

Quenchbodies That Enable One-Pot Detection of Antigens: A Structural Perspective

Abstract
:
1. Quenchbody
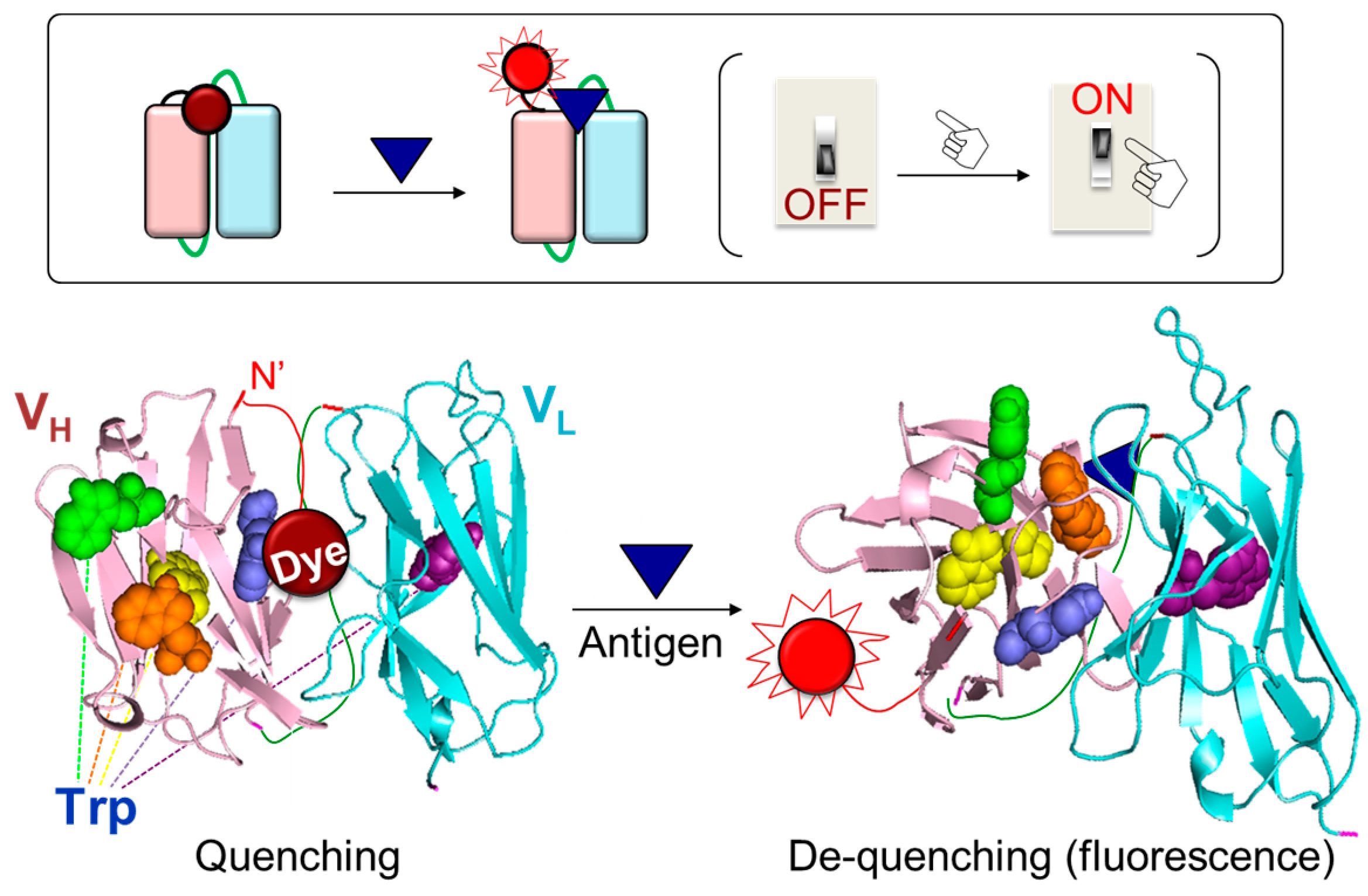
2. Reaction Mechanism of the Q-Body
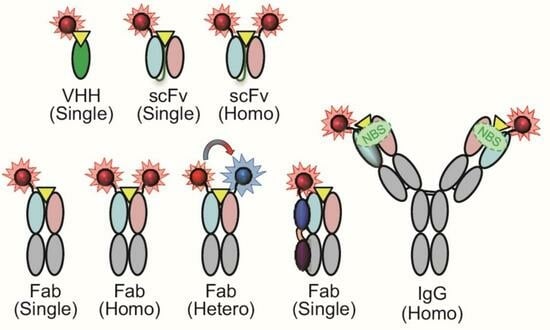
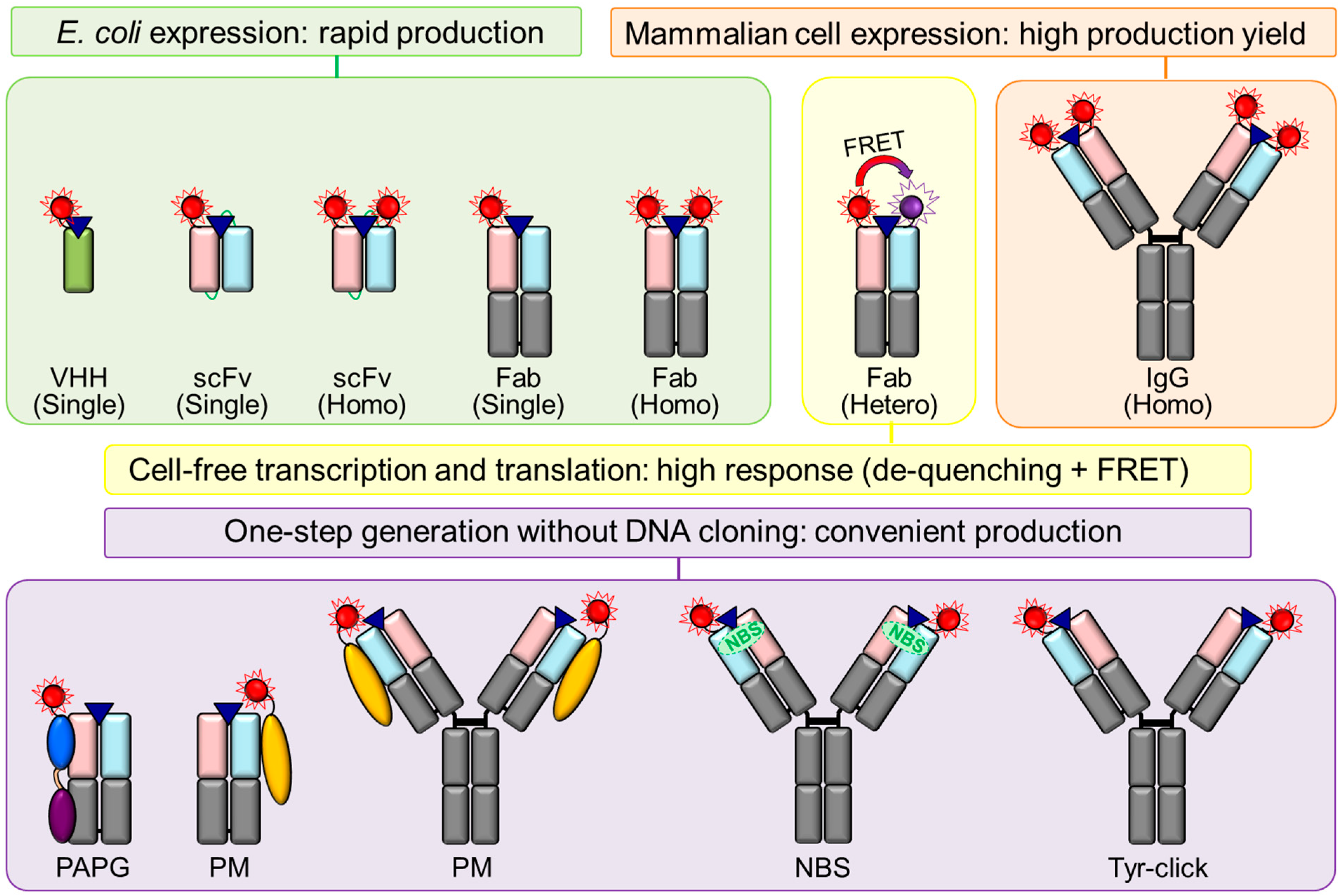
3. Structural Types of Q-Body
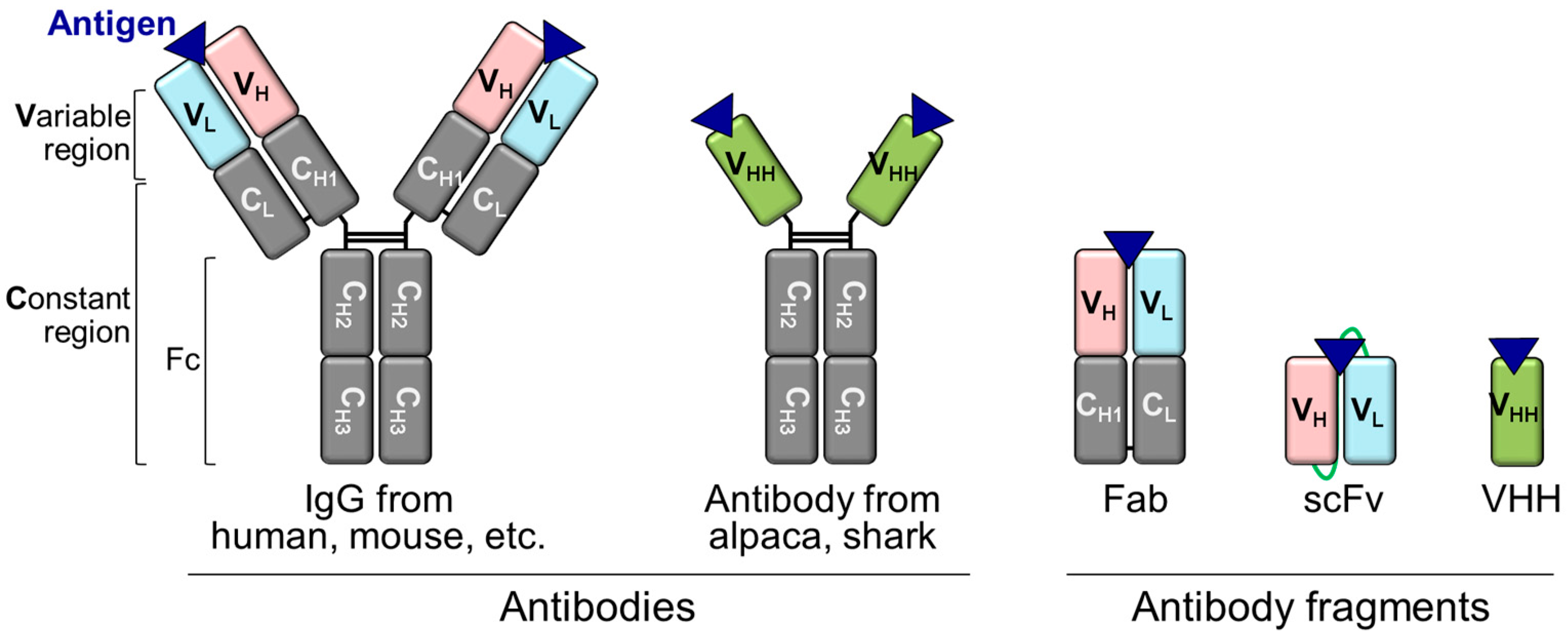
3.1. Structures of IgG and Its Fragments
3.2. scFv-Type Q-Bodies
3.3. Fab-Type Q-Bodies
3.4. IgG-Type Q-Bodies
3.5. VHH-Type Q-Bodies
3.6. Protein-Type Q-Bodies
3.7. Luciferase-Conjugated Q-Bodies
3.8. Structural Strategies for Improving the Fluorescence Response of Q-Body
4. Q-Bodies against Various Antigens of Interest
4.1. Anti-BGP Q-Body as a Model Q-Body
4.2. Q-Bodies against Small Molecules
4.3. Q-Bodies against Proteins
4.4. Q-Bodies against Cells
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kłos-Witkowska, A. The phenomenon of fluorescence in immunosensors. Acta Biochim. Pol. 2016, 63, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Yamanishi, C.D.; Chiu, J.H.; Takayama, S. Systems for multiplexing homogeneous immunoassays. Bioanalysis 2015, 7, 1545–1556. [Google Scholar] [CrossRef] [PubMed]
- Rani, A.Q.; Zhu, B.; Ueda, H.; Kitaguchi, T. Recent progress in homogeneous immunosensors based on fluorescence or bioluminescence using antibody engineering. Analyst 2023, 148, 1422–1429. [Google Scholar] [CrossRef]
- Dinis-Oliveira, R.J. Heterogeneous and homogeneous immunoassays for drug analysis. Bioanalysis 2014, 6, 2877–2896. [Google Scholar] [CrossRef]
- Renard, M.; Bedouelle, H. Improving the sensitivity and dynamic range of reagentless fluorescent immunosensors by knowledge-based design. Biochemistry 2004, 43, 15453–15462. [Google Scholar] [CrossRef] [PubMed]
- Takkinen, K.; Žvirblienė, A. Recent advances in homogenous immunoassays based on resonance energy transfer. Curr. Opin. Biotechnol. 2019, 55, 16–22. [Google Scholar] [CrossRef]
- Fang, C.; Huang, Y.; Zhao, Y. Review of FRET biosensing and its application in biomolecular detection. Am. J. Transl. Res. 2023, 15, 694–709. [Google Scholar]
- Wu, Y.; Jiang, T. Developments in FRET- and BRET-Based Biosensors. Micromachines 2022, 13, 1789. [Google Scholar] [CrossRef]
- Ray, P.C.; Fan, Z.; Crouch, R.A.; Sinha, S.S.; Pramanik, A. Nanoscopic optical rulers beyond the FRET distance limit: Fundamentals and applications. Chem. Soc. Rev. 2014, 43, 6370–6404. [Google Scholar] [CrossRef]
- Bajar, B.T.; Wang, E.S.; Zhang, S.; Lin, M.Z.; Chu, J. A Guide to Fluorescent Protein FRET Pairs. Sensors 2016, 16, 1488. [Google Scholar] [CrossRef]
- Morino, K.; Katsumi, H.; Akahori, Y.; Iba, Y.; Shinohara, M.; Ukai, Y.; Kohara, Y.; Kurosawa, Y. Antibody fusions with fluorescent proteins: A versatile reagent for profiling protein expression. J. Immunol. Methods 2001, 257, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Aoki, T.; Kaneta, M.; Onagi, H.; Takahashi, Y.; Koch, K.S.; Leffert, H.L.; Watabe, H. A simple and rapid immunoassay system using green fluorescent protein tag. J. Immunoass. 1997, 18, 321–333. [Google Scholar] [CrossRef] [PubMed]
- Casey, J.L.; Coley, A.M.; Tilley, L.M.; Foley, M. Green fluorescent antibodies: Novel in vitro tools. Protein Eng. Des. Sel. 2000, 13, 445–452. [Google Scholar] [CrossRef]
- Anderson, G.W.; Zimmerman, J.E.; Callahan, F.M. The Use of Esters of N-Hydroxysuccinimide in Peptide Synthesis. J. Am. Chem. Soc. 1964, 86, 1839–1842. [Google Scholar] [CrossRef]
- Lim, C.Y.; Owens, N.A.; Wampler, R.D.; Ying, Y.; Granger, J.H.; Porter, M.D.; Takahashi, M.; Shimazu, K. Succinimidyl Ester Surface Chemistry: Implications of the Competition between Aminolysis and Hydrolysis on Covalent Protein Immobilization. Langmuir 2014, 30, 12868–12878. [Google Scholar] [CrossRef]
- Abe, R.; Ohashi, H.; Iijima, I.; Ihara, M.; Takagi, H.; Hohsaka, T.; Ueda, H. “Quenchbodies”: Quench-based antibody probes that show antigen-dependent fluorescence. J. Am. Chem. Soc. 2011, 133, 17386–17394. [Google Scholar] [CrossRef]
