
Delamination Strength and Elastin Interlaminar Fibers Decrease with the Development of Aortic Dissection in Model Rats


Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Animal Model of Aortic Dissection
2.2. Tissue Preparation
2.3. Delamination Test
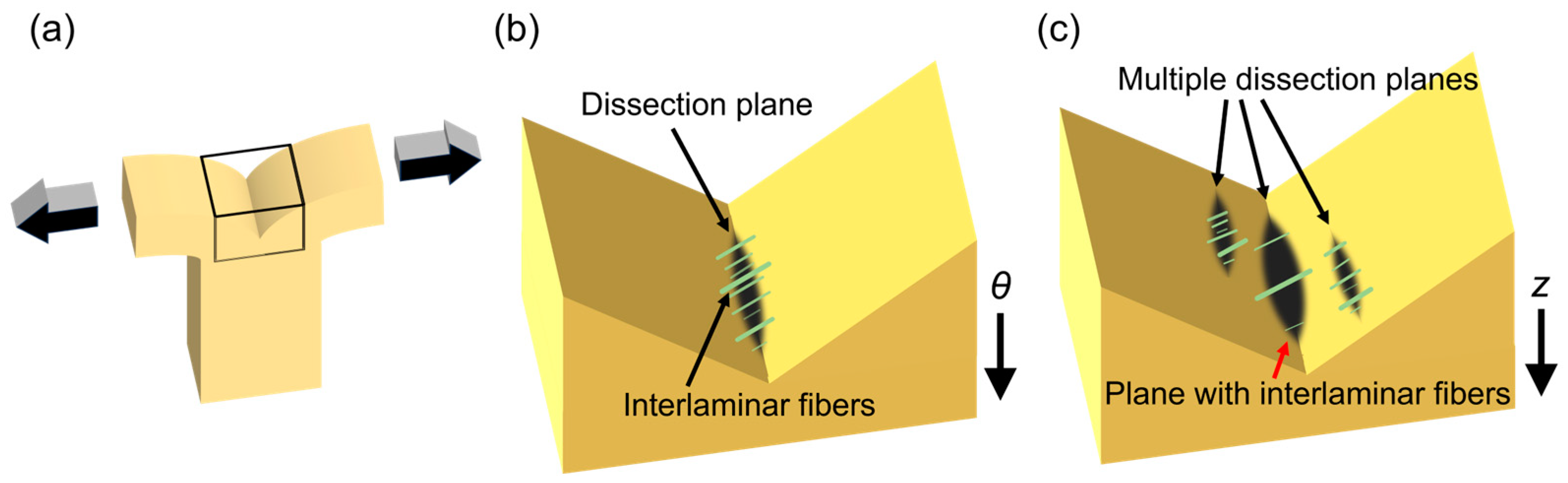
2.4. Histological Test
2.5. Analytical Method
2.6. Statistical Analysis
3. Results
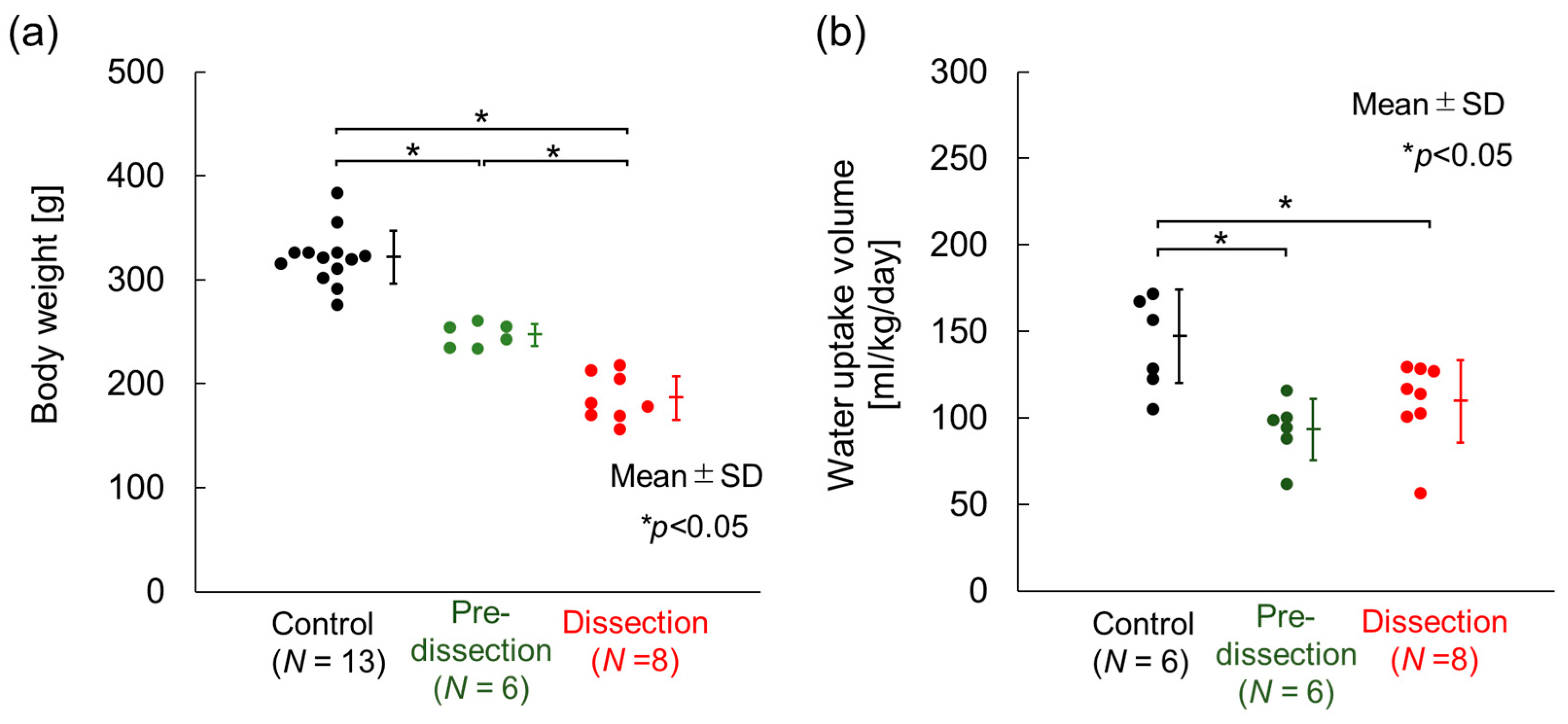
3.1. Body Weight, Water-Uptake Volume, and Aortic-Dissection Generation
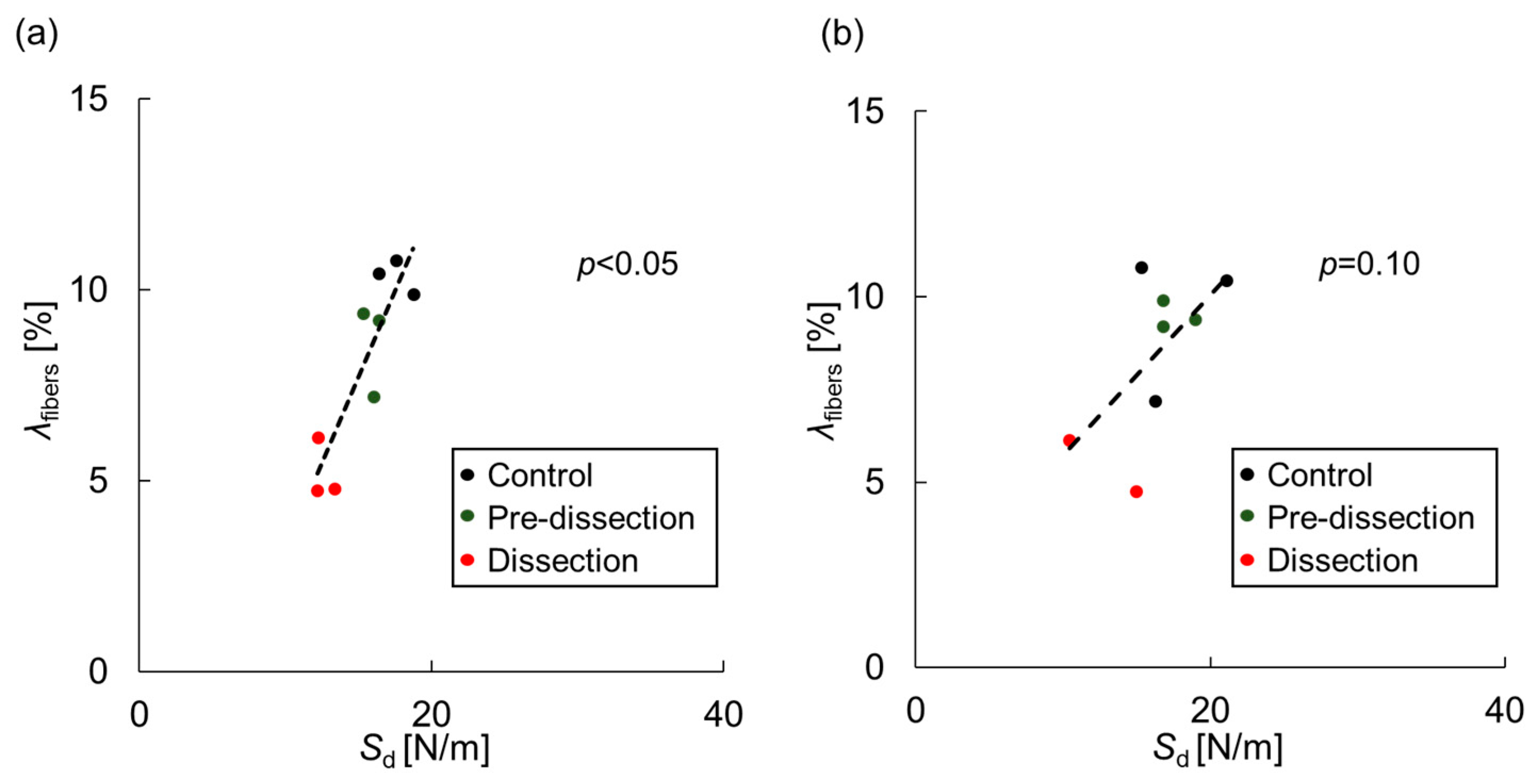
3.2. Delamination Strength
3.3. Histological Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Melvinsdottir, I.H.; Lund, S.H.; Agnarsson, B.A.; Sigvaldason, K.; Gudbjartsson, T.; Geirsson, A. The incidence and mortality of acute thoracic aortic dissection: Results from a whole nation study. Eur. J. Cardiothorac. Surg. 2016, 50, 1111–1117. [Google Scholar] [CrossRef]
- Mody, P.S.; Wang, Y.; Geirsson, A.; Kim, N.; Desai, M.M.; Gupta, A.; Dodson, J.A.; Krumholz, H.M. Trends in aortic dissection hospitalizations, interventions, and outcomes among medicare beneficiaries in the United States, 2000–2011. Circ. Cardiovasc. Qual. Outcomes 2014, 7, 920–928. