
Characterizing Soil and Bedrock Water Use of Native California Vegetation


 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Overview
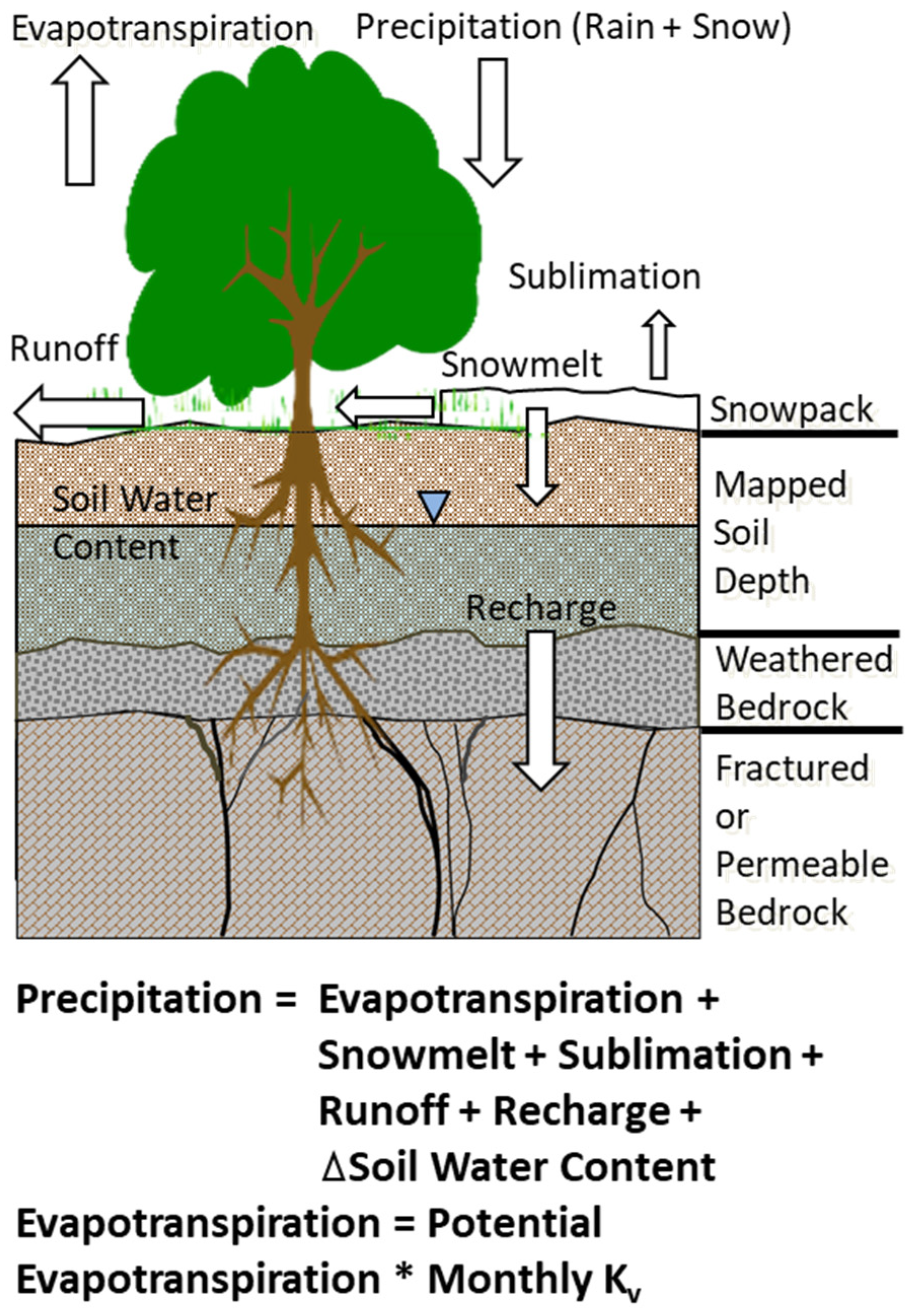
2.2. Model Description
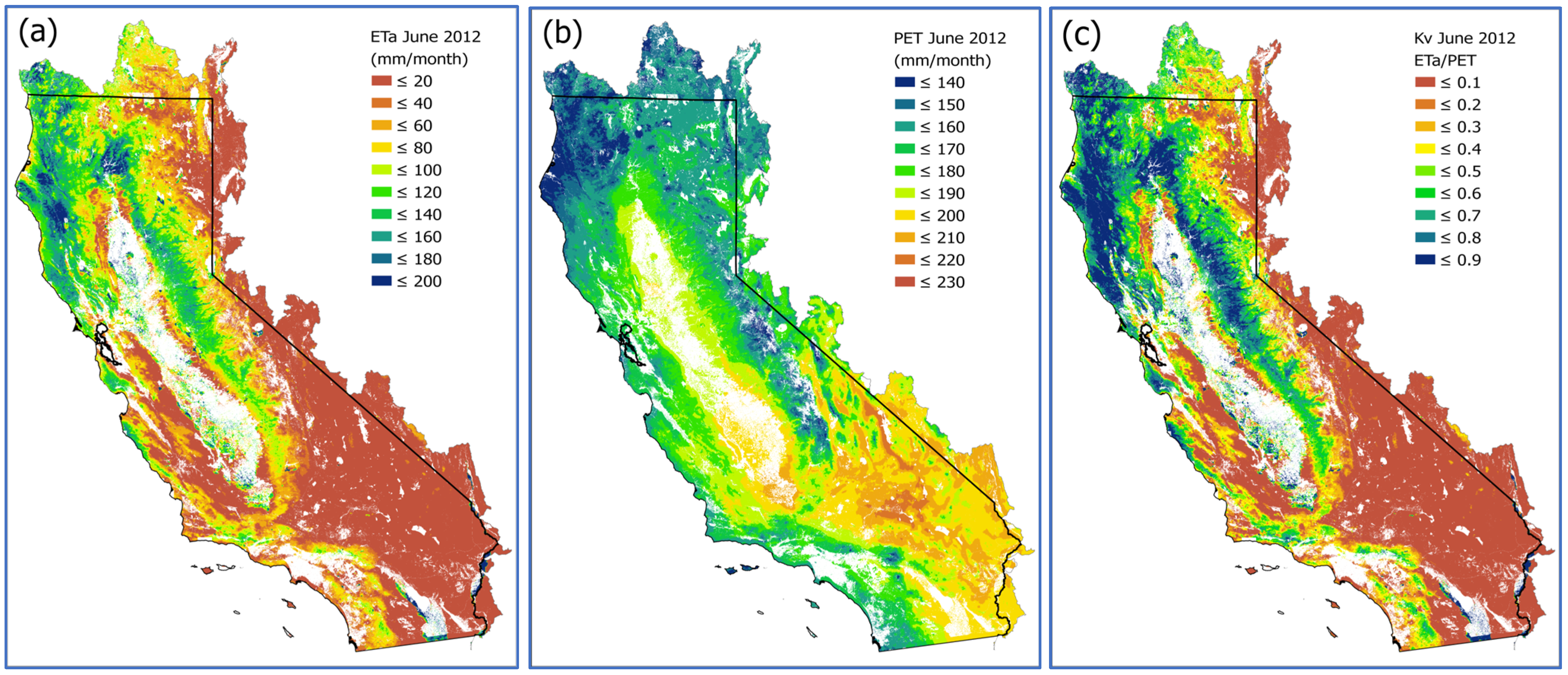
2.3. Spatially Distributed Estimates of Actual Evapotranspiration
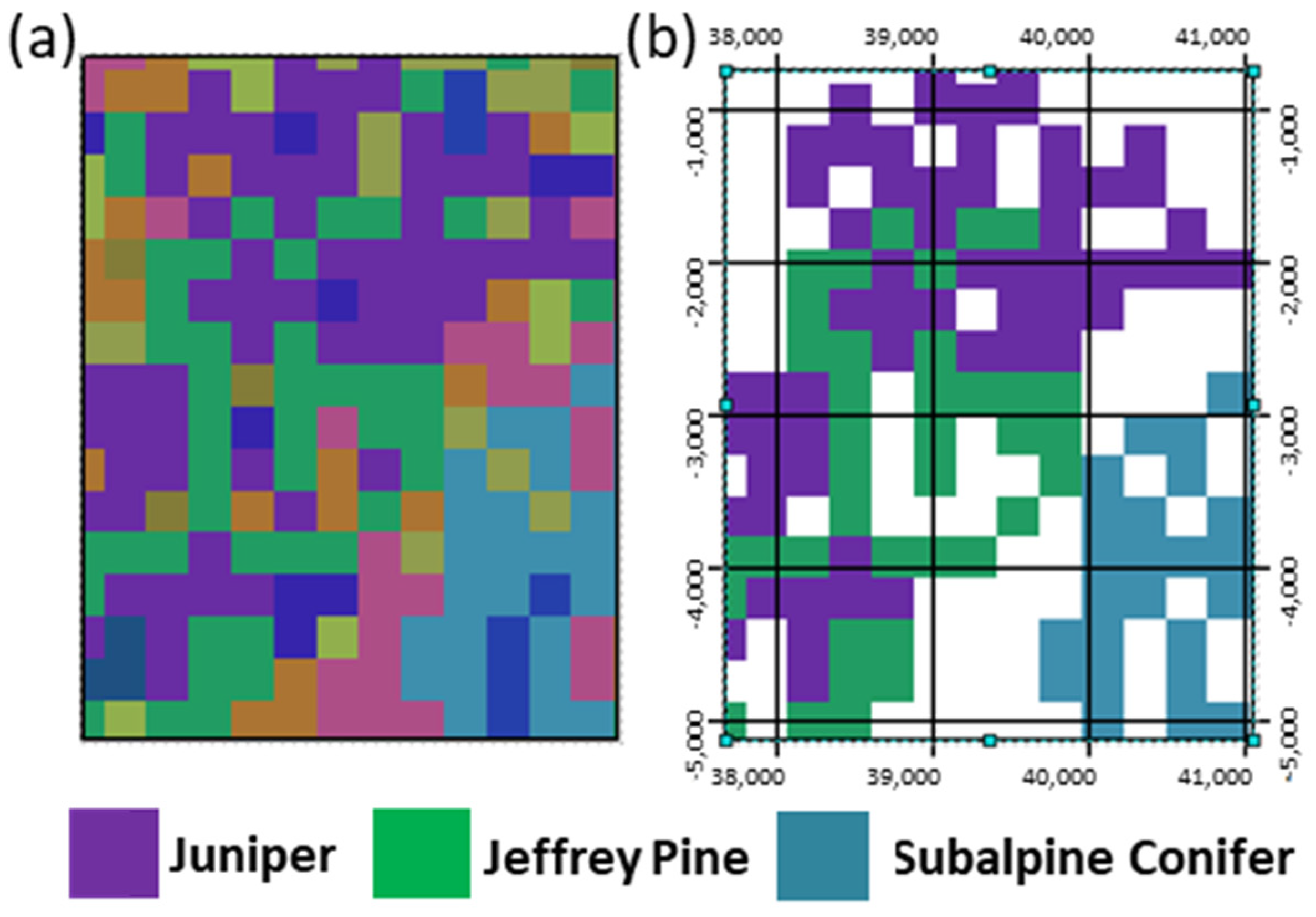
2.4. Development of Vegetation Type Map
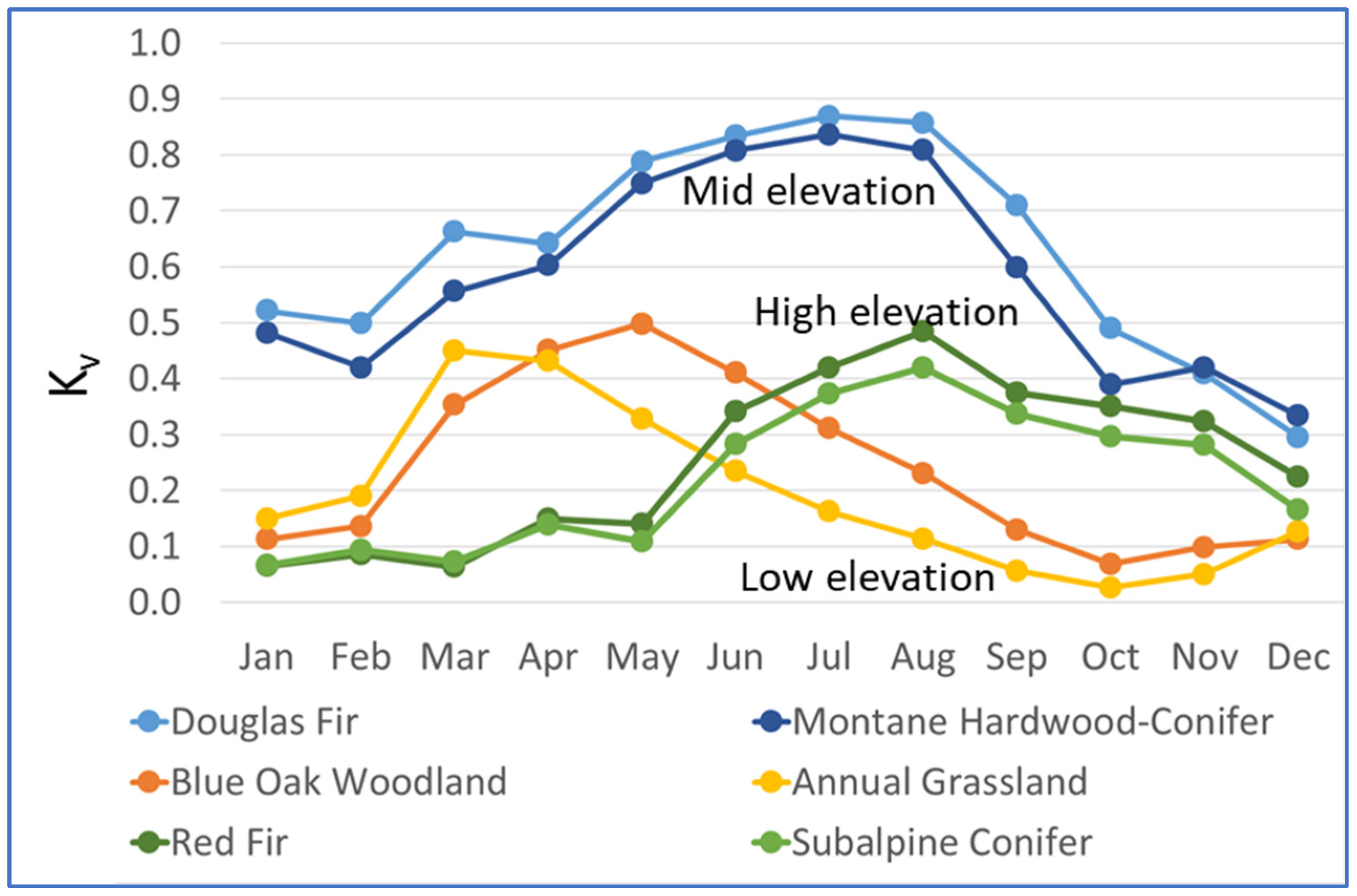
2.5. Calculation of Kv
