
Fenugreek Seed Galactomannan Aqueous and Extract Protects against Diabetic Nephropathy and Liver Damage by Targeting NF-κB and Keap1/Nrf2 Axis


Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Induction of Diabetes Mellitus
2.3. Preparation of the Fenugreek Seeds Aqueous Extract (FS-AE)
2.4. Preparation and Isolation of F-GAL
2.5. Experimental Design
2.6. Dose Selection
2.7. Urine and Blood Tissue Collection and Processing
2.8. Measurement of Glucose and Insulin Levels
2.9. Analysis of Kidney and Liver Functions
2.10. Preparation of Liver Homogenates and Biochemical Analysis
2.11. Extraction of Hepatic Lipids
2.12. Lipid Profile in the Serum and Livers
2.13. Quantitate Real-Time Polymerase Chain Reaction (q-PCR)
2.14. Histological Evaluation
2.15. Statistical Analysis
3. Results
3.1. Changes in Metabolic Parameters
3.2. Changes in Serum and Hepatic Lipids
3.3. Changes in Serum Hepatic and Renal Function Markers
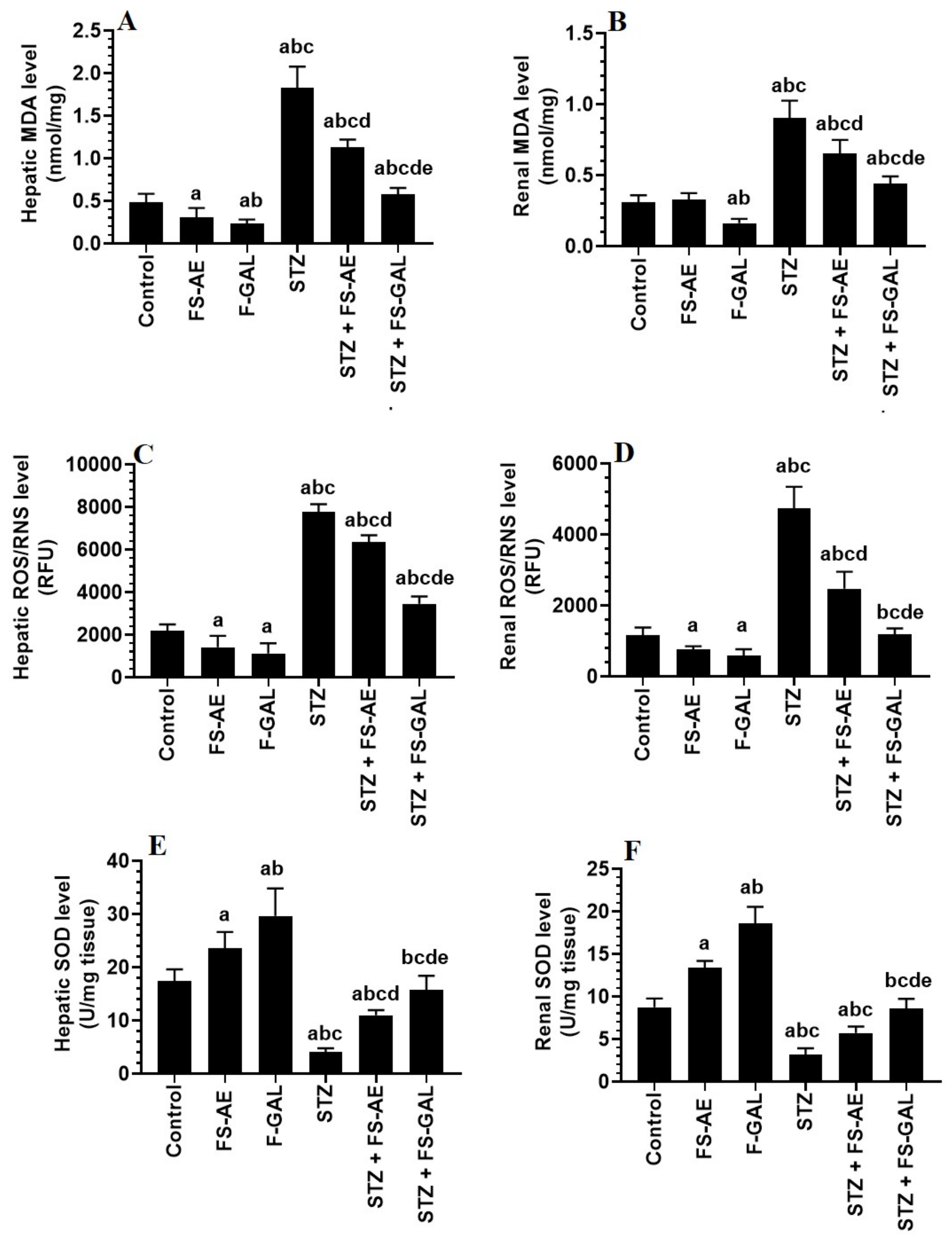
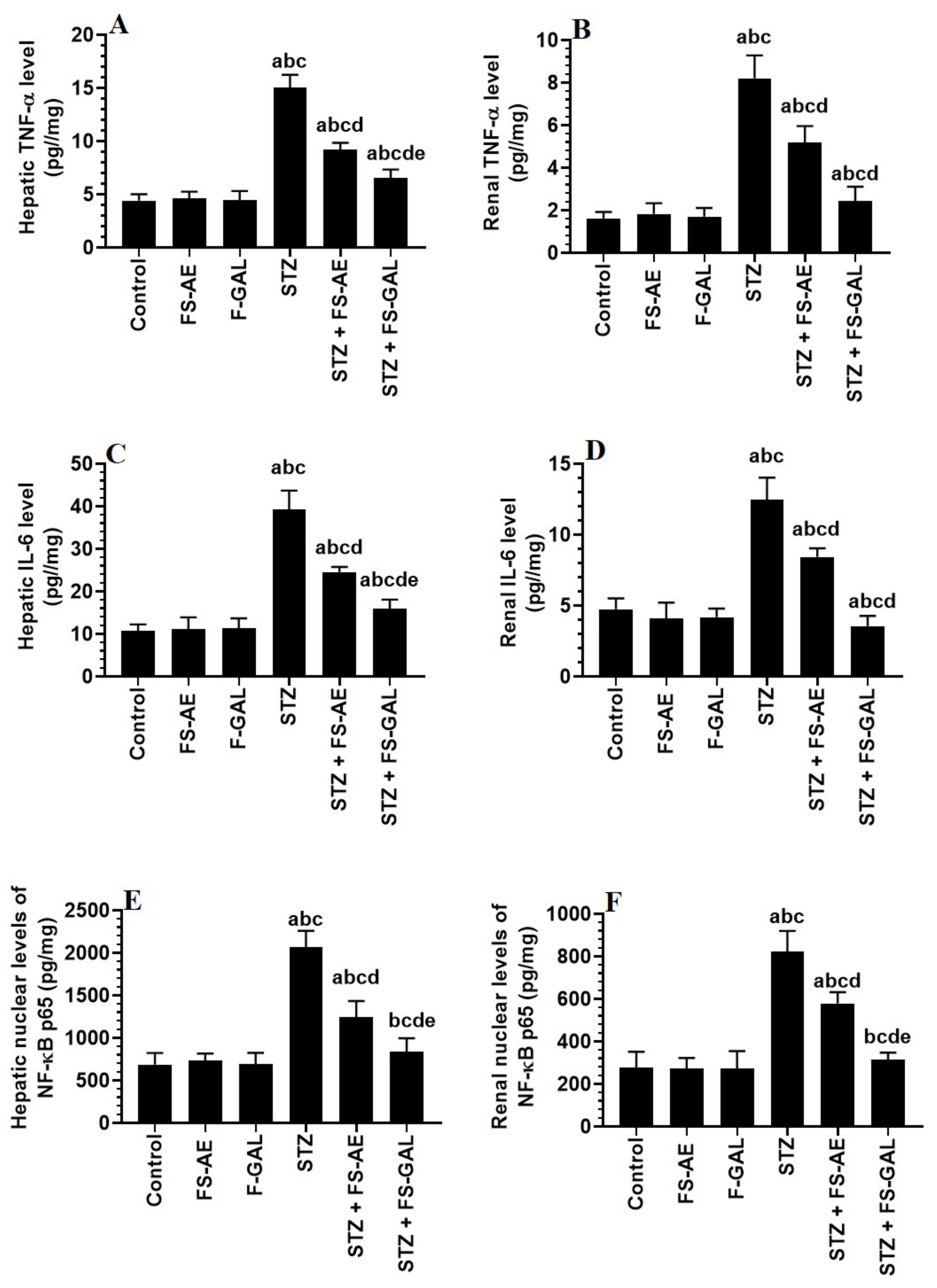
3.4. Changes in Hepatic and Renal Oxidative Stress/Antioxidant Markers
3.5. Changes in Hepatic and Renal Oxidative Stress/Antioxidant Markers
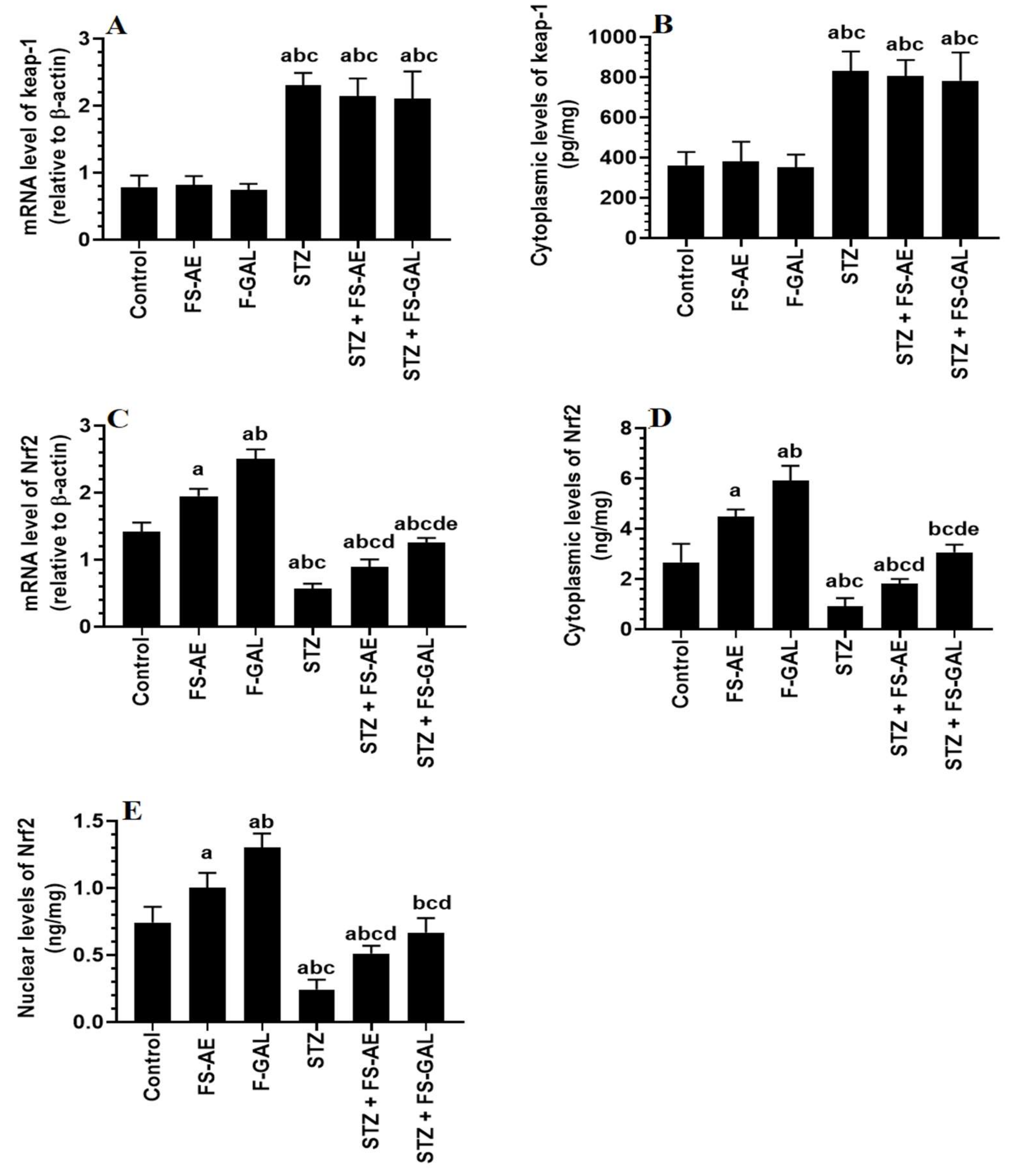
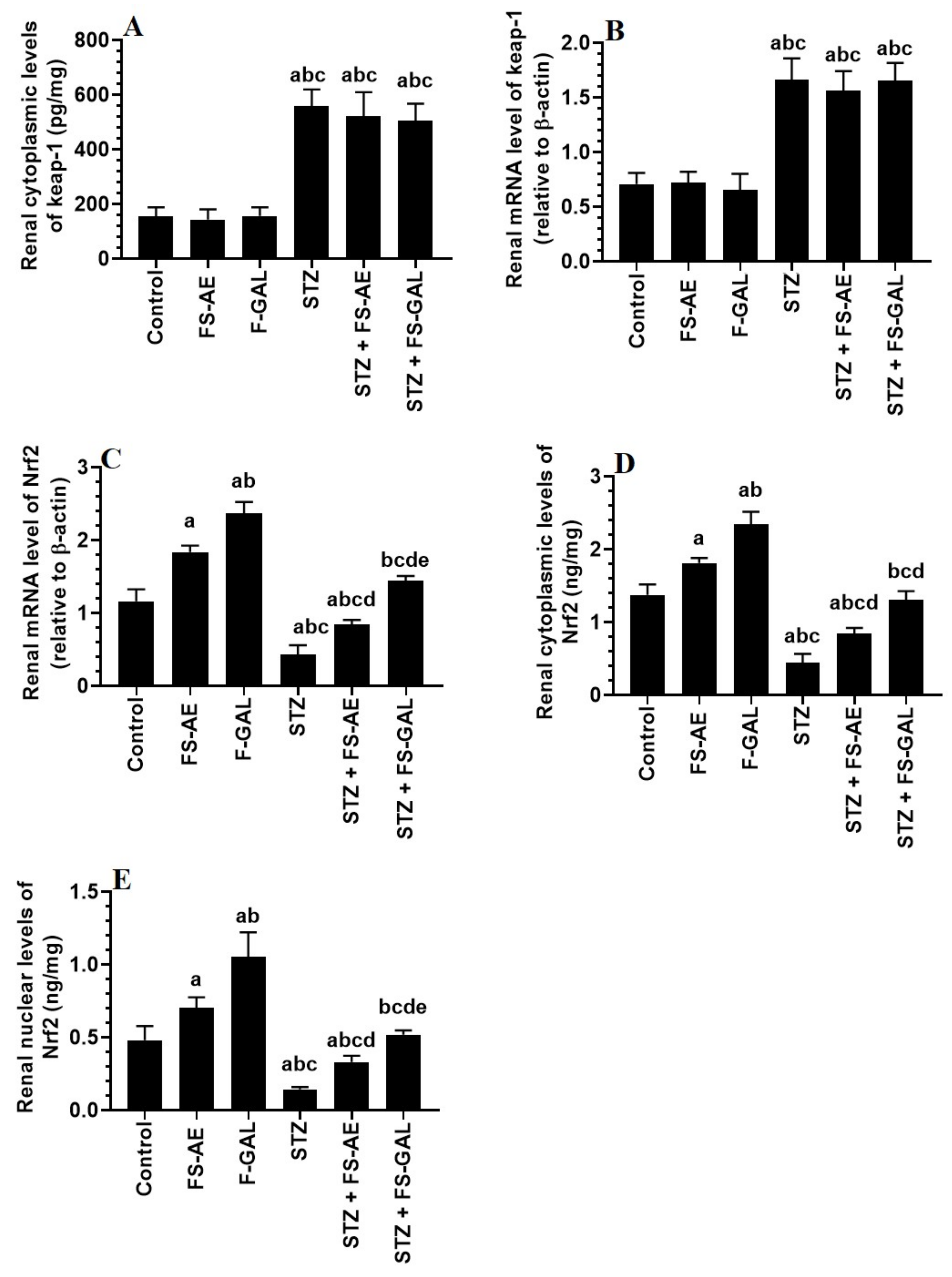
3.6. Effect on Hepatic and Renal Keap1/Nrf2 Signaling
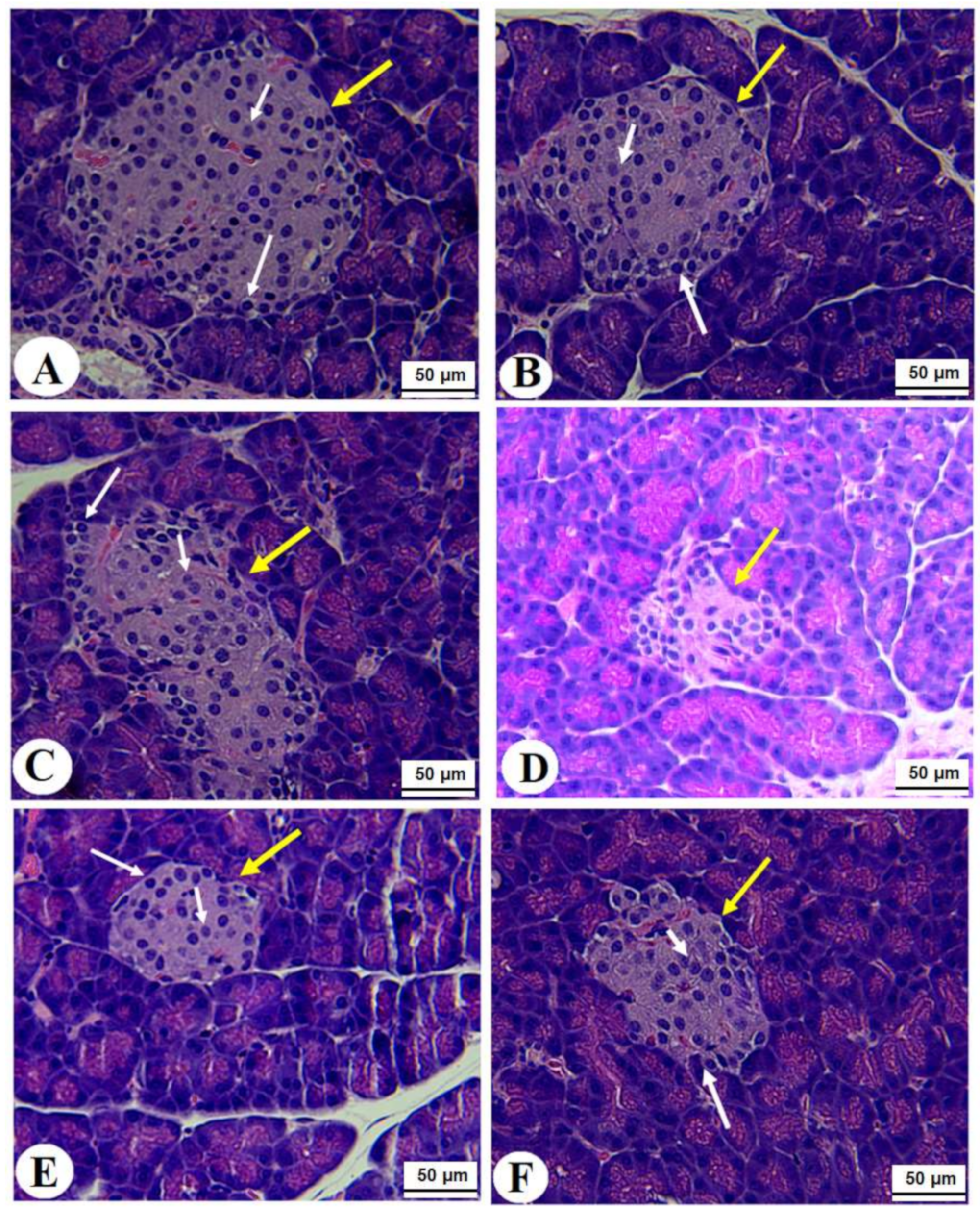
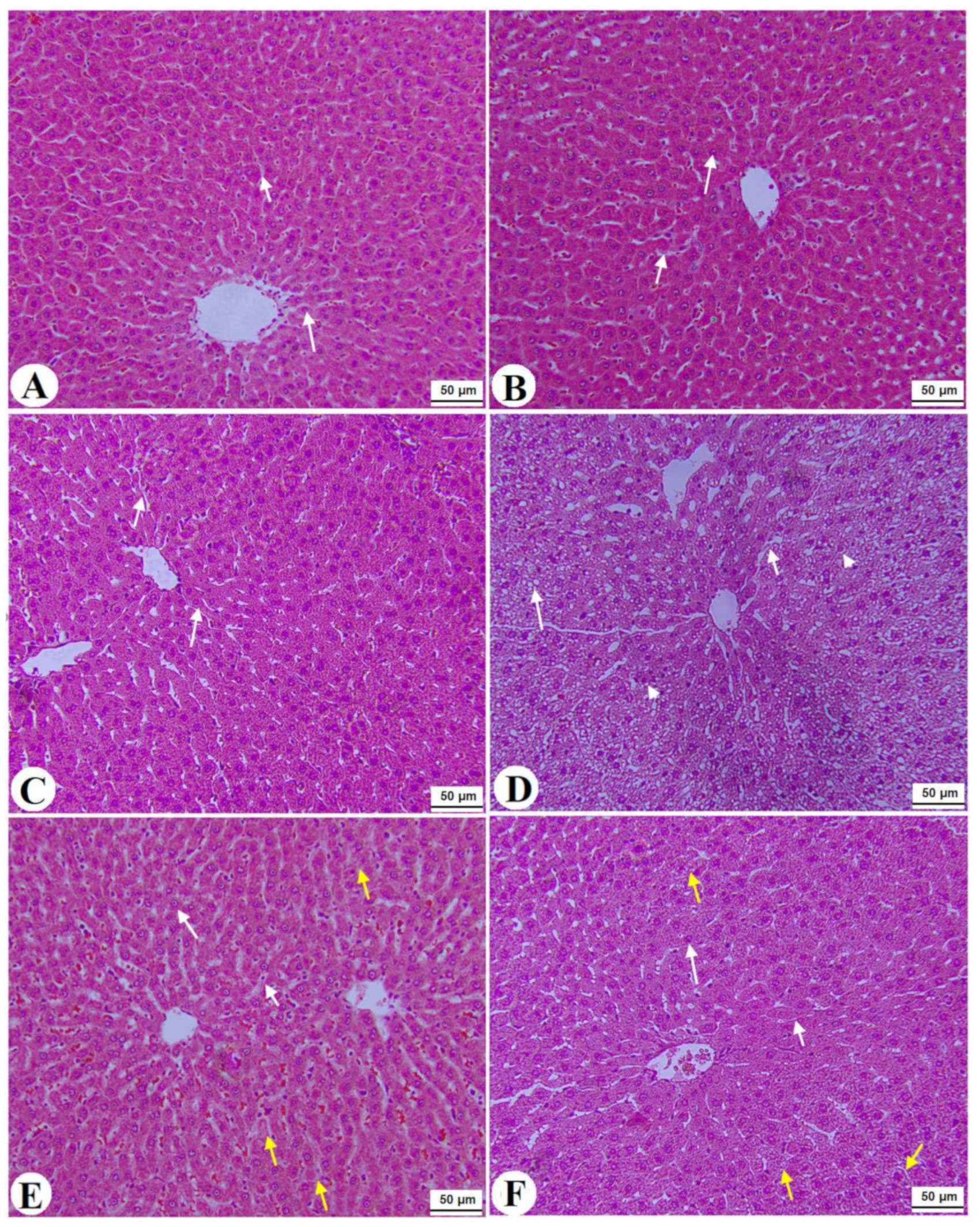
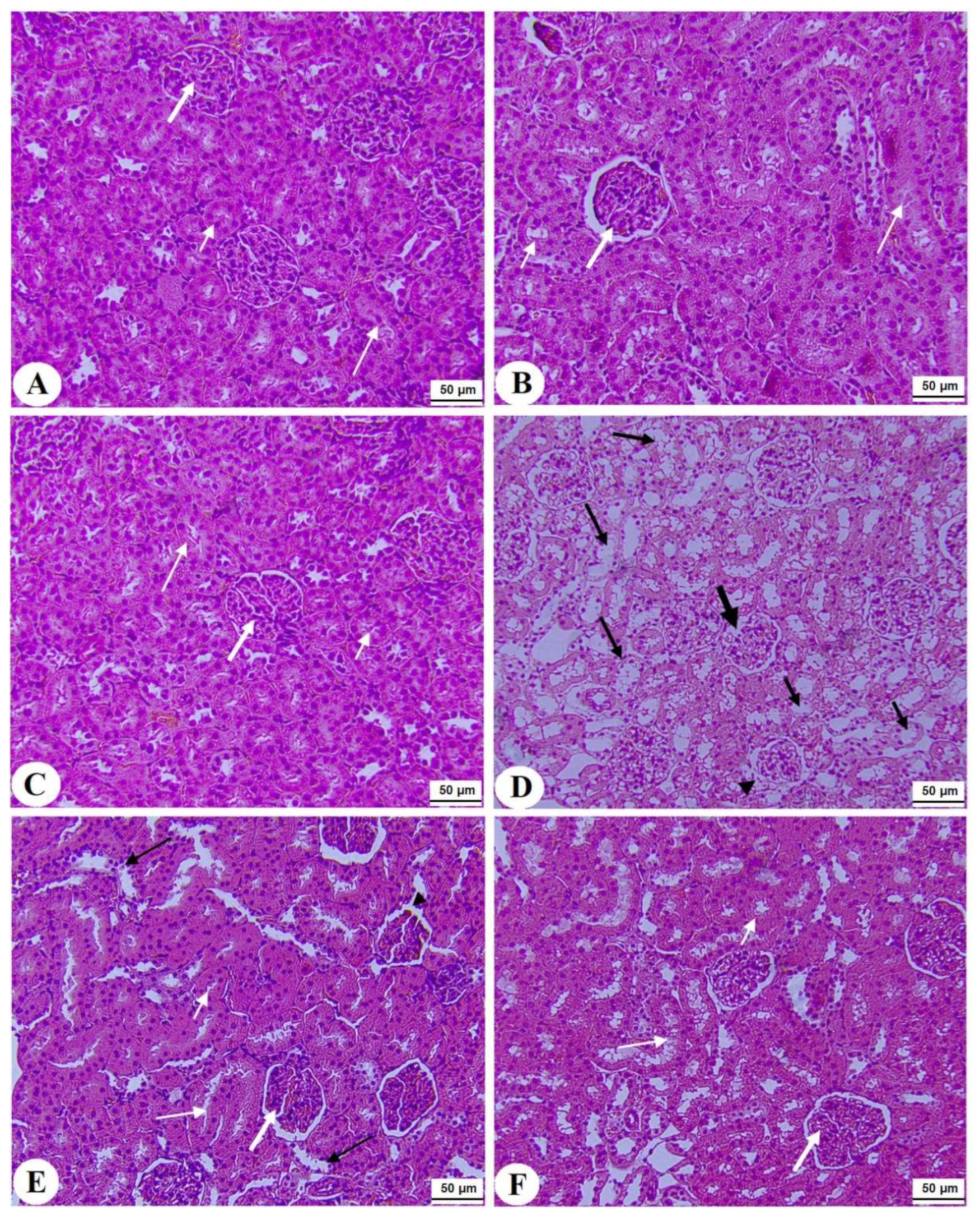
3.7. Histological Alterations
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Asmat, U.; Abad, K.; Ismail, K. Diabetes mellitus and oxidative stress-a concise review. Saudi Pharm. J. 2016, 24, 547–553. [Google Scholar] [CrossRef] [PubMed]
- Volpe, C.M.O.; Villar-Delfino, P.H.; Dos Anjos, P.M.F.; Nogueira-Machado, J.A. Cellular death, reactive oxygen species (ROS) and diabetic complications. Cell Death Dis. 2018, 9, 119. [Google Scholar] [CrossRef] [PubMed]
- Hacioglu, C.; Kar, F.; Kara, Y.; Yucel, E.; Donmez, D.B.; Sentürk, H.; Kanbak, G. Comparative effects of metformin and Cistus laurifolius L. extract in streptozotocin-induced diabetic rat model: Oxidative, inflammatory, apoptotic, and histopathological analyzes. Environ. Sci. Pollut. Res. 2021, 28, 57888–57901. [Google Scholar] [CrossRef]
- Barros, B.S.V.; Conte Santos, D.; Haas Pizarro, M.; Melo, L.G.N.D.; Brito Gomes, M. Type 1 Diabetes and Non-Alcoholic Fatty Liver Disease: When Should We Be Concerned? A Nationwide Study in Brazil. Nutrients 2017, 9, 878. [Google Scholar] [CrossRef]
- Dharmalingam, M.; Yamasandhi, P.G. Nonalcoholic Fatty Liver Disease and Type 2 Diabetes Mellitus. Indian J. Endocrinol. Metab. 2018, 22, 421–428. [Google Scholar] [CrossRef]
- Giacco, F.; Brownlee, M. Oxidative stress and diabetic complications. Circ. Res. 2010, 107, 1058–1070. [Google Scholar] [CrossRef]