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; LeMaire, S.A.; Chen, L.; Carter, S.A.; Shen, Y.H.; Gan, Y.; Bartsch, H.; Wilks, J.A.; Utama, B.; Ou, H.; et al. Decreased expression of fibulin-5 correlates with reduced elastin in thoracic aortic dissection. Surgery 2005, 138, 352–359. [Google Scholar] [CrossRef] [PubMed]
- Weis-Müller, B.T.; Modlich, O.; Drobinskaya, I.; Unay, D.; Huber, R.; Bojar, H.; Schipke, J.D.; Feindt, P.; Gams, E.; Müller, W.; et al. Gene expression in acute Stanford type A dissection: A comparative microarray study. J. Transl. Med. 2006, 4, 29. [Google Scholar] [CrossRef][Green Version]
- Wang, X.; LeMaire, S.A.; Chen, L.; Shen, Y.H.; Gan, Y.; Bartsch, H.; Carter, S.A.; Utama, B.; Ou, H.; Coselli, J.S.; et al. Increased collagen deposition and elevated expression of connective tissue growth factor in human thoracic aortic dissection. Circulation 2006, 114, I200–I205. [Google Scholar] [CrossRef]
- Nakashima, Y.; Shiokawa, Y.; Sueishi, K. Alterations of elastic architecture in human aortic dissecting aneurysm. Lab. Investig. 1990, 62, 751–760. [Google Scholar] [PubMed]
