
The Anti-Rheumatic Drug, Leflunomide, Induces Nephrotoxicity in Mice via Upregulation of TGFβ-Mediated p53/Smad2/3 Signaling

 ,
,  ,
,  , ,
, ,  and
and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
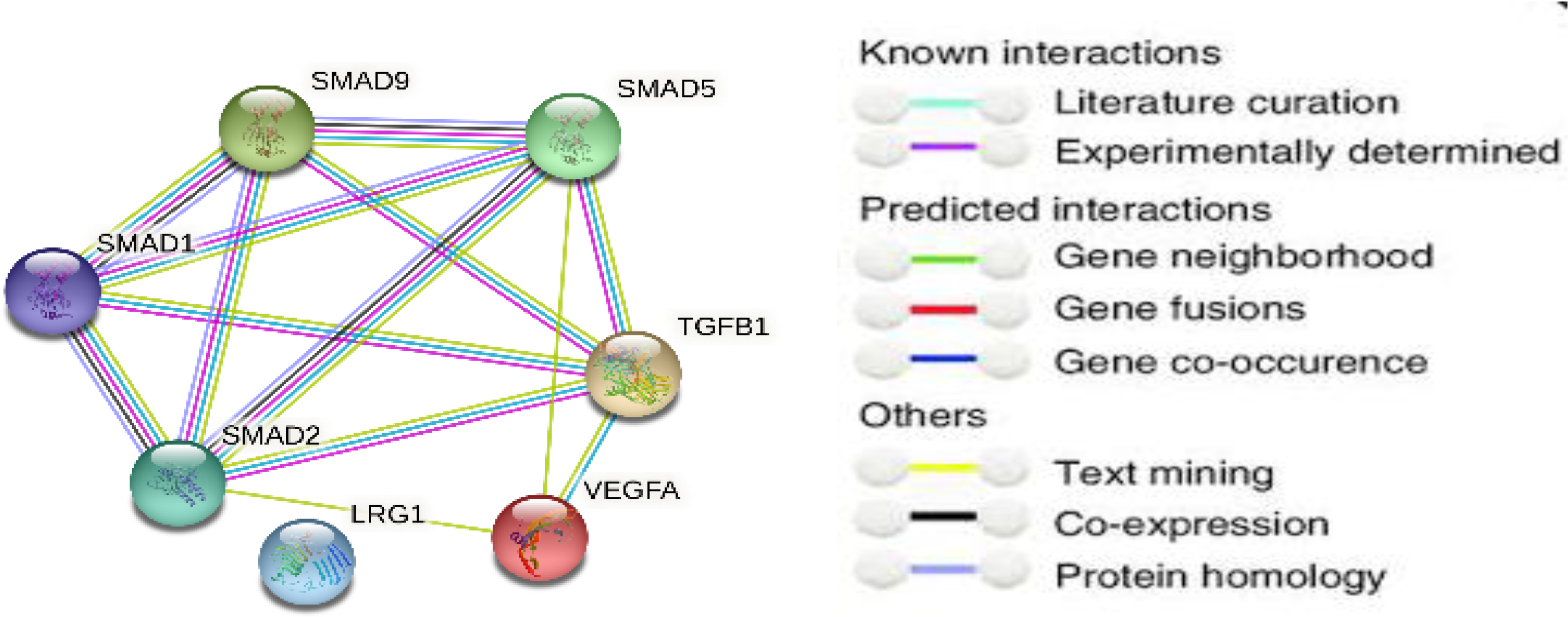
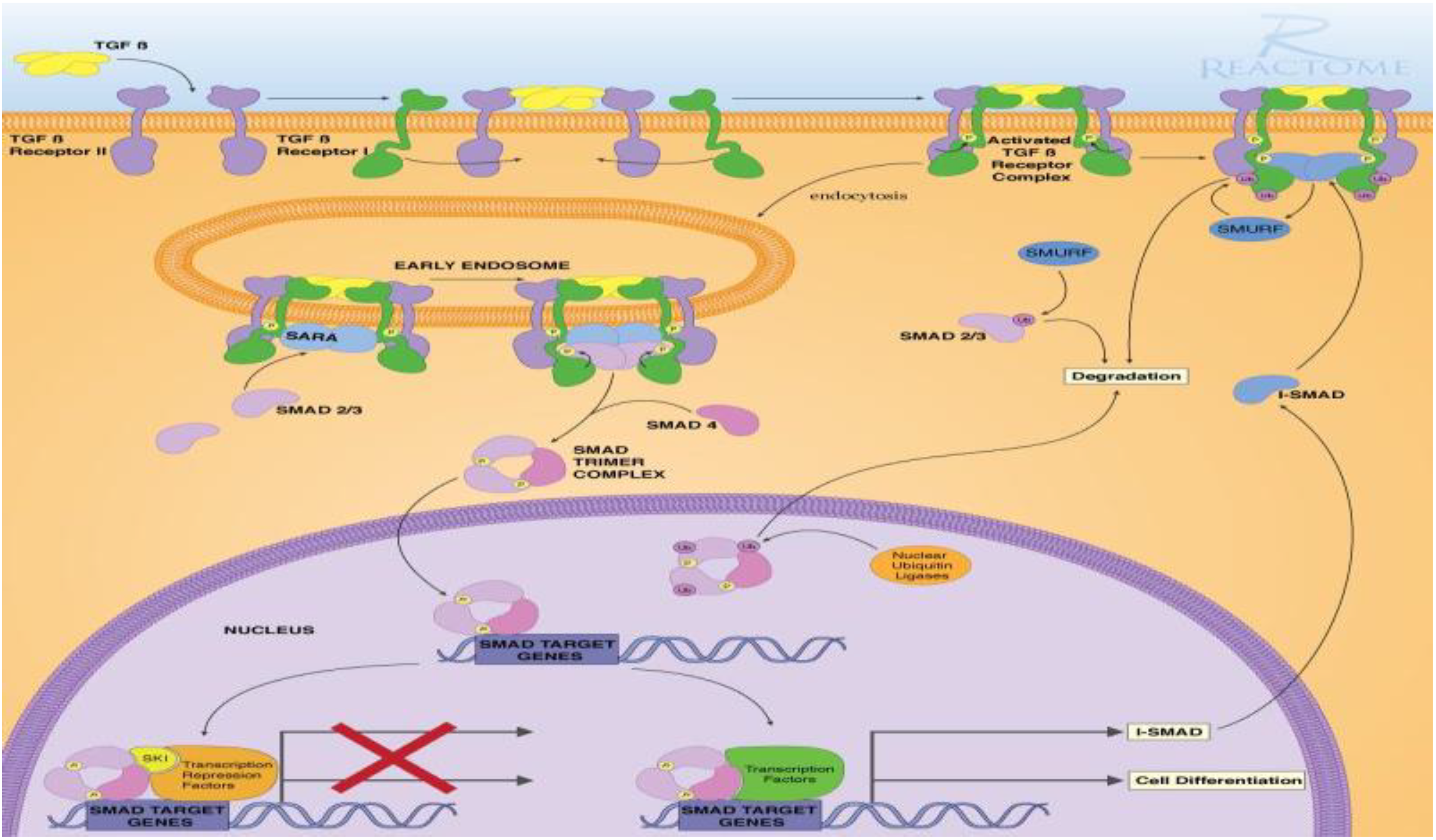
2.1. Signaling Pathway Enrichment Analysis
2.2. Animals
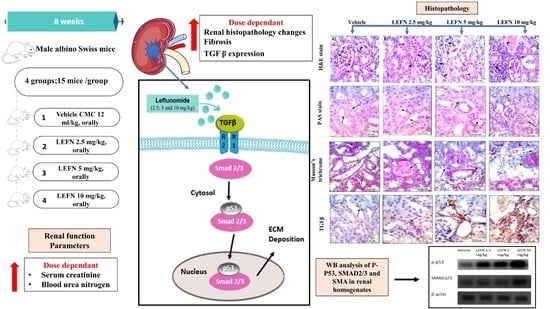
2.3. Design of the Experiment
2.4. Collection of Blood Samples and Kidneys
2.5. Estimation of Renal Function Parameters
2.6. Western Blot Analysis for p-p53 and SMAD2/3 in the Renal Homogenate
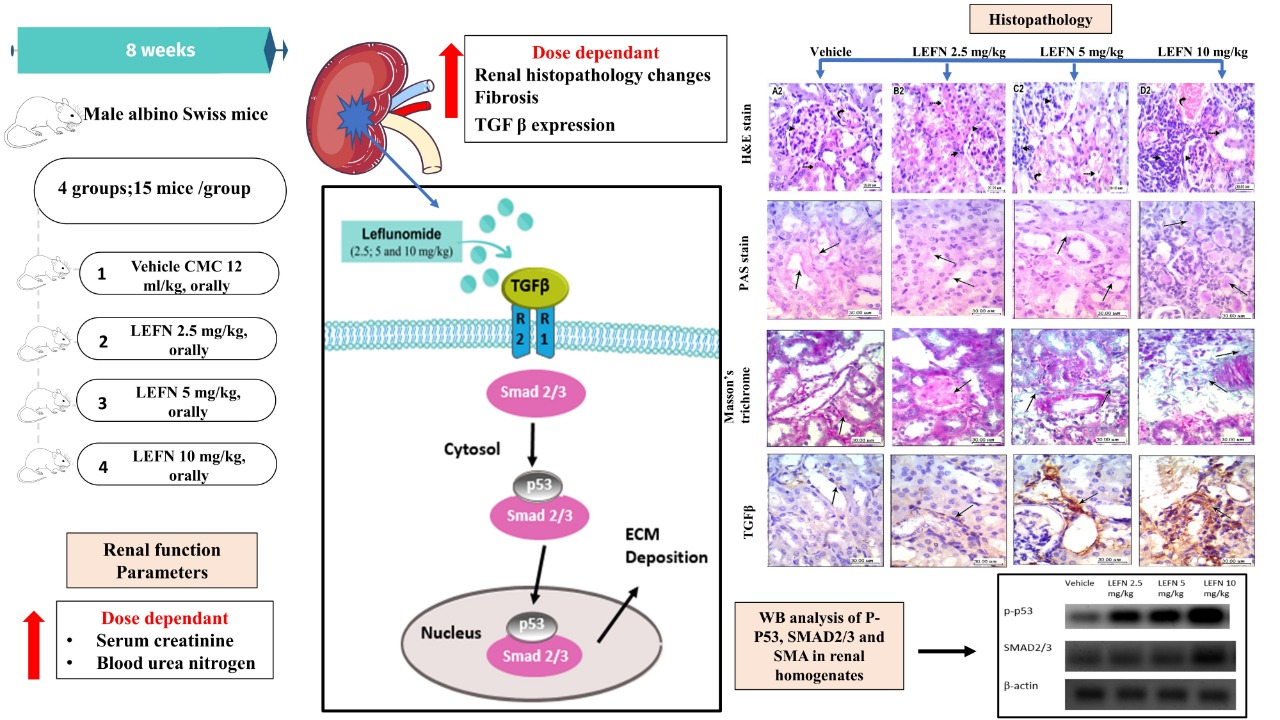
2.7. Kidney Histopathology and Immunohistochemistry
2.7.1. Morphometric Analysis of H&E-Stained Sections
2.7.2. Examination of PAS-Stained Sections
2.8. Immunohistochemistry and Image Analysis of TGFβ and p53 in Renal Tissues
2.9. Statistical Analysis
3. Results
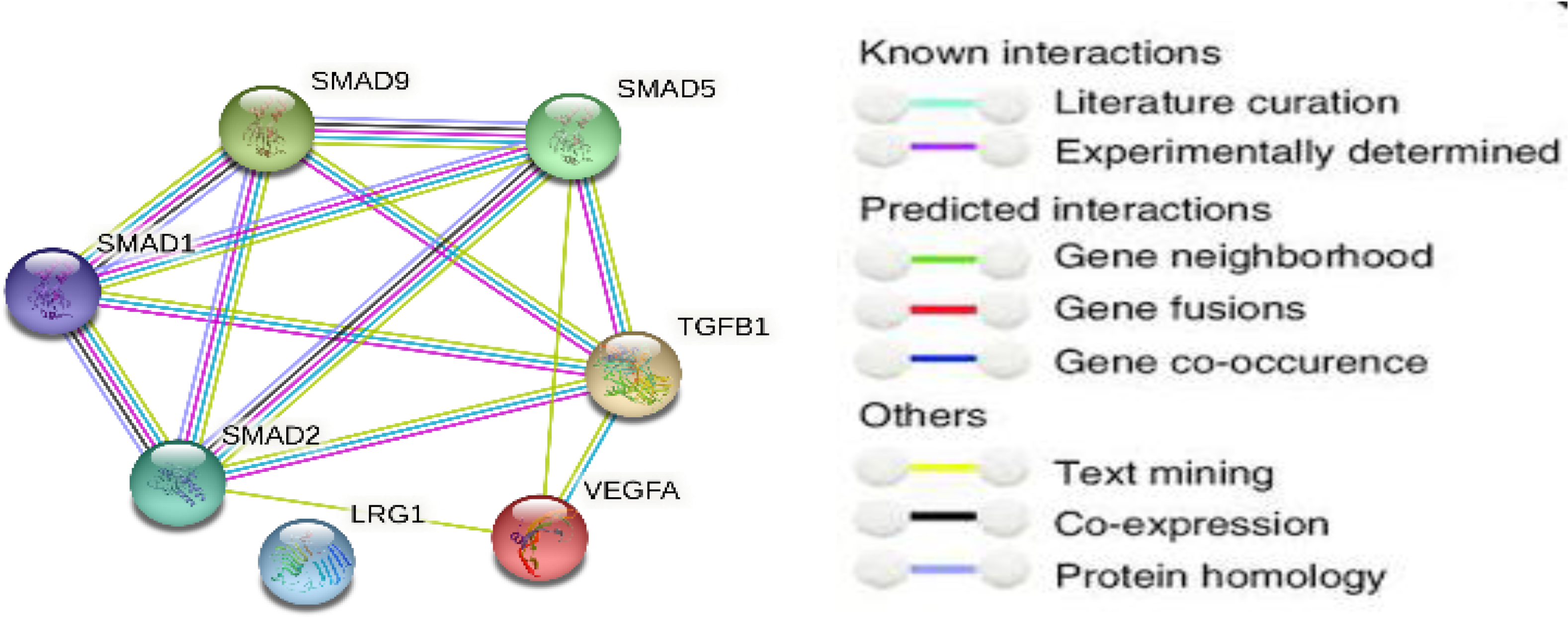
3.1. The Bioinformatic Study
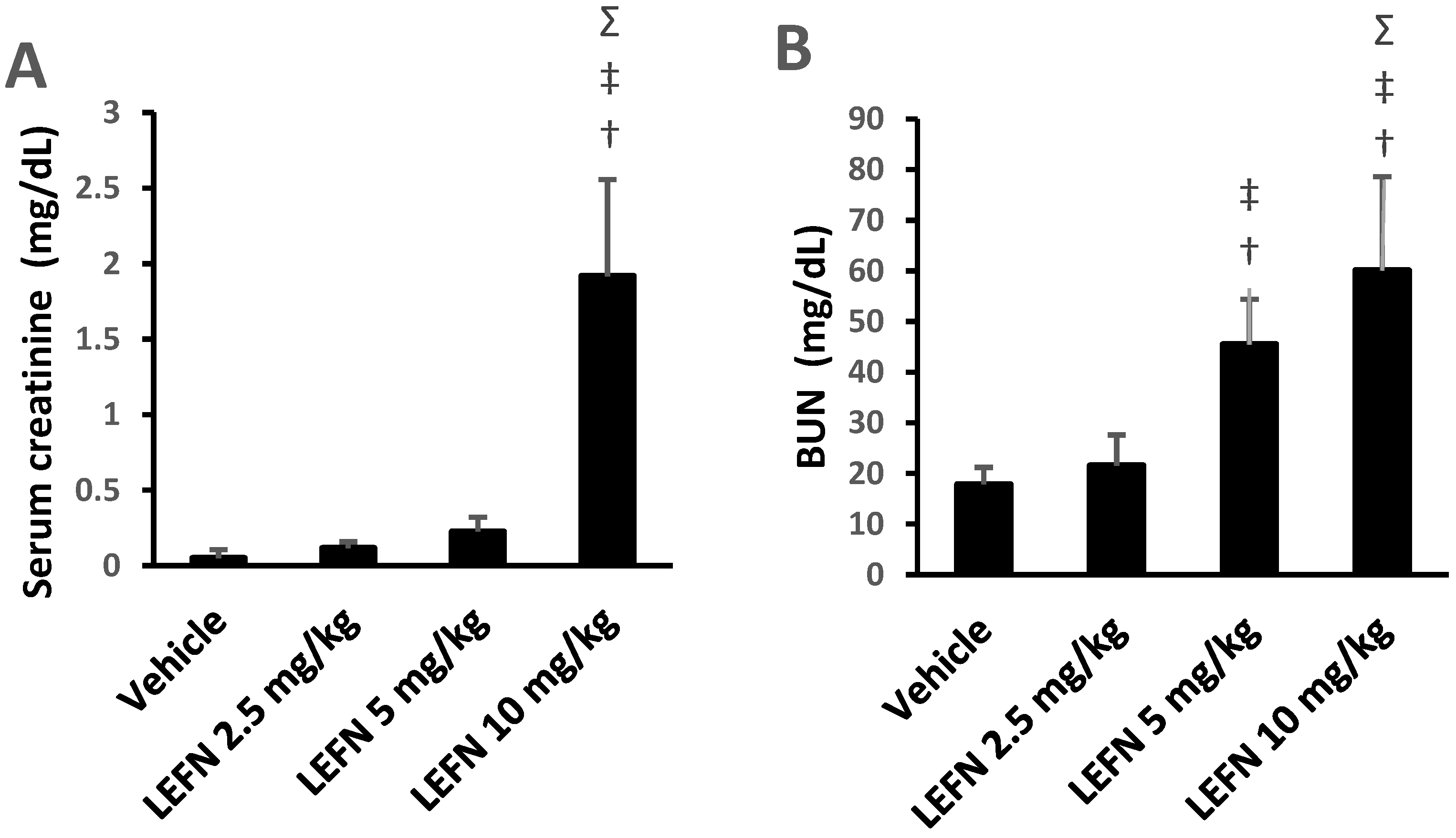
3.2. Assessment of Renal Function
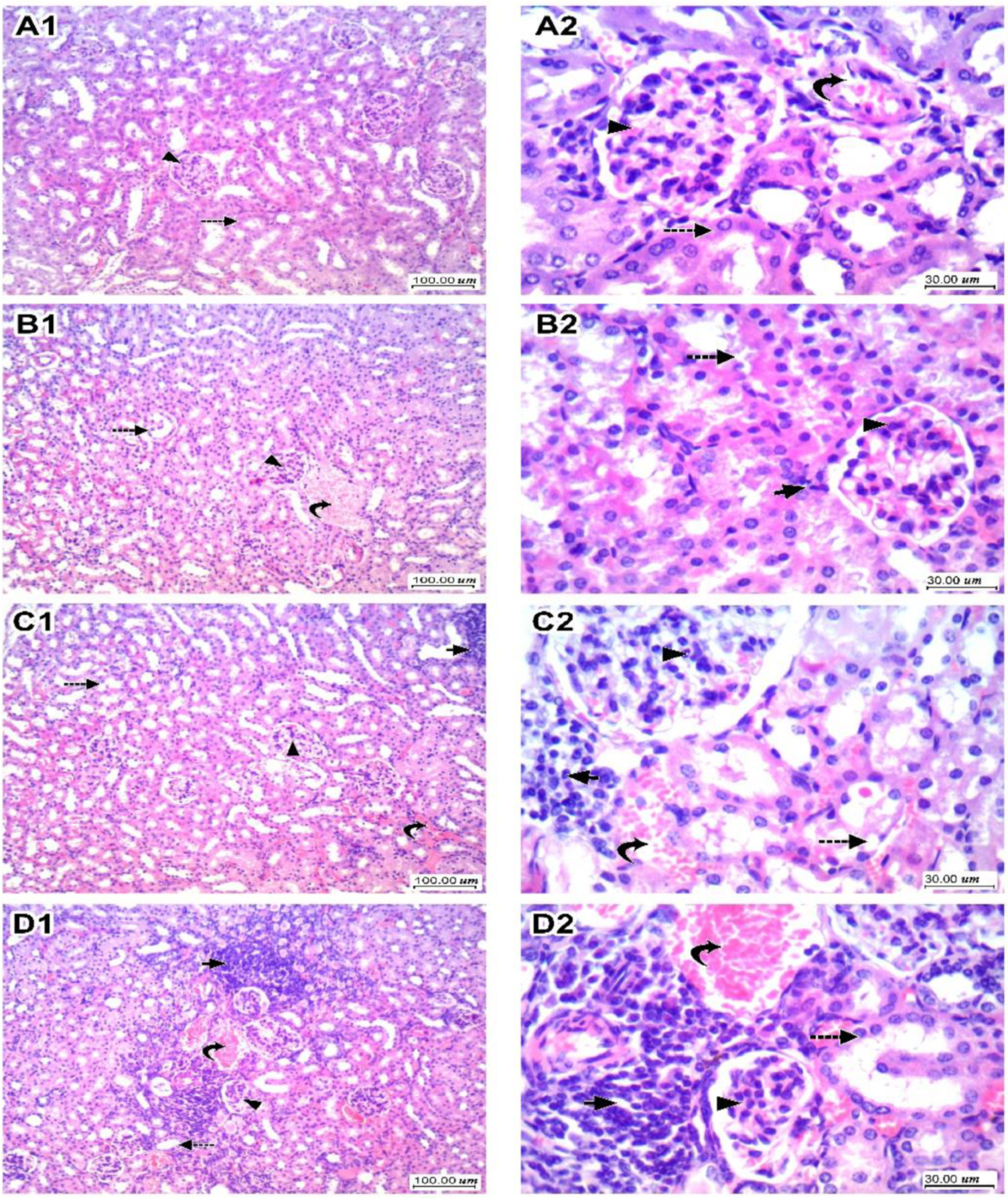
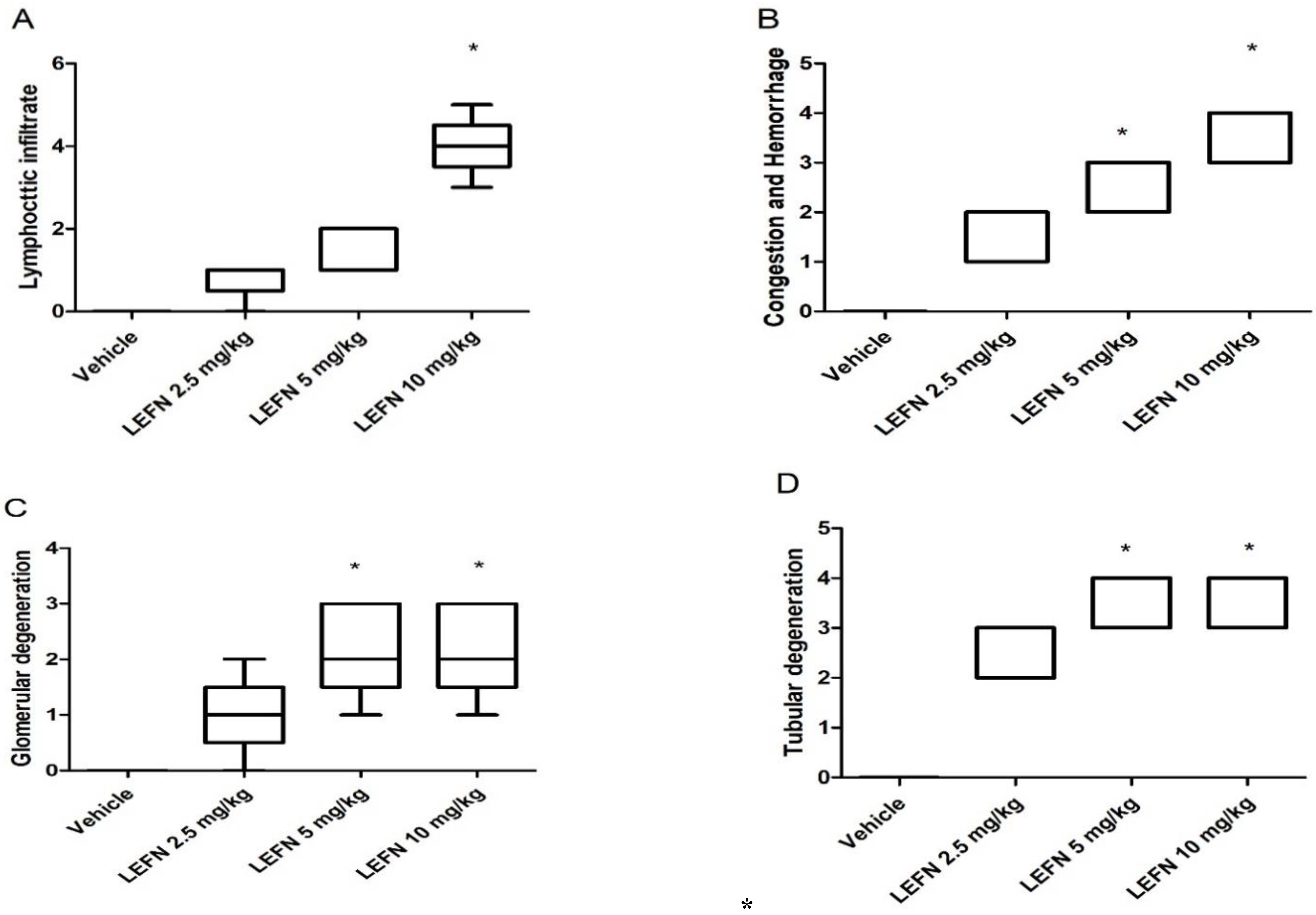
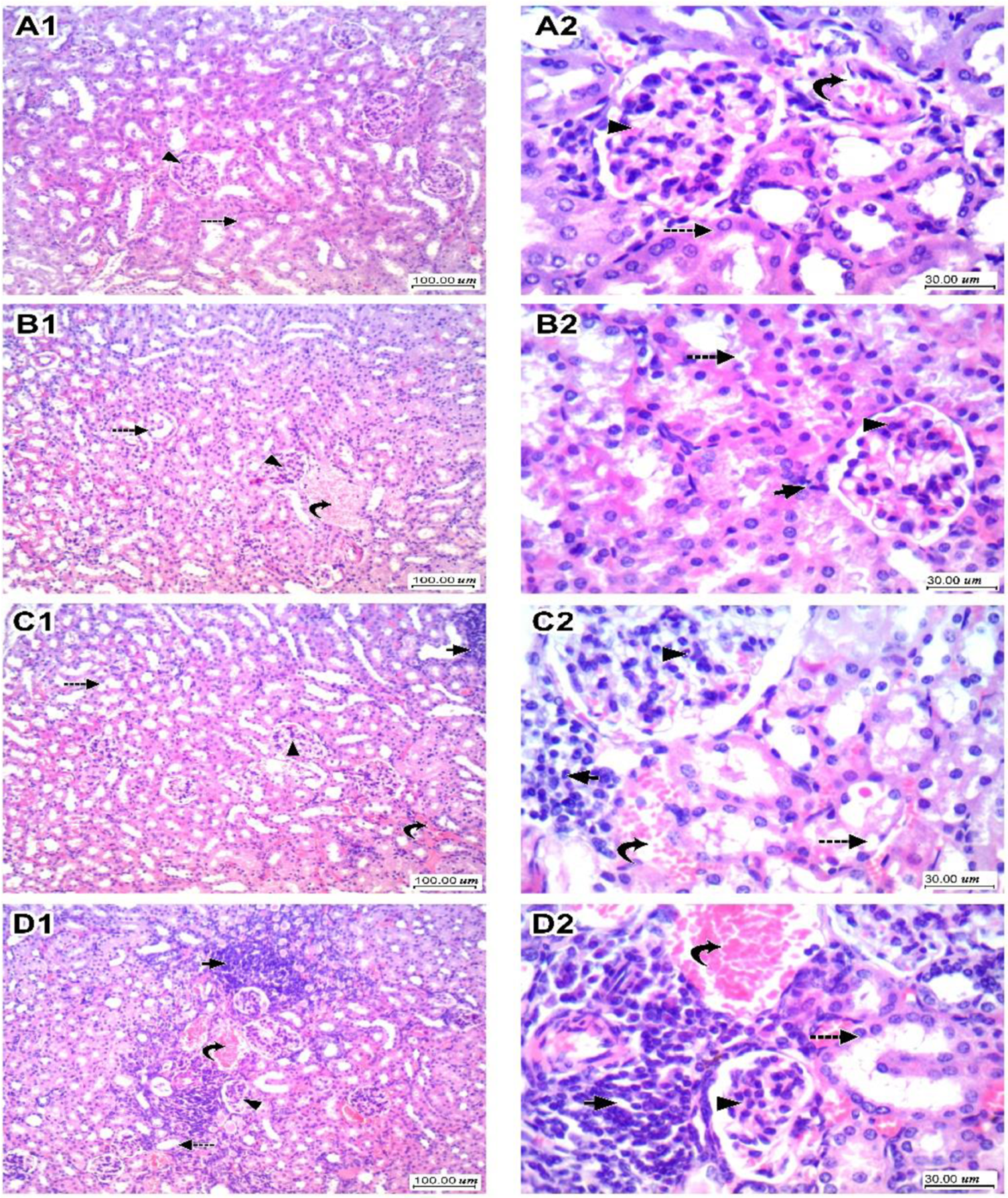
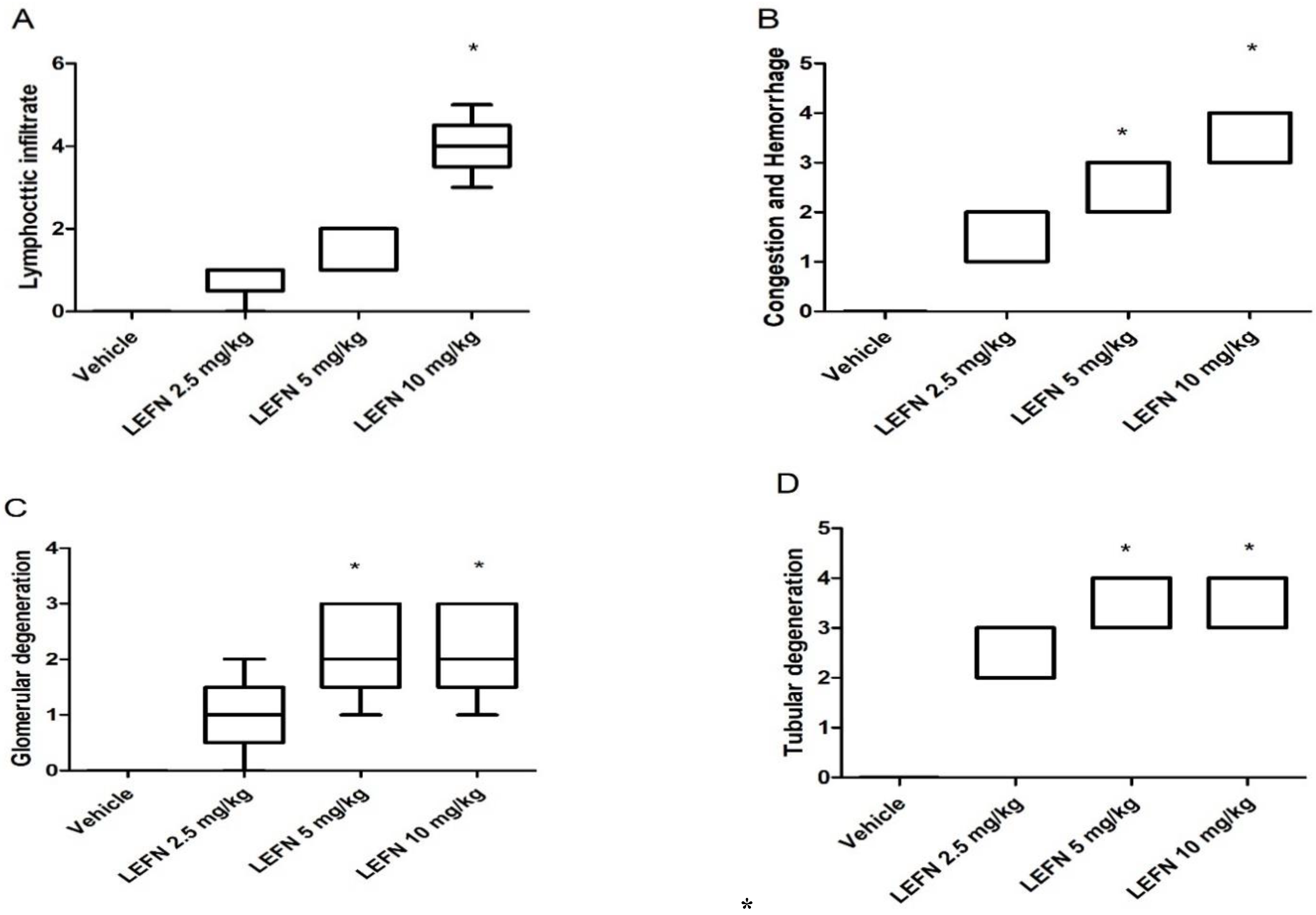
3.3. Kidney Histopathology for H&E-Stained Sections and Scoring
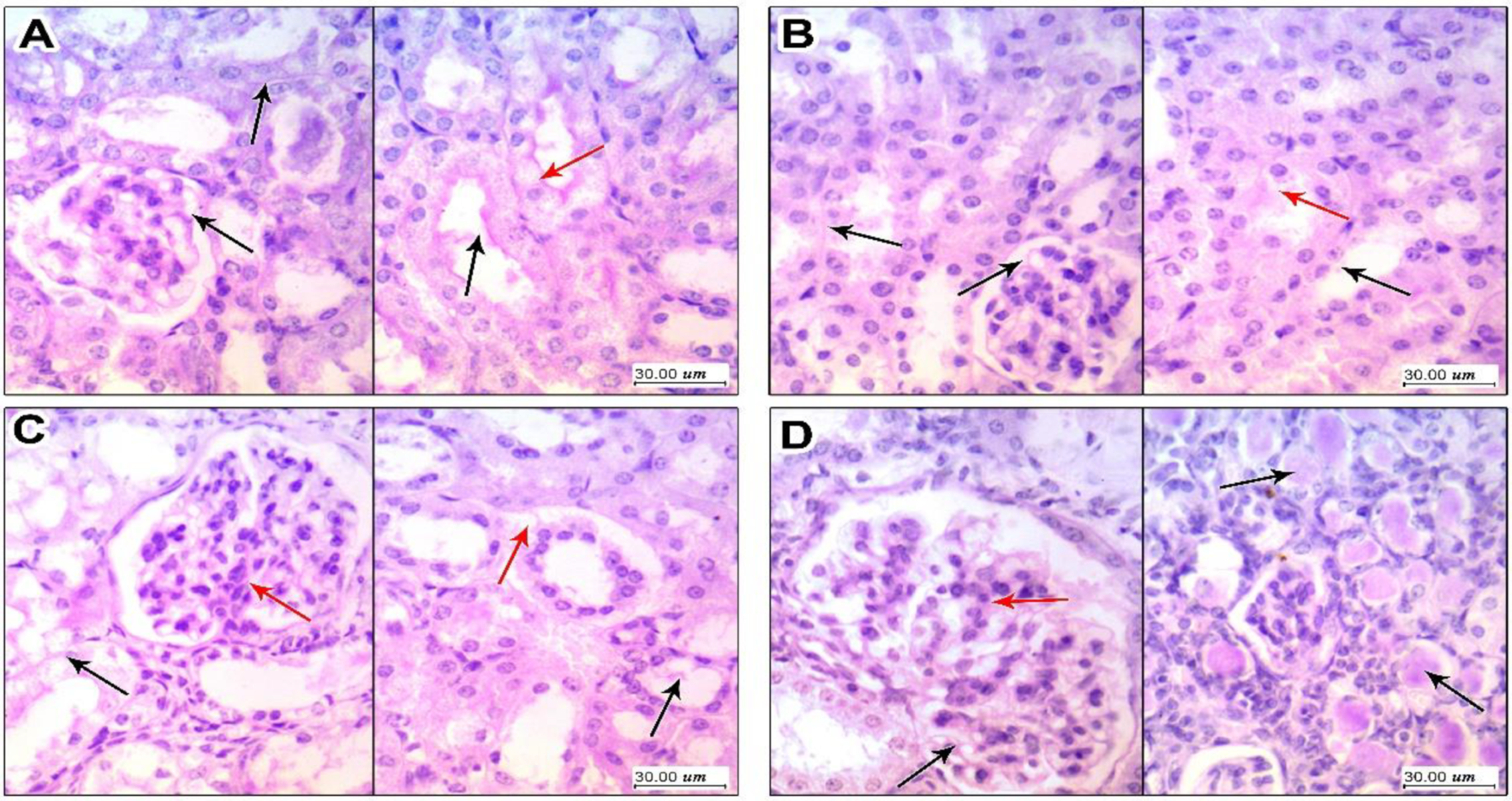
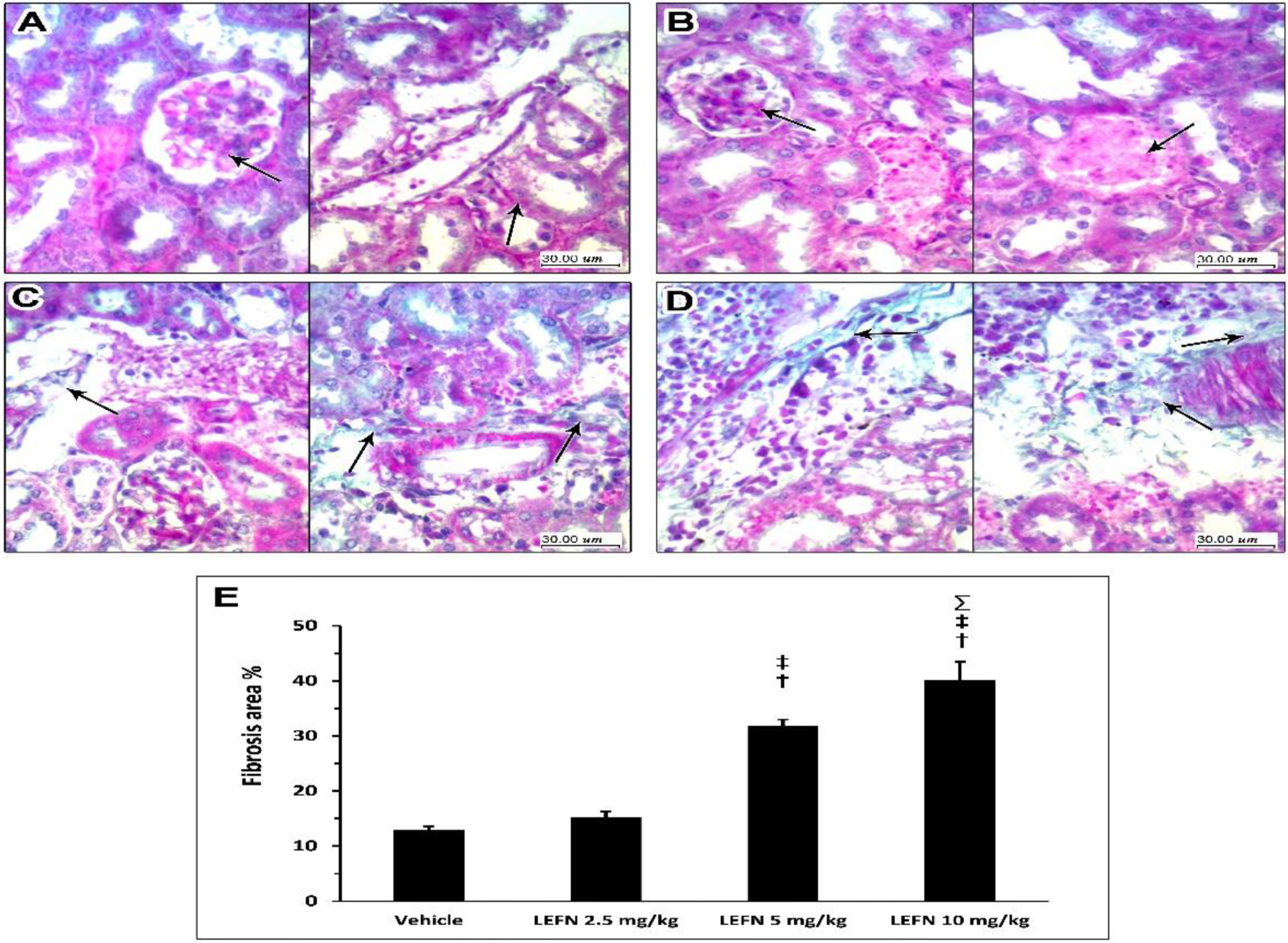
3.4. Kidney Histopathology in PAS Stained Sections
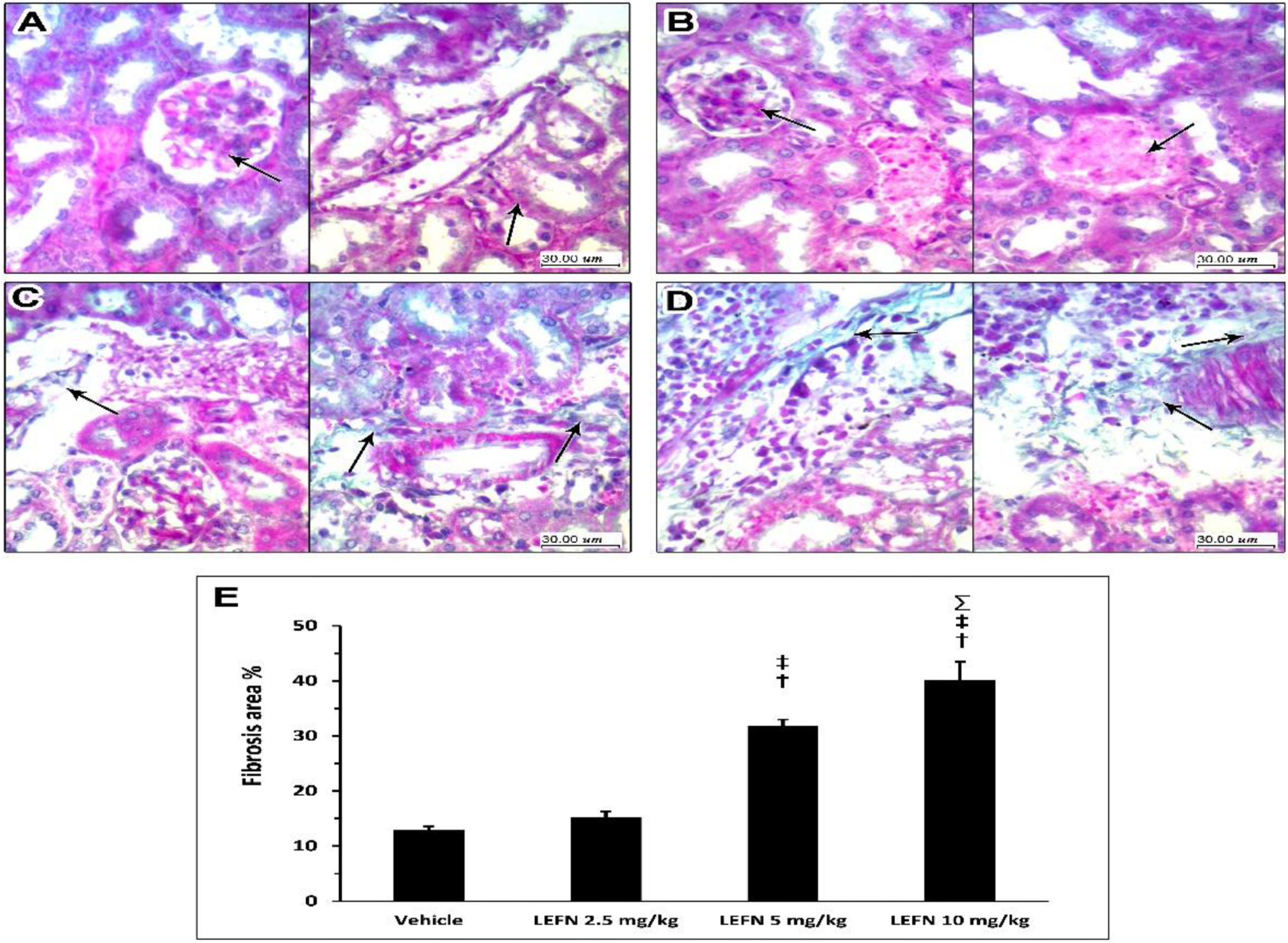
3.5. Kidney Histopathology in Masson’s Trichrome Stained Sections
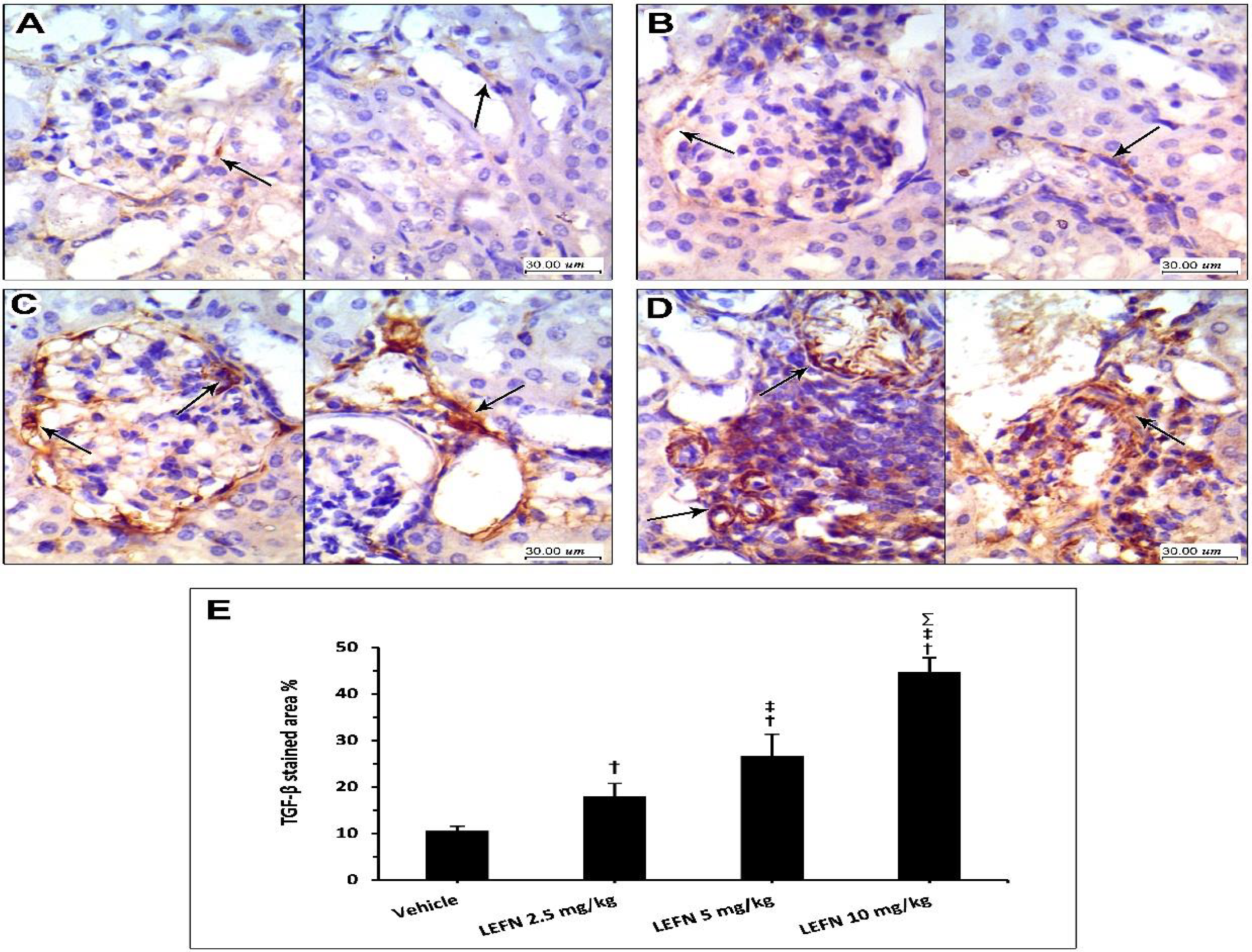
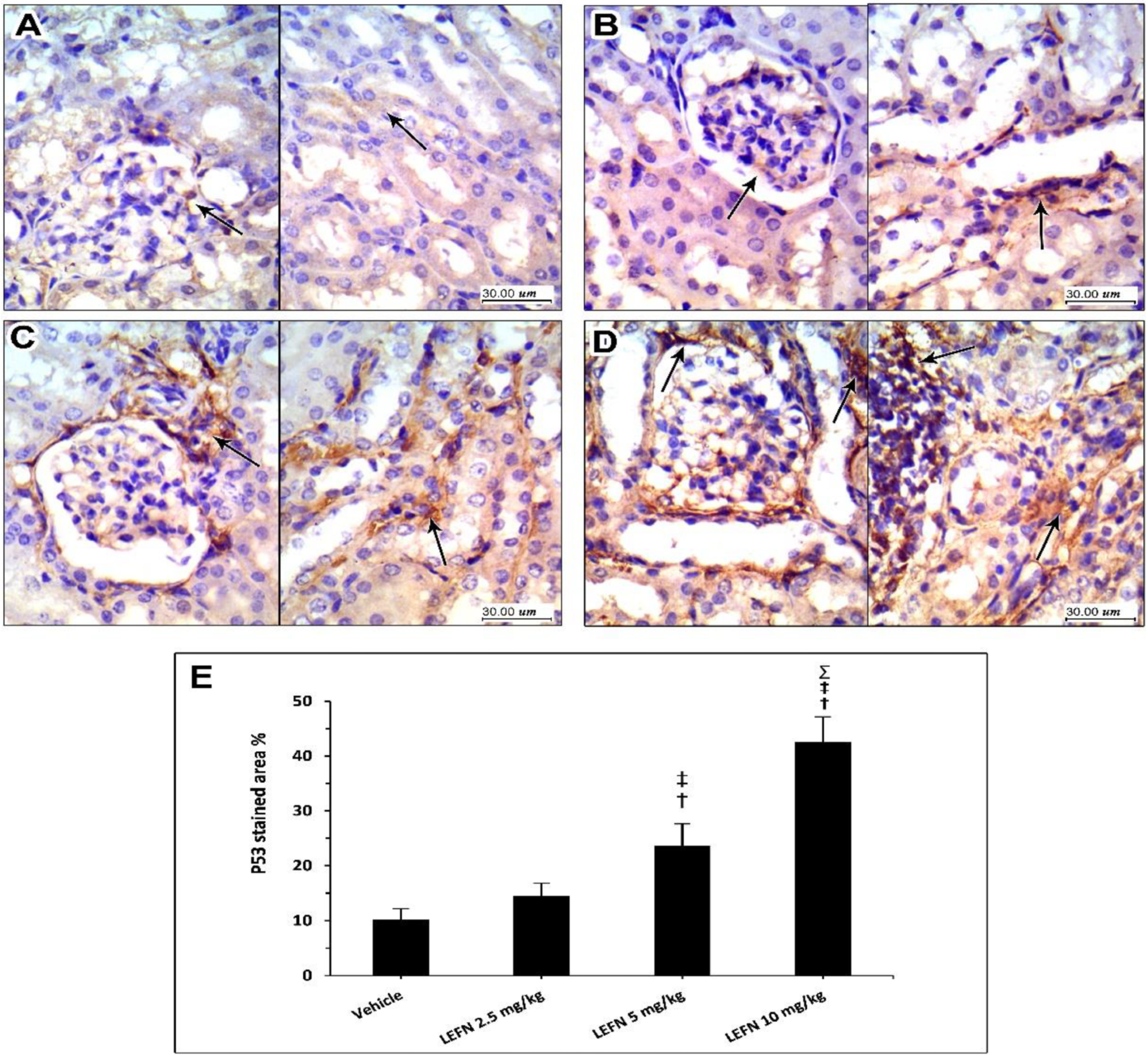
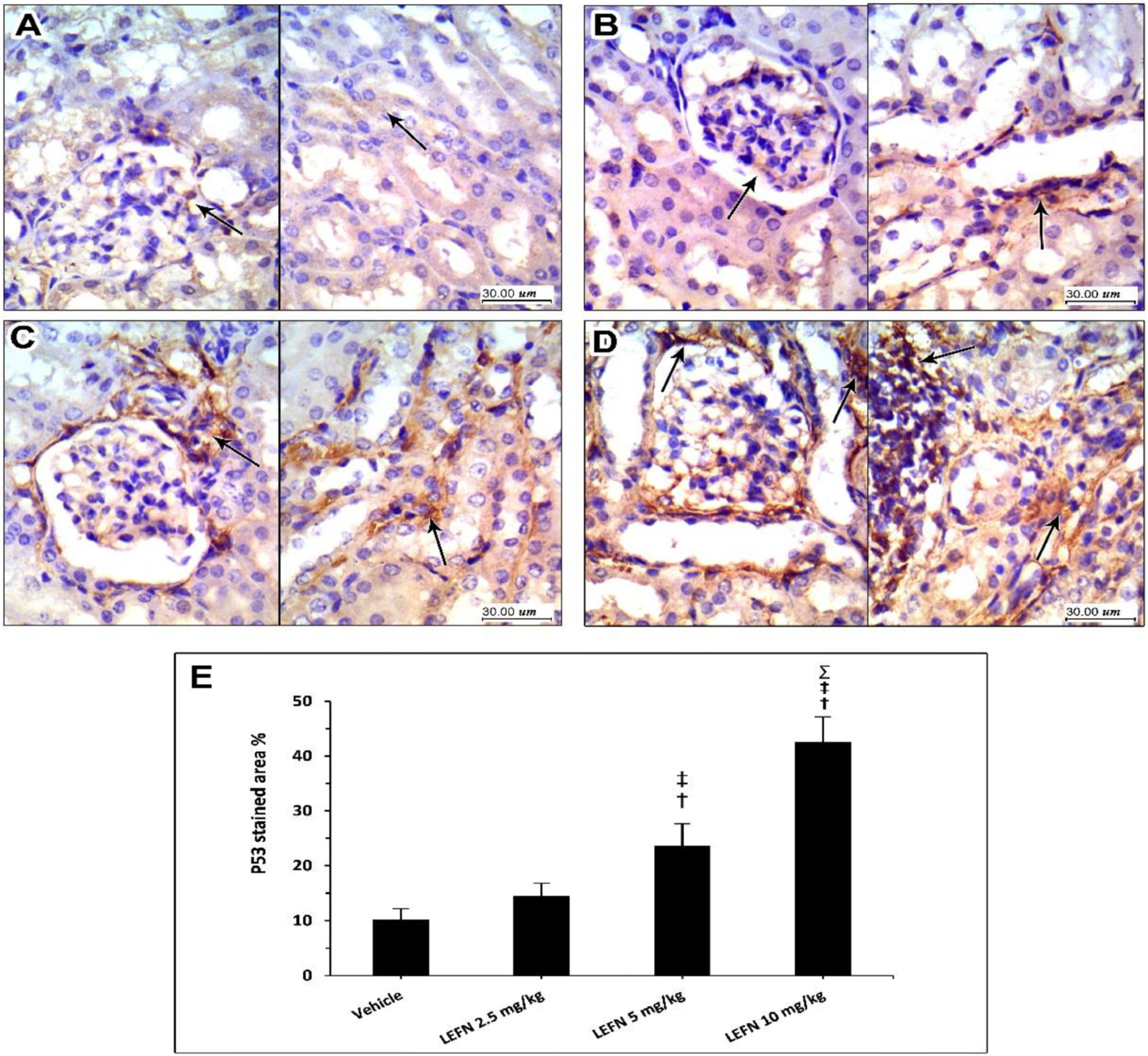
3.6. Immunohistochemical and Image Analysis of TGFβ and p53 Expression in Renal Specimen
