Abstract
Microalgal biomass has emerged as a valuable and nutrient-rich source of novel plant-based foods of the future, with several demonstrated benefits. In addition to their green and health-promoting characteristics, these foods exhibit bioactive properties that contribute to a range of physiological benefits. Photoautotrophic microalgae are particularly important as a source of food products due to their ability to biosynthesize high-value compounds. Their photosynthetic efficiency and biosynthetic activity are directly influenced by light conditions. The primary goal of this study is to track the changes in the light requirements of various high-value microalgae species and use advanced systems to regulate these conditions. Artificial intelligence (AI) and machine learning (ML) models have emerged as pivotal tools for intelligent microalgal cultivation. This approach involves the continuous monitoring of microalgal growth, along with the real-time optimization of environmental factors and light conditions. By accumulating data through cultivation experiments and training AI models, the development of intelligent microalgae cell factories is becoming increasingly feasible. This review provides a concise overview of the regulatory mechanisms that govern microalgae growth in response to light conditions, explores the utilization of microalgae-based products in plant-based foods, and highlights the potential for future research on intelligent microalgae cultivation systems.
1. Introduction
In recent years, the plant-based (PB) food market has demonstrated substantial expansion. According to some government market reports, globally valued at approximately USD 43.77 billion in 2024, this market is projected to reach USD 85 billion by 2030, growing at a combined annual growth rate (CAGR) of 9–12%. Parallel to this trend, the plant-based milk sector exceeded USD 42 billion (CNY 300 billion) in 2022 and is expected to sustain a >20% annual growth rate post-2025. Notably, plant-based meat products accounted for 12% of fast food menu offerings in 2024, with the projections indicating this share will rise to 35% by 2030, while business-to-business (B2B) procurement volumes increase at 26% annually, largely driven by mounting consumer concerns regarding the environmental, health, and ethical implications of animal-based (AB) foods. Livestock production contributes significantly across the ecological, agronomic, socioeconomic, and economic domains, but the production of meat from pastured animals remains a major contributor to greenhouse gas emission, freshwater depletion, pollution, and biodiversity loss [1]. Despite the mounting interest in PB foods, most individuals still primarily consume AB products [2]. Consequently, PB food producers are compelled to formulate PB foods that resemble and taste like AB products, including meat, seafood, eggs, and dairy products [3,4]. The nutritional gaps in PB foods have driven the incorporation of functional components, such as natural pigments (e.g., carotenoids), plant-derived proteins (e.g., pea protein), and antioxidants (e.g., polyphenols), to enhance their nutritional adequacy [5]. Microalgal biomass is regarded as a significant renewable resource for extracting various active ingredients that possess antimicrobial or antioxidant properties and are utilized to enhance the properties of food products [6,7]. As a representative example, consider ‘blue foods’, aquatic-derived food resources that have been extensively historically consumed with a high nutritional value. Among these, specific microalgae species are particularly noteworthy due to their superior protein content, comprehensive essential amino acid profiles, widespread bioavailability, and low-level allergenicity [8]. Microalgae are widely recognized as a promising resource for the development of PB food products [5], including PB meat [9], PB seafood substitutes [10], dairy products [11], and flavoring agents for PB seafood [12]. The utilization of microalgae in many cultures for the production of value-added bioproducts has attracted considerable research interest in recent years, driven by the global pursuit of sustainability [13,14]. In addition to the direct use of algae as food additives [15], the utilization of microalgae allows for the production of high-value products, including proteins, carotenoids such as astaxanthin, and polyunsaturated fatty acids (PUFAs) [16]. The principal sources of these substances are plants and animals in nature [17,18]. A growing supply–demand imbalance exists due to the escalating industrial demand and the use of unsustainable resources. To meet the market demand for these products, identifying alternative sources is of the utmost importance. Microalgae represent a promising green resource, so the selection and large-scale cultivation of high-yield algal species will be a key area of future development [19]. To ensure commercial viability, optimal cultivation conditions and techniques must be established. For photoautotrophic microalgae, light is a critical factor influencing cellular growth and biosynthetic rates. Some examples of microalgae mass cultures using natural or artificial light sources from some of the world’s leading algae companies are listed in Table 1. Thus, investigating light conditions and advancing lighting technologies are essential for enhancing productivity. In current microalgal cultivation practices, the application of multiple abiotic stresses, such as modulated light intensity, specific light wavelengths, and controlled nitrogen source concentrations, has demonstrated its potential to enhance the production of commercially valuable bioproducts [20]. This strategy could enhance industrial scalability by improving process efficiency and product yield compared to those of the traditional cultivation methods.

Table 1.
Some international companies that use natural or artificial light sources for mass cultures and their main products. (Tabulated data were compiled from official corporate portals and annual reports of respective entities.)
To obtain large-scale microalgae cultures, it is imperative to exercise careful control over light conditions. This encompasses the intensity and spectral quality of light, which are instrumental in determining the culture’s success [21]. Among these parameters, light intensity and wavelength constitute the most critical environmental regulators, directly governing photosynthetic efficiency and growth dynamics in photosynthetic microalgae [22,23]. Microalgae generally employ photosynthesis to utilize outdoor sunlight or ordinary white light provided by an artificial light source [24]. However, photosynthetic pigments selectively absorb specific wavelengths of light. The use of red or blue light sources alone for microalgae cultivation has been shown to increase the biomass productivity of various microalgae compared to that of ordinary white light [25]. The dynamic adjustment of light intensity and quality is necessary to enhance the light utilization efficiency of microalgal cells in actual production. The advent of artificial intelligence (AI) technology has precipitated a paradigm shift in various industries, with the integration of AI becoming a prevailing trend in future development. This efficacy is attributable to the fact that machine learning algorithm (MLA) approaches furnish more comprehensive insights into the uncertainty of biological processes compared to the traditional phenomenological or dynamical models [26]. Numerous studies have been conducted on the utilization of AI for the continuous monitoring and real-time dynamic adjustment of microalgae culture processes. Recent research focuses on areas such as microalgae-based wastewater treatment [27], intelligent autotrophic cultures [28], and product intelligent extraction [29], among others. AI-based monitoring and dynamic light regulation are expected to play a pivotal role in future microalgal cultivation systems. The objective of this review is to examine the impact of light conditions on the growth and synthesis of valuable microalgae products, including proteins, oils, and natural pigments that can be utilized in the production of PB food products. It is important in the formulation of new PB foods. The insights derived from this review will contribute to the optimization of microalgae cultivation methods, thereby facilitating the acceleration of biomass accumulation and the biosynthesis of high-value products.
2. Methodology
A comprehensive search strategy was implemented to identify relevant literature across three domains: (1) the light-mediated regulation of microalgal growth and biosynthesis, (2) microalgae applications in plant-based foods, and (3) the AI-driven optimization of microalgal cultivation and harvesting. A systematic literature assessment was therefore conducted to evaluate state-of-the-art knowledge. As shown in Figure 1, the search strategy employed core keywords, including ‘light parameters’, ‘microalgal cultivation’, ‘plant-based foods’, ‘microalgal biomolecules’ (encompassing proteins, fatty acids, and natural pigments), and ‘artificial intelligence’. To ensure efficient literature synthesis, core studies were systematically retrieved from multidisciplinary databases, including Web of Science, Scopus, PubMed, IEEE Xplore, ScienceDirect, SpringerLink, Wiley Online Library, Taylor & Francis Online, MDPI platforms, AGRO, Hindawi, Google Scholar, and China National Knowledge Infrastructure (CNKI). Following initial keyword searches, the identified literature underwent systematic assessment against predefined eligibility criteria for inclusion/exclusion. Articles directly aligning with the core search keywords were prioritized for retention. The metadata (titles, keywords, abstracts, authors, and references) extracted from selected publications constituted the primary dataset. Non-English publications were excluded. Ultimately, the relevant manuscript sections of 171 articles meeting all the criteria were thematically synthesized.
Figure 1.
Literature search approach.
3. Effects of Light Changes on Photosynthesis and Growth of Microalgae
3.1. Microalgal Photoadaptation and Biomass Accumulation Under Variable Light Intensities
The photosynthetic pigments present in eukaryotic algal cells form a complete vesicle network with a vesicle-like membrane; the vesicles can be stacked or penetrate through the stroma [30] (Figure 2). Changes in light have been shown to primarily alter algal cell growth by affecting electron flow and reducing power [31]. Photosynthesis is one of the most sensitive physiological processes in microalgae, undergoing substantial structural and functional changes in the photosynthetic apparatus under diverse environmental conditions. These responses include modifications in light-harvesting complexes, antenna systems, and reaction centers, along with adjustments in excitation energy distribution between photosystem I (PSI) and photosystem II (PSII) [32]. Within an optimal range, increasing light intensity enhances electron flow through both PSI and PSII, thereby promoting the generation of ATP and NADPH. However, under excessive light conditions, the photosynthetic electron transport chain may become saturated, leading to its over-reduction and potential photodamage. To mitigate this, alternative dissipation mechanisms, such as non-photochemical quenching and cyclic electron flow, are activated to safely dissipate excess excitation energy and protect the photosynthetic machinery [33]. Carotenoids located within the photosynthetic reaction center P680 play a crucial role in protecting against photooxidative damage.
