Zebrafish as an Emerging Model for Bioassay-Guided Natural Product Drug Discovery for Neurological Disorders




Abstract
1. Introduction
2. Neurodegenerative Diseases
3. Using the Zebrafish Model for Neurological Disorders
3.1. Zebrafish and Alzheimer’s Disease
3.2. Zebrafish and Parkinson’s Disease
3.3. Zebrafish and Huntington’s and Other Polyglutamine Diseases
3.4. Zebrafish and Amyotrophic Lateral Sclerosis (ALS)
3.5. Zebrafish and Schizophrenia
3.6. Zebrafish and Epilepsy
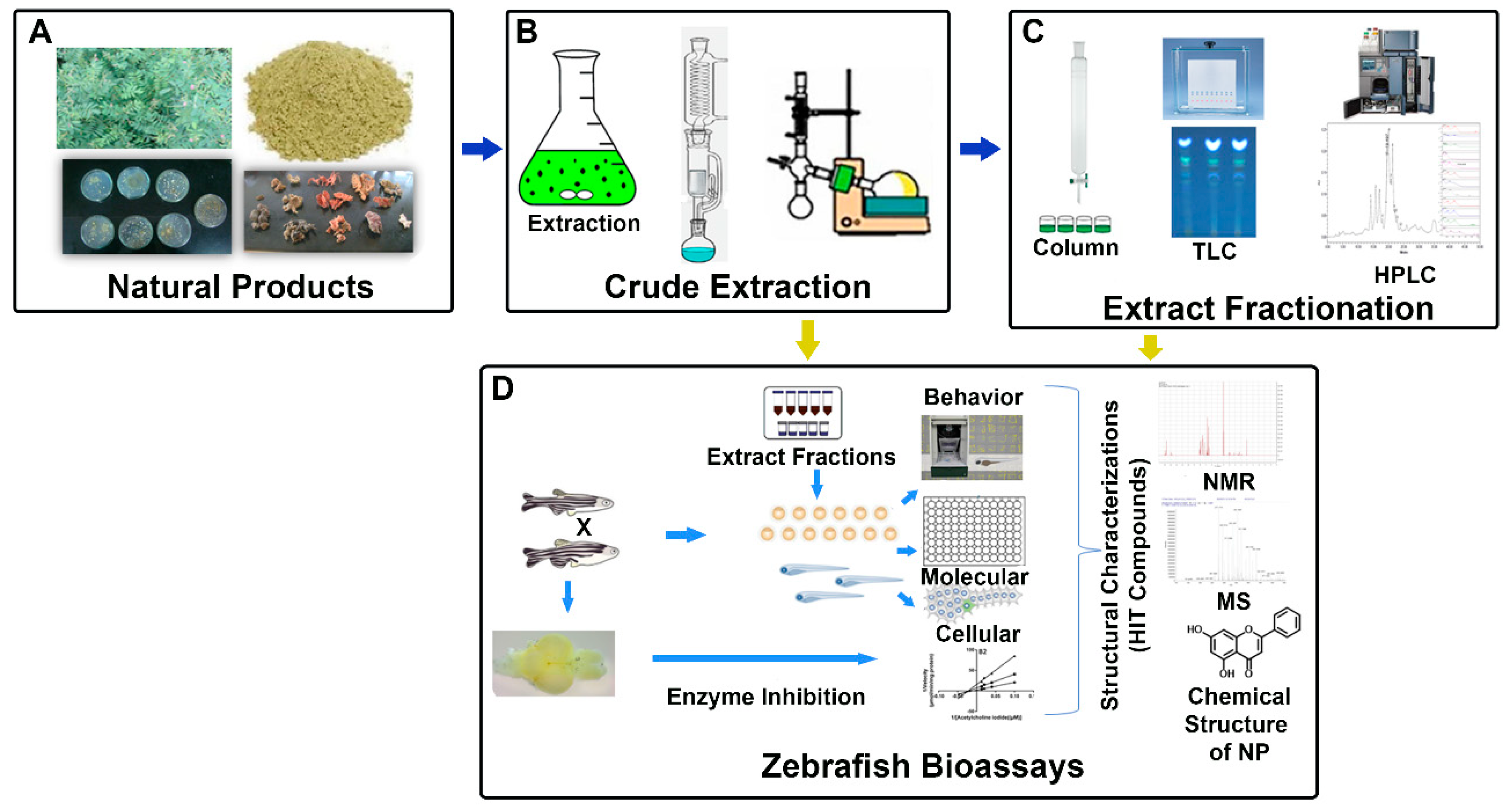
4. Zebrafish Bioassay-Guided Isolation of Natural Product Drug Discovery
5. Development of Zebrafish Models for Neurological Disorders
5.1. Transgenic Zebrafish Models
5.2. Generation of a Neurodegenerative Model Using Amyloid-β42 (Aβ42) in the Adult Zebrafish Brain
5.3. Zebrafish Cell Culture-Based Neurodegenerative Disease Models
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Devine, M.J.; Plun-Favreau, H.; Wood, N.W. Parkinson’s disease and cancer: Two wars, one front. Nat. Rev. Cancer 2011, 11, 812–823. [Google Scholar] [CrossRef]
- Tabarés-seisdedos, R.; Rubenstein, J.L. Inverse cancer comorbidity a serendipitous opportunity to gain insight into CNS disorders. Nat. Rev. Neurosci. 2013, 14, 293–304. [Google Scholar] [CrossRef]
- Behrens, M.I.; Silva, M.; Salech, F.; Ponce, D.P.; Merino, D.; Sinning, M.; Xiong, C.; Roe, M.C.; Quest, A.F.G. Inverse susceptibility to oxidative death of lymphocytes obtained from Alzheimer’s patients and skin cancer survivors: Increased apoptosis in Alzheimer’s and reduced necrosis in cancer. J. Gerontol. Ser. A Biol Sci. Med. Sci. 2012, 67, 1036–1040. [Google Scholar] [CrossRef] [PubMed]
- Hilario, E.; Álvarez, A.; Domínguez, A.; Suarez-Merino, B.; Goñi-de-Cerio, F. Central nervous system diseases and the role of the blood-brain barrier in their treatment. Neurosci. Discov. 2013, 1, 1–3. [Google Scholar]
- Gilmore, J.L.; Yi, X.; Quan, L.; Kabanov, A.V. Novel Nanomaterials for Clinical Neuroscience. J. Neuroimmune Pharmacol. 2008, 3, 83–94. [Google Scholar] [CrossRef]
- Rosén, J.; Gottfries, J.; Muresan, S.; Backlund, A.; Oprea, T.I. Novel Chemical Space Exploration via Natural Products. J. Med. Chem. 2009, 52, 1953–1962. [Google Scholar] [CrossRef]
- Koehn, F.E.; Carter, G.T. The evolving role of natural products in drug discovery. Nat. Rev. Drug Discov. 2005, 4, 206–220. [Google Scholar] [CrossRef] [PubMed]
- Harvey, A.L. Natural products in drug discovery. Drug Discov. Today 2008, 13, 894–901. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural Products As Sources of New Drugs over the 30 Years from 1981 to 2010. J. Nat. Prod. 2012, 75, 311–335. [Google Scholar] [CrossRef]
- Macarron, R.; Banks, M.N.; Bojanic, D.; Burns, D.J.; Cirovic, D.A.; Garyantes, T.; Green, D.V.S.; Hertzberg, R.P.; Janzen, W.P.; Paslay, J.W.; et al. Impact of high-throughput screening in biomedical research. Nat. Rev. Drug Discov. 2011, 10, 188–195. [Google Scholar] [CrossRef]
- Welsch, M.E.; Snyder, S.A.; Stockwell, B.R. Privileged scaffolds for library design and drug discovery. Curr. Opin. Chem. Biol. 2010, 14, 347–361. [Google Scholar] [CrossRef] [PubMed]
- Cordell, G.A. Biodiversity and drug discovery—A symbiotic relationship. Phytochemistry 2000, 55, 463–480. [Google Scholar] [CrossRef]
- Vuorela, P.; Tammela, P.; Rauha, J.; Saikku, P.; Leinonen, M.; Vuorela, H. Natural Products in the Process of Finding New Drug Candidates. Curr. Med. Chem. 2004, 11, 1375–1389. [Google Scholar] [CrossRef]
- Testa, R.; Bonfigli, A.; Genovese, S.; De Nigris, V.; Ceriello, A. The Possible Role of Flavonoids in the Prevention of Diabetic Complications. Nutrients 2016, 8, 310. [Google Scholar] [CrossRef] [PubMed]
- Zon, L.I.; Peterson, R.T. In vivo drug discovery in the zebrafish. Nat. Rev. Drug Discov. 2005, 4, 35–44. [Google Scholar] [CrossRef]
- Crawford, A.D.; Esguerra, C.V.; de Witte, P.A.M. Fishing for Drugs from Nature: Zebrafish as a Technology Platform for Natural Product Discovery. Planta Med. 2008, 74, 624–632. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.-F.; Yoo, H.-D.; Williams, C.T.; Garo, E.; Cremin, P.A.; Zeng, L.; Vervoort, H.C.; Lee, C.M.; Hart, S.M.; Goering, M.G.; et al. Miniaturization of the Structure Elucidation of Novel Natural Products—Two Trace Antibacterial Acylated Caprylic Alcohol Glycosides from Arctostaphylos pumila. Planta Med. 2005, 71, 176–180. [Google Scholar] [CrossRef] [PubMed]
- Choi, D.-K.; Lee, K.; More, S.V.; Kumar, H.; Kang, S.M.; Song, S.-Y. Advances in Neuroprotective Ingredients of Medicinal Herbs by Using Cellular and Animal Models of Parkinson’s Disease. Evid. Based Complement. Altern Med. 2013, 2013, 1–15. [Google Scholar]
- Zhang, Z.J.; Cheang, L.C.V.; Wang, M.W.; Lee, S.M.Y. Quercetin exerts a neuroprotective effect through inhibition of the iNOS/NO system and pro-inflammation gene expression in PC12 cells and in zebrafish. Int. J. Mol. Med. 2011, 27, 195–203. [Google Scholar]
- Chan, H.M.; Guo, B.J.; Sa, F.; Li, S.; Lee, S.M.Y.; Zhang, Z.J.; Zheng, Y. Pharmacokinetic Study and Optimal Formulation of New Anti-Parkinson Natural Compound Schisantherin, A. Parkinsons Dis. 2015, 2015, 1–7. [Google Scholar]