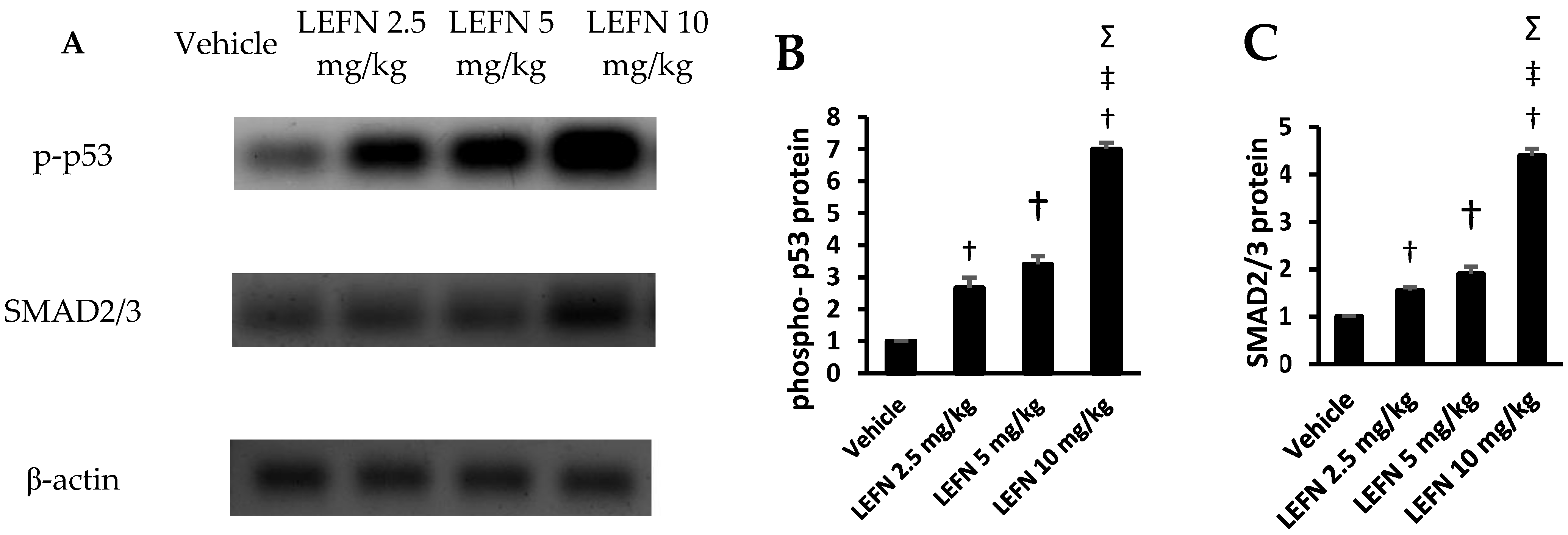
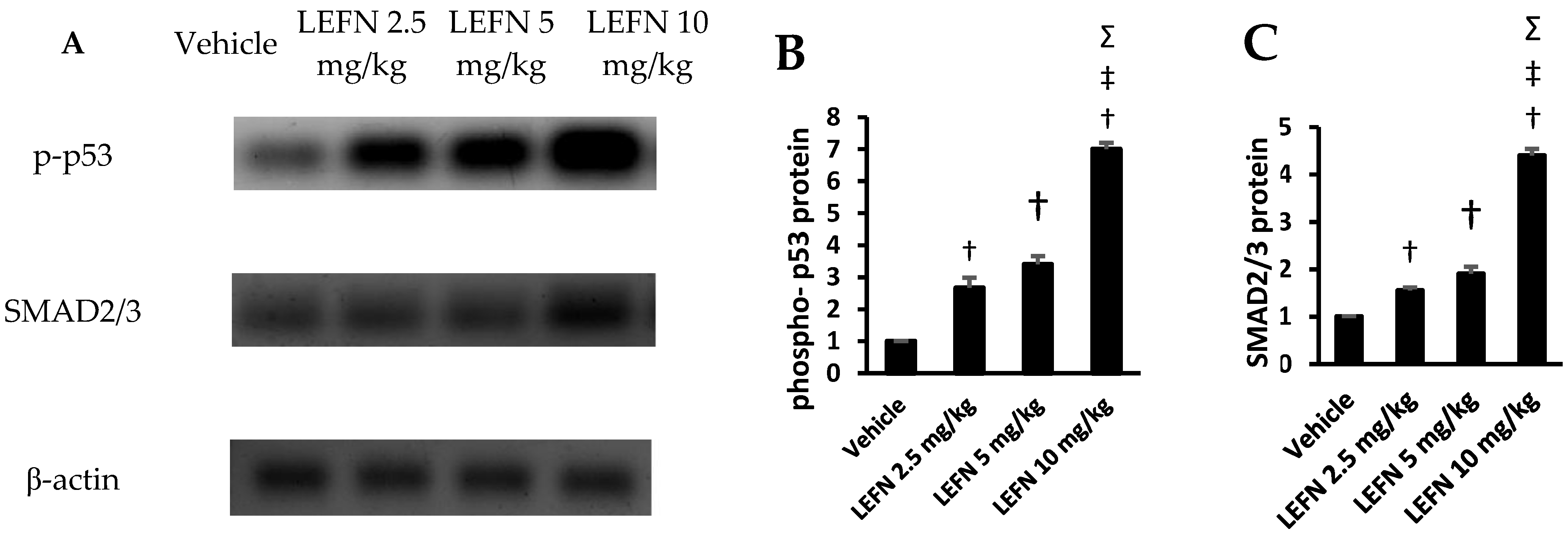
3.7. Western Blot Analysis of p-p53 and SMAD2/3 in Renal Homogenates
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alldred, A.; Emery, P. Leflunomide: A Novel DMARD for the Treatment of Rheumatoid Arthritis. Expert Opin. Pharmacother. 2001, 2, 125–137. [Google Scholar] [CrossRef]
- Kaplan, M.J. Leflunomide Aventis Pharma. Curr. Opin. Investig. Drugs 2001, 2, 222–230. [Google Scholar]
- Elshaer, R.E.; Tawfik, M.K.; Nosseir, N.; El-Ghaiesh, S.H.; Toraih, E.A.; Elsherbiny, N.M.; Zaitone, S.A. Leflunomide-Induced Liver Injury in Mice: Involvement of TLR4 Mediated Activation of PI3K/MTOR/NFκB Pathway. Life Sci. 2019, 235, 116824. [Google Scholar] [CrossRef]
- Raj, R.; Nugent, K. Leflunomide-Induced Interstitial Lung Disease (a Systematic Review). Sarcoidosis Vasc. Diffus. Lung Dis. 2013, 30, 167–176. [Google Scholar]
- Razak, S.A.; Islahudin, F.; Shamsuddin, A.F.; Shahril, N.S. A Study on Leflunomide-Induced Liver Injury in Rheumatoid Arthritis Patients. Res. J. Pharm. Technol. 2013, 6, 556–561. [Google Scholar]
- Shields, M.D.; Skelton, W.P.; Laber, D.A.; Verbosky, M.; Ashraf, N. A Novel Case of Leflunomide-Induced Thrombotic Thrombocytopenic Purpura. J. Hematol. 2021, 10, 139–142. [Google Scholar] [CrossRef]
- Villgran, V.D.; Rabold, E.; Nasrullah, A.; Disilvio, B.; Bajwa, O. Leflunomide-Induced Interstitial Pneumonitis: A Rare Occurrence in a Case without Underlying Lung Disease. Respir. Med. Case Rep. 2021, 33, 101468. [Google Scholar] [CrossRef]
- Xuan, J.; Ren, Z.; Qing, T.; Couch, L.; Shi, L.; Tolleson, W.H.; Guo, L. Mitochondrial Dysfunction Induced by Leflunomide and Its Active Metabolite. Toxicology 2018, 396–397, 33–45. [Google Scholar] [CrossRef] [Green Version]