- Oguntibeju, O.O. Type 2 diabetes mellitus, oxidative stress and inflammation: Examining the links. Int. J. Physiol. Pathophysiol. Pharmacol. 2019, 11, 45–63. [Google Scholar]
- Othman, M.S.; Khaled, A.M.; Al-Bagawi, A.H.; Fareid, M.A.; Ghany, R.A.; Habotta, O.A.; Moneim, A.E.A. Hepatorenal protective efficacy of flavonoids from Ocimum basilicum extract in diabetic albino rats: A focus on hypoglycemic, antioxidant, anti-inflammatory and anti-apoptotic activities. Biomed. Pharmacother. 2021, 144, 112287. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Lei, H.; Yang, F.; Fan, X.; Dang, Q.; Li, Y. Activation of the bile acid receptor GPBAR1 (TGR5) ameliorates interleukin-1β (IL-1β)-induced chondrocytes senescence. Biomed. Pharmacother. 2018, 106, 1713–1719. [Google Scholar] [CrossRef]
- Xu, Z.; Wei, Y.; Gong, J.; Cho, H.; Park, J.K.; Sung, E.-R.; Huang, H.; Wu, L.; Eberhart, C.; Handa, J.T. NRF2 plays a protective role in diabetic retinopathy in mice. Diabetologia 2014, 57, 204–213. [Google Scholar] [CrossRef]
- Lazaro, I.; Lopez-Sanz, L.; Bernal, S.; Oguiza, A.; Recio, C.; Melgar, A.; Jimenez-Castilla, L.; Egido, J.; Madrigal-Matute, J.; Gomez-Guerrero, C. Nrf2 activation provides atheroprotection in diabetic mice through concerted upregulation of antioxidant, anti-inflammatory, and autophagy mechanisms. Front. Pharmacol. 2018, 9, 819. [Google Scholar] [CrossRef] [PubMed]
- Giribabu, N.; Karim, K.; Kilari, E.K.; Salleh, N. Phyllanthus niruri leaves aqueous extract improves kidney functions, ameliorates kidney oxidative stress, inflammation, fibrosis and apoptosis and enhances kidney cell proliferation in adult male rats with diabetes mellitus. J. Ethnopharmacol. 2017, 205, 123–137. [Google Scholar] [CrossRef] [PubMed]
- Panasiuk, A.; Dzieciol, J.; Panasiuk, B.; Prokopowicz, D. Expression of p53, Bax and Bcl-2 proteins in hepatocytes in non-alcoholic fatty liver disease. World J. Gastroenterol. 2006, 12, 6198–6202. [Google Scholar] [CrossRef] [PubMed]
- Wagener, F.A.D.T.G.; Dekker, D.; Berden, J.H.; Scharstuhl, A.; van der Vlag, J. The role of reactive oxygen species in apoptosis of the diabetic kidney. Apoptosis 2009, 14, 1451–1458. [Google Scholar] [CrossRef] [PubMed]
- Fusco, R.; Cordaro, M.; Genovese, T.; Impellizzeri, D.; Siracusa, R.; Gugliandolo, E.; Peritore, A.F.; D’Amico, R.; Crupi, R.; Cuzzocrea, S.; et al. Adelmidrol: A New Promising Antioxidant and Anti-Inflammatory Therapeutic Tool in Pulmonary Fibrosis. Antioxidants 2020, 9, 601. [Google Scholar] [CrossRef] [PubMed]
- Fusco, R.; Cordaro, M.; Siracusa, R.; D’Amico, R.; Genovese, T.; Gugliandolo, E.; Peritore, A.F.; Crupi, R.; Impellizzeri, D.; Cuzzocrea, S.; et al. Biochemical Evaluation of the Antioxidant Effects of Hydroxytyrosol on Pancreatitis-Associated Gut Injury. Antioxidants 2020, 9, 781. [Google Scholar] [CrossRef] [PubMed]