- Löscher, W.; Potschka, H. Role of drug efflux transporters in the brain for drug disposition and treatment of brain diseases. Prog. Neurobiol. 2005, 76, 22–76. [Google Scholar] [CrossRef] [PubMed]
- Jeffrey, P.; Summerfield, S. Assessment of the blood–brain barrier in CNS drug discovery. Neurobiol. Dis. 2010, 37, 33–37. [Google Scholar] [CrossRef] [PubMed]
- Taylor, J.P.; Hardy, J.; Fischbeck, K.H. Toxic proteins in neurodegenerative disease. Science 2002, 296, 1991–1996. [Google Scholar] [CrossRef] [PubMed]
- Bandmann, O.; Burton, E.A. Genetic zebrafish models of neurodegenerative diseases. Neurobiol. Dis. 2010, 40, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Babin, P.J.; Goizet, C.; Raldúa, D. Zebrafish models of human motor neuron diseases: Advantages and limitations. Prog. Neurobiol. 2014, 118, 36–58. [Google Scholar] [CrossRef]
- Lemay, N.; Hayward, L.J.; Bosco, D.A.; Brown, R.H.; Zhou, H.; Burke, C.; Kwiatkowski, T.J.; Sapp, P., Jr.; Yasek, D.M.; Brown, R.H.; Hayward, L.J., Jr. Mutant FUS proteins that cause amyotrophic lateral sclerosis incorporate into stress granules. Hum. Mol. Genet. 2010, 19, 4160–4175. [Google Scholar]
- Lemmens, R.; Van Hoecke, A.; Hersmus, N.; Geelen, V.; D’Hollander, I.; Thijs, V.; Van Den Bosch, L.; Carmeliet, P.; Robberecht, W. Overexpression of mutant superoxide dismutase 1 causes a motor axonopathy in the zebrafish. Hum. Mol. Genet. 2007, 16, 2359–2365. [Google Scholar] [CrossRef]
- Laird, A.S.; Mackovski, N.; Rinkwitz, S.; Becker, T.S.; Giacomotto, J. Tissue-specific models of spinal muscular atrophy confirm a critical role of SMN in motor neurons from embryonic to adult stages. Hum. Mol. Genet. 2016, 25, 1728–1738. [Google Scholar] [CrossRef]
- Paquet, D.; Bhat, R.; Sydow, A.; Mandelkow, E.-M.; Berg, S.; Hellberg, S.; Fälting, J.; Schmid, B.; Haass, C. A zebrafish model of tauopathy allows in vivo imaging of neuronal cell death and drug evaluation Find the latest version: Technical advance A zebrafish model of tauopathy allows in vivo imaging of neuronal cell death and drug evaluation. J. Clin. Investig. 2009, 119, 1382–1395. [Google Scholar] [CrossRef]
- Miller, V.M.; Nelson, R.F.; Gouvion, C.M.; Williams, A.; Rodriguez-Lebron, E.; Harper, S.Q.; Davidson, B.L.; Rebagliati, M.R.; Paulson, H.L. CHIP Suppresses Polyglutamine Aggregation and Toxicity In Vitro and In Vivo. J. Neurosci. 2005, 25, 9152–9161. [Google Scholar] [CrossRef]
- McGown, A.; McDearmid, J.R.; Panagiotaki, N.; Tong, H.; Al Mashhadi, S.; Redhead, N.; Lyon, A.N.; Beattie, C.E.; Shaw, P.J.; Ramesh, T.M. Early interneuron dysfunction in ALS: Insights from a mutant sod1 zebrafish model. Ann. Neurol. 2013, 73, 246–258. [Google Scholar] [CrossRef] [PubMed]
- Kabashi, E.; Lin, L.; Tradewell, M.L.; Dion, P.A.; Bercier, V.; Bourgouin, P.; Rochefort, D.; Hadj, S.B.; Durham, H.D.; Rouleau, C.V.V.G.A.; et al. Gain and loss of function of ALS-related mutations of TARDBP (TDP-43) cause motor deficits in vivo. Hum. Mol. Genet. 2009, 19, 671–683. [Google Scholar] [CrossRef] [PubMed]
- Bai, Q.; Burton, E.A. Zebrafish models of Tauopathy HHS Public Access. Biochim. Biophys. Acta 2011, 1812, 353–363. [Google Scholar] [CrossRef]
- Hartl, F.U.; Kretzschmar, H.; Haass, C.; Hirschberger, T.; Giese, A.; Schmid, B.; Schiffer, N.W.; Broadley, S.A.; Tavan, P. Identification of Anti-prion Compounds as Efficient Inhibitors of Polyglutamine Protein Aggregation in a Zebrafish Model. J. Biol. Chem. 2006, 282, 9195–9203. [Google Scholar]
- Charles, B.K. Patterning the Brain of the Zebrafish Embryo. Annu. Rev. Neurosci. 2002, 16, 707–732. [Google Scholar] [CrossRef] [PubMed]
- Wullimann, M.F.; Rupp, B.; Reichert, H.; Verlag, B. Neuroanatomy of the Zebrafish Brain: A Topological Atlas; Birkhäuser: Basel, Switzerland, 1996. [Google Scholar]
- Brown, J.; Guerra-Assunção, J.A.; Saunders, D.; Willey, D.; Barker, D.; Ellwood, M.; Gordon, D.; Chow, W.; Clark, R.; Karotki, L.; et al. The zebrafish reference genome sequence and its relationship to the human genome. Nature 2013, 496, 498–503. [Google Scholar]
- Kumar, S.; Hedges, B.S. Amolecular timescalefor vertebrateevolution Sudhir. Nature 1998, 392, 917–920. [Google Scholar] [CrossRef]
- Clarke, A.R. Transgenesis Techniques: Principles and Protocols; Springer: Berlin, Germany, 2003; p. 561. [Google Scholar]
- Hruscha, A.; Krawitz, P.; Rechenberg, A.; Heinrich, V.; Hecht, J.; Haass, C.; Schmid, B. Efficient CRISPR/Cas9 genome editing with low off-target effects in zebrafish. Development 2013, 140, 4982–4987. [Google Scholar] [CrossRef]
- Musa, A.; Lehrach, H.; Russo, V.E. Distinct expression patterns of two zebrafish homologues of the human APP gene during embryonic development. Dev. Genes Evol. 2001, 211, 563–567. [Google Scholar] [CrossRef]
- Leimer, U.; Lun, K.; Romig, H.; Walter, J.; Grünberg, J.; Brand, M.; Haass, C. Zebrafish (Danio rerio) Presenilin Promotes Aberrant Amyloid β-Peptide Production and Requires a Critical Aspartate Residue for Its Function in Amyloidogenesis. Biochemistry 1999, 38, 13602–13609. [Google Scholar] [CrossRef]