- Ueda, H.; Dong, J. From fluorescence polarization to Quenchbody: Recent progress in fluorescent reagentless biosensors based on antibody and other binding proteins. Biochim. Biophys. Acta (BBA)—Proteins Proteom. 2014, 1844, 1951–1959. [Google Scholar] [CrossRef]
- Dong, J.; Ueda, H. Recent Advances in Quenchbody, a Fluorescent Immunosensor. Sensors 2021, 21, 1223. [Google Scholar] [CrossRef]
- Rapado, A.; de la Piedra, C.; Torres, R. BGP (osteocalcin, bone-Gla-protein) in involutional osteoporosis. Clin. Rheumatol. 1989, 8 (Suppl. S2), 30–34. [Google Scholar] [CrossRef]
- Marmé, N.; Knemeyer, J.P.; Sauer, M.; Wolfrum, J. Inter- and intramolecular fluorescence quenching of organic dyes by tryptophan. Bioconjug. Chem. 2003, 14, 1133–1139. [Google Scholar] [CrossRef]
- Jones Brunette, A.M.; Farrens, D.L. Distance Mapping in Proteins Using Fluorescence Spectroscopy: Tyrosine, like Tryptophan, Quenches Bimane Fluorescence in a Distance-Dependent Manner. Biochemistry 2014, 53, 6290–6301. [Google Scholar] [CrossRef] [PubMed]
- Doose, S.; Neuweiler, H.; Sauer, M. Fluorescence quenching by photoinduced electron transfer: A reporter for conformational dynamics of macromolecules. ChemPhysChem 2009, 10, 1389–1398. [Google Scholar] [CrossRef] [PubMed]
- Jones, G.; Lu, L.N.; Vullev, V.; Gosztola, D.J.; Greenfield, S.R.; Wasielewski, M.R. Photoinduced electron transfer for pyrenesulfonamide conjugates of tryptophan-containing peptides. Mitigation of fluoroprobe behavior in N-terminal labeling experiments. Bioorganic Med. Chem. Lett. 1995, 5, 2385–2390. [Google Scholar] [CrossRef]
- Jeong, H.-J.; Ueda, H. Strategy for Making a Superior Quenchbody to Proteins: Effect of the Fluorophore Position. Sensors 2014, 14, 13285–13297. [Google Scholar] [CrossRef] [PubMed]
- Vaiana, A.C.; Neuweiler, H.; Schulz, A.; Wolfrum, J.; Sauer, M.; Smith, J.C. Fluorescence Quenching of Dyes by Tryptophan: Interactions at Atomic Detail from Combination of Experiment and Computer Simulation. J. Am. Chem. Soc. 2003, 125, 14564–14572. [Google Scholar] [CrossRef]
- Shirai, K.; Matsuoka, M.; Fukunishi, K. Fluorescence quenching by intermolecular π–π interactions of 2,5-bis(N,N-dialkylamino)-3,6-dicyanopyrazines. Dye. Pigment. 1999, 42, 95–101. [Google Scholar] [CrossRef]
- Chiu, M.L.; Goulet, D.R.; Teplyakov, A.; Gilliland, G.L. Antibody Structure and Function: The Basis for Engineering Therapeutics. Antibodies 2019, 8, 55. [Google Scholar] [CrossRef]
- Rizi, S.Y.; Kim, S.H.; Lee, J.H.; Jeong, H.J. Generation of an scFv-type Quenchbody Against HER2 for Sensitive Detection of Human Breast Cancer. KSBB J. 2023, 38, 35–45. [Google Scholar] [CrossRef]
- Jeong, H.J.K.T.; Dong, J.; Ueda, H. Q-bodies from recombinant single-chain Fv fragment with better yield and expanded palette of fluorophores. ACS Sens. 2016, 1, 88–94. [Google Scholar] [CrossRef]
- Zhao, S.; Dong, J.; Jeong, H.J.; Okumura, K.; Ueda, H. Rapid detection of the neonicotinoid insecticide imidacloprid using a quenchbody assay. Anal. Bioanal. Chem. 2018, 410, 4219–4226. [Google Scholar] [CrossRef]
- Dong, J.; Banwait, B.; Ueda, H.; Kristensen, P. V(H)-Based Mini Q-Body: A Novel Quench-Based Immunosensor. Sensors 2023, 23, 2251. [Google Scholar] [CrossRef]
- Ohashi, H.; Matsumoto, T.; Jeong, H.J.; Dong, J.; Abe, R.; Ueda, H. Insight into the Working Mechanism of Quenchbody: Transition of the Dye around Antibody Variable Region That Fluoresces upon Antigen Binding. Bioconjug. Chem. 2016, 27, 2248–2253. [Google Scholar] [CrossRef] [PubMed]
- Mori, Y.O.H.; Watanabe, T.; Hohsaka, T. Antigen-dependent fluorescence response of anti-c-Myc Quenchbody studied by molecular dynamics simulations. Chem. Phys. Lett. 2018, 698, 223–226. [Google Scholar] [CrossRef]
- Swindells, M.B.; Porter, C.T.; Couch, M.; Hurst, J.; Abhinandan, K.R.; Nielsen, J.H.; Macindoe, G.; Hetherington, J.; Martin, A.C. abYsis: Integrated Antibody Sequence and Structure-Management, Analysis, and Prediction. J. Mol. Biol. 2017, 429, 356–364. [Google Scholar] [CrossRef]
- Doose, S.; Neuweiler, H.; Sauer, M. A close look at fluorescence quenching of organic dyes by tryptophan. ChemPhysChem 2005, 6, 2277–2285. [Google Scholar] [CrossRef]