- Watanabe, M.; Sawai, T. Alteration of cross-linking amino acids of elastin in human aorta in association with dissecting aneurysm: Analysis using high performance liquid chromatography. Tohoku J. Exp. Med. 1999, 187, 291–303. [Google Scholar] [CrossRef][Green Version]
- Myneni, M.; Rao, A.; Jiang, M.; Moreno, M.R.; Rajagopal, K.R.; Benjamin, C.C. Segmental variations in the peel characteristics of the porcine thoracic aorta. Ann. Biomed. Eng. 2020, 48, 1751–1767. [Google Scholar] [CrossRef]
- Wang, R.; Yu, X.; Gkousioudi, A.; Zhang, Y. Effect of glycation on interlamellar bonding of arterial elastin. Exp. Mech. 2021, 61, 81–94. [Google Scholar] [CrossRef]
- Ríos-Ruiz, I.; Martínez, M.; Peña, E. Is location a significant parameter in the layer dependent dissection properties of the aorta? Biomech. Model. Mechanobiol. 2022, 21, 1887–1901. [Google Scholar] [CrossRef]
- Sommer, G.; Gasser, T.C.; Regitnig, P.; Auer, M.; Holzapfel, G.A. Dissection properties of the human aortic media: An experimental study. J. Biomech. Eng. 2008, 130, 021007. [Google Scholar] [CrossRef]
- Tong, J.; Sommer, G.; Regitnig, P.; Holzapfel, G.A. Dissection properties and mechanical strength of tissue components in human carotid bifurcations. Ann. Biomed. Eng. 2011, 39, 1703–1719. [Google Scholar] [CrossRef] [PubMed]