- Lodhi, R.L.; Maity, S.; Kumar, P.; Saraf, S.A.; Kaithwas, G.; Saha, S. Evaluation of Mechanism of Hepatotoxicity of Leflunomide Using Albino Wistar Rats. Afr. J. Pharm. Pharmacol. 2013, 7, 1625–1631. [Google Scholar] [CrossRef] [Green Version]
- Quee, M.S.; Lee-Sun, N.; Eric, C.Y.C.; Urs, A.B. Oxidative Bioactivation and Toxicity of Leflunomide in Immortalized Human Hepatocytes and Kinetics of the Non-Enzymatic to Its Major Etabolitie, A77 1726. Drug Metab. Lett. 2008, 2, 153–157. [Google Scholar]
- Ren, Z.; Chen, S.; Qing, T.; Xuan, J.; Couch, L.; Yu, D.; Ning, B.; Shi, L.; Guo, L. Endoplasmic Reticulum Stress and MAPK Signaling Pathway Activation Underlie Leflunomide-Induced Toxicity in HepG2 Cells. Toxicology 2017, 392, 11–21. [Google Scholar] [CrossRef] [Green Version]
- El-Sherbiny, M.; Atef, H.; Eladl, M.A.; Mohamed, A.S.; El-Shafey, M.; Ali, H.S.; Zaitone, S.A.; Alomar, S.Y.; Alqahtani, S.A.M.; Aloyouni, S.Y.; et al. Leflunomide Induces Dose-Dependent Lung Injury in Mice via Stimulating Vimentin and NLRP3 Inflammasome Production. Front. Pharmacol. 2021, 12, 649. [Google Scholar] [CrossRef]
- Nakafero, G.; Grainge, M.J.; Card, T.; Mallen, C.D.; Zhang, W.; Doherty, M.; Taal, M.W.; Aithal, G.P.; Abhishek, A. What Is the Incidence of Methotrexate or Leflunomide Discontinuation Related to Cytopenia, Liver Enzyme Elevation or Kidney Function Decline? Rheumatology 2021, 60, 5785–5794. [Google Scholar] [CrossRef]
- Evans, S.J.; Webb, D.; Lawson, T.M.; Siebert, S. Renal Tubular Acidosis Associated with Leflunomide. Rheumatology 2007, 46, 1040–1041. [Google Scholar] [CrossRef] [Green Version]