- Zeng, F.; Luo, J.; Han, H.; Xie, W.; Wang, L.; Han, R.; Chen, H.; Cai, Y.; Huang, H.; Xia, Z. Allopurinol ameliorates liver injury in type 1 diabetic rats through activating Nrf2. Int. J. Immunopathol. Pharmacol. 2021, 35, 20587384211031417. [Google Scholar] [CrossRef] [PubMed]
- Jiang, T.; Huang, Z.; Lin, Y.; Zhang, Z.; Fang, D.; Zhang, D.D. The protective role of Nrf2 in streptozotocin-induced diabetic nephropathy. Diabetes 2010, 59, 850–860. [Google Scholar] [CrossRef] [PubMed]
- Negi, G.; Kumar, A.; Joshi, R.P.; Sharma, S.S. Oxidative stress and Nrf2 in the pathophysiology of diabetic neuropathy: Old perspective with a new angle. Biochem. Biophys. Res. Commun. 2011, 408, 1–5. [Google Scholar] [CrossRef]
- Elosta, A.; Ghous, T.; Ahmed, N. Natural products as anti-glycation agents: Possible therapeutic potential for diabetic complications. Curr. Diabetes Rev. 2012, 8, 92–108. [Google Scholar] [CrossRef]
- Babu, P.V.A.; Liu, D.; Gilbert, E.R. Recent advances in understanding the anti-diabetic actions of dietary flavonoids. J. Nutr. Biochem. 2013, 24, 1777–1789. [Google Scholar] [CrossRef] [PubMed]
- Geberemeskel, G.A.; Debebe, Y.G.; Nguse, N.A. Antidiabetic effect of fenugreek seed powder solution (Trigonella foenum-graecum L.) on hyperlipidemia in diabetic patients. J. Diabetes Res. 2019, 2019, 8507453. [Google Scholar] [CrossRef] [PubMed]
- Zargar, S. Protective effect of Trigonella foenum-graecum on thioacetamide induced hepatotoxicity in rats. Saudi J. Biol. Sci. 2014, 21, 139–145. [Google Scholar] [CrossRef]
- Mbarki, S.; Alimi, H.; Bouzenna, H.; Elfeki, A.; Hfaiedh, N. Phytochemical study and protective effect of Trigonella foenum graecum (Fenugreek seeds) against carbon tetrachloride-induced toxicity in liver and kidney of male rat. Biomed. Pharmacother. 2017, 88, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, U.N.; Chandra, D. The plant extracts of Momordica charantia and Trigonella foenum graecum have antioxidant and anti-hyperglycemic properties for cardiac tissue during diabetes mellitus. Oxid. Med. Cell. Longev. 2009, 2, 290–296. [Google Scholar] [CrossRef] [PubMed]
- Haeri, M.R.; Limaki, H.K.; White, C.J.B.; White, K.N. Non-insulin dependent anti-diabetic activity of (2S, 3R, 4S) 4-hydroxyisoleucine of fenugreek (Trigonella foenum graecum) in streptozotocin-induced type I diabetic rats. Phytomedicine 2012, 19, 571–574. [Google Scholar] [CrossRef]
- Gaddam, A.; Galla, C.; Thummisetti, S.; Marikanty, R.K.; Palanisamy, U.D.; Rao, P.V. Role of Fenugreek in the prevention of type 2 diabetes mellitus in prediabetes. J. Diabetes Metab. Disord. 2015, 14, 74. [Google Scholar] [CrossRef]