2.6. Model Calibration to Match AET
2.7. Projecting Calibrated Landscape ET to Assess Drought Stress
3. Results
3.1. Kv Calculations
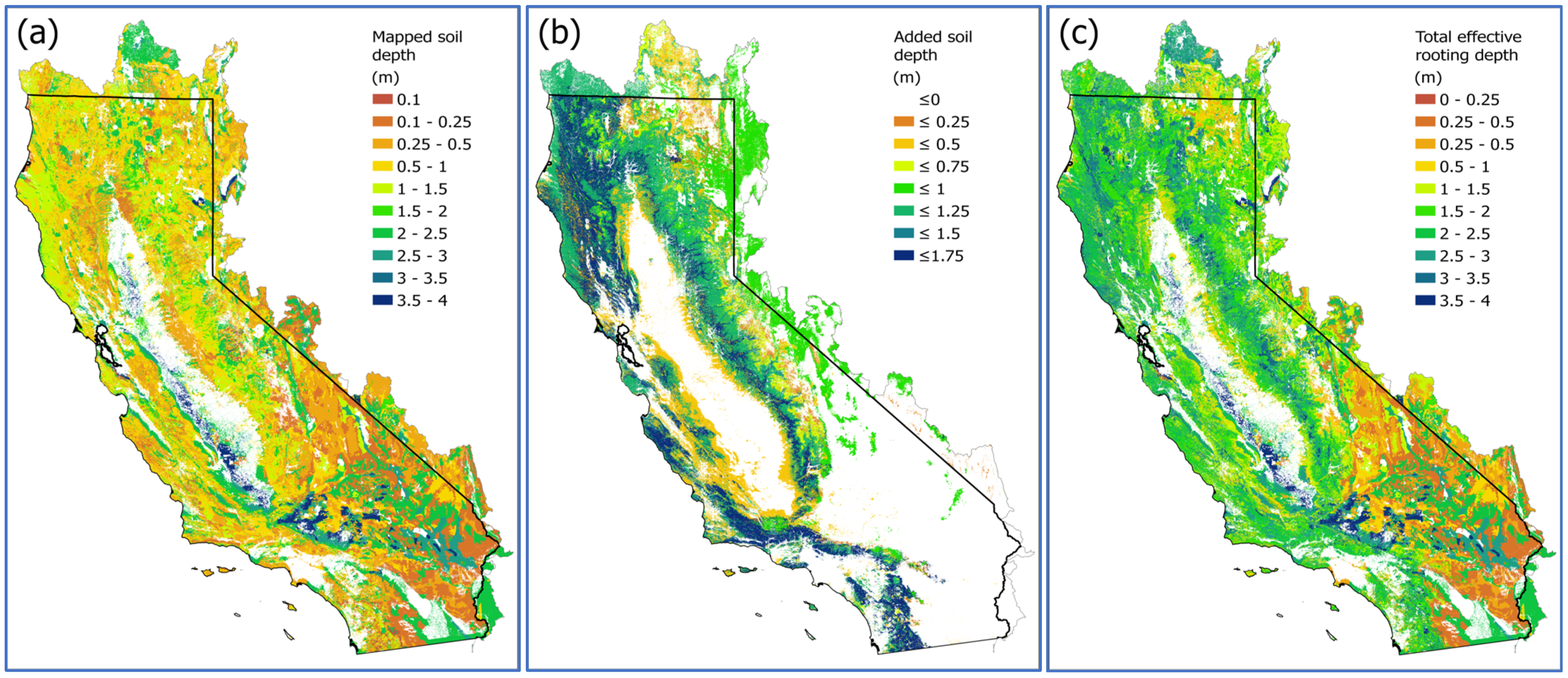
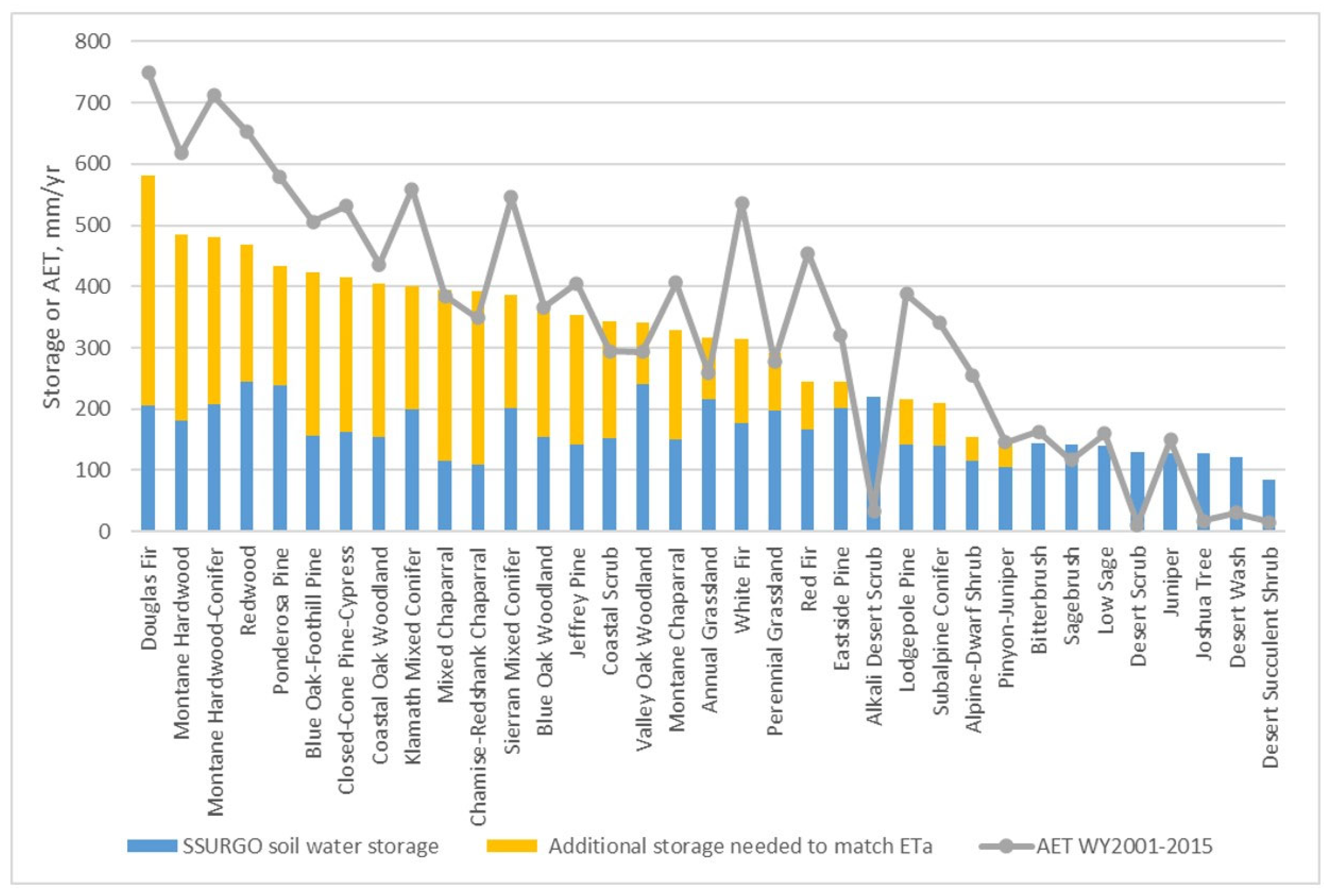
3.2. Effective Rooting Volume in Soil and Bedrock
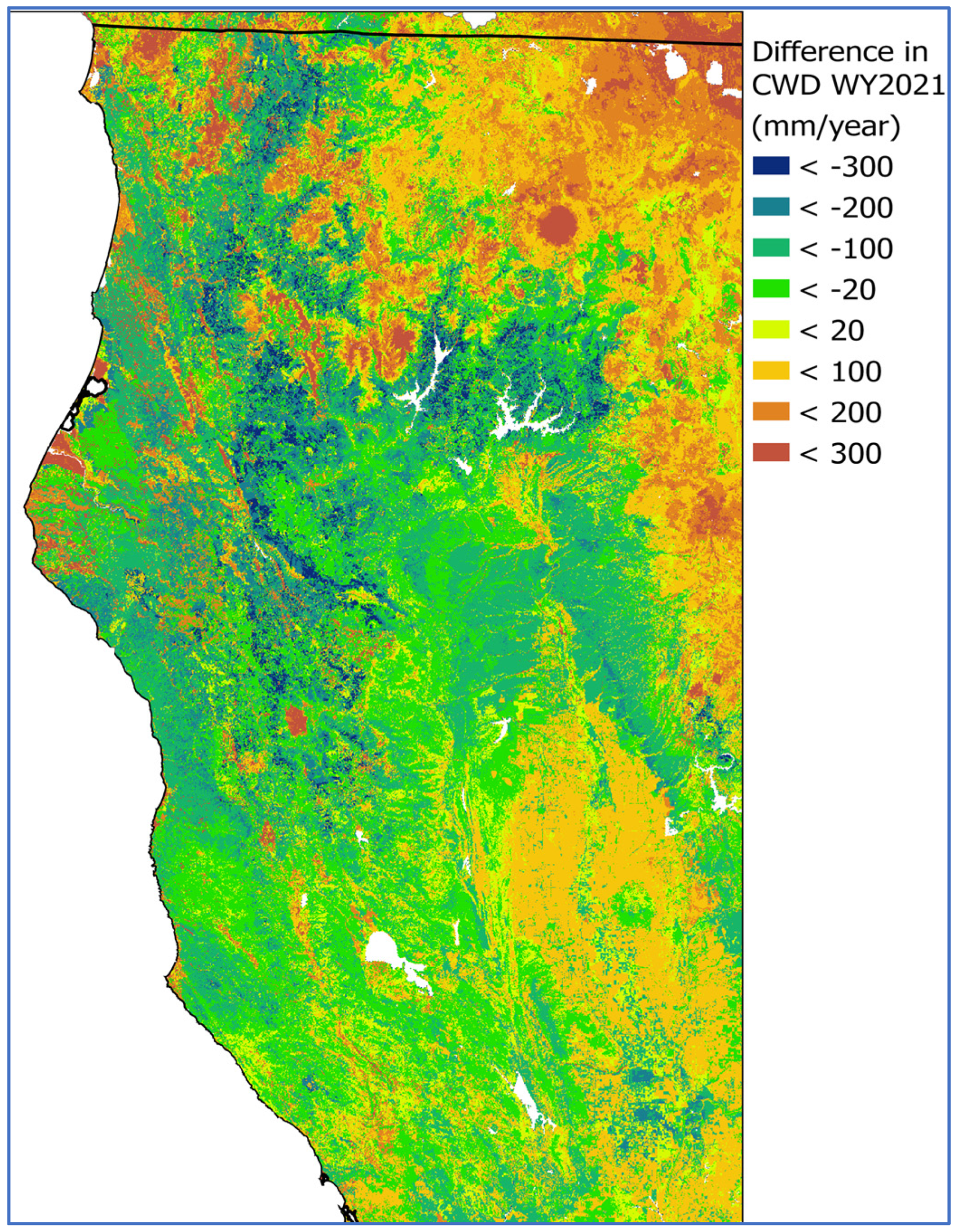
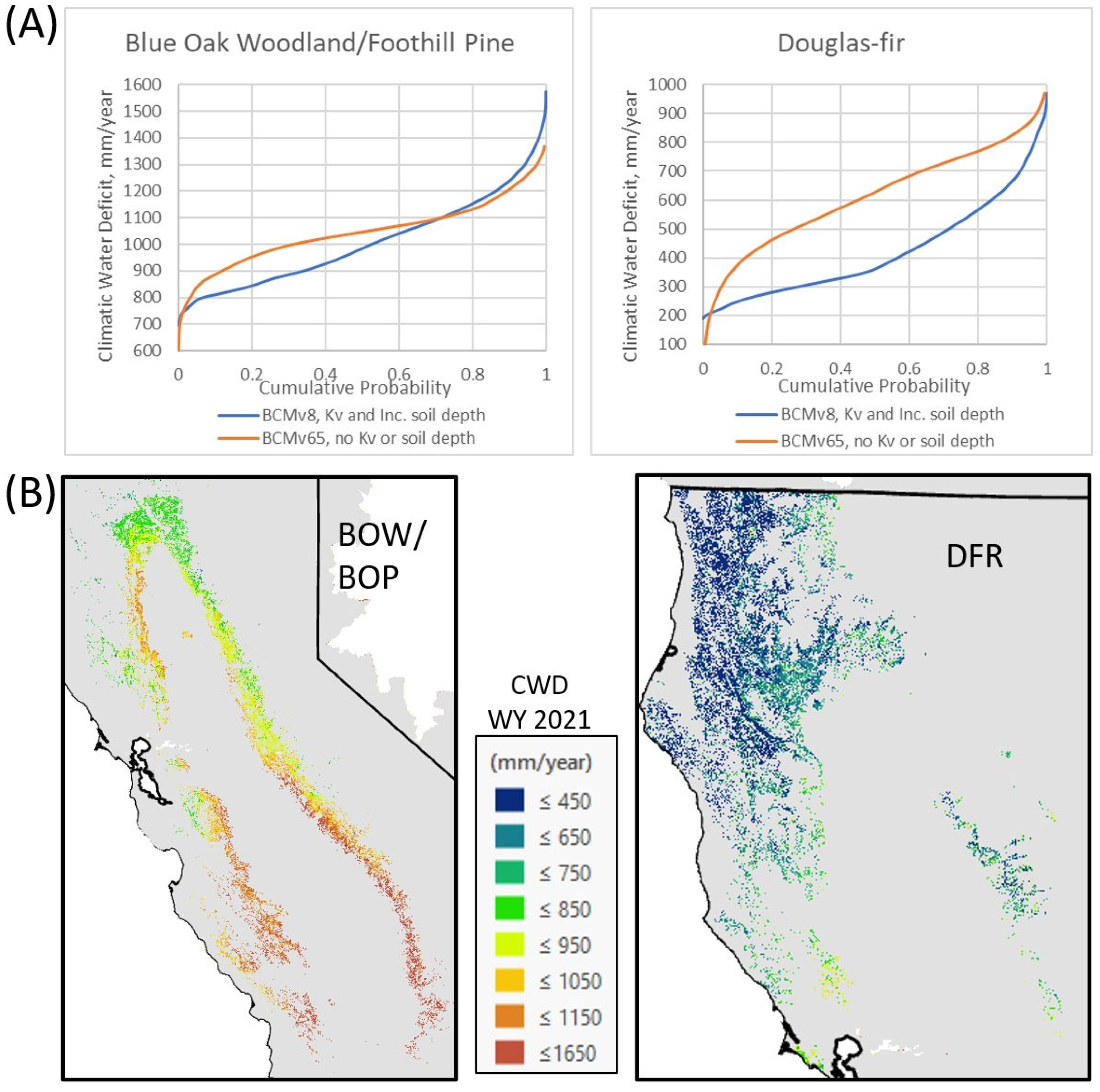
3.3. Comparisons of Model Simulation with and Without Kv Values and Addition of Effective Rooting Depth