- Haydar, A.A.; Hujairi, N.; Kirkham, B.; Hangartner, R.; Goldsmith, D.J. Chronic Overdose of Leflunomide Inducing Interstitial Nephritis. Nephrol. Dial. Transplant. 2004, 19, 1334–1335. [Google Scholar] [CrossRef]
- Hurtado, M.; Silva, O.R.B.; Santacruz, F. Nefritis túbulo intersticial por leflunomida. Rev. Colomb. Nefrol. 2016, 3, 132–136. [Google Scholar] [CrossRef]
- Pinto, B.; Dhir, V.; Krishnan, S.; Nada, R. Leflunomide-Induced DRESS Syndrome with Renal Involvement and Vasculitis. Clin. Rheumatol. 2013, 32, 689–693. [Google Scholar] [CrossRef]
- Azab, A.; Fetouh, F.; Albasha, M. Nephro-Protective Effects of Curcumin, Rosemary and Propolis against Gentamicin Induced Toxicity in Guinea Pigs: Morphological and Biochemical Study. Am. J. Clin. Exp. Med. 2014, 2, 28–35. [Google Scholar] [CrossRef] [Green Version]
- Balas, M.; Popescu Din, I.M.; Hermenean, A.; Cinteza, L.O.; Dinischiotu, A. Exposure to Iron Oxide Nanoparticles Coated with Phospholipid-Based Polymeric Micelles Induces Renal Transitory Biochemical and Histopathological Changes in Mice. Materials 2021, 14, 2605. [Google Scholar] [CrossRef]
- Haschek, W.M.; Rousseaux, C.G.; Wallig, M.A.; Bolon, B.; Ochoa, R. Haschek and Rousseaux’s Handbook of Toxicologic Pathology; Academic Press: Cambridge, MA, USA, 2013. [Google Scholar]
- Balaji, K.; Vijayakumar, J.; Sankaran, P.K.; Senthilkumar, S.; Vijayaraghavan, R.; Selvaraj, J.; Yuvaraj, M.F. Molecular Studies on the Nephroprotective Potential of Celastrus Paniculatus against Lead-Acetate-Induced Nephrotoxicity in Experimental Rats: Role of the PI3K/AKT Signaling Pathway. Molecules 2021, 26, 6647. [Google Scholar] [CrossRef]
- Min, H.S.; Lee, J.E.; Ghee, J.Y.; Kang, Y.S.; Cha, J.J.; Han, J.Y.; Han, S.Y.; Cha, D.R. A Novel Dipeptidyl Peptidase-4 Inhibitor DA-1229 Ameliorates Tubulointerstitial Fibrosis in Cyclosporine Nephrotoxicity in Mice. Life 2021, 11, 251. [Google Scholar] [CrossRef]
