
Dimethyl Sulfoxide: Morphological, Histological, and Molecular View on Developing Chicken Liver

 ,
,  ,
,
Abstract

1. Introduction
2. Materials and Methods
2.1. Histology
2.1.1. Light Microscopy
2.1.2. Transmission Electron Microscopy
2.2. Molecular Analysis—RNA Extraction and RT qPCR
2.3. Statistical Analysis
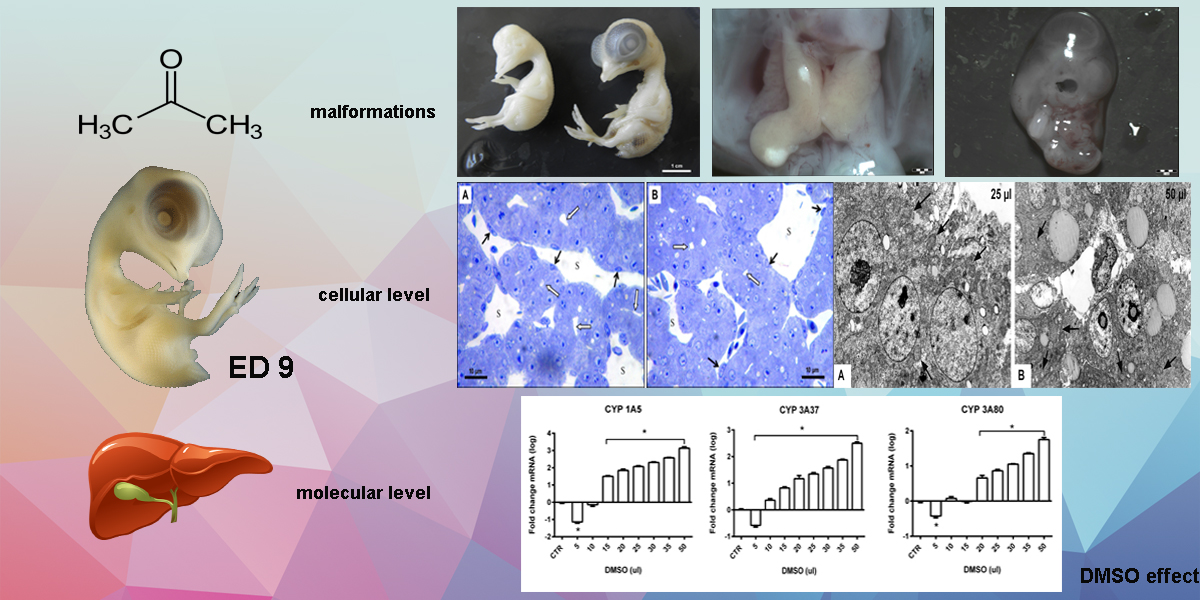
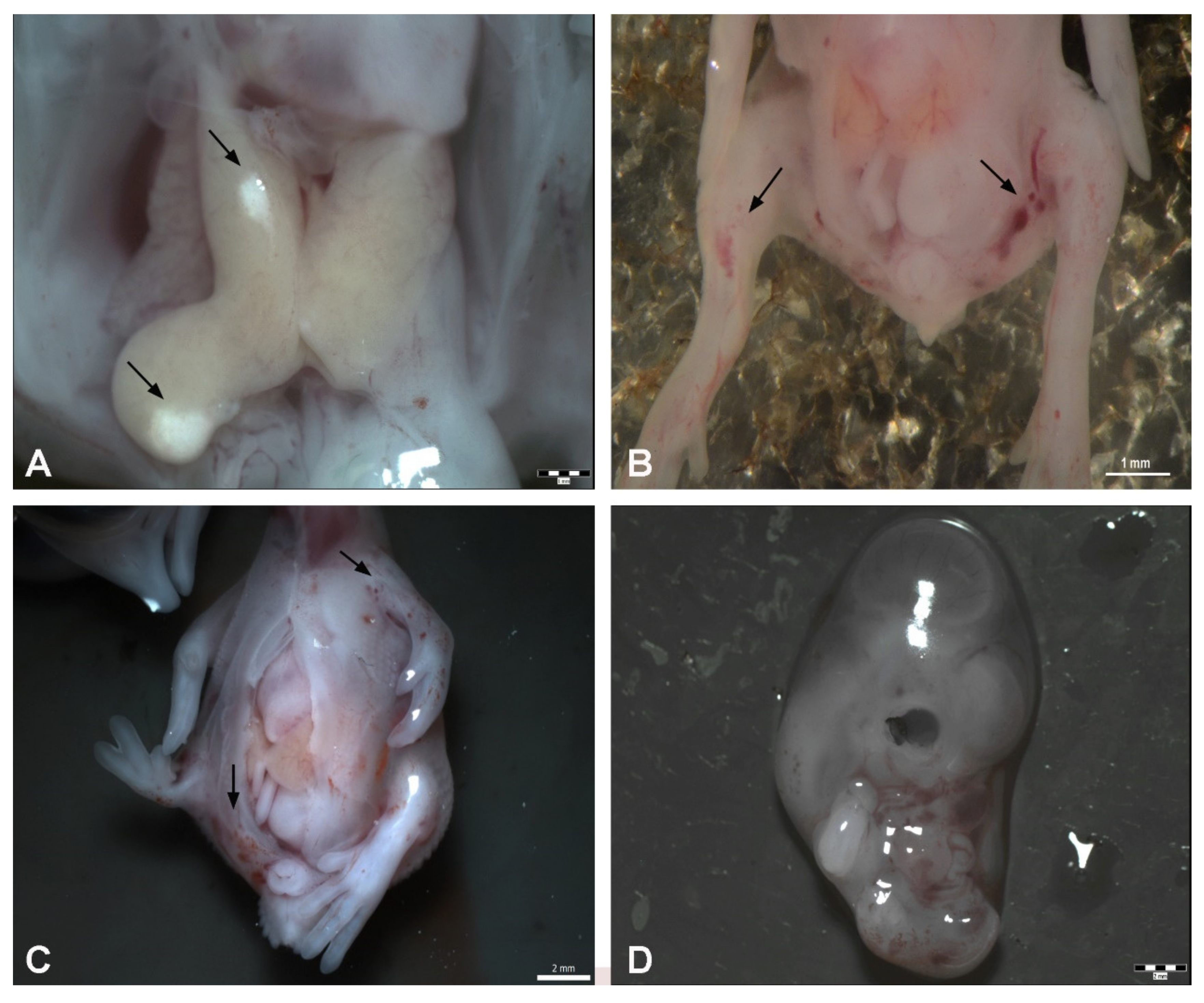
3. Results
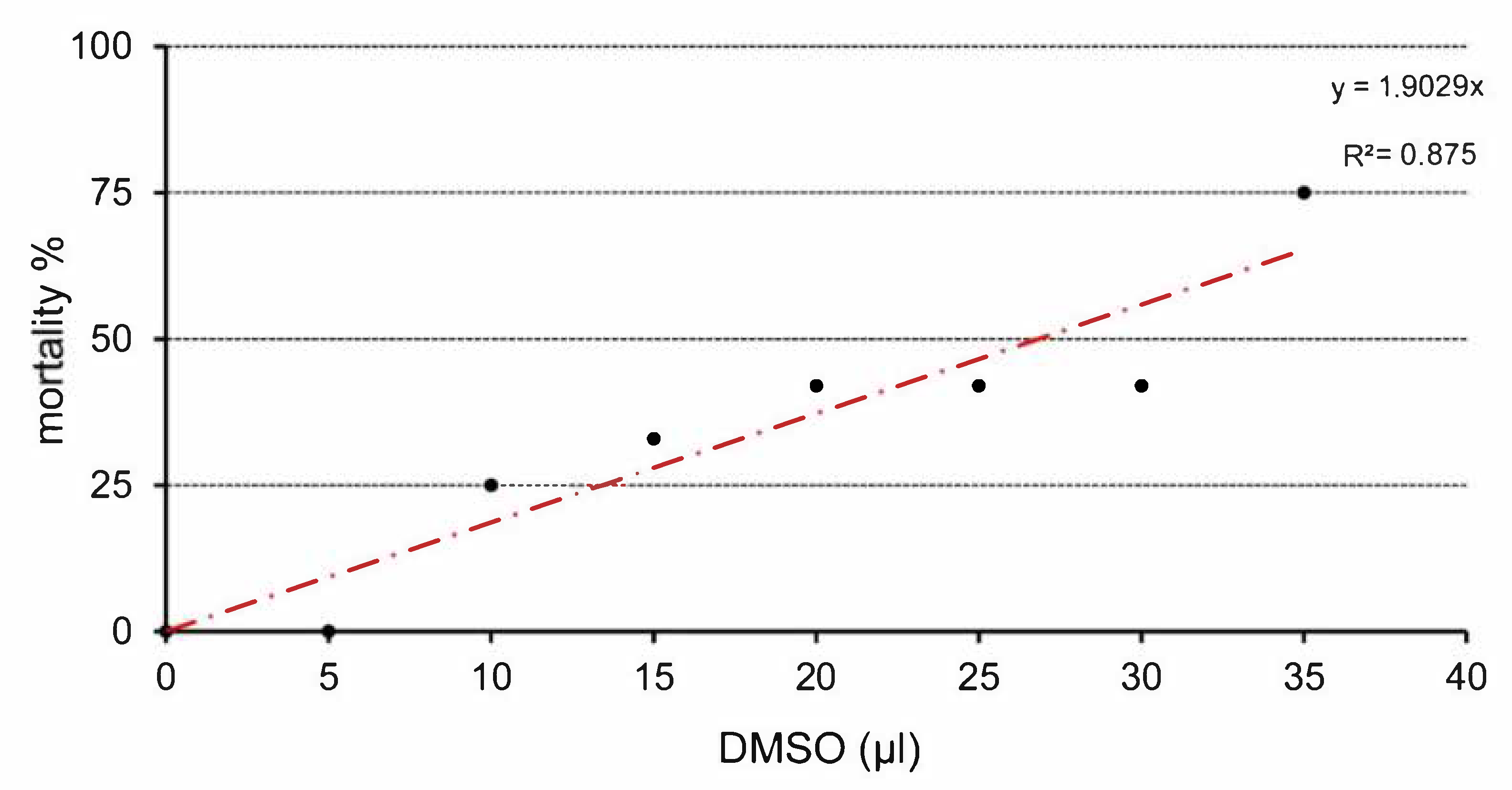
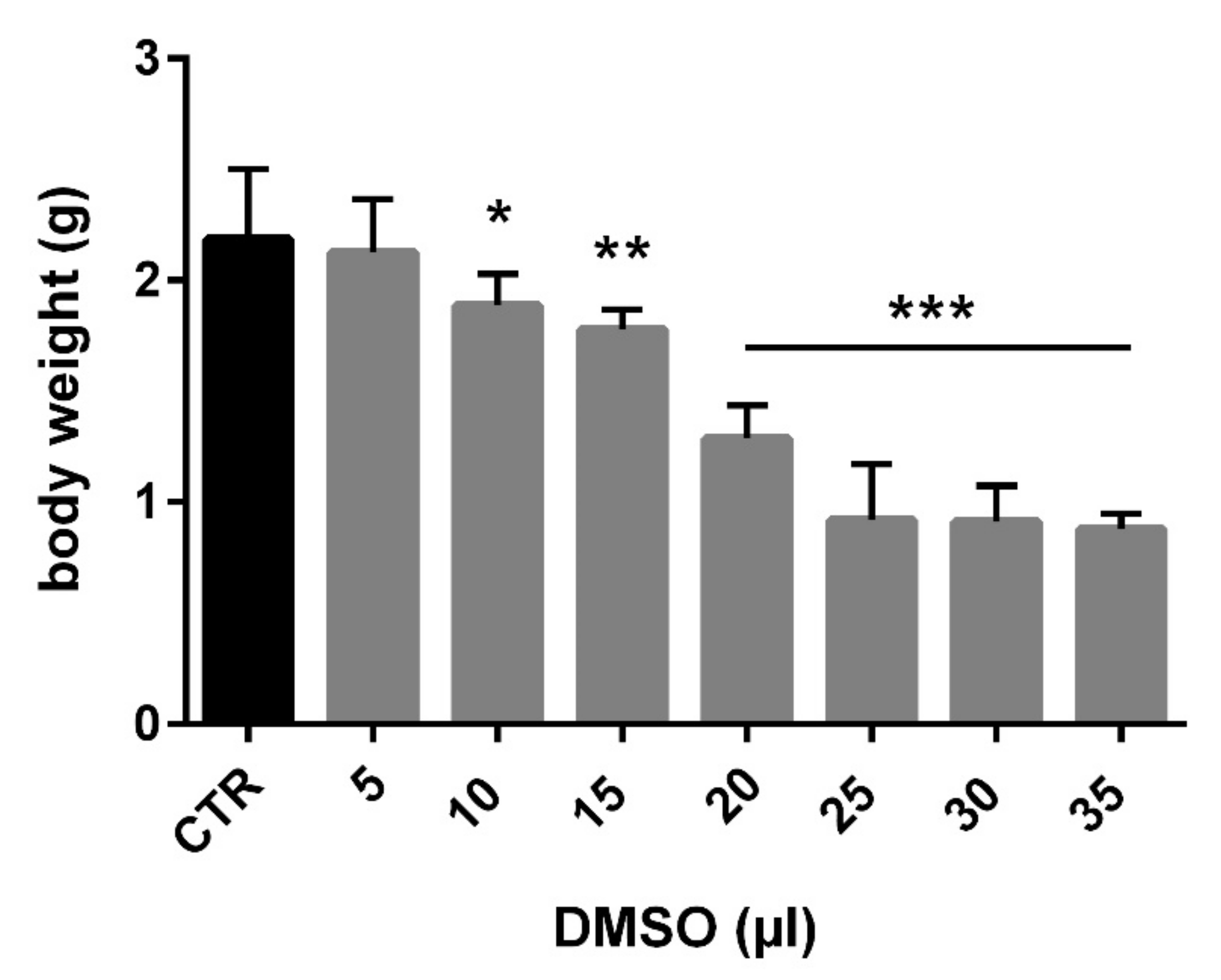
3.1. Total Embryotoxicity
3.2. Histology
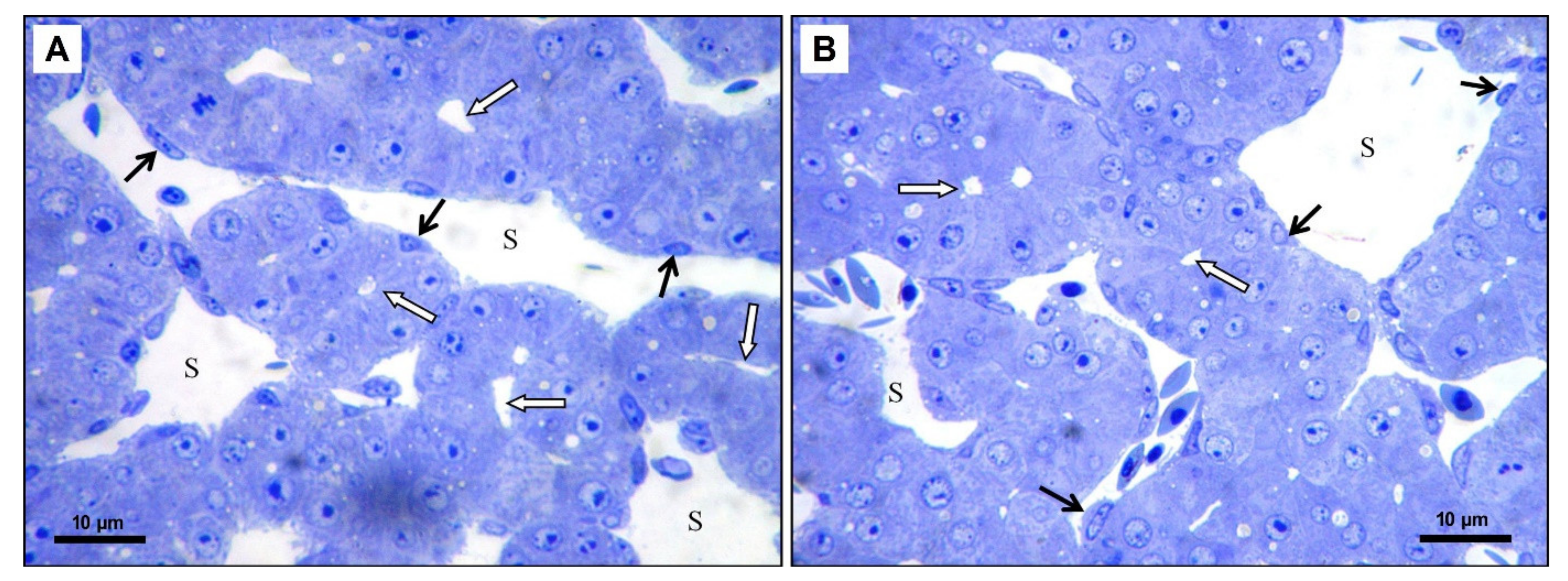
3.2.1. Light Microscopy
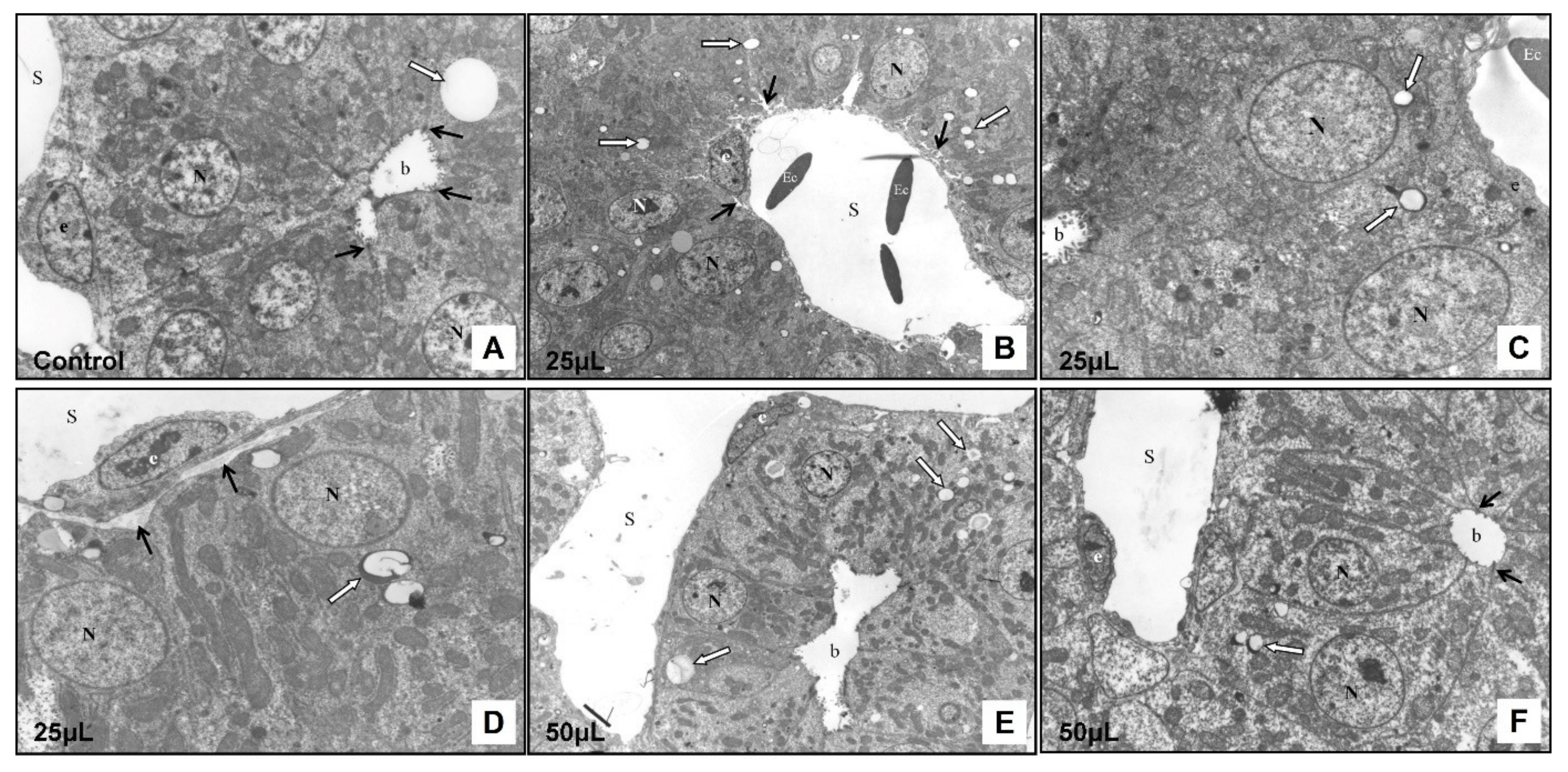
3.2.2. Transmission Electron Microscopy
3.3. Gene Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Galvao, J.; Davis, B.; Tilley, M.; Normando, E.; Duchen, M.R.; Cordeiro, M.F. Unexpected low-dose toxicity of the universal solvent DMSO. FASEB J. 2014, 28, 1317–1330. [Google Scholar] [CrossRef]
- Verheijen, M.; Lienhard, M.; Schrooders, Y.; Clayton, O.; Nudischer, R.; Boerno, S.; Timmermann, B.; Selevsek, N.; Schlapbach, R.; Gmuender, H.; et al. DMSO induces drastic changes in human cellular processes and epigenetic landscape in vitro. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef]
- Chen, T.-H.; Wang, Y.-H.; Wu, Y.-H. Developmental exposures to ethanol or dimethylsulfoxide at low concentrations alter locomotor activity in larval zebrafish: Implications for behavioral toxicity bioassays. Aquat. Toxicol. 2011, 102, 162–166. [Google Scholar] [CrossRef] [PubMed]
- Madsen, B.K.; Hilscher, M.; Zetner, D.; Rosenberg, J. Adverse reactions of dimethyl sulfoxide in humans: A systematic review. F1000 Res. 2018, 7, 1746. [Google Scholar] [CrossRef]
- Watanabe, K.P.; Kawai, Y.K.; Ikenaka, Y.; Kawata, M.; Ikushiro, S.-I.; Sakaki, T.; Ishizuka, M. Avian Cytochrome P450 (CYP) 1-3 Family Genes: Isoforms, Evolutionary Relationships, and mRNA Expression in Chicken Liver. PLoS ONE 2013, 8, e75689. [Google Scholar] [CrossRef]