- Jeong, H.J.; Itayama, S.; Ueda, H. A signal-on fluorosensor based on quench-release principle for sensitive detection of antibiotic rapamycin. Biosensors 2015, 5, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Abe, R.; Jeong, H.J.; Arakawa, D.; Dong, J.; Ohashi, H.; Kaigome, R.; Saiki, F.; Yamane, K.; Takagi, H.; Ueda, H. Ultra Q-bodies: Quench-based antibody probes that utilize dye-dye interactions with enhanced antigen-dependent fluorescence. Sci. Rep. 2014, 4, 4640. [Google Scholar] [CrossRef]
- Jeong, H.J.; Kawamura, T.; Iida, M.; Kawahigashi, Y.; Takigawa, M.; Ohmuro-Matsuyama, Y.; Chung, C.I.; Dong, J.; Kondoh, M.; Ueda, H. Development of a Quenchbody for the Detection and Imaging of the Cancer-Related Tight-Junction-Associated Membrane Protein Claudin. Anal. Chem. 2017, 89, 10783–10789. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Fujita, R.; Zako, T.; Ueda, H. Construction of Quenchbodies to detect and image amyloid β oligomers. Anal. Biochem. 2018, 550, 61–67. [Google Scholar] [CrossRef]
- Sasao, A.; Takaki, M.; Jeong, H.J.; Yonemitsu, K.; Ohtsu, Y.; Tsutsumi, H.; Furukawa, S.; Morioka, H.; Ueda, H.; Nishitani, Y. Development of a fluvoxamine detection system using a Quenchbody, a novel fluorescent biosensor. Drug Test. Anal. 2019, 11, 601–609. [Google Scholar] [CrossRef]
- Li, H.; Li, X.; Chen, L.; Li, B.; Dong, H.; Liu, H.; Yang, X.; Ueda, H.; Dong, J. Quench-Release-Based Fluorescent Immunosensor for the Rapid Detection of Tumor Necrosis Factor α. ACS Omega 2021, 6, 31009–31016. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zhu, B.; Li, B.; Chen, L.; Ning, X.; Dong, H.; Liang, J.; Yang, X.; Dong, J.; Ueda, H. Isolation of a human SARS-CoV-2 neutralizing antibody from a synthetic phage library and its conversion to fluorescent biosensors. Sci. Rep. 2022, 12, 15496. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Zhang, R.; Gao, Y.; Cheng, Y.; Zhao, S.; Li, M.; Li, H.; Dong, J. Immunosensor for Rapid and Sensitive Detection of Digoxin. ACS Omega 2023, 8, 15341–15349. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.; Nosaka, N.; Kanamaru, S.; Dong, J.; Dai, Y.; Inoue, A.; Yang, Y.; Kobayashi, K.; Kitaguchi, T.; Iwasaki, H.; et al. Rapid and sensitive SARS-CoV-2 detection using a homogeneous fluorescent immunosensor Quenchbody with crowding agents. Analyst 2022, 147, 4971–4979. [Google Scholar] [CrossRef]
- Kurumida, Y.; Hayashi, N. Development of a Novel Q-body Using an In Vivo Site-Specific Unnatural Amino Acid Incorporation System. Sensors 2018, 18, 2519. [Google Scholar] [CrossRef] [PubMed]
- Sato, S.; Matsumura, M.; Ueda, H.; Nakamura, H. Preparation of an antigen-responsive fluorogenic immunosensor by tyrosine chemical modification of the antibody complementarity determining region. Chem. Commun. 2021, 57, 9760–9763. [Google Scholar] [CrossRef]
- Zanetti-Domingues, L.C.; Tynan, C.J.; Rolfe, D.J.; Clarke, D.T.; Martin-Fernandez, M. Hydrophobic fluorescent probes introduce artifacts into single molecule tracking experiments due to non-specific binding. PLoS ONE 2013, 8, e74200. [Google Scholar] [CrossRef]
- Kowada, T.; Maeda, H.; Kikuchi, K. BODIPY-based probes for the fluorescence imaging of biomolecules in living cells. Chem. Soc. Rev. 2015, 44, 4953–4972. [Google Scholar] [CrossRef]
- Nelson, A.L. Antibody fragments: Hope and hype. MAbs 2010, 2, 77–83. [Google Scholar] [CrossRef]
- Safdari, Y.; Ahmadzadeh, V.; Khalili, M.; Jaliani, H.Z.; Zarei, V.; Erfani-Moghadam, V. Use of single chain antibody derivatives for targeted drug delivery. Mol. Med. 2016, 22, 258–270. [Google Scholar] [CrossRef]
- Ahmad, Z.A.; Yeap, S.K.; Ali, A.M.; Ho, W.Y.; Alitheen, N.B.; Hamid, M. scFv antibody: Principles and clinical application. Clin. Dev. Immunol. 2012, 2012, 980250. [Google Scholar] [CrossRef] [PubMed]
- Ito, Y.; Sasaki, R.; Asari, S.; Yasuda, T.; Ueda, H.; Kitaguchi, T. Efficient Microfluidic Screening Method Using a Fluorescent Immunosensor for Recombinant Protein Secretions. Small 2023, 19, e2207943. [Google Scholar] [CrossRef] [PubMed]