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
A.1. Calculating Potential Evapotranspiration and Seasonality of Actual Evapotranspiration
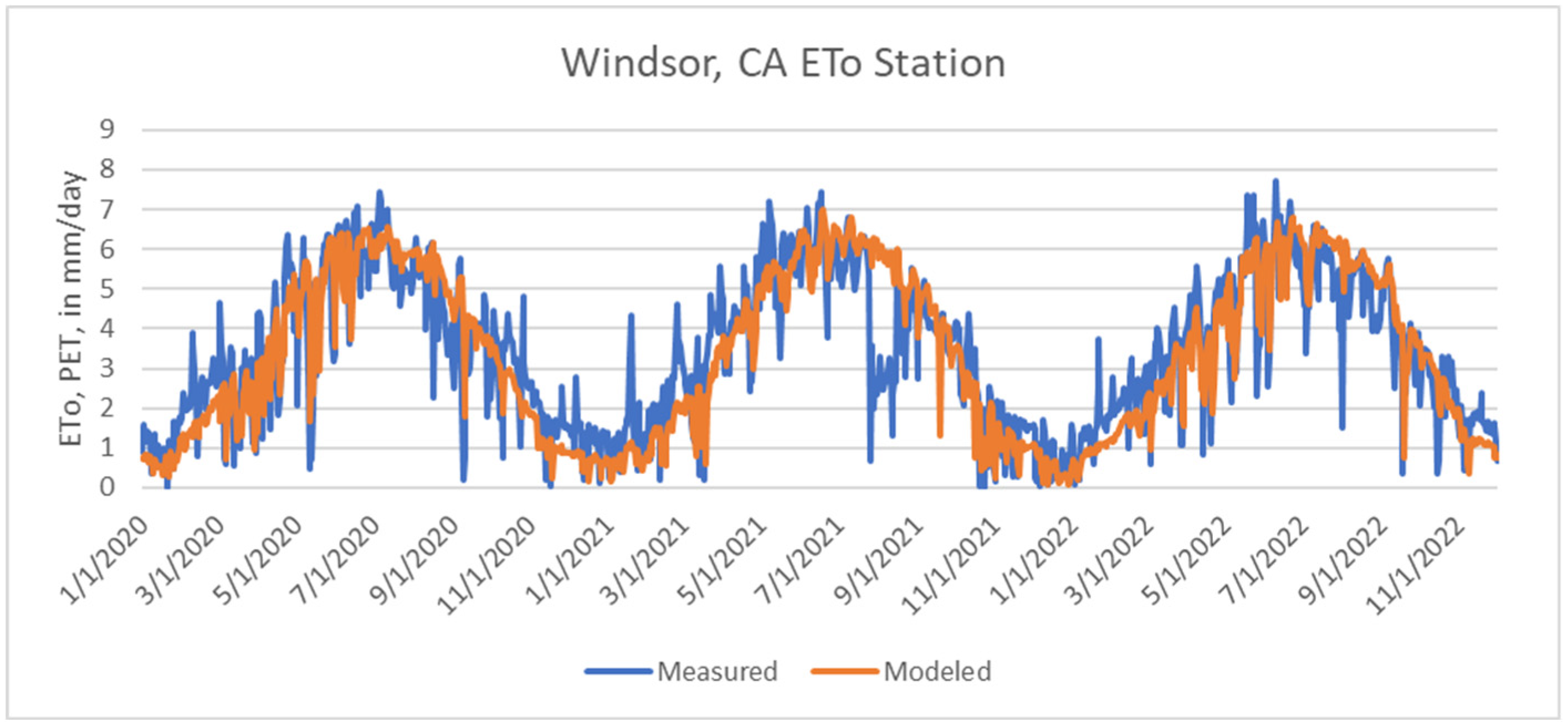
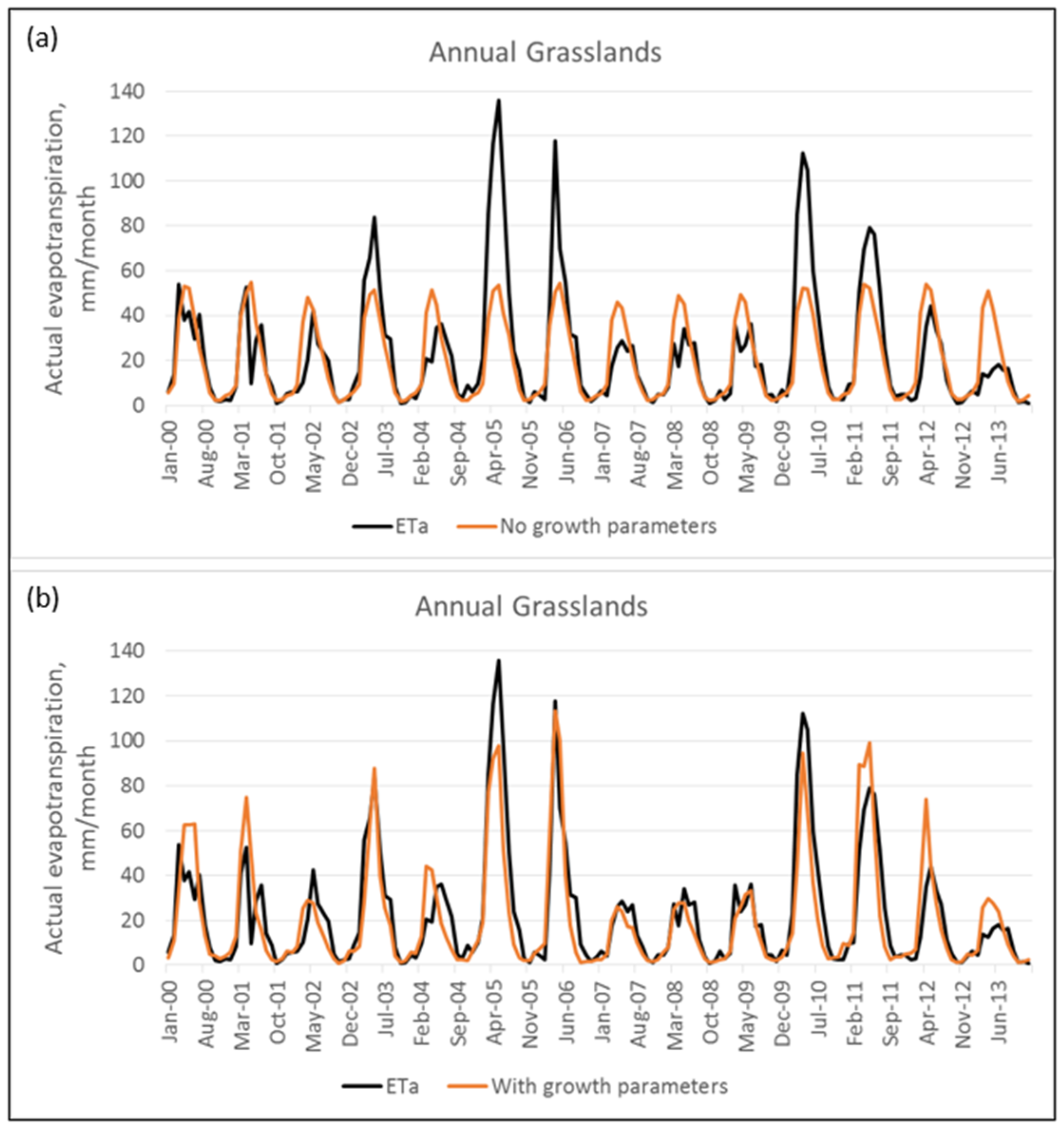

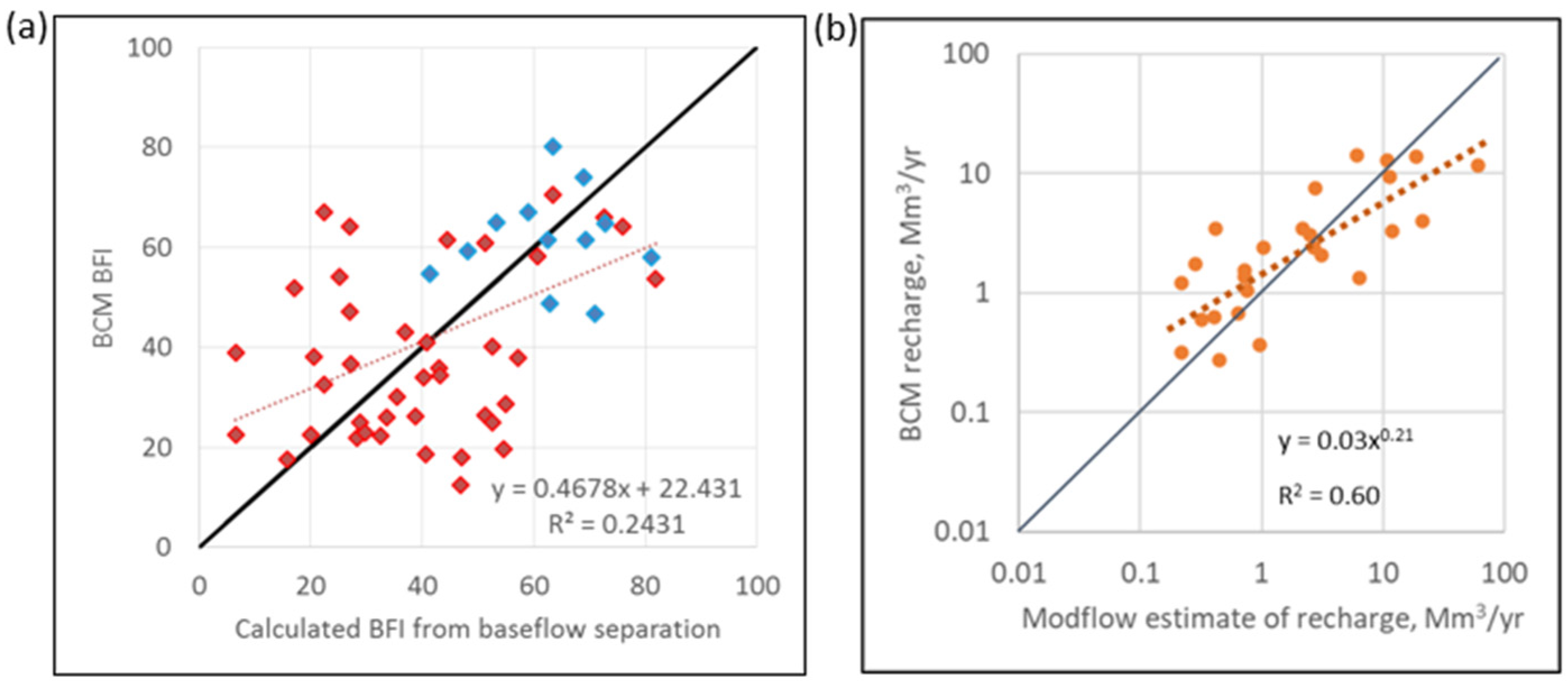
A.2. Calibration Results of BCM to Actual Evapotranspiration, Streamflow, Baseflow Index, and Groundwater Flow Model Estimates of Recharge


References
- McCormick, E.L.; Dralle, D.N.; Hahm, W.J.; Tune, A.K.; Schmidt, L.M.; Charwick, K.D.; Rempek, D.M. Widespread woody plant use of water stored in bedrock. Nature 2021, 597, 225–229. [Google Scholar] [CrossRef] [PubMed]
- McIntyre, P.J.; Thorne, J.H.; Dolanc, C.R.; Flint, A.L.; Flint, L.E.; Kelly, M.; Ackerly, D.D. 20th century shifts in forest structure in California: Denser forests, smaller trees, and increased dominance of oaks. Proc. Natl. Acad. Sci. USA 2015, 112, 1458–1463. [Google Scholar] [CrossRef] [PubMed]
- Flint, L.E.; Flint, A.L.; Stern, M.A.; Mayer, A.; Silver, W.; Casey, C.F.; Franco, F.; Byrd, K.; Sleeter, B. Increasing Soil Organic Carbon to Mitigate Greenhouse Gases and Increase Climate Resiliency for California. Calif. Nat. Resour. Agency California’s Fourth Clim. Change Assess. Tech. Rep. 2018, CCCA4-CNRA-2018-006, 83p. Available online: https://pubs.usgs.gov/publication/70199846 (accessed on 10 November 2024).