- Zanger, U.M.; Schwab, M. Cytochrome P450 enzymes in drug metabolism: Regulation of gene expression, enzyme activities, and impact of genetic variation. Pharmacol. Ther. 2013, 138, 103–141. [Google Scholar] [CrossRef] [PubMed]
- Gannon, M.; Gilday, D.; Rifkind, A.B. TCDD Induces CYP1A4 and CYP1A5 in Chick Liver and Kidney and Only CYP1A4, an Enzyme Lacking Arachidonic Acid Epoxygenase Activity, in Myocardium and Vascular Endothelium. Toxicol. Appl. Pharmacol. 2000, 164, 24–37. [Google Scholar] [CrossRef]
- Lee, J.-S.; Kim, E.-Y.; Iwata, H. Dioxin activation of CYP1A5 promoter/enhancer regions from two avian species, common cormorant (Phalacrocorax carbo) and chicken (Gallus gallus): Association with aryl hydrocarbon receptor 1 and 2 isoforms. Toxicol. Appl. Pharmacol. 2009, 234, 1–13. [Google Scholar] [CrossRef]
- Head, J.A.; Kennedy, S.W. Differential expression, induction, and stability of CYP1A4 and CYP1A5 mRNA in chicken and herring gull embryo hepatocytes. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2007, 145, 617–624. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.X.; Jones, S.P.; Iwata, H.; Kim, E.-Y.; Kennedy, S.W. Effects of co-exposure to 2,3,7,8-tetrachlorodibenzo-p-dioxin and perfluorooctane sulfonate or perfluorooctanoic acid on expression of cytochrome P450 isoforms in chicken (Gallus gallus) embryo hepatocyte cultures. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2009, 149, 605–612. [Google Scholar] [CrossRef]
- Fathi, M.A.; Han, G.; Kang, R.; Shen, D.; Shen, J.; Li, C. Disruption of cytochrome P450 enzymes in the liver and small intestine in chicken embryos in ovo exposed to glyphosate. Environ. Sci. Pollut. Res. 2020, 27, 16865–16875. [Google Scholar] [CrossRef]