- Bushau-Sprinkle, A.M.; Barati, M.T.; Zheng, Y.; Watson, W.H.; Gagnon, K.B.; Khundmiri, S.J.; Kitterman, K.T.; Clark, B.J.; Siskind, L.J.; Doll, M.A.; et al. Na/H Exchange Regulatory Factor 1 Deficient Mice Show Evidence of Oxidative Stress and Altered Cisplatin Pharmacokinetics. Antioxidants 2021, 10, 1036. [Google Scholar] [CrossRef]
- Perazella, M.A. Pharmacology behind Common Drug Nephrotoxicities. Clin. J. Am. Soc. Nephrol. 2018, 13, 1897–1908. [Google Scholar] [CrossRef]
- Roberts, A.B.; Wakefield, L.M. The Two Faces of Transforming Growth Factor β in Carcinogenesis. Proc. Natl. Acad. Sci. USA 2003, 100, 8621–8623. [Google Scholar] [CrossRef] [Green Version]
- Meulmeester, E.; ten Dijke, P. The Dynamic Roles of TGF-β in Cancer. J. Pathol. 2011, 223, 206–219. [Google Scholar] [CrossRef]
- Inman, G.J. Switching TGFβ from a Tumor Suppressor to a Tumor Promoter. Curr. Opin. Genet. Dev. 2011, 21, 93–99. [Google Scholar] [CrossRef]
- Huang, T.; David, L.; Mendoza, V.; Yang, Y.; Villarreal, M.; De, K.; Sun, L.; Fang, X.; López-Casillas, F.; Wrana, J.L.; et al. TGF-β Signalling Is Mediated by Two Autonomously Functioning TβRI:TβRII Pairs. EMBO J. 2011, 30, 1263–1276. [Google Scholar] [CrossRef] [Green Version]
- Groppe, J.; Hinck, C.S.; Samavarchi-Tehrani, P.; Zubieta, C.; Schuermann, J.P.; Taylor, A.B.; Schwarz, P.M.; Wrana, J.L.; Hinck, A.P. Cooperative Assembly of TGF-β Superfamily Signaling Complexes Is Mediated by Two Disparate Mechanisms and Distinct Modes of Receptor Binding. Mol. Cell 2008, 29, 157–168. [Google Scholar] [CrossRef]