- Bera, T.K.; Ali, K.M.; Jana, K.; Ghosh, A.; Ghosh, D. Protective effect of aqueous extract of seed of Psoralea corylifolia (Somraji) and seed of Trigonella foenum-graecum L.(Methi) in streptozotocin-induced diabetic rat: A comparative evaluation. Pharmacogn. Res. 2013, 5, 277–285. [Google Scholar]
- Joshi, D.V.; Patil, R.R.; Naik, S.R. Hydroalcohol extract of Trigonella foenum-graecum seed attenuates markers of inflammation and oxidative stress while improving exocrine function in diabetic rats. Pharm. Biol. 2015, 53, 201–211. [Google Scholar] [CrossRef]
- Shetty, A.K.; Salimath, P.V. Reno-protective effects of fenugreek (Trigonella foenum greacum) during experimental diabetes. e-SPEN Eur. e-J. Clin. Nutr. Metab. 2009, 4, e137–e142. [Google Scholar] [CrossRef][Green Version]
- Jin, Y.; Shi, Y.; Zou, Y.; Miao, C.; Sun, B.; Li, C. Fenugreek prevents the development of STZ-induced diabetic nephropathy in a rat model of diabetes. Evid. Based Complement. Altern. Med. 2014, 2014, 259368. [Google Scholar] [CrossRef] [PubMed]
- Kandhare, A.D.; Bodhankar, S.L.; Mohan, V.; Thakurdesai, P.A. Effect of glycosides based standardized fenugreek seed extract in bleomycin-induced pulmonary fibrosis in rats: Decisive role of Bax, Nrf2, NF-κB, Muc5ac, TNF-α and IL-1β. Chem. Biol. Interact. 2015, 237, 151–165. [Google Scholar] [CrossRef] [PubMed]
- Hamza, A.; Elwy, H.; Badawi, A. Fenugreek seed extract attenuates cisplatin-induced testicular damage in W istar rats. Andrologia 2016, 48, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Hannan, J.; Ali, L.; Rokeya, B.; Khaleque, J.; Akhter, M.; Flatt, P.; Abdel-Wahab, Y. Soluble dietary fibre fraction of Trigonella foenum-graecum (fenugreek) seed improves glucose homeostasis in animal models of type 1 and type 2 diabetes by delaying carbohydrate digestion and absorption, and enhancing insulin action. Br. J. Nutr. 2007, 97, 514–521. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Gao, L.; Li, P.; Kan, H.; Qu, J.; Men, L.; Liu, Z.; Liu, Z. Metabonomics study of the therapeutic mechanism of fenugreek galactomannan on diabetic hyperglycemia in rats, by ultra-performance liquid chromatography coupled with quadrupole time-of-flight mass spectrometry. J. Chromatogr. B 2017, 1044, 8–16. [Google Scholar] [CrossRef]
- Kamble, H.; Kandhare, A.D.; Bodhankar, S.; Mohan, V.; Thakurdesai, P. Effect of low molecular weight galactomannans from fenugreek seeds on animal models of diabetes mellitus. Biomed. Aging Pathol. 2013, 3, 145–151. [Google Scholar] [CrossRef]
- Anwar, S.; Desai, S.; Mandlik, R. Exploring antidiabetic mechanisms of action of galactomannan: A carbohydrate isolated from fenugreek seeds. J. Complement. Integr. Med. 2009, 6, 1553–3840. [Google Scholar] [CrossRef]
- Altamimi, J.; Alfaris, N.; Alshammari, G.; Alagal, R.; Aljabryn, D.; Aldera, H.; Alkhateeb, M.; Yahya, M. Ellagic acid protects against diabetic cardiomyopathy in rats by stimulating cardiac silent information regulator 1 signaling. J. Physiol. Pharm. 2020, 71, 891–904. [Google Scholar]