- Yang, J.; An, J.; Li, M.; Hou, X.; Qiu, X. Characterization of chicken cytochrome P450 1A4 and 1A5: Inter-paralog comparisons of substrate preference and inhibitor selectivity. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2013, 157, 337–343. [Google Scholar] [CrossRef]
- Kapelyukh, Y.; Henderson, C.J.; Scheer, N.; Rode, A.; Wolf, C.R. Defining the Contribution of CYP1A1 and CYP1A2 to Drug Metabolism Using Humanized CYP1A1/1A2 and Cyp1a1/Cyp1a2 Knockout Mice. Drug Metab. Dispos. 2019, 47, 907–918. [Google Scholar] [CrossRef] [PubMed]
- Shang, S.; Jiang, J.; Deng, Y. Chicken Cytochrome P450 1A5 Is the Key Enzyme for Metabolizing T-2 Toxin to 3’OH-T-2. Int. J. Mol. Sci. 2013, 14, 10809–10818. [Google Scholar] [CrossRef]
- Ourlin, J.-C.; Baader, M.; Fraser, D.; Halpert, J.R.; Meyer, U.A. Cloning and Functional Expression of a First Inducible Avian Cytochrome P450 of the CYP3A Subfamily (CYP3A37). Arch. Biochem. Biophys. 2000, 373, 375–384. [Google Scholar] [CrossRef]
- Yuan, Y.; Zhou, X.; Yang, J.; Li, M.; Qiu, X. T-2 toxin is hydroxylated by chicken CYP3A37. Food Chem. Toxicol. 2013, 62, 622–627. [Google Scholar] [CrossRef] [PubMed]
- Greenblatt, D.J.; Zhao, Y.; Venkatakrishnan, K.; Duan, S.X.; Harmatz, J.S.; Parent, S.J.; Court, M.H.; von Moltke, L.L. Mechanism of cytochrome P450-3A inhibition by ketoconazole. J. Pharm. Pharmacol. 2011, 63, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Fagerberg, L.; Hallström, B.M.; Oksvold, P.; Kampf, C.; Djureinovic, D.; Odeberg, J.; Habuka, M.; Tahmasebpoor, S.; Danielsson, A.; Edlund, K.; et al. Analysis of the Human Tissue-specific Expression by Genome-wide Integration of Transcriptomics and Antibody-based Proteomics. Mol. Cell. Proteom. 2014, 13, 397–406. [Google Scholar] [CrossRef]
- Uhlén, M.; Fagerberg, L.; Hallström, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, Å.; Kampf, C.; Sjöstedt, E.; Asplund, A.; et al. Tissue-based map of the human proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef] [PubMed]