- Groth, C.; Nornes, S.; McCarty, R.; Tamme, R.; Lardelli, M. Identification of a second presenilin gene in zebrafish with similarity to the human Alzheimer’s disease gene presenilin2. Dev. Genes Evol. 2002, 212, 486–490. [Google Scholar] [CrossRef]
- Moussavi Nik, S.H.; Wilson, L.; Newman, M.; Croft, K.; Mori, T.A.; Musgrave, I.; Lardelli, M. The BACE1-PSEN-AβPP regulatory axis has an ancient role in response to low oxygen/oxidative stress. J. Alzheimer’s Dis. 2012, 28, 515–530. [Google Scholar] [CrossRef] [PubMed]
- Van Bebber, F.; Hruscha, A.; Willem, M.; Schmid, B.; Haass, C. Loss of Bace2 in zebrafish affects melanocyte migration and is distinct from Bace1 knock out phenotypes. J. Neurochem. 2013, 127, 471–481. [Google Scholar] [CrossRef] [PubMed]
- Francis, R.; McGrath, G.; Zhang, J.; Ruddy, D.A.; Sym, M.; Apfeld, J.; Nicoll, M.; Maxwell, M.; Hai, B.; Ellis, M.C.; et al. aph-1 and pen-2 are required for Notch pathway signaling, γ-secretase cleavage of βAPP, and presenilin protein accumulation. Dev. Cell 2002, 3, 85–97. [Google Scholar] [CrossRef]
- Campbell, W.A.; Yang, H.; Zetterberg, H.; Baulac, S.; Sears, J.A.; Liu, T.; Wong, S.T.C.; Zhong, T.P.; Xia, W. Zebrafish lacking Alzheimer presenilin enhancer 2 (Pen-2) demonstrate excessive p53-dependent apoptosis and neuronal loss. J. Neurochem. 2006, 96, 1423–1440. [Google Scholar] [CrossRef]
- Team MGC (MGC), P. Generation and initial analysis of more than 15,000 full-length human and mouse cDNA sequences. Proc. Natl. Acad. Sci. USA 2002, 99, 16899–16903. [Google Scholar]
- Babin, P.J.; Thisse, C.; Durliat, M.; Andre, M.; Akimenko, M.-A.; Thisse, B. Both apolipoprotein E and A-I genes are present in a nonmammalian vertebrate and are highly expressed during embryonic development. Proc. Natl. Acad. Sci. USA 1997, 94, 8622–8627. [Google Scholar] [CrossRef]
- Woods, I.G.; Wilson, C.; Friedlander, B.; Chang, P.; Reyes, D.K.; Nix, R.; Kelly, P.D.; Chu, F.; Postlethwait, J.H.; Talbot, W.S.; et al. The zebrafish gene map defines ancestral vertebrate chromosomes. Genome Res. 2005, 15, 1307–1314. [Google Scholar] [CrossRef]
- Lee, J.; Peterson, S.M.; Freeman, J.L. Sex-specific characterization and evaluation of the Alzheimer’s disease genetic risk factor sorl1 in zebrafish during aging and in the adult brain following a 100 ppb embryonic lead exposure. J. Appl. Toxicol. 2017, 37, 400–407. [Google Scholar] [CrossRef]
- Bai, Q.; Mullett, S.J.; Garver, J.A.; Hinkle, D.A.; Burton, E.A. Zebrafish DJ-1 is evolutionarily conserved and expressed in dopaminergic neurons. Brain Res. 2006, 1113, 33–44. [Google Scholar] [CrossRef]
- Flinn, L.; Mortiboys, H.; Volkmann, K.; Kster, R.W.; Ingham, P.W.; Bandmann, O. Complex i deficiency and dopaminergic neuronal cell loss in parkin-deficient zebrafish (Danio rerio). Brain 2009, 132, 1613–1623. [Google Scholar] [CrossRef] [PubMed]
- Anichtchik, O.; Roach, A.; Goldsmith, P.; Diekmann, H.; Rubinsztein, D.C.; Fleming, A. Loss of PINK1 Function Affects Development and Results in Neurodegeneration in Zebrafish. J. Neurosci. 2008, 28, 8199–8207. [Google Scholar] [CrossRef]
- Ren, G.; Xin, S.; Li, S.; Zhong, H.; Lin, S. Disruption of lrrk2 does not cause specific loss of dopaminergic neurons in zebrafish. PLoS ONE 2011, 6, 2–7. [Google Scholar] [CrossRef] [PubMed]
- Karlovich, C.A.; John, R.M.; Ramirez, L.; Stainier, D.Y.; Myers, R.M. Characterization of the Huntington’s disease (HD) gene homolog in the zebrafish Danio rerio. Gene 1998, 217, 117–125. [Google Scholar] [CrossRef]
- Laboissonniere, L.A.; Smith, C.L.; Mesenbrink, J.; Chowdhury, R.; Burney, A.; Lang, M.; Sierra, M.; Stark, A.; Maldonado-Casalduc, G.; Muller, M.; et al. ALS-associated genes display CNS expression in the developing zebrafish. Gene Expr. Patterns 2018, 30, 14–31. [Google Scholar] [CrossRef]
- Schmid, B.; Hruscha, A.; Hogl, S.; Banzhaf-Strathmann, J.; Strecker, K.; van der Zee, J.; Teucke, M.; Eimer, S.; Hegermann, J.; Kittelmann, M.; et al. Loss of ALS-associated TDP-43 in zebrafish causes muscle degeneration, vascular dysfunction, and reduced motor neuron axon outgrowth. Proc. Natl. Acad. Sci. USA 2013, 110, 4986–4991. [Google Scholar] [CrossRef]
- Carlson, K.M.; Melcher, L.; Lai, S.; Zoghbi, H.Y.; Clark, H.B.; Orr, H.T. Characterization of the zebrafish atxn1/axh gene family. J. Neurogenet. 2009, 23, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Wood, J.D.; Bonath, F.; Kumar, S.; Ross, C.A.; Cunliffe, V.T. Disrupted-in-schizophrenia 1 and neuregulin 1 are required for the specification of oligodendrocytes and neurones in the zebrafish brain. Hum. Mol. Genet. 2009, 18, 391–404. [Google Scholar] [CrossRef]
- Kabashi, E.; Brustein, E.; Champagne, N.; Drapeau, P. Zebrafish models for the functional genomics of neurogenetic disorders. Biochim. Biophys. Acta 2011, 1812, 335–3345. [Google Scholar] [CrossRef]
- The State of the Art of Dementia Research: New Frontiers; World Alzheimer Report; Alzheimer’s Disease International: London, UK, 2018; pp. 1–48.