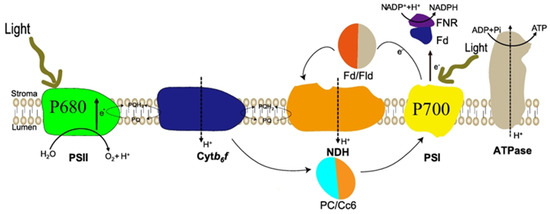
Figure 2.
Photosynthetic system of microalgae cells, PSI: photosystem I; PSII: photosystem II; FD: Ferredoxin; FNR: Fd-NADP reductase; Fld: Flavodoxin; NDH: NAD(P)H dehydrogenase complex; PC: plastocyanin; Cc6: cytochrome; Cytb6/f: plastocyanin. NDH: NAD(P)H dehydrogenase complex; PC: plastocyanin; Cc6: cytochrome; Cyt b6/f: cytochrome b6-f complex; PQ: plastoquinone.
Photosynthetically active radiation (PAR) refers to the portion of solar radiation that can be utilized by photosynthetic organisms, typically defined within the 400–700 nm wavelength range. Although chlorophylls absorb light primarily in the blue and red regions of this spectrum, their absorption capacity is limited to specific wavelengths. To broaden the usable spectrum, photosynthetic organisms employ light-harvesting complexes (LHCs), which contain numerous chlorophyll molecules and carotenoids that collectively capture and transfer excitation energy to the reaction centers [34,35]. In addition to their structural role within the photosynthetic apparatus, carotenoids act as auxiliary light-harvesting pigments and photoprotective agents, quenching reactive oxygen species (ROS) and dissipating excess excitation energy, thereby reducing photodamage and photoinhibition in photosynthetic organisms exposed to light stress [36]. The intensity of incident light plays a critical role in regulating photosynthetic activity, which, in turn, governs the growth dynamics and biomass productivity of microalgae. Variations in light intensity result in differential growth rates and accumulation patterns across microalgal species. Additionally, the contents of cytochromes and lipids, as well as the composition and ratio of fatty acids, will also differ based on these light conditions [37]. To ensure the stable production of PB-food additives from microalgae, it is necessary to provide suitable light conditions for photosynthetic algae. Ensuring an adequate supply of light radiation is a major challenge in photoautotrophic microalgae cultivation, as it is essential to provide the appropriate amount of light to prevent both light limitation and photoinhibition during growth [37]. To ascertain the optimal light intensity for the cultivation of high-value microalgae, the impact of a diverse range of light intensities on the growth and biosynthesis of various microalgal species has been studied [38,39,40]. In most experiments on well-studied high-value microalgae, the light intensities ranged from 50 to 1500 μmol photons m−2 s−1, with optimal growth typically occurring from around 60 to 200 μmol photons m−2 s−1 (Table 2).
Table 2.
Optimum light intensity values for maximum growth rates of algae of different taxonomic groups.
The growth rate of the diatomaceous microalgae Phaeodactylum tricornutum was found to be approximately 2 × 106 cells ml−1 day−1 at an initial light intensity of 20 μmol photons m−2 s−1. However, when the light intensity was increased to 200 μmol photons m−2 s−1, the growth and division rate increased threefold [55]. The impact of light on photosynthesis is significant. Upon adjusting the light intensity from low (40 μmol photons m−2 s−1) to high (350 μmol photons m−2 s−1), it was observed that photosynthesis and respiration increased, and plastid–mitochondria contact was boosted [56]. The rate of carbon fixation by cellular photosynthesis was increased, resulting in the increased accumulation of organic matter within the cells. The cellular metabolism and biosynthesis of P. tricornutum were found to be significantly increased. Nevertheless, when light intensity exceeded 500 μmol photons m−2 s−1, the growth rate ceased to increase in tandem with the intensification of illumination. Instead, a decline in growth was observed, accompanied by the emergence of a negative effect on growth stagnation. High-intensity light impedes the process of cellular photosynthesis, reducing the rate of the synthesis of photosynthetic pigments [57].
In an outdoor bioreactor, Metsoviti et al. [58] observed the notable impact of light intensity on the growth and biomass productivity of Chlorella vulgaris. Terrestrial photosynthetically active radiation (PAR) intensities exhibit substantial variability depending on geographic location, seasonal changes, and atmospheric conditions. Increased light availability during sunnier periods has been shown to promote C. vulgaris growth, particularly by enhancing specific growth rates, the maximum optical density, and the overall biomass productivity. Concurrent studies by Khoeyi and Seyfabadi yielded analogous results, observing a 20% surge in biomass productivity upon elevating the light intensity to which C. vulgaris was exposed from 65 μmol photons m−2 s−1 to 78 μmol photons m−2 s−1 [59]. C. vulgaris is relatively less sensitive to changes in light intensity and can maintain good growth under higher-intensity light. Some studies have also shown that C. vulgaris can grow under extremely intense light. Other researchers have successfully cultivated C. vulgaris at ultra-high light intensities [60] (7000 μmol photons m−2 s−1) using a strobe effect with flickering light [61] rather than continuous exposure. Additionally, the variations in light intensity also significantly influenced the photosynthetic pigment content of C. vulgaris, affecting both cell growth and division rates. The concentration of chlorophyll decreased by more than 20% with increasing light intensity, whereas the content of β-carotene exhibited a contrasting trend, increasing by approximately 30% with increasing light intensity [62]. The primary rationale for this phenomenon is that excessive light intensity can result in light saturation, which may potentially lead to cellular damage and the disruption of photosynthetic processes. Carotenoids serve as vital antioxidants that mitigate the adverse effects of light saturation, including photoinhibition, on biological systems [63]. The researchers delineated the boundaries between weak and strong light by observing alterations in the growth pattern of Haematococcus pluvialis cells and the photoinhibition point [64]. Based on the experimental results, 100 μmol photons m−2 s−1 was selected as the optimum light intensity for H. pluvialis. At this time, the biomass of H. pluvialis was 4 g/L, and the biomass productivity value was 80 mg L−1 d−1. The aforementioned high-value microalgae exhibit sensitivity to fluctuations in light intensity. Maintaining photosynthetic efficiency in microalgal cultures requires adaptive light regulation. This can be achieved through feedback control systems that continuously monitor cell density and dynamically adjust light intensity to maintain irradiance within species-specific optimal ranges.
3.2. Microalgal Photoadaptation and Biomass Accumulation Under Variable Light Quality
The metabolic processes of microalgae cells can be influenced by light of a specific spectral composition, resulting in the accumulation of proteins, carbohydrates, or lipids [65]. Consequently, different wavelengths of light have been demonstrated to alter the growth of algal cells and the synthesis of substances. Jungandreas et al. [66] examined the impact of red light (RL), blue light (BL), and natural white light (WL) on the growth rate of P. tricornutum. The findings indicated that red light could facilitate the growth of P. tricornutum during the initial phase of cultivation; however, in the subsequent phase, the impacts of the three light qualities on the density of P. tricornutum cells were nearly indistinguishable. Significant differences in primary photosynthetic activity were observed between RL- and BL-acclimated microalgal cells. Specifically, the quantum yield of PSII across a range of light intensities was markedly lower in the BL-acclimated cells, indicating reduced photochemical efficiency under BL conditions. The synthesis of photosynthetic pigments in P. tricornutum was markedly influenced by different light qualities. Both RL and BL were observed to promote photosynthetic pigment synthesis, with RL exhibiting a more pronounced effect than BL [66]. In contrast to the findings observed in the case of P. tricornutum, the study demonstrated that the utilization of BL as a light source resulted in the enhancement of the optimal light intensity required for the growth of C. vulgaris [67]. The shorter wavelength of BL activates specific photoreceptor proteins in C. vulgaris, thereby promoting C. vulgaris biomass accumulation. The concentrations of chlorophyll a, chlorophyll b, and β-carotene increased under BL conditions at the same light intensity [24]. In contrast to the positive effect of BL on the growth of C. vulgaris cells, RL has been shown to cause damage to these cells and to reduce their growth rate [68]. Both red and blue light have been demonstrated to promote the growth and reproduction of H. pluvialis, and both of these variables accumulate greater biomasses in comparison to WL. Furthermore, the astaxanthin concentration of H. pluvialis has been shown to increase significantly in response to BL, reaching levels that are eight times higher than those observed under WL and two times higher than those observed under RL [69]. As demonstrated in the relevant literature, reactive oxygen species (ROS) have been shown to function as a cellular signal, thereby activating cellular metabolism in H. pluvialis [70], thereby promoting astaxanthin synthesis. BL undoubtedly promotes astaxanthin synthesis by stimulating the metabolism of H. pluvialis, leading to the production of ROS. Consequently, it is imperative to adjust the light quality according to the microalgae species during the actual cultivation process. The utilization of a combination of light-emitting diode (LED) lights of different colors as a light source has been demonstrated to be an effective solution to this problem.