- Kawai, Y.K.; Itou, K.; Yoshino, T.; Iima, H.; Matsumoto, F.; Kubota, A. Hepatic transcriptional profile and tissue distribution of cytochrome P450 1-3 genes in the red-crowned crane Grus japonensis. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2020, 228, 108643. [Google Scholar] [CrossRef]
- Geng, W.; Long, S.L.; Chang, Y.-J.; Saxton, A.M.; Joyce, S.A.; Lin, J. Evaluation of bile salt hydrolase inhibitor efficacy for modulating host bile profile and physiology using a chicken model system. Sci. Rep. 2020, 10, 1–20. [Google Scholar] [CrossRef]
- Luo, G.; Guenthner, T.; Gan, L.-S.; Humphreys, W.G. CYP3A4 Induction by Xenobiotics: Biochemistry, Experimental Methods and Impact on Drug Discovery and Development. Curr. Drug Metab. 2004, 5, 483–505. [Google Scholar] [CrossRef]
- Doke, S.K.; Dhawale, S.C. Alternatives to animal testing: A review. Saudi Pharm. J. 2015, 23, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Ribatti, D.; Annese, T.; Tamma, R. The use of the chick embryo CAM assay in the study of angiogenic activiy of biomaterials. Microvasc. Res. 2020, 131, 104026. [Google Scholar] [CrossRef]
- Kue, C.S.; Tan, K.Y.; Lam, M.L.; Lee, H.B. Chick embryo chorioallantoic membrane (CAM): An alternative predictive model in acute toxicological studies for anti-cancer drugs. Exp. Anim. 2015, 64, 129–138. [Google Scholar] [CrossRef]
- Tavakkoli, H.; Attaran, R.; Khosravi, A.; Salari, Z.; Salarkia, E.; Dabiri, S.; Mosallanejad, S.S. Vascular alteration in relation to fosfomycine: In silico and in vivo investigations using a chick embryo model. Biomed. Pharmacother. 2019, 118, 109240. [Google Scholar] [CrossRef] [PubMed]
- Rashidi, H.; Sottile, V. The chick embryo: Hatching a model for contemporary biomedical research. BioEssays 2009, 31, 459–465. [Google Scholar] [CrossRef]