- Jeong, H.J.; Matsumoto, K.; Itayama, S.; Kodama, K.; Abe, R.; Dong, J.; Shindo, M.; Ueda, H. Construction of dye-stapled Quenchbodies by photochemical crosslinking to antibody nucleotide-binding sites. Chem. Commun. 2017, 53, 10200–10203. [Google Scholar] [CrossRef] [PubMed]
- Jeong, H.J.; Ohmuro-Matsuyama, Y.; Ohashi, H.; Ohsawa, F.; Tatsu, Y.; Inagaki, M.; Ueda, H. Detection of vimentin serine phosphorylation by multicolor Quenchbodies. Biosens. Bioelectron. 2013, 40, 17–23. [Google Scholar] [CrossRef]
- Dong, J.; Miyake, C.; Yasuda, T.; Oyama, H.; Morita, I.; Tsukahara, T.; Takahashi, M.; Jeong, H.J.; Kitaguchi, T.; Kobayashi, N.; et al. PM Q-probe: A fluorescent binding protein that converts many antibodies to a fluorescent biosensor. Biosens. Bioelectron. 2020, 165, 112425. [Google Scholar] [CrossRef]
- Ning, X.; Yasuda, T.; Kitaguchi, T.; Ueda, H. Construction of Fluorescent Immunosensor Quenchbody to Detect His-Tagged Recombinant Proteins Produced in Bioprocess. Sensors 2021, 21, 4993. [Google Scholar] [CrossRef]
- Dai, Y.; Sato, Y.; Zhu, B.; Kitaguchi, T.; Kimura, H.; Ghadessy, F.J.; Ueda, H. Intra Q-body: An antibody-based fluorogenic probe for intracellular proteins that allows live cell imaging and sorting. Chem. Sci. 2022, 13, 9739–9748. [Google Scholar] [CrossRef]
- Dong, J.; Jeong, H.J.; Ueda, H. Preparation of Quenchbodies by protein transamination reaction. J. Biosci. Bioeng. 2016, 122, 125–130. [Google Scholar] [CrossRef]
- Yasuda, T.; Inoue, A.; Kitaguchi, T.; Ueda, H. Rapid construction of fluorescence quenching-based immunosensor Q-bodies using α-helical coiled-coil peptides. Chem. Commun. 2021, 57, 8206–8209. [Google Scholar] [CrossRef]
- Yoshinari, T.; Ohashi, H.; Abe, R.; Kaigome, R.; Ohkawa, H.; Sugita-Konishi, Y. Development of a rapid method for the quantitative determination of deoxynivalenol using Quenchbody. Anal. Chim. Acta 2015, 888, 126–130. [Google Scholar] [CrossRef]
- Dong, J.; Oka, Y.; Jeong, H.J.; Ohmuro-Matsuyama, Y.; Ueda, H. Detection and destruction of HER2-positive cancer cells by Ultra Quenchbody-siRNA complex. Biotechnol. Bioeng. 2020, 117, 1259–1269. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Dong, H.; Wang, H.; Yi, Z.; Jiang, G.; Inagaki, T.; Gomez-Sanchez, C.E.; Dong, J.; Ueda, H. Creation of a quick and sensitive fluorescent immunosensor for detecting the mineralocorticoid steroid hormone aldosterone. J. Steroid Biochem. Mol. Biol. 2022, 221, 106118. [Google Scholar] [CrossRef] [PubMed]
- Ueda, H.; Dai, Y.; Ghadessy, F. Visualizing intracellular target antigens in live cells. Trends Cell Biol. 2023, 33, 277–279. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Dong, H.; Xu, F.; Li, B.; Li, H.; Chen, L.; Li, M.; Liu, Y.; Jiang, G.; Dong, J. Isolation of a Monoclonal Antibody and its Derived Immunosensor for Rapid and Sensitive Detection of 17β-Estradiol. Front. Bioeng. Biotechnol. 2022, 10, 818983. [Google Scholar] [CrossRef] [PubMed]
- Fukunaga, K.; Watanabe, T.; Novitasari, D.; Ohashi, H.; Abe, R.; Hohsaka, T. Antigen-responsive fluorescent antibody probes generated by selective N-terminal modification of IgGs. Chem. Commun. 2018, 54, 12734–12737. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Zhao, S.; Zhang, R.; Li, H.; Li, M.; Yu, B.; Dong, J. Immunosensor for realtime monitoring of the expression of recombinant proteins during bioprocess. Anal. Biochem. 2023, 665, 115069. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Li, H.; Li, B.; Li, M.; Xu, W.; Chen, L.; Wang, B.; Dong, J. Rapid conversion of IgG to biosensor using an antibody-binding protein-based probe. Analyst 2021, 146, 6114–6118. [Google Scholar] [CrossRef]
- Jeong, H.-J.; Kojima, T.; Dong, J.; Ohashi, H.; Ueda, H. One-pot construction of Quenchbodies using antibody-binding proteins. Anal. Methods 2016, 8, 7774–7779. [Google Scholar] [CrossRef]
- Inoue, A.; Ohmuro-Matsuyama, Y.; Kitaguchi, T.; Ueda, H. Creation of a Nanobody-Based Fluorescent Immunosensor Mini Q-body for Rapid Signal-On Detection of Small Hapten Methotrexate. ACS Sens. 2020, 5, 3457–3464. [Google Scholar] [CrossRef]