- Berroukche, A.; Terras, M.; Kharoubi, M.; Boudia, H.; Kharoubi, O.; Dellaoui, H.; Lansari, W.; Zerarki, I. Assessing of funegreek (Trigonella foenumgraecum) seeds pharmacological effects against glucotoxicity and lipotoxicity in streptozotocine-induced diabetic rats. Diabetes Updates 2018, 5, 1–5. [Google Scholar]
- Al-Attar, A.M.; Alsalmi, F.A. Influence of olive leaves extract on hepatorenal injury in streptozotocin diabetic rats. Saudi J. Biol. Sci. 2019, 26, 1865–1874. [Google Scholar] [CrossRef]
- Furukawa, S.; Kobayashi, Y. Leaner Women with Impaired Insulin Secretion Accounts for about 40 of Gestational Diabetes Mellitus in Japan. J. Pregnancy 2019, 2019, 7578403. [Google Scholar] [CrossRef] [PubMed]
- Heidari, R.; Taheri, V.; Rahimi, H.R.; Shirazi Yeganeh, B.; Niknahad, H.; Najibi, A. Sulfasalazine-induced renal injury in rats and the protective role of thiol-reductants. Ren. Fail. 2016, 38, 137–141. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Wu, K.; Wu, X.; Xin, C.; Minghui Zhou, M.; Jin Lei, J.; Chen, J. Punicalagin alleviates brain injury and inflammatory responses, and regulates HO-1/Nrf-2/ARE signaling in rats after experimental intracerebral haemorrhage. Trop. J. Pharm. Res. 2020, 19, 727–737. [Google Scholar] [CrossRef]
- Alsabaani, N.A.; Osman, O.M.; Dallak, M.A.; Morsy, M.D.; Al-Dhibi, H.A. Maslinic Acid Protects against Streptozotocin-Induced Diabetic Retinopathy by Activating Nrf2 and Suppressing NF-κB. J. Ophthalmol. 2022, 2022, 3044202. [Google Scholar] [CrossRef] [PubMed]
- Eid, R.A.; Zaki, M.S.A.; Alaa Eldeen, M.; Alshehri, M.M.; Shati, A.A.; El-kott, A.F. Exendin-4 protects the hearts of rats from ischaemia/reperfusion injury by boosting antioxidant levels and inhibition of JNK/p66 Shc/NADPH axis. Clin. Exp. Pharm. Physiol. 2020, 47, 1240–1253. [Google Scholar] [CrossRef] [PubMed]
- Alshehri, A.S.; El-Kott, A.F.; Eleawa, S.M.; El-Gerbed, M.S.A.; Khalifa, H.S.; El-Kenawy, A.E.; Albadrani, G.M.; Abdel-Daim, M.M. Kaempferol protects against streptozotocin-induced diabetic cardiomyopathy in rats by a hypoglycemic effect and upregulating SIRT1. J. Physiol. Pharmacol. 2021, 72, 339–355. [Google Scholar]
- Alshammari, G.M.; Al-Qahtani, W.H.; AlFaris, N.A.; Albekairi, N.A.; Alqahtani, S.; Eid, R.; Yagoub, A.E.A.; Al-Harbi, L.N.; Yahya, M.A. Quercetin alleviates cadmium chloride-induced renal damage in rats by suppressing endoplasmic reticulum stress through SIRT1-dependent deacetylation of Xbp-1s and eIF2α. Biomed. Pharmacother. 2021, 141, 111862. [Google Scholar] [CrossRef]