- Psychoyos, D.; Finnell, R. Method for Culture of Early Chick Embryos ex vivo (New Culture). J. Vis. Exp. 2008, 20, e903. [Google Scholar] [CrossRef]
- Kiecker, C. The chick embryo as a model for the effects of prenatal exposure to alcohol on craniofacial development. Dev. Biol. 2016, 415, 314–325. [Google Scholar] [CrossRef] [PubMed]
- Merckx, M.G.; Tay, M.H.; Monaco, M.M.L.; van Zandvoort, M.A.; De Spiegelaere, W.; Lambrichts, I.; Bronckaers, A. Chorioallantoic Membrane Assay as Model for Angiogenesis in Tissue Engineering: Focus on Stem Cells. Tissue Eng. Part B Rev. 2020, 26, 519–539. [Google Scholar] [CrossRef] [PubMed]
- Sedmera, D.; Hu, N.; Weiss, K.M.; Keller, B.B.; Denslow, S.; Thompson, R.P. Cellular changes in experimental left heart hypoplasia. Anat. Rec. Adv. Integr. Anat. Evol. Biol. 2002, 267, 137–145. [Google Scholar] [CrossRef]
- Clark, E.B.; Hu, N.; Dummett, J.L.; VandeKieft, G.K.; Olson, C.; Tomanek, R. Ventricular function and morphology in chick embryo from stages 18 to 29. Am. J. Physiol. Circ. Physiol. 1986, 250, H407–H413. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.L.; Zhang, J.R.; Yu, Z.G.; Zhao, J.; Mo, F.; Jiang, S.X. Effects of ionophores on liver CYP1A and 3A in male broilers. J. Vet. Pharmacol. Ther. 2010, 33, 551–557. [Google Scholar] [CrossRef]
- Caujolle, F.M.E.; Caujolle, D.H.; Cros, S.B.; Calvet, M.-M.J. Limits of toxic and teratogenic tolerance of dimethyl sulfoxide. Ann. N. Y. Acad. Sci. 1967, 141, 110–125. [Google Scholar] [CrossRef]
- Srinivas, S.; Sironmani, T.; Shanmugam, G. Dimethyl sulfoxide inhibits the expression of early growth-response genes and arrests fibroblasts at quiescence. Exp. Cell Res. 1991, 196, 279–286. [Google Scholar] [CrossRef]
- Liu, J.; Yoshikawa, H.; Nakajima, Y.; Tasaka, K. Involvement of mitochondrial permeability transition and caspase-9 activation in dimethyl sulfoxide-induced apoptosis of EL-4 lymphoma cells. Int. Immunopharmacol. 2001, 1, 63–74. [Google Scholar] [CrossRef]