4. Light Conditions for the High-Value Products of Microalgae Synthesis
The extraction of microalgae biomass and the subsequent analysis of the proteins, fatty acids, natural pigments, polysaccharides, and other substances present within this biomass are of significant importance in the realm of raw materials and food additives for PB foods (Table 3). Therefore, it is necessary to clarify the effects of changing light conditions on the synthesis and accumulation of these important nutrients in the algal cell. Microalgae productivity was found to be increased by regulating light factors (Table 4).
Table 3.
Microalgal biomass and microalgal extracts in plant-based foods.
Table 4.
Influence of light intensity and light quality on nutrient productivity of microalgae.
4.1. Long-Chain Unsaturated Fatty Acids
A plethora of studies demonstrated the favorable effects of PUFAs on human health [81]. These include the ability to regulate blood pressure, strengthen the immune system, and reduce inflammation [82]. The addition of PUFAs to PB protein foods can increase their nutritional value, improve their water-binding capacity, and enhance their fat-binding properties [83]. Microalgae fatty acids can be a high-quality source of PUFAs in plant-based foods. The synthesis of PUFAs requires desaturases and elongases, which facilitate the formation of double bonds and carbon chain extension in fatty acids [84]. This process is accompanied by a significant energy expenditure, ultimately leading to the production of PUFAs (Figure 3) [85,86]. Phaeodactylum tricornutum, an important marine diatom, has emerged as a promising candidate for industrial biotechnology in recent years. Its rapid growth, robust metabolite production, and amenability to genetic engineering have established it as a model organism in algal bioengineering [87]. The market demand for Phaeodactylum tricornutum is expected to increase in parallel with the growing consumer interest in natural and organic ingredients. This trend is further driven by the rising demand for high-value bioactive compounds, such as eicosapentaenoic acid (EPA), fucoxanthin, and exopolysaccharides, for use in functional foods, nutraceuticals, and cosmetics [88]. P. tricornutum is regarded as a novel marine resource due to its abundance of EPA [89]. Changes in light intensity have been demonstrated to directly affect the EPA content of P. tricornutum. When the light intensity was increased from 30 μmol photons m−2 s−1 to 180 μmol photons m−2 s−1, the EPA content of fatty acids was observed to decrease from 17.2 to 13.8% [75]. Moreover, an increase in light intensity from 150 μmol photons m−2s−1 to 750 μmol photons m−2 s−1 was accompanied by a decrease in EPA content, with a reduction of 5.73% [90]. The EPA biosynthesis pathway in P. tricornutum involves the transcription of six major gene products (four desaturation steps and two elongation steps) [90]. Of the four desaturase genes analyzed (PTD5α, PTD5β, PTD6, and PTD15), only PTD5α was significantly upregulated in response to high-intensity light exposure (750 μmol photons m−2s−1). Conversely, the transcription of the other desaturase genes was significantly reduced across all the treatments, which also explains the observed decrease in the EPA content of P. tricornutum under high light intensity. The wavelength of light also affects the fatty acid content; a shift from red to blue light increased the lipid content of P. tricornutum, while a shift from blue to red light promoted the synthesis of carbohydrates [76]. BL enhanced P. tricornutum’s capacity to produce fatty acids, while RL resulted in a notable elevation in the proportion of PUFAs [91], so light quality must be adjusted to increase PUFA production. The utilization of yellow and blue light was conducive to the accumulation of total lipids in Thalassiosira pseudonana, with the contents of PUFA and EPA reaching over 34% and 16%, respectively, under both light conditions [92]. Therefore, in the large-scale cultivation of microalgae for fatty acid production, specific combinations of light sources need to be tailored to the growth characteristics of microalgae to improve the efficiency of light energy utilization.
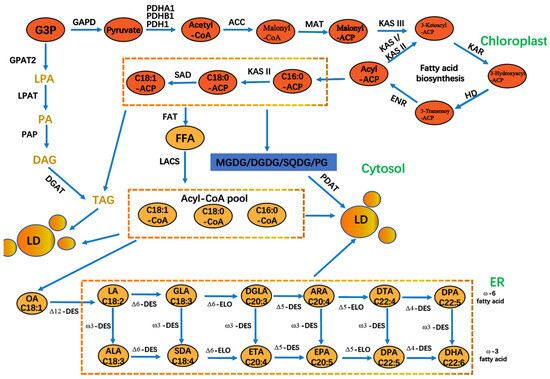
Figure 3.
The process of PUFA biosynthesis in P. tricornutum cells. GAPD: glyceraldehyde 3-phosphate dehydrogenase; PDH: pyruvate dehydrogenase; ACC: acetyl-CoA carboxylase; MAT: malonyl-CoA-acyl carrier protein transacylase; KAS: 3-ketoacyl-ACP synthase; HD: 3-hydroxyacyl-ACP dehydratase; ENR: enoyl-ACP reductase; KAR: β-ketoacyl carrier protein reductase; ADS: acyl-CoA desaturase; DGAT: acyl-CoA: diacylglycerol acyl transferase; FAT: fatty acyl-ACP thioesterase; FFA: free fatty acid; DGDG. DGDG: digalactosyldiacylglycerol; MGDG: monogalactosyldiacylglycerol; PG: phosphatidylglycerol; SQDG: sulfoquinovosyldiacylglycerol; LD: lipid droplets; PDAT: phosphatidylglycerol; SQDG: sulfoquinovosyldiacylglycerol droplets; PDAT: phospholipid diacylglycerol acyltransferase; G3P: glycerol 3 phosphate; GPAT: acyl-CoA glycerol-3-phosphate acyl transferase. LPAT: acyl-CoA lysophosphatidic acid acyl transferase; PAP: phosphatidic acid phosphatase; LACS: long chain acyl-CoA synthetase; PDH: pyruvate dehydrogenase. PDH: pyruvate dehydrogenase.
4.2. Fucoxanthin
Fucoxanthin, a marine-derived xanthophyll carotenoid, is primarily localized in the chloroplasts of brown algae (e.g., kelp and wakame) and diatoms. It plays a dual role in light harvesting and photoprotection and has attracted attention for its bioactive properties in the nutraceutical and cosmetic industries [93,94]. A multitude of in vitro and in vivo studies have substantiated the multifaceted pharmacological effects of fucoxanthin, including anticancer, antidiabetic, anti-inflammatory, and antioxidant properties [95]. Fucoxanthin is also widely used as a food additive, commonly incorporated into functional food products as a natural antioxidant [96], and also exhibits antimicrobial activity, with the potential to extend the shelf life of plant-based dairy products when incorporated into plant-based dairy alternatives [97]. The biosynthetic pathway of fucoxanthin in P. tricornutum has been largely elucidated (Figure 4). The production of fucoxanthin in P. tricornutum is highly sensitive to environmental conditions [98], with light conditions identified as a key factor influencing the content of fucoxanthin in cells [77,99]. P. tricornutum produces more fucoxanthin in low-intensity light (20 μmol photons m−2 s−1) and 100 μmol photons m−2 s−1 conditions than in high-intensity light (200 μmol photons m−2 s−1) conditions [78]. The expression of several key genes involved in carotenoid synthesis (PtPSY, PtPDS, and PtVDL1) and the formation of the fucoxanthin–chlorophyll protein (FCP) complex (PtLhcf5 and PtLhcf8) were identified by RNA sequencing and LC-MS/MS analysis in response to changes in light intensity [100]. A significant reduction in PtPDS gene expression was observed with increased light intensity, resulting in only one-tenth of the expression value under low light conditions. The expression of PtLhcf5 and PtLhcf8 exhibited a comparatively minor decline, amounting to approximately 20%. Light quality also influences the synthesis of fucoxanthin in P. tricornutum; the transition from white LED light to red LED light has been shown to enhance the expression of genes such as PSY, ZEP, VDE, and FCP-b, resulting in a notable increase in fucoxanthin accumulation [101]. In the presence of RL, the expression of genes associated with the light-harvesting protein complex was found to be upregulated, thus promoting the capture and conversion of light energy. Concurrently, fucoxanthin levels exhibited an analogous increase [102]. Under blue-green light conditions, the expression of genes involved in glycolysis, the tricarboxylic acid (TCA) cycle, and terpene skeleton biosynthesis is upregulated, thereby facilitating the synthesis of intracellular organic matter and promoting fucoxanthin accumulation. However, blue light is generally less effective than red light in enhancing fucoxanthin biosynthesis [103]. In the future, it will be necessary to optimize the ratio of red and blue LEDs to maximize the yield of fucoxanthin. A study employed LED lamps with adjustable spectral ratios as a tunable light source for the marine diatom Odontella aurita. When the red-light (RL)-to-blue-light (BL) ratio was set at 8:2, fucoxanthin productivity reached 9.41 mg L−1 d−1 [104]. These findings highlight the critical role of spectral optimization in maximizing the yield of high-value metabolites in microalgal biotechnology.