- Inoue, A.; Yasuda, T.; Zhu, B.; Kitaguchi, T.; Murakami, A.; Ueda, H. Evaluation and selection of potent fluorescent immunosensors by combining fluorescent peptide and nanobodies displayed on yeast surface. Sci. Rep. 2021, 11, 22590. [Google Scholar] [CrossRef]
- Liang, Y.F.; Li, J.D.; Fang, R.Y.; Xu, Z.L.; Luo, L.; Chen, Z.J.; Yang, J.Y.; Shen, Y.D.; Ueda, H.; Hammock, B.; et al. Design of an Antigen-Triggered Nanobody-Based Fluorescence Probe for PET Immunoassay to Detect Quinalphos in Food Samples. Anal. Chem. 2023, 95, 12321–12328. [Google Scholar] [CrossRef]
- Li, X.; Huang, X.; Zhang, L.; Cong, Y.; Zhao, G.; Liang, J.; Chen, H.; Li, H.; Chen, L.; Dong, J. Development of a fluorescent probe for the detection of hPD-L1. J. Biosci. Bioeng. 2020, 130, 431–436. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, R.; Yasuda, T.; Ohmuro-Matsuyama, Y.; Ueda, H. BRET Q-Body: A Ratiometric Quench-based Bioluminescent Immunosensor Made of Luciferase-Dye-Antibody Fusion with Enhanced Response. Anal. Chem. 2021, 93, 7571–7578. [Google Scholar] [CrossRef]
- Rodriguez, E.A.; Lester, H.A.; Dougherty, D.A. In vivo incorporation of multiple unnatural amino acids through nonsense and frameshift suppression. Proc. Natl. Acad. Sci. USA 2006, 103, 8650–8655. [Google Scholar] [CrossRef]
- Ohtsuki, T.; Manabe, T.; Sisido, M. Multiple incorporation of non-natural amino acids into a single protein using tRNAs with non-standard structures. FEBS Lett. 2005, 579, 6769–6774. [Google Scholar] [CrossRef] [PubMed]
- Hohsaka, T.; Kajihara, D.; Ashizuka, Y.; Murakami, H.; Sisido, M. Efficient Incorporation of Nonnatural Amino Acids with Large Aromatic Groups into Streptavidin in In Vitro Protein Synthesizing Systems. J. Am. Chem. Soc. 1999, 121, 34–40. [Google Scholar] [CrossRef]
- Abe, R.; Shiraga, K.; Ebisu, S.; Takagi, H.; Hohsaka, T. Incorporation of fluorescent non-natural amino acids into N-terminal tag of proteins in cell-free translation and its dependence on position and neighboring codons. J. Biosci. Bioeng. 2010, 110, 32–38. [Google Scholar] [CrossRef]
- Northrop, B.H.; Frayne, S.H.; Choudhary, U. Thiol–maleimide “click” chemistry: Evaluating the influence of solvent, initiator, and thiol on the reaction mechanism, kinetics, and selectivity. Polym. Chem. 2015, 6, 3415–3430. [Google Scholar] [CrossRef]
- Popp, M.W.; Antos, J.M.; Grotenbreg, G.M.; Spooner, E.; Ploegh, H.L. Sortagging: A versatile method for protein labeling. Nat. Chem. Biol. 2007, 3, 707–708. [Google Scholar] [CrossRef]
- Yun, H.; Ueda, H.; Jeong, H.-J. Development of a Spacer-optimized Quenchbody against Tumor Necrosis Factor Alpha. Biotechnol. Bioprocess Eng. 2022, 27, 846–856. [Google Scholar] [CrossRef]
- Taira, H.; Fukushima, M.; Hohsaka, T.; Sisido, M. Four-base codon-mediated incorporation of non-natural amino acids into proteins in a eukaryotic cell-free translation system. J. Biosci. Bioeng. 2005, 99, 473–476. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, M.; Kosaka, N.; Choyke, P.L.; Kobayashi, H. H-Type Dimer Formation of Fluorophores: A Mechanism for Activatable, in Vivo Optical Molecular Imaging. ACS Chem. Biol. 2009, 4, 535–546. [Google Scholar] [CrossRef] [PubMed]
- Bollmann, S.; Löllmann, M.; Sauer, M.; Doose, S. Dimer formation of organic fluorophores reports on biomolecular dynamics under denaturing conditions. Phys. Chem. Chem. Phys. 2011, 13, 12874–12882. [Google Scholar] [CrossRef]
- KÖHler, G.; Milstein, C. Continuous cultures of fused cells secreting antibody of predefined specificity. Nature 1975, 256, 495–497. [Google Scholar] [CrossRef]
- McCafferty, J.; Griffiths, A.D.; Winter, G.; Chiswell, D.J. Phage antibodies: Filamentous phage displaying antibody variable domains. Nature 1990, 348, 552–554. [Google Scholar] [CrossRef]
- Clackson, T.; Hoogenboom, H.R.; Griffiths, A.D.; Winter, G. Making antibody fragments using phage display libraries. Nature 1991, 352, 624–628. [Google Scholar] [CrossRef]
- Forsgren, A.; Sjöquist, J. “Protein A” from S. Aureus: I. Pseudo-Immune Reaction with Human γ-Globulin. J. Immunol. 1966, 97, 822–827. [Google Scholar] [CrossRef] [PubMed]