- Pal, R.; Mamidi, M.K.; Das, A.K.; Bhonde, R. Diverse effects of dimethyl sulfoxide (DMSO) on the differentiation potential of human embryonic stem cells. Arch. Toxicol. 2012, 86, 651–661. [Google Scholar] [CrossRef]
- Biagioli, M.; Pifferi, S.; Ragghianti, M.; Bucci, S.; Rizzuto, R.; Pinton, P. Endoplasmic reticulum stress and alteration in calcium homeostasis are involved in cadmium-induced apoptosis. Cell Calcium 2008, 43, 184–195. [Google Scholar] [CrossRef] [PubMed]
- La Rovere, R.M.; Roest, G.; Bultynck, G.; Parys, J.B. Intracellular Ca2+ signaling and Ca2+ microdomains in the control of cell survival, apoptosis and autophagy. Cell Calcium 2016, 60, 74–87. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.-H.; Das, J.; Gurunathan, S.; Park, H.-W.; Song, H.; Park, C.; Kim, J.-H. The cytotoxic effects of dimethyl sulfoxide in mouse preimplantation embryos: A mechanistic study. Theranostics 2017, 7, 4735–4752. [Google Scholar] [CrossRef]
- Welte, M.A. Proteins under new management: Lipid droplets deliver. Trends Cell Biol. 2007, 17, 363–369. [Google Scholar] [CrossRef] [PubMed]
- Murphy, S.; Martin, S.; Parton, R.G. Lipid droplet-organelle interactions; sharing the fats. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2009, 1791, 441–447. [Google Scholar] [CrossRef] [PubMed]
- Goto, Y.; Noda, Y.; Mori, T.; Nakano, M. Increased generation of reactive oxygen species in embryos cultured in vitro. Free Radic. Biol. Med. 1993, 15, 69–75. [Google Scholar] [CrossRef]
- Sadowska-Bartosz, I.; Pączka, A.; Mołoń, M.; Bartosz, G. Dimethyl sulfoxide induces oxidative stress in the yeast Saccharomyces cerevisiae. FEMS Yeast Res. 2013, 13, 820–830. [Google Scholar] [CrossRef]