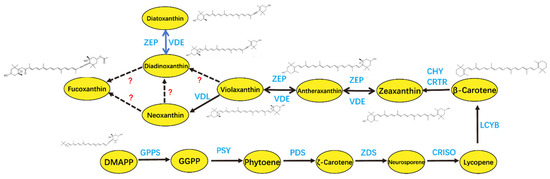
Figure 4.
Fucoxanthin biosynthesis in P. tricornutum with recently investigated genes. DMAPP: dimethylallyl pyrophosphate; GPPS: geranylgeranyl pyrophosphate synthase; GGPP: geranylgeranyl pyrophosphate; PSY: phytoene synthase; PDS: phytoene desaturase; ZDS: ζ-carotene desaturase; CRISO: carotene isomerase; LCYB: lycopene B-cyclase; CHY: carotene hydroxylase; CRTR: carotene hydroxylase; VDE: violaxanthin de-epoxidase; ZEP: zeaxanthin epoxidase; VDL: violaxanthin de-epoxidase-like.
4.3. Microalgal Protein
Microalgae are characterized by elevated protein levels, in addition to a comprehensive array of amino acids that are indispensable for the human body. The amino acid values are comparable to those found in high-quality protein sources [105], including egg whites, lactoglobulin, and soybean [106]. A significant number of PB meat and seafood products that contain microalgae proteins have already been developed [10,107]. Microalgal proteins have minimal adverse effects on the nutritional value and flavor of plant foods. In addition, these proteins have excellent solubility, emulsification, gelation, and foaming properties, and therefore have a wide range of uses in the food industry [108]. The primary microalgal species used for protein production include Spirulina sp., C. vulgaris, and Dunaliella salina. Spirulina sp., a cyanobacterium rich in phycocyanin and essential amino acids, is widely recognized for its nutritional and medicinal value. Its high protein content and abundance of bioactive compounds makes it a valuable functional ingredient in human health foods, aquaculture feed, and livestock nutrition. Notably, it exhibits well-documented antioxidant and immunomodulatory properties [109,110]. Owing to its favorable safety profile and robust productivity, Spirulina sp. has been extensively studied as a sustainable alternative to the traditional protein sources in both food and feed industries. Spirulina sp. biomass is characterized by high carbohydrate and protein contents. Spirulina sp. is a blue-green bacterium, with phycocyanin being of particular significance due to its functional properties [111]. In one study, the maximum protein content (37%) in Spirulina sp. was achieved at the lowest light intensity (9 μmol photons m−2 s−1) [112].
Furthermore, the biomass growth and protein content of Spirulina sp. are influenced by the photoperiod [113]. The results indicated that the highest biomass concentration (0.51 g/L) and the maximum protein content (26.2%) were attained under a 14L:10D photoperiod. Spirulina sp. was also found to have a preference for different wavelengths of light; the highest dry cell weight of 0.343 g/L was recorded for a white light-emitting diode. In contrast, the highest protein content of 64.10 ± 0.44% was registered with a blue light-emitting diode [114]. C. vulgaris is a species of edible algae that is notable for its high protein content [115]. It has been demonstrated that protein content is less sensitive to fluctuations in light intensity [116]. Similar to Spirulina sp., the protein content of C. vulgaris under BL conditions was found to be higher than that under red or white light conditions [117]. Using 14C-labeled amino acids, some researchers observed that blue light increased the incorporation rate of labeled amino acids into proteins, suggesting enhanced protein synthesis efficiency under blue light conditions. In the context of algae-based protein production, a two-stage cultivation process can be employed, with the quality of light being a pivotal factor in achieving the desired outcome. A recent study has demonstrated that the production of biomass and protein in D. salina can be significantly enhanced through the regulation of light quality and the implementation of a two-stage cultivation method. In the initial phase, the utilization of a light source comprising dynamically enhanced WL is paramount. After accumulating a certain amount of biomass using blue LED or red LED light as a light source to continue cultivation, the second stage of this method led to a notable enhancement in protein production, with the utilization of either red or blue light proving to be particularly effective [118]. Algal protein synthesis exhibits relatively low sensitivity to fluctuations in light intensity. Achieving high protein yields depends on three fundamental strategies: preventing photoinhibition, providing adequate blue light (BL), and ensuring sufficient nitrogen availability. These conditions collectively enhance nitrogen assimilation and promote the activity of protein biosynthetic pathways, making them essential for optimizing microalgal cultivation for protein-rich biomass.
4.4. Astaxanthin
Astaxanthin (3,3′-dihydroxy-β, β-1-carotene-4,4′-dione) is a natural carotenoid [119] with superior antioxidant properties compared to those of other carotenoids and vitamin E [120]. Astaxanthin can provide vibrant colors to plant-based foods. In addition to this capacity, it has been shown to possess antioxidant, antineoplastic, and hypotensive properties when incorporated into functional foods [121,122]. H. pluvialis is one of the most important sources of natural astaxanthin, and the pathway of its biosynthesis has been fully elucidated (Figure 5). The two most critical factors affecting growth and astaxanthin synthesis in H. pluvialis SAG 34-7 are light conditions and nitrogen concentration [123]. It is hypothesized that nitrogen starvation in combination with strong light is the optimal method for triggering the hyperaccumulation of astaxanthin in H. pluvialis (strain H2) [80,124]. However, one of the adverse effects of nitrogen starvation is a reduction in chlorophyll concentration within cells. Alterations in the concentration of photosynthetic pigments significantly influence the light absorption of H. pluvialis, which, in turn, affects the growth and reproduction rate of algal cells. The proposal of the two-step incubation method effectively addresses this issue [125] (Figure 6). The initial phase of the astaxanthin production process entails the cultivation of the green cells of H. pluvialis to accumulate a sufficient biomass. H. pluvialis exhibited optimal growth at a light intensity of 70 μmol photons m−2 s−1, which facilitated the expeditious accumulation of biomass [126,127]. However, in the red phase of H. pluvialis, intense light exceeding 100 μmol photons m−2 s−1 enhances natural astaxanthin accumulation [128]. Additionally, ROS generated as a consequence of photodamage act as a cellular signal to activate cellular metabolism in H. pluvialis [129], thereby promoting astaxanthin synthesis. At the gene transcription level, genes encoding specific enzymes required for β-carotene synthesis (psy,pds,zds,crt-iso,lycB) and related genes encoding key enzymes for bio-astaxanthin synthesis (bkt and crtr-b) exhibited increased expression upon increased light intensity [130,131,132]. Light saturation is uncommon in large-scale outdoor culture conditions, particularly during the red stage, which may be attributed to light scattering and mutual shading caused by high-cell-density cultures, which reduces the actual absorbed light irradiance. Therefore, higher astaxanthin accumulation in H. pluvialis can be achieved by combining nitrogen starvation with intense light in a large-scale culture. The cultivation of microalgae for astaxanthin production is also contingent upon the quality of light. In a particular study [79], astaxanthin production in H. pluvialis was observed to be most prolific under 3R4B (three red and four blue LED units) treatment, with a yield of 15.28 g/L. This yield was 2.2 times higher than that obtained under white light (WL) control conditions. In the context of future astaxanthin production in H. pluvialis, the use of combined red and blue light sources warrants consideration. This lighting strategy shows potential for enhancing the yield of a target compound.
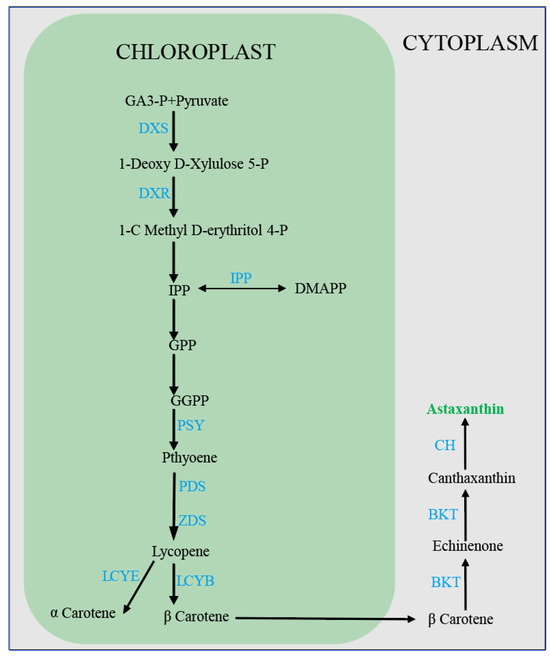
Figure 5.