- Tian, W.; Liu, X.; Liu, C.; Bai, P. Investigation and simulations of changes in the relationship of precipitation-runoff in drought years. J. Hydrol. 2018, 565, 95–105. [Google Scholar] [CrossRef]
- Hahm, W.J.; Dralle, D.N.; Sanders, M.; Bryk, A.B.; Fauria, K.E.; Huang, M.H.; Hudson-Rasmussen, B.; Nelson, M.D.; Pedrazas, M.A.; Schmidt, L.; et al. Bedrock vadose zone storage dynamics under extreme drought: Consequences for plant water availability, recharge, and runoff. Water Resour. Res. 2022, 58, e2021WR031781. [Google Scholar] [CrossRef]
- Park, I.W.; Mann, M.L.; Flint, L.E.; Flint, A.L.; Moritz, M. Relationships of climate, human activity, and fire history to spatiotemporal variation in annual fire probability across California. PLoS ONE 2022, 16, e0254723. [Google Scholar]
- Thorne, J.H.; Moody, T.; Rosenberg, M. Forest Tree Mortality. In Indicators of Climate Change in California; Milanes, C., Kadir, T., Lock, B., Miller, G., Monserrat, L., Randles, K., Eds.; California Environmental Protection Agency: Sacramento, CA, USA, 2022. Available online: https://oehha.ca.gov/media/04treemortality.pdf (accessed on 10 November 2024).
- Chen, J.; McGuire, K.J.; Stewart, R.D. Effect of soil water-repellent layer depth on post-wildfire hydrological processes. Hydrol. Proc. 2020, 34, 270–283. [Google Scholar] [CrossRef]
- Maina, F.Z.; Siirila-Woodburn, E.R.; Vahmani, P. On the sensitivity of meteorological forcing resolution on hydrologic metrics. Energy 2019, 7, 4–5. [Google Scholar]
- Thorne, J.H.; Boynton, R.M.; Flint, L.E.; Flint, A.L. The magnitude and spatial patterns of historical and future hydrologic change in California’s watersheds. Ecosphere 2015, 6. Available online: https://esajournals.onlinelibrary.wiley.com/doi/10.1890/ES14-00300.1 (accessed on 10 November 2024).
- Thorne, J.H.; Morgan, B.J.; Kennedy, J.A. Vegetation Change over 60 Years in the Central Sierra Nevada. Madroño 2008, 55, 223–237. [Google Scholar] [CrossRef]
- Goulden, M.L.; Bales, R.C. Mountain runoff vulnerability to increased evapotranspiration with vegetation expansion. Proc. Natl. Acad. Sci. USA 2014, 111, 14071–14075. [Google Scholar] [CrossRef]
- Ehlert, R.S.; Hahm, W.J.; Dralle, D.N.; Rempe, D.M.; Allen, D.M. Bedrock controls on water and energy partitioning. Water Resour. Res. 2024, 60, e2023WR036719. [Google Scholar] [CrossRef]
- Hahm, W.J.; Dralle, D.N.; Lapides, D.A.; Ehlert, R.S.; Rempe, D.M. Geologic controls on apparent root-zone storage capacity. Water Resour. Res. 2024, 60, e2023WR035362. [Google Scholar] [CrossRef]
- Hahm, W.J.; Rempe, D.M.; Dralle, D.N.; Dawson, T.E.; Lovill, S.M.; Bryk, A.B. Lithologically controlled subsurface critical zone thickness and water storage capacity determine regional plant community composition. Water Resour. Res. 2019, 55, 3028–3055. [Google Scholar] [CrossRef]
- Eggemeyer, K.D.; Schwinning, S. Biogeography of woody encroachment: Why is mesquite excluded from shallow soils? Ecohydrology 2009, 2, 81–87. [Google Scholar] [CrossRef]
- Cui, G.; Guo, W.; Goulden, M.; Bales, R. MODIS-based modeling of evapotranspiration from woody vegetation supported by root-zone water storage. Remote Sens. Environ. 2024, 303, 114000. [Google Scholar] [CrossRef]
- Doorenbos, J.; Pruitt, W.O. Crop Water Requirements; FAO Irrigation Drainage Paper 24 (Rev.); Food and Agriculture Organization of the United Nations: Rome, Italy, 1977. [Google Scholar]
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. Crop Evapotranspiration–Guidelines for Computing Crop Water Requirements; FAO Irrigation Drainage Paper 56; Food and Agriculture Organization of the United Nations: Rome, Italy, 1998. [Google Scholar]
- Lascano, R.J. A general system to measure and calculate daily crop water use. Agron. J. 2000, 92, 821–832. [Google Scholar] [CrossRef]
- Mata-González, R.; McLendon, T.; Martin, D.W. The Inappropriate Use of Crop Transpiration Coefficients (Kc) to Estimate Evapotranspiration in Arid Ecosystems: A Review. Arid Land Res. Manag. 2005, 19, 285–295. [Google Scholar] [CrossRef]
- Wyatt, B.M.; Ochsner, T.E.; Zou, C.B. Estimating root zone soil moisture across diverse land cover types by integrating in-situ and remotely sensed data. Agric. For. Meteor. 2021, 15, 108471. [Google Scholar] [CrossRef]
- Klos, P.; Goulden, M.; Riebe, C.; Tague, C.; O’Geen, A.; Flinchum, B.; Safeeq, M.; Conklin, M.; Hart, S.; Berhe, A.; et al. Subsurface plant-accessible water in mountain ecosystems with a Mediterranean climate. WIREs Water 2018, 5, e1277. [Google Scholar] [CrossRef]
- Flint, A.L.; Childs, S.W. Physical properties of rock fragments and their effect on available water in skeletal soils. In Erosion and Productivity of Soils Containing Rock Fragments; Special Publication; Soil Science Society America: Madison, WI, USA, 1984; Volume 13, pp. 91–103. [Google Scholar]
- Ichii, K.; Wang, W.; Hashimoto, H.; Yang, F.; Votava, P.; Michaelis, A.; Nemani, R. Refinement of rooting depths using satellite-based evapotranspiration seasonality for ecosystem modeling in California. Agric. For. Meteor. 2009, 149, 1907–1918. [Google Scholar] [CrossRef]
- Reitz, M.; Sanford, W.E.; Senay, G.B.; Cazenas, J. Annual Estimates of Recharge, Quick-Flow Runoff, and Evapotranspiration for the Contiguous US Using Empirical Regression Equations. J. Amer. Water Resour. Ass. 2017, 53, 961–983. [Google Scholar] [CrossRef]
- Flint, L.E.; Flint, A.L.; Stern, M.A. The Basin Characterization Model version 8–A Regional Water Balance Software Package. U.S. Geol. Surv. Tech. Methods 2021, 6–H1, 85. [Google Scholar] [CrossRef]
- Daly, C.; Halbleib, M.; Smith, J.I.; Gibson, W.P.; Doggett, M.K.; Taylor, G.H.; Curtis, J.; Pasteris, P.P. Physiographically sensitive mapping of climatological temperature and precipitation across the conterminous United States. Inter. J. Climatol. J. R. Meteor. Soc. 2008, 28, 2031–2064. [Google Scholar] [CrossRef]
- Flint, L.E.; Flint, A.L. Downscaling climate change scenarios for ecologic applications. Ecol. Process 2012, 1, 1–15. [Google Scholar]
- Saxton, K.E.; Rawls, W.J. Soil water characteristic estimates by texture and organic matter for hydrologic solutions. Soil Sci. Soc. Am. J. 2006, 70, 1569–1578. [Google Scholar] [CrossRef]
- Flint, L.E.; Flint, A.L.; Thorne, J.H.; Boynton, R. Fine-scale hydrological modeling for climate change applications; using watershed calibrations to assess model performance for landscape projections. Ecol. Process. 2013, 2, 1–25. [Google Scholar] [CrossRef]
- Mu, Q.; Zhao, M.; Running, S.W. Improvements to a MODIS global terrestrial evapotranspiration algorithm. Remote Sens. Environ. 2011, 115, 1781–1800. [Google Scholar] [CrossRef]
- Senay, G.B.; Bohms, S.; Singh, R.K.; Gowda, P.H.; Velpuri, N.M.; Alemu, H.; Verdin, J.P. Operational evapotranspiration mapping using remote sensing and weather datasets: A new parameterization for the SSEB approach. J. Am. Water Resour. Assoc. 2013, 49, 577–591. [Google Scholar] [CrossRef]
- California Department of Forestry and Fire Protection. California Vegetation by Wildlife Habitat Relationship Type; 2015. Available online: https://www.fire.ca.gov/Home/What-We-Do/Fire-Resource-Assessment-Program/GIS-Mapping-and-Data-Analytics (accessed on 11 November 2024).