- Feng, X.-H.; Derynck, R. Specificity and versatility in tgf-β signaling through smads. Annu. Rev. Cell Dev. Biol. 2005, 21, 659–693. [Google Scholar] [CrossRef] [Green Version]
- Heldin, C.-H.; Moustakas, A. Role of Smads in TGFβ Signaling. Cell Tissue Res. 2012, 347, 21–36. [Google Scholar] [CrossRef]
- Massagué, J.; Seoane, J.; Wotton, D. Smad Transcription Factors. Genes Dev. 2005, 19, 2783–2810. [Google Scholar] [CrossRef] [Green Version]
- Ramesh, S.; Qi, X.-J.; Wildey, G.M.; Robinson, J.; Molkentin, J.; Letterio, J.; Howe, P.H. TGFβ-Mediated BIM Expression and Apoptosis Are Regulated through SMAD3-Dependent Expression of the MAPK Phosphatase MKP2. EMBO Rep. 2008, 9, 990–997. [Google Scholar] [CrossRef] [Green Version]
- Spender, L.C.; O’Brien, D.I.; Simpson, D.; Dutt, D.; Gregory, C.D.; Allday, M.J.; Clark, L.J.; Inman, G.J. TGF-β Induces Apoptosis in Human B Cells by Transcriptional Regulation of BIK and BCL-XL. Cell Death Differ. 2009, 16, 593–602. [Google Scholar] [CrossRef] [Green Version]
- Meng, X.; Nikolic-Paterson, D.J.; Lan, H.Y. TGF-β: The Master Regulator of Fibrosis. Nat. Rev. Nephrol. 2016, 12, 325–338. [Google Scholar] [CrossRef]
- Sharma, K.; Ziyadeh, F.N.; Alzahabi, B.; McGowan, T.A.; Kapoor, S.; Kurnik, B.R.C.; Kurnik, P.B.; Weisberg, L.S. Increased Renal Production of Transforming Growth Factor-Β1 in Patients with Type II Diabetes. Diabetes 1997, 46, 854–859. [Google Scholar] [CrossRef]
- Beckerman, R.; Prives, C. Transcriptional Regulation by P53. Cold Spring Harb. Perspect. Biol. 2010, 2, a000935. [Google Scholar] [CrossRef] [Green Version]
- Riley, T.; Sontag, E.; Chen, P.; Levine, A. Transcriptional Control of Human P53-Regulated Genes. Nat. Rev. Mol. Cell Biol. 2008, 9, 402–412. [Google Scholar] [CrossRef]