- Al-Qahtani, W.H.; Alshammari, G.M.; Ajarem, J.S.; Al-Zahrani, A.Y.; Alzuwaydi, A.; Eid, R.; Yahya, M.A. Isoliquiritigenin prevents Doxorubicin-induced hepatic damage in rats by upregulating and activating SIRT1. Biomed. Pharmacother. 2022, 146, 112594. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Lorenzati, B.; Zucco, C.; Miglietta, S.; Lamberti, F.; Bruno, G. Oral hypoglycemic drugs: Pathophysiological basis of their mechanism of actionoral hypoglycemic drugs: Pathophysiological basis of their mechanism of action. Pharmaceuticals 2010, 3, 3005–3020. [Google Scholar] [CrossRef]
- Saltiel, A.R.; Kahn, C.R. Insulin signalling and the regulation of glucose and lipid metabolism. Nature 2001, 414, 799–806. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Yan, L.-J. Streptozotocin-induced type 1 diabetes in rodents as a model for studying mitochondrial mechanisms of diabetic β cell glucotoxicity. Diabetes Metab. Syndr. Obes. Targets Ther. 2015, 8, 181. [Google Scholar]
- Matthews, D.; Hosker, J.; Rudenski, A.; Naylor, B.; Treacher, D.; Turner, R.C. Homeostasis model assessment: Insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef] [PubMed]
- Sauvaire, Y.; Petit, P.; Broca, C.; Manteghetti, M.; Baissac, Y.; Fernandez-Alvarez, J.; Gross, R.; Roye, M.; Leconte, A.; Gomis, R. 4-Hydroxyisoleucine: A novel amino acid potentiator of insulin secretion. Diabetes 1998, 47, 206–210. [Google Scholar] [CrossRef] [PubMed]
- Fuller, S.; Stephens, J.M. Diosgenin, 4-hydroxyisoleucine, and fiber from fenugreek: Mechanisms of actions and potential effects on metabolic syndrome. Adv. Nutr. 2015, 6, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Kalailingam, P.; Kannaian, B.; Tamilmani, E.; Kaliaperumal, R. Efficacy of natural diosgenin on cardiovascular risk, insulin secretion, and beta cells in streptozotocin (STZ)-induced diabetic rats. Phytomedicine 2014, 21, 1154–1161. [Google Scholar] [CrossRef]
- Anwar, S.; Desai, S.; Eidi, M.; Eidi, A. Antidiabetic Activities of Fenugreek (Trigonella Foenum-Graecum) Seeds. In Nuts and Seeds in Health and Disease Prevention; Elsevier: Amsterdam, The Netherlands, 2011; pp. 469–478. [Google Scholar]
- Srichamroen, A.; Field, C.J.; Thomson, A.B.; Basu, T.K. The modifying effects of galactomannan from Canadian-grown fenugreek (Trigonella foenum-graecum L.) on the glycemic and lipidemic status in rats. J. Clin. Biochem. Nutr. 2008, 43, 167–174. [Google Scholar] [CrossRef]
- Faria, A.; Persaud, S.J. Cardiac oxidative stress in diabetes: Mechanisms and therapeutic potential. Pharmacol. Ther. 2017, 172, 50–62. [Google Scholar] [CrossRef]
- Wang, G.G.; Lu, X.H.; Li, W.; Zhao, X.; Zhang, C. Protective effects of luteolin on diabetic nephropathy in STZ-induced diabetic rats. Evid. Based Complement. Altern. Med. 2011, 2011, 323171. [Google Scholar] [CrossRef]