- California Department of Fish and Wildlife and California Interagency Wildlife Task Group. Standards and Guidelines for CWHR Species Models. California Wildlife Habitat Relationship System; Technical Report No. 31; California Department of Fish and Game: Sacramento, CA, USA, 2014.
- Federal Geographic Data Committee. National Vegetation Classification Standard, Version 2; FGDC Document Number FGDC-STD-005-2008; U.S. Geological Survey: Reston, VA, USA, 2008. Available online: https://fgdc.gov/standards/projects/vegetation/NVCS_V2_FINAL_2008-02.pdf (accessed on 11 November 2024).
- U.S. Geological Survey. Gap Analysis Project (GAP), GAP/LANDFIRE National Terrestrial Ecosystems. U.S. Geol. Surv. Data Release. 2016. Available online: https://www.sciencebase.gov/catalog/item/573cc51be4b0dae0d5e4b0c5 (accessed on 4 April 2013).
- Goulden, M.L.; Anderson, R.G.; Bales, R.C.; Kelly, A.E.; Meadows, M.; Winston, G.C. Evapotranspiration along an elevation gradient in California’s Sierra Nevada. J. Geophys. Res. Biogeosci. 2012, 117, 1–13. [Google Scholar] [CrossRef]
- Cannon, W.A. Specialization in vegetation and in environment in California. In Plant World; Wiley: Hoboken, NJ, USA, 1914; Volume 17, pp. 223–237. [Google Scholar]
- McMinn, R.G. Characteristics of Douglas-fir root systems. Can. J. Bot. 1963, 41, 105–122. [Google Scholar] [CrossRef]
- Canadell, J.; Jackson, R.; Ehleringer, J.; Mooney, H.; Sala, O.; Shulze, E. Maximum rooting depth of vegetation types at the global scale. Oecologia 1996, 108, 583–595. [Google Scholar] [CrossRef]
- Howes, D.J.; Fox, P.; Hutton, P.H. Evapotranspiration from natural vegetation in the Central Valley of California: Monthly grass reference-based vegetation coefficients and the dual crop coefficient approach. J. Hydrol. Eng. 2015, 20, 04015004. [Google Scholar] [CrossRef]
- Bales, R.C.; Hopmans, J.W.; O’Geen, A.T.; Meadows, M.; Hartsough, P.C.; Kirchner, P.; Hunsaker, C.T.; Beaudette, D. Soil moisture response to snowmelt and rainfall in a Sierra Nevada mixed-conifer forest. Vadose Zone J. 2011, 10, 786–799. [Google Scholar] [CrossRef]
- McLaughlin, B.C.; Blakey, R.; Weitz, A.P.; Feng, X.; Brown, B.J.; Ackerly, D.D.; Dawson, T.E.; Thompson, S.E. Weather underground: Subsurface hydrologic processes mediate tree vulnerability to extreme climatic drought. Glob. Change Biol. 2020, 26, 3091–3107. [Google Scholar] [CrossRef] [PubMed]
- Thorne, J.H.; Choe, H.; Boynton, R.M.; Bjorkman, J.; Albright, W.; Nydick, K.; Flint, A.L.; Flint, L.E.; Schwartz, M.W. The impact of climate change uncertainty on California’s vegetation and adaptation management. Ecosphere 2017, 8, e02021. [Google Scholar] [CrossRef]
- Rapacciuolo, G.; Maher, S.P.; Schneider, A.C.; Hammond, T.T.; Jabis, M.D.; Walsh, R.E.; Iknayan, K.J.; Walden, G.K.; Oldfather, M.F.; Ackerly, D.D.; et al. Beyond a warming fingerprint: Individualistic biogeographic responses to heterogeneous climate change in California. Glob. Change Biol. 2014, 20, 2841–2855. [Google Scholar] [CrossRef]
- Wieczorek, M.E. Area- and Depth-Weighted Averages of Selected SSURGO Variables for the Conterminous United States and District of Columbia. USGS Data Ser. 866 2014. Available online: http://water.usgs.gov/lookup/getspatial?ds866_ssurgo_variables (accessed on 3 June 2017).
- Wieczorek, M.E. STATSGO2 Area- and Depth-Weighted Averages of Selected STATSGO2 Variables. Sci. Base-Cat. 2015. Available online: https://www.sciencebase.gov/catalog/item/54b97608e4b043905e00fc9d (accessed on 3 June 2017).