- Voisin, T.; Vellas, B. Diagnosis and Treatment of Patients with Severe Alzheimer’s Disease. Drugs Aging 2009, 26, 135–144. [Google Scholar] [CrossRef]
- Regeur, L.; Badsberg Jensen, G.; Pakkenberg, H.; Evans, S.M.; Pakkenberg, B. No global neocortical nerve cell loss in brains from patients with senile dementia of Alzheimer’s type. Neurobiol. Aging 1994, 15, 347–352. [Google Scholar] [CrossRef]
- Mark, J.; Paul, D.; Dorothy, G.; Juan, C. Differences in the pattern of hippocampal neuronal loss in normal ageing and Alzheimer â€TM s disease. Lancet 1994, 344, 769–772. [Google Scholar]
- Blennow, K.; De Leon, M.J.; Zetterberg, H. SeminarAlzheimer’s disease. Lancet 2006, 368, 1–17. [Google Scholar] [CrossRef]
- Jacobsen, L.; Madsen, P.; Jacobsen, C.; Nielsen, M.S.; Gliemann, J.; Petersen, C.M. Activation and Functional Characterization of the Mosaic Receptor SorLA/LR11. J. Biol. Chem. 2001, 276, 22788–22796. [Google Scholar] [CrossRef]
- Chen, M.; Martins, R.N.; Lardelli, M. Complex splicing and neural expression of duplicated tau genes in zebrafish embryos. J. Alzheimer’s Dis. 2009, 18, 305–317. [Google Scholar] [CrossRef]
- Abeliovich, A.; Flint Beal, M. Parkinsonism genes: Culprits and clues. J. Neurochem. 2006, 99, 1062–1072. [Google Scholar] [CrossRef]
- Galvin, J.E. Cognitive change in Parkinson disease. Alzheimer Dis. Assoc. Disord. 2006, 20, 302–310. [Google Scholar] [CrossRef] [PubMed]
- Huntington, T.; Macdonald, M.E.; Ambrose, C.M.; Duyao, M.P.; Myers, R.H.; Lin, C.; Srinidhi, L.; Barnes, G.; Taylor, S.A.; James, M.; et al. A Novel Gene Containing a Trinucleotide Repeat That Is Expanded and Unstable on Huntington’s Disease Chromosomes. Cell 1993, 72, 971–983. [Google Scholar]
- Walker, F.O. Huntington’s disease. Lancet 2007, 369, 218–228. [Google Scholar] [CrossRef]
- Lumsden, A.L.; Henshall, T.L.; Dayan, S.; Lardelli, M.T.; Richards, R.I. Huntingtin-deficient zebrafish exhibit defects in iron utilization and development. Hum. Mol. Genet. 2007, 16, 1905–1920. [Google Scholar] [CrossRef]
- Diekmann, H.; Fleming, A.; Rubinsztein, D.C.; Roach, A.; Anichtchik, O.; Goldsmith, P.; Futter, M. Decreased BDNF Levels Are a Major Contributor to the Embryonic Phenotype of Huntingtin Knockdown Zebrafish. J. Neurosci. 2009, 29, 1343–1349. [Google Scholar] [CrossRef]
- Deng, H.X.; Hentati, A.; Tainer, J.A.; Iqbal, Z.; Cayabyab, A.; Hung, W.Y.; Getzoff, E.D.; Hu, P.; Herzfeldt, B.; Roos, R.P. Amyotrophic lateral sclerosis and structural defects in Cu,Zn superoxide dismutase. Science 1993, 261, 1047–1051. [Google Scholar] [CrossRef]
- Rosen, D.R.; Siddiquet, T.; Pattersont, D.; Figlewicz, D.A.; Ii, P.S.; Hentatit, A.; Donaldsont, D.; Goto, J.; Ii, J.P.O.R.; Dengt, H.; et al. Mutations in Cu/Zn superoxide dismutase gene are associated with familial amyotrophic lateral sclerosis. Nature 1993, 362, 59–62. [Google Scholar] [CrossRef] [PubMed]
- Turner, B.J.; Talbot, K. Transgenics, toxicity and therapeutics in rodent models of mutant SOD1-mediated familial ALS. Prog. Neurobiol. 2008, 85, 94–134. [Google Scholar] [CrossRef]
- Weinberger, D. From neuropathology to neurodevelopment. Lancet 1995, 346, 552–557. [Google Scholar] [CrossRef]
- Nuechterlein, K.H.; Barch, M.; Gold, J.M.; Goldberg, T.E.; Green, M.F.; Heaton, R.K. Identification of separable cognitive factors in schizophrenia. Schizophr. Res. 2004, 72, 29–39. [Google Scholar] [CrossRef]
- Swerdlow, N.R.; Butters, N.; Geyer, M.A.; Paulsen, J.; Swenson, M.R.; Braff, D.L. Impaired prepulse inhibition of acoustic and tactile startle response in patients with Huntington’s disease. J. Neurol. Neurosurg. Psychiatry 2008, 58, 192–200. [Google Scholar] [CrossRef] [PubMed]
- Braff, D.L.; Geyer, M.A.; Swerdlow, N.R. Human studies of prepulse inhibition of startle: Normal subjects, patient groups, and pharmacological studies. Psychopharmacology 2001, 156, 234–258. [Google Scholar] [CrossRef]
- Braff, D.; Schork, N.J. Gottesman, I.I. Endophenotyping schizophrenia. Am. J. Psychiatry 2007, 164, 705–707. [Google Scholar] [CrossRef] [PubMed]
- Burgess, H.A.; Granato, M. Sensorimotor Gating in Larval Zebrafish. J. Neurosci. 2007, 27, 4984–4994. [Google Scholar] [CrossRef]
- Sullivan, P.F.; Kendler, K.S.; Neale, M.C. Schizophrenia as a Complex Trait. Arch. Gen. Psychiatry 2003, 60, 1187–1192. [Google Scholar] [CrossRef] [PubMed]
- Van Rensburg, E.J.; Xu, B.; Karayiorgou, M.; Levy, S.; Roos, J.L.; Gogos, J.A. Strong association of de novo copy number mutations with sporadic schizophrenia. Nat. Genet. 2008, 40, 880–885. [Google Scholar]
- Tarabeux, J.; Champagne, N.; Brustein, E.; Hamdan, F.F.; Gauthier, J.; Lapointe, M.; Maios, C.; Piton, A.; Spiegelman, D.; Henrion, É.M.; Millet, B.; et al. De Novo Truncating Mutation in Kinesin 17 Associated with Schizophrenia. Biol. Psychiatry 2010, 68, 649–656. [Google Scholar] [CrossRef]
- Girard, S.L.; Dion, P.A.; Rouleau, G.A. Schizophrenia genetics: Putting all the pieces together. Curr. Neurol. Neurosci. Rep. 2012, 12, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Morris, J.A. Zebrafish: A model system to examine the neurodevelopmental basis of schizophrenia. Prog. Brain Res. 2009, 179, 97–106. [Google Scholar] [PubMed]
- Burgess, H.A.; Granato, M. Modulation of locomotor activity in larval zebrafish during light adaptation. J. Exp. Biol. 2007, 210, 2526–2539. [Google Scholar] [CrossRef] [PubMed]