- Roben, P.W.; Salem, A.N.; Silverman, G.J. VH3 family antibodies bind domain D of staphylococcal protein A. J. Immunol. 1995, 154, 6437–6445. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Kojima, T.; Ohashi, H.; Ueda, H. Optimal fusion of antibody binding domains resulted in higher affinity and wider specificity. J. Biosci. Bioeng. 2015, 120, 504–509. [Google Scholar] [CrossRef]
- Björck, L.; Kronvall, G. Purification and some properties of streptococcal protein G, a novel IgG-binding reagent. J. Immunol. 1984, 133, 969–974. [Google Scholar] [CrossRef]
- Sjöbring, U.; Björck, L.; Kastern, W. Streptococcal protein G. Gene structure and protein binding properties. J. Biol. Chem. 1991, 266, 399–405. [Google Scholar] [CrossRef]
- Grover, R.K.; Zhu, X.; Nieusma, T.; Jones, T.; Boero, I.; MacLeod, A.S.; Mark, A.; Niessen, S.; Kim, H.J.; Kong, L.; et al. A Structurally Distinct Human Mycoplasma Protein that Generically Blocks Antigen-Antibody Union. Science 2014, 343, 656–661. [Google Scholar] [CrossRef] [PubMed]
- Alves, N.J.; Stimple, S.D.; Handlogten, M.W.; Ashley, J.D.; Kiziltepe, T.; Bilgicer, B. Small-molecule-based affinity chromatography method for antibody purification via nucleotide binding site targeting. Anal. Chem. 2012, 84, 7721–7728. [Google Scholar] [CrossRef] [PubMed]
- Alves, N.J.; Kiziltepe, T.; Bilgicer, B. Oriented surface immobilization of antibodies at the conserved nucleotide binding site for enhanced antigen detection. Langmuir 2012, 28, 9640–9648. [Google Scholar] [CrossRef] [PubMed]
- Alves, N.J.; Champion, M.M.; Stefanick, J.F.; Handlogten, M.W.; Moustakas, D.T.; Shi, Y.; Shaw, B.F.; Navari, R.M.; Kiziltepe, T.; Bilgicer, B. Selective photocrosslinking of functional ligands to antibodies via the conserved nucleotide binding site. Biomaterials 2013, 34, 5700–5710. [Google Scholar] [CrossRef]
- Jin, B.K.; Odongo, S.; Radwanska, M.; Magez, S. Nanobodies: A Review of Generation, Diagnostics and Therapeutics. Int. J. Mol. Sci. 2023, 24, 5994. [Google Scholar] [CrossRef]
- Arbabi-Ghahroudi, M. Camelid Single-Domain Antibodies: Promises and Challenges as Lifesaving Treatments. Int. J. Mol. Sci. 2022, 23, 5009. [Google Scholar] [CrossRef]
- England, C.G.; Ehlerding, E.B.; Cai, W. NanoLuc: A Small Luciferase Is Brightening Up the Field of Bioluminescence. Bioconjugate Chem. 2016, 27, 1175–1187. [Google Scholar] [CrossRef]
- Kobayashi, H.; Picard, L.-P.; Schönegge, A.-M.; Bouvier, M. Bioluminescence resonance energy transfer–based imaging of protein–protein interactions in living cells. Nat. Protoc. 2019, 14, 1084–1107. [Google Scholar] [CrossRef]
- Xu, X.; Soutto, M.; Xie, Q.; Servick, S.; Subramanian, C.; von Arnim, A.G.; Johnson, C.H. Imaging protein interactions with bioluminescence resonance energy transfer (BRET) in plant and mammalian cells and tissues. Proc. Natl. Acad. Sci. USA 2007, 104, 10264–10269. [Google Scholar] [CrossRef]
- Milligan, G. Applications of bioluminescence- and fluorescence resonance energy transfer to drug discovery at G protein-coupled receptors. Eur. J. Pharm. Sci. 2004, 21, 397–405. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.K.; Sowa, H.; Hinoi, E.; Ferron, M.; Ahn, J.D.; Confavreux, C.; Dacquin, R.; Mee, P.J.; McKee, M.D.; Jung, D.Y.; et al. Endocrine regulation of energy metabolism by the skeleton. Cell 2007, 130, 456–469. [Google Scholar] [CrossRef] [PubMed]
- Oury, F.; Sumara, G.; Sumara, O.; Ferron, M.; Chang, H.; Smith, C.E.; Hermo, L.; Suarez, S.; Roth, B.L.; Ducy, P.; et al. Endocrine regulation of male fertility by the skeleton. Cell 2011, 144, 796–809. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.L.; Ichinose, H.; Shinoda, T.; Ueda, H. Noncompetitive detection of low molecular weight peptides by open sandwich immunoassay. Anal. Chem. 2007, 79, 6193–6200. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, C.; Ueda, H.; Mahoney, W.; Nagamune, T. Open sandwich enzyme-linked immunosorbent assay for the quantitation of small haptens. Anal. Biochem. 2000, 286, 238–246. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Fei, J.; Wu, W.; Dou, L.; Liu, M.; Shao, S.; Yu, W.; Wen, K.; Shen, J.; Wang, Z. Minimum Distance Between Two Epitopes in Sandwich Immunoassays for Small Molecules. Anal. Chem. 2022, 94, 17843–17852. [Google Scholar] [CrossRef] [PubMed]