- Abolfathi, A.A.; Mohajeri, D.; Rezaie, A.; Nazeri, M. Protective effects of green tea extract against hepatic tissue injury in streptozotocin-induced diabetic rats. Evid. Based Complement. Altern. Med. 2012, 2012, 740671. [Google Scholar] [CrossRef]
- Belaïd-Nouira, Y.; Bakhta, H.; Haouas, Z.; Flehi-Slim, I.; Neffati, F.; Najjar, M.F.; Cheikh, H.B. Fenugreek seeds, a hepatoprotector forage crop against chronic AlCl 3 toxicity. BMC Vet. Res. 2013, 9, 22. [Google Scholar] [CrossRef] [PubMed]
- Laroubi, A.; Touhami, M.; Farouk, L.; Zrara, I.; Aboufatima, R.; Benharref, A.; Chait, A. Prophylaxis effect of Trigonella foenum graecum L. seeds on renal stone formation in rats. Phytother. Res. 2007, 21, 921–925. [Google Scholar] [CrossRef] [PubMed]
- Shekha, M.S.; Qadir, A.B.; Ali, H.H.; Selim, X.E. Effect of fenugreek (Trigonella foenum-graecum) on ethylene glycol induced kidney stone in rats. Jordan J. Biol. Sci. 2014, 7, 257–260. [Google Scholar] [CrossRef][Green Version]
- Arroyave-Ospina, J.C.; Wu, Z.; Geng, Y.; Moshage, H. Role of oxidative stress in the pathogenesis of non-alcoholic fatty liver disease: Implications for prevention and therapy. Antioxidants 2021, 10, 174. [Google Scholar] [CrossRef]
- Sangeetha, M.; Mal, N.S.; Atmaja, K.; Sali, V.K.; Vasanthi, H.R. PPAR’s and Diosgenin a chemico biological insight in NIDDM. Chem. Biol. Interact. 2013, 206, 403–410. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primers (5′→3′) | GenBank Accession | Product Length |
|---|---|---|---|
| Nrf2 | F: -AAAATCATTAACCTCCCTGTTGAT R: R: ′-CGGCGACTTTATTCTTACCTCTC | NM_031789 | 118 |
| Keap-1 | 5′-TATGAGCCAGAGCGGGACGA-3′ 5′-TCATCCGCCACTCATTCCTCT-3′ | AF304364.1 | 172 |
| B-actin | F: GACCTCTATGCCAACACAGT R: CACCAATCCACACAGAGTAC | NM_031144 | 154 |
| STD | FS-AE | F-GAL | STZ | STZ + FS-AE | STZ + F-GAL | |
|---|---|---|---|---|---|---|
| Final body weight (g) | 418 ± 22 | 422 ± 26 | 409 ± 32 | 313 ± 19 abc | 379 ± 18 abcd | 411 ± 20 de |
| Weekly food intake (g/rat) (Last 2 weeks) | 192 ± 16 | 185 ± 22 | 198 ± 23 | 274 ± 28 abc | 217± 15 abcd | 186 ± 25 de |
| Liver weight (g) | 14.7 ± 1.1 | 13.1 ± 1.6 | 14.7 ± 1.2 | 18.7 ± 2.0 abc | 16.4 ± 0.8 abcd | 14.2 ± 1.1 de |
| Kidney weight (g) | 1.46 ± 0.23 | 1.49 ± 0.32 | 1.36 ± 0.29 | 1.03 ± 0.18 abc | 1.24 ± 0.23 abcd | 1.44 ± 0.22 de |
| Plasma fasting glucose (mg/dL) | 105.6 ± 7.7 | 91.3 ± 6.3 a | 87.1 ± 5.7 a | 302.1 ± 34 abc | 174.6 ± 21 abcd | 124.5 ± 12.1 abcde |
| Plasma fasting insulin (µIU/mL) | 5.5 ± 0.53 | 5.1 ± 0.62 | 5.3 ± 0.59 | 1.8 ± 0.33 abc | 2.87 ± 0.46 abcd | 3.41 ± 0.63 abcde |
| HOMA-β index | 47.2 ± 6.3 | 66.5 ± 6.6 a | 76.5 ± 9.2 ab | 2.8 ± 0.53 abc | 9.8 ± 1.1 abcd | 22.1 ± 3.5 abcde |
| STD | FS-AE | F-GAL | STZ | STZ + FS-AE | STZ + F-GAL | |
|---|---|---|---|---|---|---|
| Serum | ||||||
| TGs (mg/dL) | 54.5 ± 5.4 | 58.9 ± 7.2 | 55.3 ± 8.8 | 121 ± 11.4 abc | 85.4 ± 7.5 abcd | 61.2 ± 5.9 acde |