Synthesis process of astaxanthin in H. pluvialis cells. GA3-p: glyceraldehyde-3-phosphate; DXS: 1-deoxy-D-xylulose 5-phosphate synthase; DXR: 1-Deoxy- d-xylulose-5-phosphate reductoisomerase; IPP. isopentenyl pyrophosphate; DMAPP: dimethylallyl pyrophosphate; GPP: geranyl pyrophosphate; GGPP: geranylgeranyl pyrophosphate; PSY: phytoene synthase; PDS: phytoene desaturase; ZDS: ζ-carotene desaturase; LCYB: lycopene β-cyclase; LCYE: lycopene ε-cyclase; BKT: β-carotene ketolase; CH: -carotene hydroxylase; GPP: geranyl pyro phosphate BKT: β-carotene ketolase; CH: β-carotene hydroxylase.
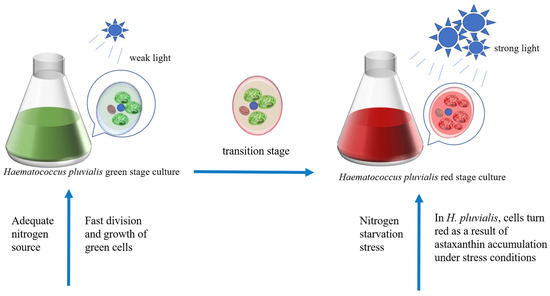
Figure 6.
The two-step incubation method of H. pluvialis. The green phase provides sufficient nitrogen and suitable light conditions for rapid biomass accumulation, and the red phase promotes the accumulation of astaxanthin in the cells of H. pluvialis under the combined stress of nitrogen starvation and high-intensity light.
4.5. Microalgal Polysaccharides
Plant-based meat substitutes are meat analogues that are typically characterized by their traditional and readily available nature, with their composition typically comprising plant proteins, polysaccharides, water, and oils [133]. Tissue-organized vegetable protein (TVP) is a structured vegetable protein that is produced through an extrusion process, the purpose of which is to mimic the fibrous structure of meat [134]. TVP can incorporate polysaccharides and non-textured proteins as a binder, thereby enhancing the water-holding capacity (WHC) and textural properties of PB meat analogues. Polysaccharides play a pivotal role as a suitable binder for the formation of stable gels [135]. Microalgal polysaccharides have been widely used in the food industry [136], and their incorporation has been shown to enhance the rheological characteristics of food substances, thereby contributing to their functionality and practical applications [137]. The utilization of PB foodstuffs fortified with microalgae polysaccharides has emerged as a novel approach to weight management. Research demonstrated that the administration of microalgae polysaccharides to mice with extreme obesity leads to the regulation of body weight and blood lipid levels, in addition to the restoration of beneficial intestinal bacteria [138]. Microalgal polysaccharides could play an important role in future PB foods, improving both their physical structure and nutritional value. Microalgal polysaccharide synthesis is also affected by light conditions. Frick et al. [139] observed that the highest chrysolaminarin content was attained under 50 μmol photons m−2 s−1 growth conditions of P. tricornutum. In addition to P. tricornutum, β-glucan was likewise produced mainly as chrysolaminarin in Haptophyta and Heterokontophyta. Light intensity was identified as a significant factor influencing the production of chrysolaminarin in these microalgae [140]. The transcript levels of genes were also compared between the light and dark cycles in O. aurita, and the expression levels of genes involved in the biosynthesis of chrysolaminarin, including UGPase/PGM, UGP1-2, UGPT, BGS, and TGS1-3, were observed to be higher at the beginning of the light period than in the dark period [141]. In Isochrysis zhangjiangensis, a marked elevation in the concentration of chrysolaminarin was discerned under high-intensity light in comparison to the corresponding cultures under low-intensity light, particularly on the initial day of incubation [142]. Similarly, Christian Schulze et al. [143] observed that the β-glucan content in Spirulina sp. platensis cells increased from 6.4% to 19.5% following an increase in light intensity from 50 μmol photons m−2 s−1 to 150 μmol photons m−2 s−1; however, when the light intensity was excessively high, there was a subsequent decrease in the content. Consequently, the light intensity should be regulated within a range of 200 μmol photons m−2 s−1 and adapted temporally by the concentration of algal cells.
5. Co-Regulatory Effects of Light and Other Environmental Factors
The physiological processes of microalgae are influenced by a combination of environmental factors. In addition to light conditions, the key variables include temperature; nitrogen availability; and the presence of exogenous additives, such as growth regulators and stress inducers [144,145]. Understanding the interactive effects of these factors is essential for optimizing microalgal cultivation systems aimed at maximizing biomass yield and target metabolite production. In general, the sensitivity of photosynthetic plants to light conditions is subject to changes in response to adjustments in ambient temperature [146], as do microalgae. There is evidence to suggest that the light saturation point of photosynthetic algae varies with temperature and decreases with a reduction in temperature [147]. A mathematical model has been developed to evaluate the photoautotrophic growth of P. tricornutum under a variety of incubation conditions, specifically regarding temperature and light intensity [148]. The optimum cultivation temperature and light intensity conditions were confirmed to be 21 °C and 496 μmol m−2s−1 at a constant aeration rate, respectively. It is noteworthy that a separate study demonstrated that the maximum biomass concentration, triacylglyceride (TAG) content, and EPA content of P. tricornutum under nitrogen starvation were each influenced by light intensity independently [149]. This indicates that light intensity and nitrogen content may be important interrelated factors affecting PUFA biosynthesis in P. tricornutum. The results of mass balance analyses demonstrated that under nitrogen starvation, P. tricornutum broke down some of the photosynthetic membrane lipids for the synthesis of EPA. However, the result is still a decrease in EPA content due to nitrogen starvation [150]; ensuring adequate nitrogen sources can therefore increase EPA yields. A study found that the activity of H. pluvialis cells dropped a lot in the first three days under strong light; lowering the nitrogen concentration in the culture can help restore their activity [151]. The results demonstrate that nitrogen starvation enhances the adaptive capacity of H. pluvialis to fluctuations in light intensity. The growth and astaxanthin accumulation of H. pluvialis are significantly influenced by the interaction between light intensity and nitrogen source concentration in a medium [152]. Nitrogen starvation is a prerequisite for the accumulation of astaxanthin in H. pluvialis; the effect of light factors on astaxanthin production in H. pluvialis needs to be investigated on this basis.
H. pluvialis can produce more astaxanthin during its red cell phase when affected by intense light, nitrogen deficiency, high salt conditions, or high temperatures. However, intense light also causes too many ROS to form, which can damage cells [153,154]. The incorporation of amino acids into the culture medium is anticipated to mitigate the level of ROS and enhance the biomass and astaxanthin production of H. pluvialis [155]. Previous research has demonstrated that the exogenous administration of arginine can augment the chlorophyll content and the expression of antioxidant activity genes in H. pluvialis, thereby reducing the detrimental effects of ROS [156]. Furthermore, in the process of culturing Isochrysis sp. for fucoxanthin production, it was observed that the incorporation of a low concentration of spermidine could stimulate algal cell proliferation and enhance fucoxanthin production. The addition of spermidine at a low light intensity of 20 μmol m−2 s−1 led to a maximal fucoxanthin yield of 6.11 mg/g [157]. In general, optimizing light conditions alone is insufficient to maximize yield and economic efficiency in microalgal production. To effectively regulate microalgal physiological activities and achieve synergistic improvements, other environmental factors must also be taken into account. These include factors such as temperature, pH, nutrient availability, and CO2 concentration, which collectively influence growth kinetics and metabolite accumulation.