- Millar, J.K.; Christie, S.; Semple, C.A.M.; Porteous, D.J. Chromosomal Location and Genomic Structure of the Human Translin-Associated Factor X Gene (TRAX; TSNAX) Revealed by Intergenic Splicing to DISC1, a Gene Disrupted by a Translocation Segregating with Schizophrenia. Genomics 2000, 67, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Zashikhina, A. Juvenile Chronic Physical Illness in Northern Russia: Studies on Mental Health, Health-Related Quality of Life, and Family Functioning. Ph.D. Thesis, Umeå Universitet, Umeå, Sweden, 2014. [Google Scholar]
- Galanopoulou, A.S.; Buckmaster, P.S.; Staley, K.J.; Moshe, S.L.; Perucca, E.; Engel, J.; Löscher, W.; Noebels, J.L.; Pitkänen, A.; Stables, J.; White, H.S.; et al. Identification of new epilepsy treatments: Issues in preclinical methodology. Epilepsia 2012, 53, 571–582. [Google Scholar] [CrossRef]
- Desmond, D.; Kyzar, E.; Gaikwad, S.; Green, J.; Riehl, R.; Roth, A.; Stewart, A.M.; Kalueffet, A.V. Assessing epilepsy-related behavioral phenotypes in adult zebrafish. In Zebrafish Protocols for Neurobehavioral Research; Springer: Berlin, Germany, 2012; pp. 313–322. [Google Scholar]
- Crawford, A.D.; Liekens, S.; Kamuhabwa, A.R.; Maes, J.; Munck, S.; Busson, R.; Rozenski, J.; Esguerra, C.V.; de Witte, P.A.M. Zebrafish bioassay-guided natural product discovery: Isolation of angiogenesis inhibitors from East African medicinal plants. PLoS ONE 2011, 6, e14694. [Google Scholar] [CrossRef] [PubMed]
- Marcourt, L.; Bock, M.; Maljevic, S.; Dayrit, F.M.; Challal, S.; Queiroz, E.F.; de Witte, P.A.M.; Crawford, A.D.; Harvey, A.L.; Buenafe, O.E.M.; et al. Zebrafish Bioassay-Guided Microfractionation Identifies Anticonvulsant Steroid Glycosides from the Philippine Medicinal Plant Solanum torvum. ACS Chem. Neurosci. 2014, 5, 993–1004. [Google Scholar]
- Pitchai, A.; Nagarajan, N.; Vincent, S.G.P.; Rajaretinam, R.K. Zebrafish bio-assay guided isolation of human acetylcholinesterase inhibitory trans-tephrostachin from Tephrosia purpurea (L.) Pers. Neurosci. Lett. 2018, 687, 268–275. [Google Scholar] [CrossRef] [PubMed]
- Buenafe, O.E.; Orellana-Paucar, A.; Maes, J.; Huang, H.; Ying, X.; De Borggraeve, W.; Crawford, A.D.; Luyten, W.; Esguerra, C.V.; De Witte, P. Tanshinone IIA exhibits anticonvulsant activity in zebrafish and mouse seizure models. ACS Chem. Neurosci. 2013, 4, 1479–1487. [Google Scholar] [CrossRef] [PubMed]
- Esguerra, C.V.; Kamuhabwa, A.R.; Wolfender, J.-L.; Moshi, M.J.; Siverio-Mota, D.; Marcourt, L.; Maes, J.; Bohni, N.; de Witte, P.A.M.; Cordero-Maldonado, M.L.; et al. Integration of Microfractionation, qNMR and Zebrafish Screening for the In Vivo Bioassay-Guided Isolation and Quantitative Bioactivity Analysis of Natural Products. PLoS ONE 2013, 8, e64006. [Google Scholar]
- Jones, R.W.; Huffman, M.N. Fish Embryos as Bio-Assay Material in Testing Chemicals for Effects on Cell Division and Differentiation. Trans. Am. Microsc. Soc. 1957, 76, 177–183. [Google Scholar] [CrossRef]
- Liu, M.; Copmans, D.; Lu, J.G.; Yang, M.R.; Sourbron, J.; Ny, A.; Jiang, Z.H.; de Witte, P.A.M.; Luyten, W. Bioassay-guided isolation of anti-seizure principles from Semen Pharbitidis using a zebrafish pentylenetetrazol seizure model. J. Ethnopharmacol. 2019, 232, 130–134. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.L.; Cheng, L.; Kwok, H.F.; Ko, C.H.; Lau, T.W.; Koon, C.M.; Zhao, M.; Lau, C.P.; Lau, K.M.; Wong, C.W.; et al. Bioassay-guided isolation of norviburtinal from the root of Rehmannia glutinosa, exhibited angiogenesis effect in zebrafish embryo model. J. Ethnopharmacol. 2011, 137, 1323–1327. [Google Scholar] [CrossRef]
- Liang, F.; Han, Y.; Gao, H.; Xin, S.; Chen, S.; Wang, N.; Qin, W.; Zhong, H.; Lin, S.; Yao, X.; et al. Kaempferol Identified by Zebrafish Assay and Fine Fractionations Strategy from Dysosma versipellis Inhibits Angiogenesis through VEGF and FGF Pathways. Sci. Rep. 2015, 5, 1–10. [Google Scholar] [CrossRef]
- Cheng, M.-C.; Lee, T.-H.; Chu, Y.-T.; Syu, L.-L.; Hsu, S.-J.; Cheng, C.-H.; Lee, J.; Wu, C.-K. Melanogenesis Inhibitors from the Rhizoma of Ligusticum Sinense in B16-F10 Melanoma Cells In Vitro and Zebrafish In Vivo. Int. J. Mol. Sci. 2018, 19, 3994. [Google Scholar] [CrossRef]
- Brillatz, T.; Lauritano, C.; Jacmin, M.; Khamma, S.; Marcourt, L.; Righi, D.; Romano, G.; Esposito, F.; Ianora, A.; Queiroz, E.F.; et al. Zebrafish-based identification of the antiseizure nucleoside inosine from the marine diatom Skeletonema marinoi. PLoS ONE 2018, 13, e0196195. [Google Scholar] [CrossRef]
- Thermes, V.; Grabher, C.; Ristoratore, F.; Bourrat, F.; Choulika, A.; Wittbrodt, J.; Joly, J.-S.; Thermes, V.; Grabher, C.; Ristoratore, F.; et al. I-SceI meganuclease mediates highly efficient transgenesis in fish. Mech. Dev. 2002, 118, 91–98. [Google Scholar] [CrossRef]
- Kawakami, K.; Shima, A.; Kawakami, N. Identification of a functional transposase of the Tol2 element, an Ac-like element from the Japanese medaka fish, and its transposition in the zebrafish germ lineage. Proc. Natl. Acad. Sci. USA 2002, 97, 11403–11408. [Google Scholar] [CrossRef]
- Hans, S.; Kaslin, J.; Freudenreich, D.; Brand, M. Temporally-controlled site-specific recombination in zebrafish. PLoS ONE 2009, 4, e4640. [Google Scholar] [CrossRef]