- Baylet, A.; Vyumvuhore, R.; Laclaverie, M.; Marchand, L.; Mainzer, C.; Bordes, S.; Closs-Gonthier, B.; Delpy, L. Transcutaneous penetration of a single-chain variable fragment (scFv) compared to a full-size antibody: Potential tool for atopic dermatitis (AD) treatment. Allergy Asthma Clin. Immunol. 2021, 17, 73. [Google Scholar] [CrossRef]
- Neri, D. Antibody-Cytokine Fusions: Versatile Products for the Modulation of Anticancer Immunity. Cancer Immunol. Res. 2019, 7, 348–354. [Google Scholar] [CrossRef]
- Jin, S.; Sun, Y.; Liang, X.; Gu, X.; Ning, J.; Xu, Y.; Chen, S.; Pan, L. Emerging new therapeutic antibody derivatives for cancer treatment. Signal Transduct. Target. Ther. 2022, 7, 39. [Google Scholar] [CrossRef]
- Costantini, L.M.; Snapp, E.L. Going Viral with Fluorescent Proteins. J. Virol. 2015, 89, 9706–9708. [Google Scholar] [CrossRef]
- Costantini, L.M.; Baloban, M.; Markwardt, M.L.; Rizzo, M.A.; Guo, F.; Verkhusha, V.V.; Snapp, E.L. A palette of fluorescent proteins optimized for diverse cellular environments. Nat. Commun. 2015, 6, 7670. [Google Scholar] [CrossRef] [PubMed]
- Kuo, T.T.; Aveson, V.G. Neonatal Fc receptor and IgG-based therapeutics. MAbs 2011, 3, 422–430. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Ishii-Watabe, A.; Tada, M.; Kobayashi, T.; Kanayasu-Toyoda, T.; Kawanishi, T.; Yamaguchi, T. Importance of Neonatal FcR in Regulating the Serum Half-Life of Therapeutic Proteins Containing the Fc Domain of Human IgG1: A Comparative Study of the Affinity of Monoclonal Antibodies and Fc-Fusion Proteins to Human Neonatal FcR. J. Immunol. 2010, 184, 1968–1976. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Q-Body Type | Antigen | Reference |
|---|---|---|
| scFv | Bone gla protein (BGP) or BGP peptide | [16,29,32,52,53] |
| Bisphenol A | [16,32] | |
| Hen egg lysozyme (HEL) | [16,24] | |
| Bovine serum albumin (BSA) | [16] | |
| Human serum albumin (HSA) | [16] | |
| Estradiol | [16] | |
| Morphine | [16] | |
| Heroin | [16] | |
| Codeine | [16] | |
| Phosphorylated vimentin peptide | [54] | |
| HIV antigen Nef | [45] | |
| Fluvoxamine | [40] | |
| Cortisol | [55] | |
| His-tag | [56] | |
| p53 tumor suppressor | [57] | |
| Fab | BGP or BGP peptide | [37,58,59] |
| HSA | [37] | |
| Morphine | [37] | |
| Methamphetamine | [32,37] | |
| Cocaine | [37] | |
| Deoxynivalenol | [32,60] | |
| Claudin4 | [38] | |
| Amyloid β oligomer | [39] | |
| HER2+ cells | [61] | |
| His-tag | [56] | |
| Tumor Necrosis Factor-α | [41] | |
| Aldosterone | [62] | |
| p53 tumor suppressor | [57,63] | |
| SARS-CoV-2 S1 protein | [42] | |
| SARS-CoV-2 pseudovirus | [42] | |
| SARS-CoV-2 N protein | [44] | |
| Digoxin | [43] | |
| 17β-estradiol | [64] | |
| IgG | HSA | [53] |
| FLAG-tag | [65,66] | |
| His-tag | [65] | |
| HA-tag | [65] | |
| Thyroxine | [65] | |
| Testosterone | [67] | |
| CD20+ cells | [46] | |
| HER2+ cells | [46] | |
| SARS-CoV-2 S1 protein | [42] | |
| PAPG | BGP or BGP peptide | [68] |
| Vimentin | [68] | |
| PM | BGP or BGP peptide | [55] |
| Progesterone | [55] | |
| Testosterone | [55] | |
| Thyroxine4 | [55] | |
| VHH | Methotrexate | [59,69,70] |
| HSA | [70] | |
| HEL | [31] | |
| Quinalphos | [71] | |
| Protein | Rapamycin | [36] |
| PD-L1 | [72] | |
| Luciferase-fusion | BGP | [73] |
| Antibody Format | Labeling Method | Dye (Position) | Response (Fold) | EC50 (nM) | Reference |
|---|---|---|---|---|---|
| scFv | Cell-free | TMR-C6 | 5.6 | 25 | [16] |
| TMR | 5.8 | 38.0 | [54] | ||
| R6G | 7.7 | 84.6 | [54] | ||
| AT520 | 2.9 | 17.7 | [54] | ||
| Maleimide–thiol | TMR-C0 | 2.9 | n.d. | [29] | |
| TMR-C2 | 2.0 | n.d. | [29] | ||
| TMR-C5 | 4.0 | 4.4 | [29] | ||
| AT495 | 1.1 | n.d. | [29] | ||
| AT520 | 2.7 | 6.3 | [29] | ||
| R6G | 5.0 | 11.4 | [29] | ||
| Rho | 1.7 | 70.9 | [29] | ||