- Flint, A.L.; Childs, S.W. Calculation of solar radiation in mountainous terrain. Agric. For. Meteorol. 1987, 40, 233–249. [Google Scholar] [CrossRef]
- Priestley, C.H.B.; Taylor, R.J. On the assessment of surface heat flux and evaporation using large-scale parameters. Mon. Weather Rev. 1972, 100, 81–92. [Google Scholar] [CrossRef]
- Bristow, K.L.; Campbell, G.S. On the relationship between incoming solar radiation and daily maximum and minimum temperature. Agric. For. Meteorol. 1984, 31, 159–166. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vegetation Type | Percent of Area | Added Soil Depth (m) | Monthly Kv Coefficients | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Oct | Nov | Dec | Jan | Feb | Mar | Apr | May | Jun | Jul | Aug | Sep | |||
| Alkali Desert Scrub | 2.7 | 0.00 | 0.066 | 0.086 | 0.125 | 0.142 | 0.193 | 0.445 | 0.568 | 0.626 | 0.516 | 0.391 | 0.290 | 0.164 |
| Alpine-Dwarf Shrub | 0.1 | 0.25 | 0.214 | 0.139 | 0.048 | 0.077 | 0.212 | 0.262 | 0.426 | 0.365 | 0.340 | 0.213 | 0.197 | 0.227 |
| Annual Grassland | 9.0 | 2.00 | 0.219 | 0.176 | 0.308 | 0.246 | 0.658 | 0.858 | 0.830 | 0.681 | 0.601 | 0.576 | 0.063 | 0.358 |
| Bitterbrush | 0.5 | 0.00 | 0.022 | 0.137 | 0.302 | 0.303 | 0.207 | 0.342 | 0.184 | 0.191 | 0.222 | 0.170 | 0.098 | 0.036 |
| Blue Oak Woodland | 2.3 | 1.30 | 0.078 | 0.120 | 0.173 | 0.188 | 0.206 | 0.533 | 0.650 | 0.734 | 0.582 | 0.467 | 0.342 | 0.189 |
| Blue Oak-Foothill Pine | 0.8 | 1.25 | 0.121 | 0.131 | 0.123 | 0.170 | 0.149 | 0.337 | 0.448 | 0.621 | 0.558 | 0.496 | 0.413 | 0.276 |
| Chamise-Redshank Chaparral | 1.1 | 2.10 | 0.170 | 0.180 | 0.250 | 0.320 | 0.410 | 0.760 | 0.790 | 0.880 | 0.910 | 0.80 | 0.700 | 0.520 |
| Closed-Cone Pine-Cypress | 0.1 | 1.50 | 0.415 | 0.348 | 0.233 | 0.417 | 0.510 | 0.676 | 0.722 | 0.897 | 0.949 | 0.875 | 0.819 | 0.627 |
| Coastal Oak Woodland | 1.1 | 1.25 | 0.130 | 0.227 | 0.311 | 0.382 | 0.352 | 0.491 | 0.515 | 0.488 | 0.421 | 0.327 | 0.270 | 0.158 |
| Coastal Scrub | 1.5 | 1.50 | 0.059 | 0.062 | 0.146 | 0.177 | 0.211 | 0.353 | 0.346 | 0.307 | 0.262 | 0.230 | 0.171 | 0.097 |
| Desert Scrub | 19.6 | 1.00 | 0.440 | 0.340 | 0.371 | 0.244 | 0.274 | 0.486 | 0.504 | 0.714 | 0.840 | 0.840 | 0.714 | 0.645 |
| Desert Succulent Shrub | 0.1 | 0.00 | 0.001 | 0.005 | 0.012 | 0.023 | 0.026 | 0.056 | 0.021 | 0.005 | 0.002 | 0.002 | 0.004 | 0.006 |
| Desert Wash | 0.7 | 1.50 | 0.043 | 0.042 | 0.197 | 0.224 | 0.129 | 0.231 | 0.184 | 0.188 | 0.217 | 0.215 | 0.189 | 0.119 |
| Douglas Fir | 2.7 | 2.00 | 0.496 | 0.457 | 0.339 | 0.556 | 0.521 | 0.693 | 0.647 | 0.797 | 0.820 | 0.874 | 0.864 | 0.717 |
| Eastside Pine | 2.1 | 0.50 | 0.067 | 0.182 | 0.188 | 0.342 | 0.272 | 0.449 | 0.330 | 0.390 | 0.371 | 0.374 | 0.321 | 0.167 |
| Jeffrey Pine | 0.4 | 1.50 | 0.002 | 0.086 | 0.131 | 0.272 | 0.221 | 0.449 | 0.395 | 0.435 | 0.428 | 0.450 | 0.409 | 0.288 |
| Joshua Tree | 0.8 | 1.80 | 0.625 | 0.566 | 0.166 | 0.404 | 0.338 | 0.658 | 0.574 | 0.604 | 0.629 | 0.625 | 0.625 | 0.625 |
| Juniper | 0.9 | 1.50 | 0.014 | 0.131 | 0.133 | 0.230 | 0.216 | 0.283 | 0.185 | 0.277 | 0.215 | 0.162 | 0.090 | 0.032 |
| Klamath Mixed Conifer | 2.2 | 1.00 | 0.280 | 0.236 | 0.204 | 0.392 | 0.353 | 0.547 | 0.544 | 0.640 | .0620 | 0.696 | 0.668 | 0.533 |
| Lodgepole Pine | 0.4 | 0.75 | 0.083 | 0.144 | 0.126 | 0.191 | 0.151 | 0.428 | 0.569 | 0.613 | 0.438 | 0.419 | 0.403 | 0.239 |
| Low Sage | 1. | 0.40 | 0.040 | 0.052 | 0.028 | 0.099 | 0.092 | 0.191 | 0.303 | .0136 | 0.106 | 0.111 | 0.109 | 0.086 |
| Mixed Chaparral | 4.6 | 0.75 | 0.126 | 0.107 | 0.172 | 0.222 | 0.257 | 0.437 | 0.472 | 0.576 | 0.551 | 0.482 | 0.379 | 0.245 |
| Montane Chaparral | 1.5 | 0.75 | 0.096 | 0.136 | 0.141 | 0.224 | 0.185 | 0.363 | 0.381 | 0.506 | 0.484 | 0.504 | 0.436 | 0.276 |
| Montane Hardwood | 3.2 | 1.00 | 0.270 | 0.224 | 0.158 | 0.227 | 0.282 | 0.471 | 0.534 | 0.741 | 0.758 | 0.718 | 0.652 | 0.471 |
| Montane Hardwood-Conifer | 2.5 | 1.25 | 0.398 | 0.472 | 0.379 | 0.533 | 0.448 | 0.589 | 0.610 | 0.755 | 0.793 | 0.832 | 0.803 | 0.609 |
| Perennial Grassland | 0.3 | 0.75 | 0.034 | 0.105 | 0.127 | 0.227 | 0.242 | 0.377 | 0.298 | 0.377 | 0.314 | 0.277 | 0.193 | 0.099 |
| Pinyon Juniper | 2.2 | 1.20 | 0.034 | 0.044 | 0.118 | 0.173 | 0.152 | 0.248 | 0.148 | 0.218 | 0.225 | 0.212 | 0.157 | 0.077 |
| Ponderosa Pine | 0.6 | 1.40 | 0.214 | 0.171 | 0.116 | 0.232 | 0.273 | 0.477 | 0.437 | .0309 | 0.631 | 0.646 | 0.607 | 0.472 |
| Red Fir | 1.2 | 0.75 | 0.101 | 0.133 | 0.121 | 0.212 | 0.193 | 0.482 | 0.591 | .0689 | 0.520 | 0.513 | 0.478 | 0.324 |
| Redwood | 1. | 1.40 | 0.485 | 0.474 | 0.363 | 0.438 | 0.475 | 0.565 | 0.561 | 0.668 | 0.71 | .0777 | 0.807 | 0.657 |
| Sagebrush | 5.9 | 1.50 | 0.017 | 0.138 | 0.298 | 0.034 | 0.266 | 0.838 | 0.900 | 0.890 | 0.881 | 0.487 | 0.686 | 0.619 |
| Sierran Mixed Conifer | 3.9 | 0.50 | 0.206 | 0.167 | 0.147 | 0.282 | 0.308 | 0.554 | 0.491 | 0.580 | 0.569 | 0.594 | 0.582 | 0.455 |
| Subalpine Conifer | 0.9 | 0.75 | 0.089 | 0.122 | 0.098 | 0.163 | 0.125 | 0.347 | 0.462 | 0.529 | 0.410 | 0.394 | 0.378 | 0.215 |
| Valley Oak Woodland | 0.1 | 0.75 | 0.063 | 0.058 | 0.087 | 0.093 | 0.108 | 0.218 | 0.267 | 0.363 | .0317 | 0.275 | 0.212 | 0.132 |
| White Fir | 0.8 | 0.75 | 0.212 | 0.202 | 0.174 | 0.391 | 0.367 | 0.633 | 0.600 | 0.657 | 0.568 | 0.622 | 0.607 | 0.474 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Flint, A.L.; Flint, L.E.; Stern, M.A.; Ackerly, D.D.; Boynton, R.; Thorne, J.H. Characterizing Soil and Bedrock Water Use of Native California Vegetation. Hydrology 2024, 11, 211. https://doi.org/10.3390/hydrology11120211
Flint AL, Flint LE, Stern MA, Ackerly DD, Boynton R, Thorne JH. Characterizing Soil and Bedrock Water Use of Native California Vegetation. Hydrology. 2024; 11(12):211. https://doi.org/10.3390/hydrology11120211
Chicago/Turabian StyleFlint, Alan L., Lorraine E. Flint, Michelle A. Stern, David D. Ackerly, Ryan Boynton, and James H. Thorne. 2024. "Characterizing Soil and Bedrock Water Use of Native California Vegetation" Hydrology 11, no. 12: 211. https://doi.org/10.3390/hydrology11120211
APA StyleFlint, A. L., Flint, L. E., Stern, M. A., Ackerly, D. D., Boynton, R., & Thorne, J. H. (2024). Characterizing Soil and Bedrock Water Use of Native California Vegetation. Hydrology, 11(12), 211. https://doi.org/10.3390/hydrology11120211