6. Safety of Microalgae in Food Applications
The issue of food safety has always represented a primary concern for a variety of regulatory agencies. Consequently, any potential consumer hazards must be considered when promoting microalgae-based foods [158]. The safety of microalgal foods depends on the type of microalgae used. In the development and production of plant-based foods containing microalgae or microalgae derivatives, selecting the appropriate microalgae species following the existing food safety regulations is necessary. In Europe, the use of microalgae in food applications is strictly regulated under the Novel Foods Regulation (EU) 2015/2283 [159]. By 2024, only two species, C. vulgaris (a green microalga) and Spirulina sp. (a cyanobacterium commonly referred to as blue-green algae), will be authorized for human consumption. Under Novel Food legislation in Europe, the consumption of Odontella aurita, Tetraselmis chuii, and Euglena gracilis (a euglenozoan, but classified as microalgae for commercial purposes) is authorized. Other microalgal species are not permitted as food unless they undergo the novel food approval process, which requires comprehensive scientific safety assessments. The safety of microalgae is also contingent on the conditions of their culture and processing, which are frequently disregarded. These organisms can accumulate toxic compounds from various sources, including water and commercial fertilizers. In some cases, microalgae can adsorb toxic substances in water, including heavy metals, dyes, and antibiotics. Furthermore, open cultivation systems for microalgae present inherent contamination risks due to invasive microalgae, potentially jeopardizing product safety in scaled operations. To mitigate these safety risks, implementing a comprehensive water environment monitoring program is essential [160]. This initiative should focus on detecting emerging pollutants and harmful algal species, while establishing correlations between the microalgal product safety profiles and the cultivation/processing conditions. It is imperative to analyze the relationship between the toxicity/safety of microalgae products and the culture and processing conditions for the establishment of safe and efficient regulatory systems. Nethravathy MU et al. [161] pointed out some “checkpoints” that are very important for the safety of microalgae products: (1) the open environment must be regularly checked for biological and non-biological pollutants, (2) water quality must be checked, (3) potential health problems caused by a high nucleic acid content must be assessed, (4) the potential allergic risks of microalgae must be assessed, and (5) the safety of the entire production process must be assessed from upstream to downstream. The safe application of microalgae in the food industry can be ensured through rigorous safety testing in the selection of algal species, water quality control, biomass harvesting, and product extraction. In the future, the large-scale cultivation of microalgae for PB food production will require strict compliance with the existing laws and regulations, alongside relevant scientific literature. This is essential to ensure the safety of cultivation practices, the proper utilization of microalgal biomass, and the overall legality and acceptability of microalgae-derived food products. As the regulatory landscape continues to evolve, interdisciplinary collaboration among researchers, policy makers, and industry stakeholders will be critical to the safe and sustainable integration of microalgae into the human food chain.
7. Prospects
7.1. Application of AI in Microalgae Cultivation
Artificially illuminated plant factory technology has matured in recent years. The integration of artificial intelligence with LED lighting has proven to offer several key benefits. First, these facilities exhibit high levels of resource utilization, resulting in enhanced productivity per unit area of land. Secondly, they demonstrate resilience to weather conditions, thereby ensuring a consistent performance regardless of external factors [162]. AI is also becoming more popular in the study of microalgae because AI algorithms can provide useful information in situations with a lot of uncertainty. In general, AI algorithms can be divided into three types: machine learning, metaheuristics, and expert systems [163]. Before using AI to detect the microalgae growth process and dynamically adjust real-time culture parameters, a substantial volume of extant data concerning the growth of microalgae influenced by environmental factors must be entered into the AI system. This data is essential for training the AI to develop an accurate predictive model [164]. AI can analyze large amounts of data collected by sensors, which include temperature, humidity, carbon dioxide concentration, pH, and nitrogen and phosphorus concentrations [165]. These data can be used to learn about the growth of microalgae and the build-up of target products. In addition to the analysis of environmental factors, it is imperative to analyze the microalgal biomass concentration, photosynthetic activity, and cell color in conjunction with hardware facilities [166]. Subsequently, the decision information is analyzed and evaluated by the artificial neural network and the decision tree, and the computer then regulates the temperature, humidity, light intensity, and light quality of the control port of the microalgae cell factory (Figure 7). Currently, only one kind of lamp is usually used to provide the light for algal cell photosynthesis in microalgae mass cultures. However, the growth process of algal cells is dynamic, and fixed light conditions cannot meet the needs of algal cells at the different stages of development. The utilization of AI for the analysis of microalgae growth, with the subsequent adjustment of light intensity and the employment of LED lamps of varying colors to provide microalgae with different light qualities are indicative of a broad range of potential applications. Optimal illumination can effectively enhance the accumulation of microalgal biomass and the production of valuable products. With recent advancements in artificial intelligence, the integration of AI-driven technologies capable of analyzing the algal growth dynamics is being explored. The subsequent real-time adjustment of environmental parameters, such as light intensity, temperature, and humidity, holds considerable potential for enhancing the efficiency and sustainability of microalgae cultivation. Integrating AI-based solutions promises to reduce labor requirements, material consumption, and optimize the operational efficiency of microalgae production. More significantly, AI-regulated microalgae cell factories hold strong potential as a major future source of PB food. Their inherent intelligence and capacity to minimize contamination risks directly address critical food safety concerns.
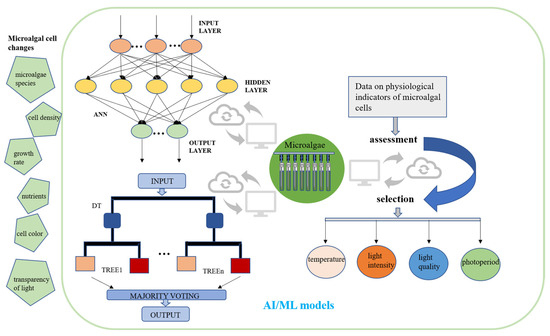
Figure 7.
A conceptual framework for integrating artificial intelligence and machine learning in optimizing large-scale microalgae cultivation systems. Using artificial intelligence to analyze the growth of microalgae cells and physiological indicators, such as microalgae growth rate and cell density, and then regulate the temperature and light conditions. AI, artificial intelligence; ML, machine learning; ANN, artificial neural network; DT, decision tree.
7.2. Limitations and Solutions
Microalgae are seen as a valuable source of nutrients for new PB foods. Microalgae biomass is rich in essential nutrients needed for good health, along with some active ingredients that offer health benefits. These factors will be a key part of the development of plant-based foods in the future. Despite recent advancements in research on breeding and cultivating high-value microalgae, the large-scale culture of microalgae and the harvesting of biomass for subsequent industrial production remain in their infancy. The selection of suitable and cost-effective light solutions for the growth of microalgae in large-scale cultivation is of paramount importance for the expansion of the microalgae industry. The current state of research on the impact of light conditions on the growth and productivity regulation of microalgae is characterized by the following limitations:
1. The research methods employed to study the impact of light on microalgae growth and productivity exhibit considerable inconsistency, resulting in a notable decline in the reliability of experimental outcomes. To ensure consistency, it is essential to provide comprehensive information in experimental studies, such as details on the light parameters, the microalgae concentration, the medium volume, the composition, as well as the concentration and the flow rate of CO2.
2. Conventionally, the cultivation of microalgae is predominantly characterized by constant light intensity and wavelength. However, in the future, it is imperative to formulate dynamic light intensity and wavelength changes, taking into account the unique characteristics of various microalgae species. This approach is more in line with the dynamic process of microalgae growth. To solve these problems, it is first necessary to accumulate a large amount of data from a large number of microalgae cultivation experiments. This data must then be analyzed through artificial intelligence, and the results must be used to train an AI model to determine the optimal environmental factors required during the growth of a certain type of microalgae. It should be noted that these factors are subject to change.
8. Conclusions
Microalgae are widely recognized as a vital component of plant-based (PB) foods and a sustainable source of diverse bioactive compounds. Microalgal cells are rich in high-quality proteins, essential amino acids, polyunsaturated fatty acids, and natural pigments, which can effectively compensate for the nutritional deficiencies commonly associated with the traditional PB foods. By precisely regulating cultivation conditions, specific functional components in microalgae can be enriched, allowing for targeted nutritional enhancement. Due to their highly efficient photosynthetic autotrophy, the biomass yield of microalgae per unit of land area can exceed that of conventional crops by more than tenfold. Moreover, their growth requires only sunlight, water, and carbon dioxide, significantly reducing reliance on freshwater and arable land. Currently, microalgae have been successfully applied in plant-based meat texture enhancement, dairy alternatives, and functional food additives. Natural pigments extracted from microalgae also serve as sustainable replacements for synthetic colorants in food formulations. Light is a fundamental requirement for the photoautotrophic growth of microalgae. The key parameters, such as light intensity, spectral composition, and the photoperiod, serve as effective tools for regulating microalgal growth, metabolic pathways, and the biosynthesis of biochemically valuable compounds. The optimal light intensity for most microalgal species typically ranges from 50 to 400 μmol photons m−2 s−1. However, a few species can tolerate and grow under intensities exceeding 3000 μmol photons m−2 s−1 due to their strong photo-regulatory capabilities. Variations in light intensity and wavelength can significantly influence lipid accumulation, fatty acid profiles, pigment synthesis, as well as protein and polysaccharide contents. Accordingly, light conditions can be dynamically adjusted, either in response to real-time growth status or tailored to the biosynthetic characteristics of target compounds, to improve light energy utilization efficiency.