| NBS | IBA-C8-TMR | 9 | 116.8 ± 32.0 | [53] | |
| BRET | AT520 | 1.9 | n.d. | [73] | |
| R6G | 4.8 | n.d. | [73] | ||
| TMR | 7.2 | n.d. | [73] | ||
| Fab | Cell-free | TMR (H) | 9.6 | 21 | [37] |
| TMR (L) | 1.8 | 7.1 | [37] | ||
| TMR (H), TMR (L) | 21.2 | 10,000 | [37] | ||
| R110 (H) | 1.1 | 7.8 | [37] | ||
| R110 (L) | 1.3 | 7.2 | [37] | ||
| R110 (H), R110 (L) | 2.4 | 7.6 | [37] | ||
| AT655 (H) | 4.6 | 4.6 | [37] | ||
| AT655 (L) | 2.7 | 9.8 | [37] | ||
| AT655 (H), AT655 (L) | 10.6 | 44 | [37] | ||
| TMR (H), R110 (L) | 2.7 | 8.8 | [37] | ||
| R110 (H), TMR (L) | 19 | 110 | [37] | ||
| TMR (H), AT655 (L) | 35 | 61 | [37] | ||
| AT655 (H), TMR (L) | 50 | 110 | [37] | ||
| Maleimide–thiol | AT520 (H, L) | 11 | 77 | [37] | |
| AT520 | 3.6 | n.d. | [37] | ||
| TMR-C5 (H, L) | 2.4 | n.d. | [37] | ||
| TMR-C5 | 7.0 | 19 | [37] | ||
| Transamination | TMR-C5 | 2.75 | n.d. | [58] | |
| Aminooxy-5(6)-TMR | 1.70 | n.d. | [58] | ||
| PM | TMR-C6 | 2.0 | 81 ± 8 | [55] | |
| Coiled-coil | TMR | 2.7 | 17 | [46] | |
| IgG | PM | TMR-C6 | ~1.9 | 7.3 ± 0.6 | [55] |
| Target Antigen | Antibody Format | Maximum Response (Fold) | EC50 | LOD | Reference |
|---|---|---|---|---|---|
| Bisphenol A | scFv | 2.0 | 20 nM | n.d. | [16] |
| BSA | scFv | 1.5 | n.d. | n.d. | [16] |
| 17β-estradiol | scFv | 4.5 | n.d. | n.d. | [16] |
| Codeine | scFv | ~1.5 | 19 nM | n.d. | [16] |
| Heroin | scFv | ~1.5 | 37 nM | n.d. | [16] |
| Vimentin PS71 | scFv | 3.95 ± 0.4 | n.d. | n.d. | [54] |
| Vimentin PS82 | scFv | 6.7 ± 0.2 | 3.8 nM | n.d. | [54] |
| HSA | Fab | 7.8 | n.d. | n.d. | [37] |
| Influenza HA protein | Fab | 7 | n.d. | n.d. | [37] |
| Morphine | Fab | 7.2 | n.d. | n.d. | [37] |
| Methamphetamine | Fab | 7.2 | n.d. | n.d. | [37] |
| Cocaine | Fab | 3.5 | n.d. | n.d. | [37] |
| HEL | VHH | ~4.5 | 397 nM | n.d. | [31] |
| DON | Fab | 3.9 | 101 nM | 5000 nM | [60] |
| Rapamycin | Protein | 1.5 | 3.6 nM | 0.65 nM | [36] |
| Claudin | Fab | 3.0 | n.d. | 1.9 ± 0.7 nM | [38] |
| Amyloid β peptide | Fab | 2.2 | 1090 ± 40 nM | 200 nM | [39] |
| ADDL | Fab | 2.8 | 33,000 ± 26,000 nM | 300 nM | [39] |
| Bio-DAE10 | Fab | 2.1 | n.d. | n.d. | [39] |
| FLV | scFv | 1.5 | 31.4 nM | n.d. | [40] |
| HER2 | Fab | 3.6 | 1.1 nM | 0.08 nM | [61] |
| Progesterone | IgG | ~1.3 | 110 ± 4 nM | 0.96 nM | [55] |
| Testosterone | IgG | ~2.0 | 1.37± 0.05 nM | 0.35 nM | [55] |
| Thyroxine T4 | IgG | ~2.8 | 15.8 ± 1.2 nM | 4.1 nM | [55] |
| Cortisol | scFv | 2.46 | 45,200 nM | n.d. | [55] |
| Methotrexate | VHH | 6.33 | 37.6 ± 11 nM | 0.56 nM | [69] |
| c-Myc | scFv | 2.2 | n.d. | n.d. | [33] |
| Thyroxine T3 | IgG | 1.3 ± 0.05 | n.d. | n.d. | [65] |
| FLAG | IgG | 9.5 ± 0.5 | n.d. | n.d. | [65] |
| hPDL1 | Protein | 1.43 | 110 nM | 0.1 nM | [72] |
| Nef (HIV protein) | scFv | 2 | n.d. | n.d. | [45] |
| His-tag peptide | Fab | ~5 | 132,000 ± 34,000 nM | 3500 nM | [56] |
| CD20 | IgG | ~3.5 | n.d. | n.d. | [46] |
| Tumor necrosis factor α | Fab | 4.05 | 65.6 ng/mL | 0.419 ng/mL | [41] |
| Aldosterone | Fab | 5 | 42.3 ng/mL | 24.1 pg/mL | [62] |
| p53 | Fab | 27 | 60 nM | 0.72 nM | [57] |
| SARS-CoV-2 spike protein | Fab | ~1.5 | n.d. | 0.11 nM | [42] |
| SARS-CoV-2 nucleocapsid protein | Fab | n.d. | n.d. | 0.191 nM | [44] |
| Digoxin | Fab | 2.2 | 0.256 ng/mL | 0.023 ng/mL | [43] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeong, H.-J. Quenchbodies That Enable One-Pot Detection of Antigens: A Structural Perspective. Bioengineering 2023, 10, 1262. https://doi.org/10.3390/bioengineering10111262
Jeong H-J. Quenchbodies That Enable One-Pot Detection of Antigens: A Structural Perspective. Bioengineering. 2023; 10(11):1262. https://doi.org/10.3390/bioengineering10111262
Chicago/Turabian StyleJeong, Hee-Jin. 2023. "Quenchbodies That Enable One-Pot Detection of Antigens: A Structural Perspective" Bioengineering 10, no. 11: 1262. https://doi.org/10.3390/bioengineering10111262
APA StyleJeong, H.-J. (2023). Quenchbodies That Enable One-Pot Detection of Antigens: A Structural Perspective. Bioengineering, 10(11), 1262. https://doi.org/10.3390/bioengineering10111262