- Halpern, M.E.; Rhee, J.; Goll, M.G.; Akitake, C.M.; Parsons, M.; Leach, S.D. Gal4/UAS Transgenic Tools and Their Application to Zebrafish. Zebrafish 2008, 5, 97–110. [Google Scholar] [CrossRef]
- Schmid, B.; Haass, C. Genomic editing opens new avenues for zebrafish as a model for neurodegeneration. J. Neurochem. 2013, 127, 461–470. [Google Scholar] [CrossRef]
- Hwang, W.Y.; Fu, Y.; Reyon, D.; Maeder, ML.; Tsai, S.Q.; Sander, J.D.; Joung, J.K.; Peterson, R.T.; Yeh, J.R.J. Efficient genome editing in zebrafish using a CRISPR-Cas system. Nat. Biotechnol. 2013, 31, 227–229. [Google Scholar] [CrossRef]
- Burns, C.G.; Milan, D.J.; Grande, E.J.; Rottbauer, W.; MacRae, C.A.; Fishman, M.C. High-throughput assay for small molecules that modulate zebrafish embryonic heart rate. Nat. Chem. Biol. 2005, 1, 263–264. [Google Scholar] [CrossRef] [PubMed]
- Goldsmith, P. Zebrafish as a pharmacological tool: The how, why and when. Curr. Opin. Pharmacol. 2004, 4, 504–512. [Google Scholar] [CrossRef]
- Sun, Z.; Chen, W.; Farrington, S.; Haldi, M.; Hopkins, N.; Amsterdam, A.; Golling, G.; Townsend, K.; Burgess, S. A large-scale insertional mutagenesis screen in zebrafish. Genes Dev. 2002, 13, 2713–2724. [Google Scholar]
- Guo, S.; Wilson, S.W.; Cooke, S.; Chitnis, A.B.; Driever, W.; Rosenthal, A. Mutations in the Zebrafish Unmask Shared Regulatory Pathways Controlling the Development of Catecholaminergic Neurons. Dev. Biol. 1999, 208, 473–487. [Google Scholar] [CrossRef]
- Amatruda, J.F.; Tallarico, J.A.; Weber, G.; Zon, L.I.; Pfaff, K.L.; King, R.W.; Straub, C.T.; Shepard, J.L.; Murphey, R.D.; Stern, H.M. Small molecules that delay S phase suppress a zebrafish bmyb mutant. Nat. Chem. Biol. 2005, 1, 366–370. [Google Scholar]
- MacRae, C.A.; Milan, D.J.; Fishman, M.C.; Schreiber, S.L.; Peterson, R.T.; Peterson, T.A.; Shaw, S.Y.; Zhong, T.P. Chemical suppression of a genetic mutation in a zebrafish model of aortic coarctation. Nat. Biotechnol. 2004, 22, 595–599. [Google Scholar]
- Song, P.; Pimplikar, S.W. Knockdown of amyloid precursor protein in zebrafish causes defects in motor axon outgrowth. PLoS ONE 2012, 7, e34209. [Google Scholar] [CrossRef] [PubMed]
- Prabhudesai, S.; Bensabeur, F.Z.; Abdullah, R.; Basak, I.; Baez, S.; Alves, G.; Holtzman, N.G.; Larsen, J.P.; Møller, S.G. LRRK2 knockdown in zebrafish causes developmental defects, neuronal loss, and synuclein aggregation. J. Neurosci. Res. 2016, 94, 717–735. [Google Scholar] [CrossRef]
- Wang, W.Y.; Fu, Y.; Reyon, D.; Maeder, M.L.; Kaini, P.; Sander, J.D.; Joung, J.K.; Peterson, R.T.; Yeh, J.R.J. Heritable and Precise Zebrafish Genome Editing Using a CRISPR-Cas System. PLoS ONE 2013, 8, e68708. [Google Scholar]
- Armstrong, G.A.B.; Liao, M.; You, Z.; Lissouba, A.; Chen, B.E.; Drapeau, P. Homology directed knockin of point mutations in the zebrafish tardbp and fus genes in ALS using the CRISPR/Cas9 system. PLoS ONE 2016, 11, e0150188. [Google Scholar] [CrossRef] [PubMed]
- Nery, L.R.; Silva, N.E.; Fonseca, R.; Vianna, M.R.M. Presenilin-1 Targeted Morpholino Induces Cognitive Deficits, Increased Brain Aβ1−42 and Decreased Synaptic Marker PSD-95 in Zebrafish Larvae. Neurochem. Res. 2017, 42, 2959–2967. [Google Scholar] [CrossRef] [PubMed]
- Peri, F.; Nüsslein-Volhard, C. Live Imaging of Neuronal Degradation by Microglia Reveals a Role for v0-ATPase a1 in Phagosomal Fusion In Vivo. Cell 2008, 133, 916–927. [Google Scholar] [CrossRef]
- Bretaud, S.; Allen, C.; Ingham, P.W.; Bandmann, O. p53-dependent neuronal cell death in a DJ-1-deficient zebrafish model of Parkinson’s disease. J. Neurochem. 2007, 100, 1626–1635. [Google Scholar] [CrossRef]
- Fett, M.E.; Pilsl, A.; Paquet, D.; van Bebber, F.; Haass, C.; Tatzelt, J.; Schmid, B.; Winklhofer, K.F. Parkin is protective against proteotoxic stress in a transgenic zebrafish model. PLoS ONE 2010, 5, e11783. [Google Scholar] [CrossRef]
- Zhang, Y.; Nguyen, D.T.; Olzomer, E.M.; Poon, G.P.; Cole, N.J.; Puvanendran, A.; Phillips, B.R.; Hesselson, D. Rescue of Pink1 Deficiency by Stress-Dependent Activation of Autophagy. Cell Chem. Biol. 2017, 24, 471–480. [Google Scholar] [CrossRef]
- Tanaka, E.M.; Ferretti, P. Considering the evolution of regeneration in the central nervous system. Nat. Rev. Neurosci. 2009, 10, 713–723. [Google Scholar] [CrossRef]
- Chapouton, P.; Jagasia, R.; Bally-Cuif, L. Adult neurogenesis in non-mammalian vertebrates. BioEssays 2007, 29, 745–757. [Google Scholar] [CrossRef]
- Kizil, C.; Kaslin, J.; Kroehne, V.; Brand, M. Adult neurogenesis and brain regeneration in zebrafish. Dev. Neurobiol. 2012, 72, 429–461. [Google Scholar] [CrossRef] [PubMed]
- Cosacak, M.I.; Papadimitriou, C.; Kizil, C. Regeneration, Plasticity, and Induced Molecular Programs in Adult Zebrafish Brain. Biomed. Res. Int. 2015, 2015, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Basu, A. Inflammation: A new candidate in modulating adult neurogenesis. J. Neurosci. Res. 2008, 86, 1199–1208. [Google Scholar] [CrossRef] [PubMed]
- Kizil, C.; Kyritsis, N.; Brand, M. Effects of inflammation on stem cells: Together they strive? EMBO Rep. 2015, 16, 416–426. [Google Scholar] [CrossRef] [PubMed]
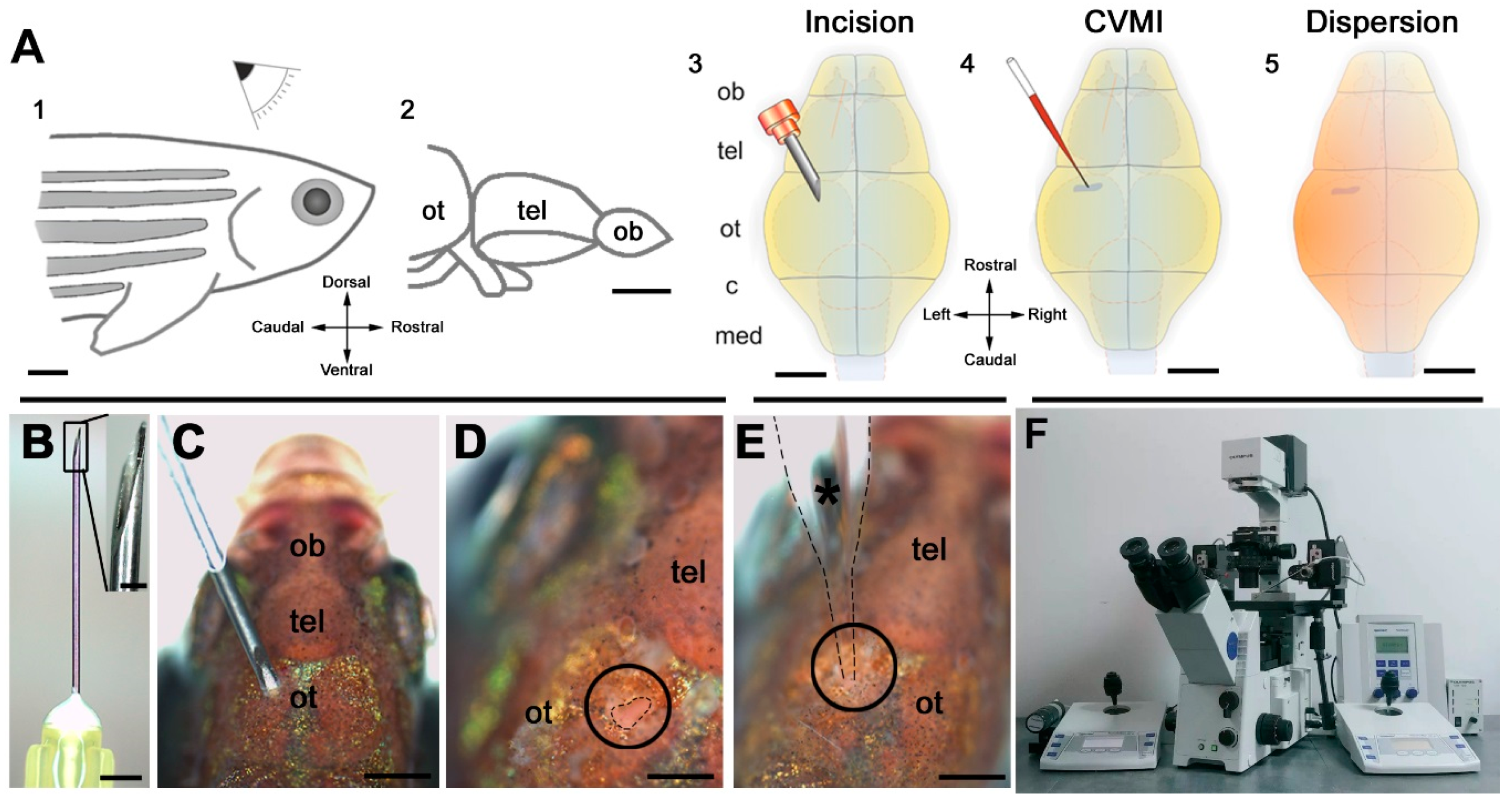
- Kizil, C.; Brand, M. Cerebroventricular microinjection (CVMI) into adult zebrafish brain is an efficient misexpression method for forebrain ventricular cells. PLoS ONE 2011, 6, e27395. [Google Scholar] [CrossRef] [PubMed]