- Soto-Cruz, I.; Legorreta-Herrera, M. Analysis of a P53 Mutation Associated with Cancer Susceptibility for Biochemistry and Genetic Laboratory Courses. Biochem. Mol. Biol. Educ. 2009, 37, 236–242. [Google Scholar] [CrossRef]
- Levine, A.J. Reviewing the Future of the P53 Field. Cell Death Differ. 2018, 25, 1–2. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, M.S.; Stojanov, P.; Mermel, C.H.; Robinson, J.T.; Garraway, L.A.; Golub, T.R.; Meyerson, M.; Gabriel, S.B.; Lander, E.S.; Getz, G. Discovery and Saturation Analysis of Cancer Genes across 21 Tumour Types. Nature 2014, 505, 495–501. [Google Scholar] [CrossRef] [Green Version]
- Levine, A.J.; Oren, M. The First 30 Years of P53: Growing Ever More Complex. Nat. Rev. Cancer 2009, 9, 749–758. [Google Scholar] [CrossRef] [Green Version]
- Vogelstein, B.; Lane, D.; Levine, A.J. Surfing the P53 Network. Nature 2000, 408, 307–310. [Google Scholar] [CrossRef]
- Elston, R.; Inman, G.J. Crosstalk between P53 and TGF-β Signalling. J. Signal Transduct. 2012, 2012, e294097. [Google Scholar] [CrossRef] [Green Version]
- Wyllie, F.S.; Dawson, T.; Bond, J.A.; Goretzki, P.; Game, S.; Prime, S.; Wynford-Thomas, D. Correlated Abnormalities of Transforming Growth Factor-Β1 Response and P53 Expression in Thyroid Epithelial Cell Transformation. Mol. Cell. Endocrinol. 1991, 76, 13–21. [Google Scholar] [CrossRef]
- Higgins, S.P.; Tang, Y.; Higgins, C.E.; Mian, B.; Zhang, W.; Czekay, R.-P.; Samarakoon, R.; Conti, D.J.; Higgins, P.J. TGF-Β1/P53 Signaling in Renal Fibrogenesis. Cell. Signal. 2018, 43, 1–10. [Google Scholar] [CrossRef]
- KEGG: Kyoto Encyclopedia of Genes and Genomes. Available online: https://www.genome.jp/kegg/ (accessed on 12 May 2022).