To advance the development of microalgal cell factories, smart light regulation within a plant factory framework is essential for optimizing light energy utilization and enhancing biomass productivity. Artificial intelligence (AI) models trained on cultivation data can dynamically adjust light intensity according to specific growth stages, while programmable LED systems can synergistically accelerate microalgal growth and improve energy efficiency. Integrating such intelligent lighting strategies with environmental sensors and metabolic models may enable fully autonomous and energy-efficient algal cultivation systems in the future.
Author Contributions
Conceptualization, Y.C. and S.H.; software, F.W. and R.W.; validation, X.H. (Xinjuan Hu), F.Z. and L.X.; formal analysis, X.H. (Xiang Huang); writing—original draft preparation, X.H. (Xiang Huang); writing—review and editing, X.H. (Xiang Huang), O.U.R., Y.C. and S.H.; visualization, O.U.R. and R.W.; supervision, S.H. project administration, S.H. and R.W.; funding acquisition, S.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the National Natural Science Foundation of China (32361143786 and 32370387) and the Jiangsu Funding Program for Excellent Postdoctoral Talent (2024ZB884).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- McClements, D.J.; Grossmann, L. Next-Generation Plant-Based Foods: Challenges and Opportunities. Annu. Rev. Food Sci. Technol. 2024, 15, 79–101. [Google Scholar] [CrossRef] [PubMed]
- Tonsor, G.T.; Lusk, J.L. U.S. Perspective: Meat Demand Outdoes Meat Avoidance. Meat Sci. 2022, 190, 108843. [Google Scholar] [CrossRef] [PubMed]
- McClements, D.J.; Grossmann, L. (Eds.) Meat and Fish Alternatives. In Next-Generation Plant-Based Foods: Design, Production, and Properties; Springer International Publishing: Cham, Switzerland, 2022; pp. 285–339. ISBN 978-3-030-96764-2. [Google Scholar]
- McClements, D.J.; Grossmann, L. (Eds.) Eggs and Egg Products. In Next-Generation Plant-Based Foods: Design, Production, and Properties; Springer International Publishing: Cham, Switzerland, 2022; pp. 341–388. ISBN 978-3-030-96764-2. [Google Scholar]
- Alcorta, A.; Porta, A.; Tárrega, A.; Alvarez, M.D.; Vaquero, M.P. Foods for Plant-Based Diets: Challenges and Innovations. Foods 2021, 10, 293. [Google Scholar] [CrossRef]
- Shah, M.A.R.; Zhang, Y.; Cui, Y.; Hu, X.; Zhu, F.; Kumar, S.; Li, G.; Kubar, A.A.; Mehmood, S.; Huo, S. Ultrasonic-Assisted Green Extraction and Incorporation of Spirulina platensis Bioactive Components into Turmeric Essential Oil-in-Water Nanoemulsion for Enhanced Antioxidant and Antimicrobial Activities. Food Chem. 2024, 452, 139561. [Google Scholar] [CrossRef] [PubMed]
- Surendhiran, D.; Li, C.; Cui, H.; Lin, L. Marine Algae as Efficacious Bioresources Housing Antimicrobial Compounds for Preserving Foods—A Review. Int. J. Food Microbiol. 2021, 358, 109416. [Google Scholar] [CrossRef]
- Li, Y.; Xiang, N.; Zhu, Y.; Yang, M.; Shi, C.; Tang, Y.; Sun, W.; Sheng, K.; Liu, D.; Zhang, X. Blue Source-Based Food Alternative Proteins: Exploring Aquatic Plant-Based and Cell-Based Sources for Sustainable Nutrition. Trends Food Sci. Technol. 2024, 147, 104439. [Google Scholar] [CrossRef]
- Zhu, J.; Xiao, X.; Du, W.; Cai, Y.; Yang, Z.; Yin, Y.; Wakisaka, M.; Wang, J.; Zhou, Z.; Liu, D.; et al. Leveraging Microalgae as a Sustainable Ingredient for Meat Analogues. Food Chem. 2024, 450, 139360. [Google Scholar] [CrossRef]
- Siddiqui, S.A.; Ucak, İ.; Afreen, M.; Sasidharan, A.; Yunusa, B.M.; Bhowmik, S.; Pandiselvam, R.; Ambartsumov, T.G.; Shah, M.A. Microalgae as a Potential Raw Material for Plant-Based Seafood Alternatives: A Comprehensive Review. Food Sci. Nutr. 2024, 12, 8559–8593. [Google Scholar] [CrossRef]
- Bhatnagar, P.; Gururani, P.; Parveen, A.; Gautam, P.; Chandra Joshi, N.; Tomar, M.S.; Nanda, M.; Vlaskin, M.S.; Kumar, V. Algae: A Promising and Sustainable Protein-Rich Food Ingredient for Bakery and Dairy Products. Food Chem. 2024, 441, 138322. [Google Scholar] [CrossRef]
- Coleman, B.; Van Poucke, C.; Dewitte, B.; Ruttens, A.; Moerdijk-Poortvliet, T.; Latsos, C.; De Reu, K.; Blommaert, L.; Duquenne, B.; Timmermans, K.; et al. Potential of Microalgae as Flavoring Agents for Plant-Based Seafood Alternatives. Future Foods 2022, 5, 100139. [Google Scholar] [CrossRef]
- Garcia-Perez, P.; Cassani, L.; Garcia-Oliveira, P.; Xiao, J.; Simal-Gandara, J.; Prieto, M.A.; Lucini, L. Algal Nutraceuticals: A Perspective on Metabolic Diversity, Current Food Applications, and Prospects in the Field of Metabolomics. Food Chem. 2023, 409, 135295. [Google Scholar] [CrossRef] [PubMed]
- Chamorro, F.; Carpena, M.; Fraga-Corral, M.; Echave, J.; Riaz Rajoka, M.S.; Barba, F.J.; Cao, H.; Xiao, J.; Prieto, M.A.; Simal-Gandara, J. Valorization of Kiwi Agricultural Waste and Industry By-Products by Recovering Bioactive Compounds and Applications as Food Additives: A Circular Economy Model. Food Chem. 2022, 370, 131315. [Google Scholar] [CrossRef] [PubMed]
- Akyil, S.; İlter, I.; Koç, M.; Ertekin, F. Recent Trends in Extraction Techniques for High Value Compounds from Algae as Food Additives. Turk. J. Agric. Food Sci. Technol. 2018, 6, 1008–1014. [Google Scholar] [CrossRef]
- Shitanaka, T.; Fujioka, H.; Khan, M.; Kaur, M.; Du, Z.-Y.; Khanal, S.K. Recent Advances in Microalgal Production, Harvesting, Prediction, Optimization, and Control Strategies. Bioresour. Technol. 2024, 391, 129924. [Google Scholar] [CrossRef]
- Jing, H.; Nie, M.; Dai, Z.; Xiao, Y.; Song, J.; Zhang, Z.; Zhou, C.; Li, D. Identification of Carotenoids from Fruits and Vegetables with or without Saponification and Evaluation of Their Antioxidant Activities. J. Food Sci. 2023, 88, 2693–2703. [Google Scholar] [CrossRef]
- Ahmad, N.; Hussain, A.; Khan, S.; Korma, S.A.; Hussain, G.; Aadil, R.M.; Siddique, R.; Ali, A.; Shabbir, U.; Haq, A.U.; et al. Impact of Thermal Extrusion and Microwave Vacuum Drying on Fatty Acids Profile during Fish Powder Preparation. Food Sci. Nutr. 2021, 9, 2743–2753. [Google Scholar] [CrossRef]
- Khan, M.I.; Shin, J.H.; Kim, J.D. The Promising Future of Microalgae: Current Status, Challenges, and Optimization of a Sustainable and Renewable Industry for Biofuels, Feed, and Other Products. Microb. Cell Fact. 2018, 17, 36. [Google Scholar] [CrossRef]
- Song, X.; Liu, B.-F.; Kong, F.; Ren, N.-Q.; Ren, H.-Y. Overview on Stress-Induced Strategies for Enhanced Microalgae Lipid Production: Application, Mechanisms and Challenges. Resour. Conserv. Recycl. 2022, 183, 106355. [Google Scholar] [CrossRef]
- Maltsev, Y.; Maltseva, K.; Kulikovskiy, M.; Maltseva, S. Influence of Light Conditions on Microalgae Growth and Content of Lipids, Carotenoids, and Fatty Acid Composition. Biology 2021, 10, 1060. [Google Scholar] [CrossRef]