- Bhattarai, P.; Thomas, A.K.; Cosacak, M.I.; Papadimitriou, C.; Mashkaryan, V.; Froc, C.; Reinhardt, S.; Kurth, T.; Dahl, T.; Zhang, Y.; et al. IL4/STAT6 Signaling Activates Neural Stem Cell Proliferation and Neurogenesis upon Amyloid-β42 Aggregation in Adult Zebrafish Brain. Cell Rep. 2016, 17, 941–948. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.W.; Quaranta, V.; Glenner, G.G. Neuritic plaques and cerebrovascular amyloid in Alzheimer disease are antigenically related. Proc. Natl. Acad. Sci. USA 1985, 82, 8729–8732. [Google Scholar] [CrossRef]
- Masters, C.L.; Simms, G.; Weinman, N.A.; Multhaup, G.; McDonald, B.L.; Beyreuther, K. Amyloid plaque core protein in Alzheimer disease and Down syndrome. Proc. Natl. Acad. Sci. USA 1985, 82, 4245–4249. [Google Scholar] [CrossRef]
- Younkin, S.G. The role of Aβ42 in Alzheimer’s disease. J. Physiol. Paris 1998, 92, 289–292. [Google Scholar] [CrossRef]
- Liu, W.; Guan, Y.; Collodi, P. A Zebrafish Cell Culture Assay for the Identification of MicroRNA Targets. Zebrafish 2010, 7, 343–348. [Google Scholar] [CrossRef] [PubMed]
- Myhre, J.L.; Pilgrim, D.B. Cellular Differentiation in Primary Cell Cultures from Single Zebrafish Embryos as a Model for the Study of Myogenesis. Zebrafish 2010, 7, 255–266. [Google Scholar] [CrossRef]
- Robles, V.; Martí, M.; Belmonte, J.C.I. Study of Pluripotency Markers in Zebrafish Embryos and Transient Embryonic Stem Cell Cultures. Zebrafish 2011, 8, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Sakowski, S.A.; Lunn, J.S.; Busta, A.S.; Palmer, M.; Dowling, J.J.; Feldman, E.L. A novel approach to study motor neurons from zebrafish embryos and larvae in culture. J. Neurosci. Methods 2012, 205, 277–282. [Google Scholar] [CrossRef] [PubMed]
- Ciarlo, C.A.; Zon, L. Embryonic cell culture in zebrafish. Methods Cell Biol. 2016, 133, 1–10. [Google Scholar]
- Sassen, W.A.; Lehne, F.; Russo, G.; Wargenau, S.; Dübel, S.; Köster, R.W. Embryonic zebrafish primary cell culture for transfection and live cellular and subcellular imaging. Dev. Biol. 2017, 430, 18–31. [Google Scholar] [CrossRef]
- Giros, B.; Fassier, C.; Nothias, F.; Lumsden, A.; Hazan, J.; Scholpp, S.; Hutt, J.A.; Houart, C.; Schneider-Maunoury, S. Zebrafish atlastin controls motility and spinal motor axon architecture via inhibition of the BMP pathway. Nat. Neurosci. 2010, 13, 1380–1387. [Google Scholar]
- Liu, D.; Westerfield, M. Clustering of muscle acetylcholine receptors requires motoneurons in live embryos, but not in cell culture. J. Neurosci. 2018, 12, 1859–1866. [Google Scholar] [CrossRef]
- Ghosh, C.; Liu, Y.; Ma, C.; Collodi, P. Cell cultures derived from early zebrafish embryos differentiate in vitro into neurons and astrocytes. Cytotechnology 1997, 23, 221–230. [Google Scholar] [CrossRef]
- Andersen, S.S.L. Preparation of dissociated Zebrafish spinal neuron cultures. Methods Cell Sci. 2001, 23, 205–209. [Google Scholar] [CrossRef] [PubMed]
- Beattie, C.E. Control of motor axon guidance in the zebrafish embryo. Brain Res. Bull. 2000, 53, 489–500. [Google Scholar] [CrossRef]
- Eisen, J.S. Motoneuronal development in the embryonic zebrafish. Development 1991, 2, 141–147. [Google Scholar]
- Hendricks, M.; Jesuthasan, S. Electroporation-based methods for in vivo, whole mount and primary culture analysis of zebrafish brain development. Neural Dev. 2007, 2, 1–10. [Google Scholar] [CrossRef]
- Saint-amant, L.; Sprague, S.M.; Hirata, H.; Li, Q.; Cui, W.W.; Zhou, W.; Poudou, O.; Hume, R.I.; Kuwada, J.Y. The Zebrafish ennui Behavioral Mutation Disrupts Acetylcholine Receptor Localization and Motor Axon Stability. Dev. Neurobiol. 2007, 68, 45–61. [Google Scholar] [CrossRef]
- Currie, P.D.; Ferguson, C.; Westerfield, M.; Key, B.; Nixon, S.J.; Méry, P.-F.; Parton, R.G.; Hancock, J.F.; Wegner, J. Zebrafish as a model for caveolin-associated muscle disease; caveolin-3 is required for myofibril organization and muscle cell patterning. Hum. Mol. Genet. 2005, 14, 1727–1743. [Google Scholar]
- Vreede, A.P.; Dowling, J.J.; Low, S.E.; Feldman, E.L.; Bonnemann, C.G.; Gibbs, E.M.; Kuwada, J.Y. Loss of Myotubularin Function Results in T-Tubule Disorganization in Zebrafish and Human Myotubular Myopathy. PLoS Genet. 2009, 5, e1000372. [Google Scholar]
- Don, E.K.; Watchon, M.; Yuan, K.C.; Fifita, J.A.; Blair, I.P.; Nicholson, G.A.; Laird, A.S.; Acosta, J.R.; Goldsbury, C.; Svahn, A.J.; et al. Neuronal cell culture from transgenic zebrafish models of neurodegenerative disease. Biol. Open 2018, 7, bio036475. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Disease | Protein | Human Gene | Zebrafish Gene | Amino Acid Similarity (%) | Reference |
|---|---|---|---|---|---|
| Alzheimer’s Disease | Amyloid precursor protein | APP GeneID: 351 Locus: 21q21.2 Protein length: 695 | appa GeneID: 58083 Chromosome: 1 Protein length: 738 | 74 | [41] |
| appb GeneID: 170846 Chromosome: 9 Protein length: 694 | 77 | ||||
| Presenilin-1 | PSEN1 GeneID: 5663 Locus: 14q24.3 Protein length: 467 | psen1 GeneID: 30221 Chromosome: 17 Protein length: 456 | 75 | [42] | |
| Presenilin-2 | PSEN2 GeneID: 5664 Locus: 1q31-q42 Protein length: 448 | psen2 GeneID: 58026 Chromosome: 1 Protein length: 441 | 76 | [43] | |
| β-secretase | BACE1 GeneID: 23621 Locus: 11q23.2-q23.3 Protein length: 501 | bace1 zgc:77409 GeneID: 403005 Chromosome: 15 Protein length: 505 | 82 | [44] | |