- Pasta, S.; Phillippi, J.A.; Gleason, T.G.; Vorp, D.A. Effect of aneurysm on the mechanical dissection properties of the human ascending thoracic aorta. J. Thorac. Cardiovasc. Surg. 2012, 143, 460–467. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.A.; Beck, C.; Barbour, J.R.; Zavadzkas, J.A.; Mukherjee, R.; Spinale, F.G.; Ikonomidis, J.S. Alterations in aortic cellular constituents during thoracic aortic aneurysm development: Myofibroblast-mediated vascular remodeling. Am. J. Pathol. 2009, 175, 1746–1756. [Google Scholar] [CrossRef] [PubMed]
- He, C.M.; Roach, M.R. The composition and mechanical properties of abdominal aortic aneurysms. J. Vasc. Surg. 1994, 20, 6–13. [Google Scholar] [CrossRef] [PubMed]
- Noble, C.; Smulders, N.; Lewis, R.; Carré, M.J.; Franklin, S.E.; MacNeil, S.; Taylor, Z.A. Controlled peel testing of a model tissue for diseased aorta. J. Biomech. 2016, 49, 3667–3675. [Google Scholar] [CrossRef][Green Version]
- Kurihara, G.; Ujihara, Y.; Nakamura, M.; Sugita, S. Effect of elastin on delamination strength in the porcine aortic tunica media. J. Biorheol. 2023, in press.