- He, L.; He, T.; Farrar, S.; Ji, L.; Liu, T.; Ma, X. Antioxidants Maintain Cellular Redox Homeostasis by Elimination of Reactive Oxygen Species. Cell. Physiol. Biochem. 2017, 44, 532–553. [Google Scholar] [CrossRef]
- Jarc, E.; Petan, T. Lipid Droplets and the Management of Cellular Stress. Yale J. Biol. Med. 2019, 92, 435–452. [Google Scholar]
- Rambold, A.S.; Cohen, S.; Lippincott-Schwartz, J. Fatty Acid Trafficking in Starved Cells: Regulation by Lipid Droplet Lipolysis, Autophagy, and Mitochondrial Fusion Dynamics. Dev. Cell 2015, 32, 678–692. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.B.; Louie, S.M.; Daniele, J.R.; Tran, Q.; Dillin, A.; Zoncu, R.; Nomura, D.K.; Olzmann, J.A. DGAT1-Dependent Lipid Droplet Biogenesis Protects Mitochondrial Function during Starvation-Induced Autophagy. Dev. Cell 2017, 42, 9–21.e5. [Google Scholar] [CrossRef]
- Tunçer, S.; Gurbanov, R.; Sheraj, I.; Solel, E.; Esenturk, O.; Banerjee, S. Low dose dimethyl sulfoxide driven gross molecular changes have the potential to interfere with various cellular processes. Sci. Rep. 2018, 8, 14828. [Google Scholar] [CrossRef] [PubMed]
- Fujii-Kuriyama, Y.; Mimura, J. Molecular mechanisms of AhR functions in the regulation of cytochrome P450 genes. Biochem. Biophys. Res. Commun. 2005, 338, 311–317. [Google Scholar] [CrossRef]
- Zhang, H.F.; Lin, X.H.; Yang, H.; Zhou, L.C.; Guo, Y.L.; Barnett, J.V.; Guo, Z.M. Regulation of the Activity and Expression of Aryl Hydrocarbon Receptor by Ethanol in Mouse Hepatic Stellate Cells. Alcohol. Clin. Exp. Res. 2012, 36, 1873–1881. [Google Scholar] [CrossRef]
- Su, T.; Waxman, D.J. Impact of dimethyl sulfoxide on expression of nuclear receptors and drug-inducible cytochromes P450 in primary rat hepatocytes. Arch. Biochem. Biophys. 2004, 424, 226–234. [Google Scholar] [CrossRef] [PubMed]
- Honkakoski, P.; Negishi, M. Regulation of cytochrome P450 (CYP) genes by nuclear receptors. Biochem. J. 2000, 347, 321–337. [Google Scholar] [CrossRef] [PubMed]