| BACE2 GeneID: 25825 Locus: 21q22.2-q22.3 Protein length: 518 | bace2 zgc:103530 GeneID: 449818 chromosome: 15 Protein length: 462 | [45] | |||
| γ-secretase | PSENEN GeneID: 55851 Locus: 19q13.12 Protein length: 101 | psenen GeneID: 402810 chromosome: 15 Protein length: 101 | 91 | [46,47] | |
| NCSTN Gene ID: 23385 Locus: 1q23.2 Protein length: 709 | ncstn GeneID: 494449 chromosome: 2 Protein length: 707 | 56 | [48] | ||
| APH1b Gene ID: 83464 Locus: 15q22.2 Protein length: 257 | aph1p Gene ID: 386808 chromosome: 7 Protein length: 258 | [46] | |||
| Apolipoprotein E (ApoE) | APOE GeneID:348 Locus: 19q13.32 Protein length: 317 | apoea Gene ID: 553587 chromosome: 19 Protein length: 269 | 27.5 | [49] | |
| apoeb Gene ID: 30314 chromosome: 16 Protein length: 281 | [50] | ||||
| Sortilin related receptor 1 (Sorl1) | SORL1 GeneID: 6653, Locus:11q24.1 Protein length: 2214 | sorl1 Gene ID: 497306, chromosome: 15 Protein length: 2213 | 64 | [51] | |
| Parkinson’s Disease | DJ-1 | DJ-1 Gene ID: 11315 Locus: PARK7 1p36.23 Protein length: 189 | dj-1 Gene ID: 449674 Chromosome: 11 Protein length: 189 | 83 | [52] |
| Parkin | PRKN Gene ID: 5071 Locus: PARK2 6q25.2-q27 Protein length: 465 | prkn Gene ID: 550328 Chromosome: 13 Protein length: 458 | 62 | [53] | |
| PTEN-induced kinase 1 (PINK-1) | PINK1 Gene ID: 65018 Locus: PARK6 1p36 Protein length: 581 | pink1 Gene ID: 494085 Chromosome: 6 Protein length: 574 | 54 | [54] | |
| Leucine-rich repeat kinase2 (LRRK2) | LRRK2 GeneID: 120892 Locus: PARK8 12q12 Protein length: 2527 | lrrk2 GeneID: 559366 Chromosome: 25 Protein length: 1985 | 38 | [55] | |
| Huntington’s Disease | Huntingtin | HTT GeneID: 3064 Locus: 4q16.3 Protein length: 3144 | htt GeneID: 30214 Chromosome: 1 Protein length: 3121 | 70 | [56] |
| Amyotrophic Lateral Sclerosis (ALS) | Fused in sarcoma | FUS GeneID: 2521 Locus: 16p11.2 Protein length: 526 | fus Gene ID: 394058 Chromosome: 3 Protein length: 541 | 63 | [57] |
| Tar DNA binding protein of 43 (TDP-43) | TARDBP GeneID: 23435 Locus: 1p36.22 Protein length: 414 | tardpb GeneID: 325052 Chromosome: 6 Protein length: 412 | 71 | [58] | |
| Spinocerebellar Ataxia Type 1 | Ataxin 1 | ATXN1 GeneID: 6310 Locus: 6p23 Protein length: 815 | atxn1a GeneID: 565841 Chromosome: 16 Protein length: 827 | 32 | [59] |
| atxn1b GeneID: 557340 Chromosome: 19 Protein length: 781 | 42 | ||||
| Schizophrenia | Dystrobrevin binding protein | DISC1 Gene ID: 27185 Locus: 1q42.2 Protein length:854 | disc1 GeneID: 407621 Chromosome: 13 Protein length: 994 | 53 | [60] |
| Kinesin family member 17 | KIF17 Gene ID: 57576 Locus: 1p36.12 Protein length:1029 | kif17 GeneID:557863 Chromosome: 11 Protein length: 823 | 83 | [61] |
| Source | Disease/Targets | Molecules | References |
|---|---|---|---|
| Pharbitis nil (Seeds) | anti-seizure | Pharbitin | [100] |
| Rehmannia glutinosa (Root) | angiogenesis effect | Norviburtinal | [101] |
| Rhynchosia viscosa (Whole Plant) | angiogenesis effect | Rhynchoviscin | [98] |
| Dysosma versipellis | anti-angiogenesis | Kaempferol | [102] |
| Ligusticum sinense (Rhizoma) | anti-melanogenesis | 1. Lignan 2. cis-4-pentylcyclohex-3-ene-1,2-diol | [103] |
| Tephrosia purpurea (Leaves) | anti-acetylcholinesterase | trans-Tephrostachin | [96] |
| Solanum torvum (leaves) | anti-convulsant | Paniculonin A Paniculonin B | [95] |
| Skeletonema marinoi | anti-seizure | Inosine | [104] |
| Disease | Gene | Technique | References |
|---|---|---|---|
| Alzheimer’s Disease | appb | Morpholino injection - knockdown | [117] |
| psen1 | Morpholino injections - knockdown | [121] | |
| bace2 | Zinc-finger nucleases (ZFNs)—knockout | [45] | |
| psenen | morpholino injection—knockdown | [47] | |
| apoea | CRISPR-Cas—knockout | [119] | |
| apoe | morpholino injection (live cell imaging) | [122] | |
| Parkinson’s Disease | djJ-1 | morpholino injection—knockdown | [123] |
| prkn | gripNAsTM-mediated knockdown | [124] | |
| pink1 | transcription activator-like effector nucleases (TALENs)—knockdown | [125] | |
| lrrk2 | morpholino injection—knockdown | [118] | |
| Amyotrophic Lateral Sclerosis | tardbp (bpt1) | CRISPR-Cas—knockin | [40] |
| fus | CRISPR-Cas—knockin | [120] | |
| Huntington’s Disease | htt | morpholino injection—knockdown | [71] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pitchai, A.; Rajaretinam, R.K.; Freeman, J.L. Zebrafish as an Emerging Model for Bioassay-Guided Natural Product Drug Discovery for Neurological Disorders. Medicines 2019, 6, 61. https://doi.org/10.3390/medicines6020061
Pitchai A, Rajaretinam RK, Freeman JL. Zebrafish as an Emerging Model for Bioassay-Guided Natural Product Drug Discovery for Neurological Disorders. Medicines. 2019; 6(2):61. https://doi.org/10.3390/medicines6020061
Chicago/Turabian StylePitchai, Arjun, Rajesh Kannan Rajaretinam, and Jennifer L. Freeman. 2019. "Zebrafish as an Emerging Model for Bioassay-Guided Natural Product Drug Discovery for Neurological Disorders" Medicines 6, no. 2: 61. https://doi.org/10.3390/medicines6020061
APA StylePitchai, A., Rajaretinam, R. K., & Freeman, J. L. (2019). Zebrafish as an Emerging Model for Bioassay-Guided Natural Product Drug Discovery for Neurological Disorders. Medicines, 6(2), 61. https://doi.org/10.3390/medicines6020061