| CHOL (mg/dL) | 77.5 ± 9.4 | 83.3 ± 7.5 | 79.5 ± 8.1 | 143 ± 12.3 abc | 104 ± 3.2 abcd | 81.4 ± 6.9 de |
| LDL-c (mg/dL) | 33.2 ± 4.3 | 29.6 ± 4.1 | 31.5 ± 4.8 | 71.5 ± 7.7 abc | 53.1 ± 6.8 abcd | 35.7 ± 5.4 bcde |
| HDL-c (mg/dL) | 24.5 ± 4.3 | 35.2 ± 5.1 | 23.7 ± 4.8 | 11.1 ± 2.3 abc | 17.8 ± 2.7 abcd | 25.4 ± 3.1 de |
| FFAs (µmol/L) | 224 ± 14.5 | 216 ± 19.4 | 227 ± 17.6 | 646 ± 35.7 abc | 342 ± 24.7 abcd | 271 ± 19.7 abcde |
| Liver | ||||||
| Triglycerides (µg/g) | 4642 ± 220 | 4593 ± 198 | 4721 ± 318 | 9135 ± 615 abc | 6619 ± 472 abd | 5492 ± 301 abcde |
| CHOL (µg/g) | 1594 ± 152 | 1499 ± 178 | 1518 ± 159 | 4021 ± 328 abc | 2821± 219 abcd | 1612 ± 171 bcde |
| FFAs (µmol/g) | 63.7 ± 7.5 | 59.6 ± 5.9 | 62.5 ± 6.9 | 125.1 ± 10.5 abc | 89.3 ± 6.8 acbd | 71.8 ± 6.6 abcde |
| STD | FS-AE | F-GAL | STZ | STZ + FS-AE | STZ + F-GAL | |
|---|---|---|---|---|---|---|
| ALT (U/L) | 34.3 ± 3.5 | 35.6 ± 4.1 | 32.5 ± 3.9 | 79.7 ± 7.4 abc | 53.4 ± 6.3 abcd | 38.9 ± 3.5 abcde |
| AST (U/L) | 48.7 ± 5.3 | 41.5 ± 6.4 | 47.6 ± 5.9 | 132.1 ± 9.5 abc | 86.5 ± 8.7 abcd | 55.6 ± 5.1 abcde |
| Albumin (g/dL) | 5.9 ± 0.93 | 6.1 ± 1.2 | 5.5 ± 0.89 | 2.3 ± 0.43 abc | 4.3 ± 0.69 abcd | 5.8 ± 0.83 de |
| Urea (mg/dL) | 4.3 ± 0.47 | 4.1 ± 0.61 | 3.9 ± 0.42 | 9.3 ± 1.7 abc | 6.5 ± 1.4 abcd | 4.2 ± 0.79 de |
| Creatinine (mg/dL) | 0.73 ± 0.04 | 0.69 ± 0.05 a | 0.64 ± 0.06 | 2.56 ± 0.71 abc | 1.25 ± 0.16 abcd | 0.95 ± 0.11 abcde |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alsuliam, S.M.; Albadr, N.A.; Almaiman, S.A.; Al-Khalifah, A.S.; Alkhaldy, N.S.; Alshammari, G.M. Fenugreek Seed Galactomannan Aqueous and Extract Protects against Diabetic Nephropathy and Liver Damage by Targeting NF-κB and Keap1/Nrf2 Axis. Toxics 2022, 10, 362. https://doi.org/10.3390/toxics10070362
Alsuliam SM, Albadr NA, Almaiman SA, Al-Khalifah AS, Alkhaldy NS, Alshammari GM. Fenugreek Seed Galactomannan Aqueous and Extract Protects against Diabetic Nephropathy and Liver Damage by Targeting NF-κB and Keap1/Nrf2 Axis. Toxics. 2022; 10(7):362. https://doi.org/10.3390/toxics10070362
Chicago/Turabian StyleAlsuliam, Sarah M., Nawal A. Albadr, Salah A. Almaiman, Abdullrahman S. Al-Khalifah, Noorah S. Alkhaldy, and Ghedeir M. Alshammari. 2022. "Fenugreek Seed Galactomannan Aqueous and Extract Protects against Diabetic Nephropathy and Liver Damage by Targeting NF-κB and Keap1/Nrf2 Axis" Toxics 10, no. 7: 362. https://doi.org/10.3390/toxics10070362
APA StyleAlsuliam, S. M., Albadr, N. A., Almaiman, S. A., Al-Khalifah, A. S., Alkhaldy, N. S., & Alshammari, G. M. (2022). Fenugreek Seed Galactomannan Aqueous and Extract Protects against Diabetic Nephropathy and Liver Damage by Targeting NF-κB and Keap1/Nrf2 Axis. Toxics, 10(7), 362. https://doi.org/10.3390/toxics10070362