- Handschin, C.; Podvinec, M.; Meyer, U.A. CXR, a chicken xenobiotic-sensing orphan nuclear receptor, is related to both mammalian pregnane X receptor (PXR) and constitutive androstane receptor (CAR). Proc. Natl. Acad. Sci. USA 2000, 97, 10769–10774. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dose (µL) | N | Dead | Mortality (%) | Malformations | Mean Body Weight (g) | Body Weight SD |
|---|---|---|---|---|---|---|
| 0 | 12 | 0 | 0 | 0 | 2.185 | 0.316 |
| 5 | 11 | 0 | 0 | 0 | 2.126 | 0.241 |
| 10 | 12 | 3 | 25 | 0 | 1.886 | 0.142 |
| 15 | 12 | 4 | 33 | 1 | 1.779 | 0.091 |
| 20 | 12 | 5 | 42 | 0 | 1.285 | 0.150 |
| 25 | 12 | 5 | 42 | 5 | 0.918 | 0.252 |
| 30 | 12 | 5 | 42 | 6 | 0.911 | 0.164 |
| 35 | 12 | 9 | 75 | 3 | 0.880 | 0.069 |
| 95 | 31 | 33 | 15 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luptakova, L.; Dvorcakova, S.; Demcisakova, Z.; Belbahri, L.; Holovska, K.; Petrovova, E. Dimethyl Sulfoxide: Morphological, Histological, and Molecular View on Developing Chicken Liver. Toxics 2021, 9, 55. https://doi.org/10.3390/toxics9030055
Luptakova L, Dvorcakova S, Demcisakova Z, Belbahri L, Holovska K, Petrovova E. Dimethyl Sulfoxide: Morphological, Histological, and Molecular View on Developing Chicken Liver. Toxics. 2021; 9(3):55. https://doi.org/10.3390/toxics9030055
Chicago/Turabian StyleLuptakova, Lenka, Simona Dvorcakova, Zuzana Demcisakova, Lassaad Belbahri, Katarina Holovska, and Eva Petrovova. 2021. "Dimethyl Sulfoxide: Morphological, Histological, and Molecular View on Developing Chicken Liver" Toxics 9, no. 3: 55. https://doi.org/10.3390/toxics9030055
APA StyleLuptakova, L., Dvorcakova, S., Demcisakova, Z., Belbahri, L., Holovska, K., & Petrovova, E. (2021). Dimethyl Sulfoxide: Morphological, Histological, and Molecular View on Developing Chicken Liver. Toxics, 9(3), 55. https://doi.org/10.3390/toxics9030055