- Cai, Y.L.; Wang, Z.W. The expression and significance of IL-6, IFN-γ, SM22α, and MMP-2 in rat model of aortic dissection. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 560–568. [Google Scholar]
- Luo, B.Y.; Zhou, J.; Guo, D.; Yang, Q.; Tian, Q.; Cai, D.P.; Zhou, R.M.; Xu, Z.Z.; Wang, H.J.; Chen, S.Y.; et al. Methamphetamine induces thoracic aortic aneurysm/dissection through C/EBPβ. Biochimica Et Biophysica Acta—Mol. Basis Dis. 2022, 1868, 166447. [Google Scholar] [CrossRef]
- Guo, L.L.; Wu, M.T.; Zhang, L.W.; Chu, Y.X.; Tian, P.; Jing, Z.P.; Li, J.S.; Sun, Y.D.; Yeung, K.K.; Zhang, L. Blocking Interleukin-1 Beta Reduces the Evolution of Thoracic Aortic Dissection in a Rodent Model. Eur. J. Vasc. Endovasc. Surg. 2020, 60, 916–924. [Google Scholar] [CrossRef]
- Chen, T.; Jiang, N.; Zhang, S.; Chen, Q.; Guo, Z. BAPN-induced rodent model of aortic dissecting aneurysm and related complications. J. Thorac. Dis. 2021, 13, 3643–3651. [Google Scholar] [CrossRef] [PubMed]
- Kurihara, T.; Shimizu-Hirota, R.; Shimoda, M.; Adachi, T.; Shimizu, H.; Weiss, S.J.; Itoh, H.; Hori, S.; Aikawa, N.; Okada, Y. Neutrophil-derived matrix metalloproteinase 9 triggers acute aortic dissection. Circulation 2012, 126, 3070–3080. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, Y.; Sueishi, K. Alteration of elastic architecture in the lathyritic rat aorta implies the pathogenesis of aortic dissecting aneurysm. Am. J. Pathol. 1992, 140, 959–969. [Google Scholar] [PubMed]
- Zhou, H.; Ren, Y.; Xiao, J.; He, J.; Zhang, Y.; Qiu, Z.; Huang, Q.; Hu, Y.; Chen, L. Changes in aortic collagen in β-aminopropionitrile-induced acute aortic dissection. Ann. Transl. Med. 2021, 9, 1574. [Google Scholar] [CrossRef]
- Sugita, S.; Matsumoto, T. Multiphoton microscopy observations of 3D elastin and collagen fiber microstructure changes during pressurization in aortic media. Biomech. Model. Mechanobiol. 2017, 16, 763–773. [Google Scholar] [CrossRef]
- Sugita, S.; Kato, M.; Wataru, F.; Nakamura, M. Three-dimensional analysis of the thoracic aorta microscopic deformation during intraluminal pressurization. Biomech. Model. Mechanobiol. 2020, 19, 147–157. [Google Scholar] [CrossRef]
- Kono, K.; Hayakawa, M.; Kuzuya, F. Effect of vitamin B6 on dissecting aneurysm of rats fed β-aminopropionitrile (Part 1). J. Jpn. Atheroscler. Soc. 1987, 14, 1225–1229. [Google Scholar] [CrossRef]
- Pinnell, S.R.; Martin, G.R. The cross-linking of collagen and elastin: Enzymatic conversion of lysine in peptide linkage to alpha-aminoadipic-delta-semialdehyde (allysine) by an extract from bone. Proc. Natl. Acad. Sci. USA 1968, 61, 708–716. [Google Scholar] [CrossRef]
- O’Connell, M.K.; Murthy, S.; Phan, S.; Xu, C.; Buchanan, J.; Spilker, R.; Dalman, R.L.; Zarins, C.K.; Denk, W.; Taylor, C.A. The three-dimensional micro- and nanostructure of the aortic medial lamellar unit measured using 3D confocal and electron microscopy imaging. Matrix Biol. 2008, 27, 171–181. [Google Scholar] [CrossRef]
- Yu, X.; Suki, B.; Zhang, Y. Avalanches and power law behavior in aortic dissection propagation. Sci. Adv. 2020, 6, eaaz1173. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kurihara, G.; Ujihara, Y.; Nakamura, M.; Sugita, S. Delamination Strength and Elastin Interlaminar Fibers Decrease with the Development of Aortic Dissection in Model Rats. Bioengineering 2023, 10, 1292. https://doi.org/10.3390/bioengineering10111292
Kurihara G, Ujihara Y, Nakamura M, Sugita S. Delamination Strength and Elastin Interlaminar Fibers Decrease with the Development of Aortic Dissection in Model Rats. Bioengineering. 2023; 10(11):1292. https://doi.org/10.3390/bioengineering10111292
Chicago/Turabian StyleKurihara, Genki, Yoshihiro Ujihara, Masanori Nakamura, and Shukei Sugita. 2023. "Delamination Strength and Elastin Interlaminar Fibers Decrease with the Development of Aortic Dissection in Model Rats" Bioengineering 10, no. 11: 1292. https://doi.org/10.3390/bioengineering10111292
APA StyleKurihara, G., Ujihara, Y., Nakamura, M., & Sugita, S. (2023). Delamination Strength and Elastin Interlaminar Fibers Decrease with the Development of Aortic Dissection in Model Rats. Bioengineering, 10(11), 1292. https://doi.org/10.3390/bioengineering10111292