- Home-Reactome Pathway Database. Available online: https://reactome.org/ (accessed on 12 May 2022).
- Xu, Q.; Ming, Z.; Dart, A.M.; Du, X.-J. Optimizing Dosage of Ketamine and Xylazine in Murine Echocardiography. Clin. Exp. Pharmacol. Physiol. 2007, 34, 499–507. [Google Scholar] [CrossRef]
- Gargiulo, S.; Greco, A.; Gramanzini, M.; Esposito, S.; Affuso, A.; Brunetti, A.; Vesce, G. Mice Anesthesia, Analgesia, and Care, Part I: Anesthetic Considerations in Preclinical Research. ILAR J. 2012, 53, E55–E69. [Google Scholar] [CrossRef] [Green Version]
- Bancroft, J.D.; Gamble, M. Theory and Practice of Histological Techniques; Elsevier: Amsterdam, The Netherlands, 2008. [Google Scholar]
- Klopfleisch, R. Multiparametric and Semiquantitative Scoring Systems for the Evaluation of Mouse Model Histopathology—A Systematic Review. BMC Vet. Res. 2013, 9, 123. [Google Scholar] [CrossRef] [Green Version]
- Asa, S.A.; EL-Nahass, E.-S.; Moselhy, W.A.; Hassan, N.E.-H.Y.; Hassan, A.A. Aflatoxin Induced Renal Toxicity in Albino Rats and the Ameliorative Effect of Green Tea Aqueous Extract: Histological, Morphometric and Immuno-Histochemical Study. J. Exp. Appl. Anim. Sci. 2018, 2, 273–285. [Google Scholar] [CrossRef]
- Cekmen, M.; Otunctemur, A.; Ozbek, E.; Cakir, S.S.; Dursun, M.; Polat, E.C.; Somay, A.; Ozbay, N. Pomegranate Extract Attenuates Gentamicin-Induced Nephrotoxicity in Rats by Reducing Oxidative Stress. Ren. Fail. 2013, 35, 268–274. [Google Scholar] [CrossRef] [Green Version]
- Breedveld, F.C.; Dayer, J.-M. Leflunomide: Mode of Action in the Treatment of Rheumatoid Arthritis. Ann. Rheum. Dis. 2000, 59, 841–849. [Google Scholar] [CrossRef]
- Van Riel, P.L.C.M.; Smolen, J.S.; Emery, P.; Kalden, J.R.; Dougados, M.; Strand, C.V.; Breedveld, F.C. Leflunomide: A Manageable Safety Profile. J. Rheumatol. Suppl. 2004, 71, 21–24. [Google Scholar]
- Toyokawa, Y.; Kingetsu, I.; Yasuda, C.; Yasuda, J.; Yoshida, K.; Kurosaka, D.; Yamada, A. Pancytopenia, Including Macrocytic Anemia, Associated with Leflunomide in a Rheumatoid Arthritis Patient. Mod. Rheumatol. 2007, 17, 436–440. [Google Scholar] [CrossRef]
- Kemp, E.; Nielsen, H.; Petersen, L.J.; Gam, A.N.; Dahlager, J.; Horn, T.; Larsen, S.; Olsen, S. Newer Immunomodulating Drugs in Rheumatoid Arthritis May Precipitate Glomerulonephritis. Clin. Nephrol. 2001, 55, 87–88. [Google Scholar]
- Pliquett, R.U.; Lübbert, C.; Schäfer, C.; Girndt, M. Thrombotic Microangiopathy and Liver Toxicity Due to a Combination Therapy of Leflunomide and Methotrexate: A Case Report. J. Med. Case Rep. 2020, 14, 26. [Google Scholar] [CrossRef]
- Rozman, B. Clinical Pharmacokinetics of Leflunomide. Clin. Pharmacokinet. 2002, 41, 421–430. [Google Scholar] [CrossRef]
- Boelsterli, U.A.; Lim, P.L.K. Mitochondrial Abnormalities—A Link to Idiosyncratic Drug Hepatotoxicity? Toxicol. Appl. Pharmacol. 2007, 220, 92–107. [Google Scholar] [CrossRef]
- Suissa, S.; Ernst, P.; Hudson, M.; Bitton, A.; Kezouh, A. Newer Disease-Modifying Antirheumatic Drugs and the Risk of Serious Hepatic Adverse Events in Patients with Rheumatoid Arthritis. Am. J. Med. 2004, 117, 87–92. [Google Scholar] [CrossRef]
- Van Roon, E.N.; Jansen, T.L.T.A.; Houtman, N.M.; Spoelstra, P.; Brouwers, J.R. Leflunomide for the Treatment of Rheumatoid Arthritis in Clinical Practice. Drug Saf. 2004, 27, 345–352. [Google Scholar] [CrossRef]
- Weinblatt, M.E.; Dixon, J.A.; Falchuk, K.R. Serious Liver Disease in a Patient Receiving Methotrexate and Leflunomide. Arthritis Rheum. 2000, 43, 2609–2611. [Google Scholar] [CrossRef] [Green Version]
- Fabregat, I.; Moreno-Càceres, J.; Sánchez, A.; Dooley, S.; Dewidar, B.; Giannelli, G.; ten Dijke, P.; on behalf of the IT-LIVER Consortium. TGF-β Signalling and Liver Disease. FEBS J. 2016, 283, 2219–2232. [Google Scholar] [CrossRef] [Green Version]
- Tsuchida, T.; Friedman, S.L. Mechanisms of Hepatic Stellate Cell Activation. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 397–411. [Google Scholar] [CrossRef]
- Lee, S.-W.; Park, H.-J.; Kim, B.K.; Han, K.-H.; Lee, S.-K.; Kim, S.U.; Park, Y.-B. Leflunomide Increases the Risk of Silent Liver Fibrosis in Patients with Rheumatoid Arthritis Receiving Methotrexate. Arthritis Res. Ther. 2012, 14, R232. [Google Scholar] [CrossRef] [Green Version]
- Chang, K.-W.; Zhang, X.; Lin, S.-C.; Lin, Y.-C.; Li, C.-H.; Akhrymuk, I.; Lin, S.-H.; Lin, C.-C. Atractylodin Suppresses TGF-β-Mediated Epithelial-Mesenchymal Transition in Alveolar Epithelial Cells and Attenuates Bleomycin-Induced Pulmonary Fibrosis in Mice. Int. J. Mol. Sci. 2021, 22, 11152. [Google Scholar] [CrossRef]
- Schiff, M.H.; Whelton, A. Renal Toxicity Associated with Disease-Modifying Antirheumatic Drugs Used for the Treatment of Rheumatoid Arthritis. Semin. Arthritis Rheum. 2000, 30, 196–208. [Google Scholar] [CrossRef]
- Clark, D.A.; Coker, R. Transforming Growth Factor-Beta (TGF-Beta). Int. J. Biochem. Cell Biol. 1998, 30, 293–298. [Google Scholar] [CrossRef]
- Ma, T.-T.; Meng, X.-M. TGF-β/Smad and Renal Fibrosis. In Renal Fibrosis: Mechanisms and Therapies; Liu, B.-C., Lan, H.-Y., Lv, L.-L., Eds.; Advances in Experimental Medicine and Biology; Springer: Singapore, 2019; pp. 347–364. ISBN 9789811388712. [Google Scholar]
- Isaka, Y. Targeting TGF-β Signaling in Kidney Fibrosis. Int. J. Mol. Sci. 2018, 19, 2532. [Google Scholar] [CrossRef] [Green Version]
- Dewald, O.; Zymek, P.; Winkelmann, K.; Koerting, A.; Ren, G.; Abou-Khamis, T.; Michael, L.H.; Rollins, B.J.; Entman, M.L.; Frangogiannis, N.G. CCL2/Monocyte Chemoattractant Protein-1 Regulates Inflammatory Responses Critical to Healing Myocardial Infarcts. Circ. Res. 2005, 96, 881–889. [Google Scholar] [CrossRef] [Green Version]
- Böttinger, E.P.; Bitzer, M. TGF-ß Signaling in Renal Disease. J. Am. Soc. Nephrol. 2002, 13, 2600–2610. [Google Scholar] [CrossRef] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aljohani, A.A.; Alqarni, Y.S.; Alrashidi, M.N.; Aljuhani, M.H.; Shehata, S.A.; El-Kherbetawy, M.K.; Prabahar, K.; Alshaman, R.; Alattar, A.; Helaly, A.M.N.; et al. The Anti-Rheumatic Drug, Leflunomide, Induces Nephrotoxicity in Mice via Upregulation of TGFβ-Mediated p53/Smad2/3 Signaling. Toxics 2022, 10, 274. https://doi.org/10.3390/toxics10050274
Aljohani AA, Alqarni YS, Alrashidi MN, Aljuhani MH, Shehata SA, El-Kherbetawy MK, Prabahar K, Alshaman R, Alattar A, Helaly AMN, et al. The Anti-Rheumatic Drug, Leflunomide, Induces Nephrotoxicity in Mice via Upregulation of TGFβ-Mediated p53/Smad2/3 Signaling. Toxics. 2022; 10(5):274. https://doi.org/10.3390/toxics10050274
Chicago/Turabian StyleAljohani, Alhanouf A., Yasmeen S. Alqarni, Maram N. Alrashidi, Maha H. Aljuhani, Shaimaa A. Shehata, Mohamed K. El-Kherbetawy, Kousalya Prabahar, Reem Alshaman, Abdullah Alattar, Ahmed M. N. Helaly, and et al. 2022. "The Anti-Rheumatic Drug, Leflunomide, Induces Nephrotoxicity in Mice via Upregulation of TGFβ-Mediated p53/Smad2/3 Signaling" Toxics 10, no. 5: 274. https://doi.org/10.3390/toxics10050274
APA StyleAljohani, A. A., Alqarni, Y. S., Alrashidi, M. N., Aljuhani, M. H., Shehata, S. A., El-Kherbetawy, M. K., Prabahar, K., Alshaman, R., Alattar, A., Helaly, A. M. N., Ateyya, H., Ismail, E. A., & Zaitone, S. A. (2022). The Anti-Rheumatic Drug, Leflunomide, Induces Nephrotoxicity in Mice via Upregulation of TGFβ-Mediated p53/Smad2/3 Signaling. Toxics, 10(5), 274. https://doi.org/10.3390/toxics10050274