacchi G, Focarete ML. Electrospun Poly(L-lactide-co-ε-caprolactone) Scaffold Potentiates C2C12 Myoblast Bioactivity and Acts as a Stimulus for Cell Commitment in Skeletal Muscle Myogenesis. Bioengineering. 2023; 10(2):239. https://doi.org/10.3390/bioengineering10020239
Chicago/Turabian StylePacilio, Serafina, Roberta Costa, Valentina Papa, Maria Teresa Rodia, Carlo Gotti, Giorgia Pagnotta, Giovanna Cenacchi, and Maria Letizia Focarete. 2023. "Electrospun Poly(L-lactide-co-ε-caprolactone) Scaffold Potentiates C2C12 Myoblast Bioactivity and Acts as a Stimulus for Cell Commitment in Skeletal Muscle Myogenesis" Bioengineering 10, no. 2: 239. https://doi.org/10.3390/bioengineering10020239
APA StylePacilio, S., Costa, R., Papa, V., Rodia, M. T., Gotti, C., Pagnotta, G., Cenacchi, G., & Focarete, M. L. (2023). Electrospun Poly(L-lactide-co-ε-caprolactone) Scaffold Potentiates C2C12 Myoblast Bioactivity and Acts as a Stimulus for Cell Commitment in Skeletal Muscle Myogenesis. Bioengineering, 10(2), 239. https://doi.org/10.3390/bioengineering10020239