- Parveen, A.; Bhatnagar, P.; Gautam, P.; Bisht, B.; Nanda, M.; Kumar, S.; Vlaskin, M.S.; Kumar, V. Enhancing the Bio-Prospective of Microalgae by Different Light Systems and Photoperiods. Photochem. Photobiol. Sci. 2023, 22, 2687–2698. [Google Scholar] [CrossRef]
- Chávez-Fuentes, P.; Ruiz-Marin, A.; Canedo-López, Y. Biodiesel Synthesis from Chlorella vulgaris under Effect of Nitrogen Limitation, Intensity and Quality Light: Estimation on the Based Fatty Acids Profiles. Mol. Biol. Rep. 2018, 45, 1145–1154. [Google Scholar] [CrossRef] [PubMed]
- Baidya, A.; Akter, T.; Islam, M.R.; Shah, A.K.M.A.; Hossain, M.A.; Salam, M.A.; Paul, S.I. Effect of Different Wavelengths of LED Light on the Growth, Chlorophyll, β-Carotene Content and Proximate Composition of Chlorella Ellipsoidea. Heliyon 2021, 7, e08525. [Google Scholar] [CrossRef] [PubMed]
- Schulze, P.S.C.; Barreira, L.A.; Pereira, H.G.C.; Perales, J.A.; Varela, J.C.S. Light Emitting Diodes (LEDs) Applied to Microalgal Production. Trends Biotechnol. 2014, 32, 422–430. [Google Scholar] [CrossRef] [PubMed]
- Sundui, B.; Ramirez Calderon, O.A.; Abdeldayem, O.M.; Lázaro-Gil, J.; Rene, E.R.; Sambuu, U. Applications of Machine Learning Algorithms for Biological Wastewater Treatment: Updates and Perspectives. Clean. Technol. Environ. Policy 2021, 23, 127–143. [Google Scholar] [CrossRef]
- Oruganti, R.K.; Biji, A.P.; Lanuyanger, T.; Show, P.L.; Sriariyanun, M.; Upadhyayula, V.K.K.; Gadhamshetty, V.; Bhattacharyya, D. Artificial Intelligence and Machine Learning Tools for High-Performance Microalgal Wastewater Treatment and Algal Biorefinery: A Critical Review. Sci. Total Environ. 2023, 876, 162797. [Google Scholar] [CrossRef]
- Chong, J.W.R.; Khoo, K.S.; Chew, K.W.; Ting, H.-Y.; Iwamoto, K.; Ruan, R.; Ma, Z.; Show, P.L. Artificial Intelligence-Driven Microalgae Autotrophic Batch Cultivation: A Comparative Study of Machine and Deep Learning-Based Image Classification Models. Algal Res. 2024, 79, 103400. [Google Scholar] [CrossRef]
- Chong, J.W.R.; Tang, D.Y.Y.; Leong, H.Y.; Khoo, K.S.; Show, P.L.; Chew, K.W. Bridging Artificial Intelligence and Fucoxanthin for the Recovery and Quantification from Microalgae. Bioengineered 2023, 14, 2244232. [Google Scholar] [CrossRef]
- HÄDER, D.-P. Photosynthesis in Plants and Algae. Anticancer Res. 2022, 42, 5035–5041. [Google Scholar] [CrossRef]
- Lehmuskero, A.; Skogen Chauton, M.; Boström, T. Light and Photosynthetic Microalgae: A Review of Cellular- and Molecular-Scale Optical Processes. Prog. Oceanogr. 2018, 168, 43–56. [Google Scholar] [CrossRef]
- Rym, B.D. Photosynthetic Behavior of Microalgae in Response to Environmental Factors. In Applied Photosynthesis; IntechOpen: London, UK, 2012; ISBN 978-953-51-0061-4. [Google Scholar][Green Version]
- Esteves, A.F.; Salgado, E.M.; Vilar, V.J.P.; Gonçalves, A.L.; Pires, J.C.M. A Growth Phase Analysis on the Influence of Light Intensity on Microalgal Stress and Potential Biofuel Production. Energy Convers. Manag. 2024, 311, 118511. [Google Scholar] [CrossRef]
- Mirkovic, T.; Ostroumov, E.E.; Anna, J.M.; van Grondelle, R.; Govindjee; Scholes, G.D. Light Absorption and Energy Transfer in the Antenna Complexes of Photosynthetic Organisms. Chem. Rev. 2017, 117, 249–293. [Google Scholar] [CrossRef]
- Zhao, L.-S.; Wang, P.; Li, K.; Zhang, Q.-B.; He, F.-Y.; Li, C.-Y.; Su, H.-N.; Chen, X.-L.; Liu, L.-N.; Zhang, Y.-Z. Structural Basis and Evolution of the Photosystem I-Light-Harvesting Supercomplex of Cryptophyte Algae. Plant Cell 2023, 35, 2449–2463. [Google Scholar] [CrossRef] [PubMed]
- Dmitrieva, V.A.; Tyutereva, E.V.; Voitsekhovskaja, O.V. Singlet Oxygen in Plants: Generation, Detection, and Signaling Roles. Int. J. Mol. Sci. 2020, 21, 3237. [Google Scholar] [CrossRef]
- Ramanna, L.; Rawat, I.; Bux, F. Light Enhancement Strategies Improve Microalgal Biomass Productivity. Renew. Sustain. Energy Rev. 2017, 80, 765–773. [Google Scholar] [CrossRef]
- Levin, G.; Yasmin, M.; Liveanu, V.; Burstein, C.; Hanna, R.; Kleifeld, O.; Simanowitz, M.C.; Meir, A.; Tadmor, Y.; Hirschberg, J.; et al. A Desert Chlorella sp. That Thrives at Extreme High-Light Intensities Using a Unique Photoinhibition Protection Mechanism. Plant J. 2023, 115, 510–528. [Google Scholar] [CrossRef]
- Wahidin, S.; Idris, A.; Shaleh, S.R.M. The Influence of Light Intensity and Photoperiod on the Growth and Lipid Content of Microalgae Nannochloropsis sp. Bioresour. Technol. 2013, 129, 7–11. [Google Scholar] [CrossRef]
- Virtanen, O.; Khorobrykh, S.; Tyystjärvi, E. Acclimation of Chlamydomonas Reinhardtii to Extremely Strong Light. Photosynth. Res. 2021, 147, 91–106. [Google Scholar] [CrossRef]
- Remmers, I.M.; Martens, D.E.; Wijffels, R.H.; Lamers, P.P. Dynamics of Triacylglycerol and EPA Production in Phaeodactylum tricornutum under Nitrogen Starvation at Different Light Intensities. PLoS ONE 2017, 12, e0175630. [Google Scholar] [CrossRef]
- Liyanaarachchi, V.C.; Nishshanka, G.K.S.H.; Premaratne, R.G.M.M.; Ariyadasa, T.U.; Nimarshana, P.H.V.; Malik, A. Astaxanthin Accumulation in the Green Microalga Haematococcus pluvialis: Effect of Initial Phosphate Concentration and Stepwise/Continuous Light Stress. Biotechnol. Rep. 2020, 28, e00538. [Google Scholar] [CrossRef]
- Gudvilovich, I.N.; Lelekov, A.S.; Maltsev, E.I.; Kulikovskii, M.S.; Borovkov, A.B. Growth of Porphyridium Purpureum (Porphyridiales, Rhodophyta) and Production of B-Phycoerythrin under Varying Illumination. Russ. J. Plant Physiol. 2021, 68, 188–196. [Google Scholar] [CrossRef]
- Mishra, N.; Prasad, S.M.; Mishra, N. Influence of High Light Intensity and Nitrate Deprivation on Growth and Biochemical Composition of the Marine Microalgae Isochrysis Galbana. Braz. Arch. Biol. Technol. 2019, 62, e19180398. [Google Scholar] [CrossRef]
- Kebede, E.; Ahlgren, G. Optimum Growth Conditions and Light Utilization Efficiency of Spirulina Platensis (=Arthrospira Fusiformis) (Cyanophyta) from Lake Chitu, Ethiopia. Hydrobiologia 1996, 332, 99–109. [Google Scholar] [CrossRef]
- Oostlander, P.C.; van Houcke, J.; Wijffels, R.H.; Barbosa, M.J. Optimization of Rhodomonas sp. under Continuous Cultivation for Industrial Applications in Aquaculture. Algal Res. 2020, 47, 101889. [Google Scholar] [CrossRef]
- Włodarczyk, A.; Selão, T.T.; Norling, B.; Nixon, P.J. Newly Discovered Synechococcus sp. PCC 11901 Is a Robust Cyanobacterial Strain for High Biomass Production. Commun. Biol. 2020, 3, 215. [Google Scholar] [CrossRef]
- Van Wagenen, J.; Miller, T.W.; Hobbs, S.; Hook, P.; Crowe, B.; Huesemann, M. Effects of Light and Temperature on Fatty Acid Production in Nannochloropsis Salina. Energies 2012, 5, 731–740. [Google Scholar] [CrossRef]
- Solovchenko, A.E.; Khozin-Goldberg, I.; Didi-Cohen, S.; Cohen, Z.; Merzlyak, M.N. Effects of Light Intensity and Nitrogen Starvation on Growth, Total Fatty Acids and Arachidonic Acid in the Green Microalga Parietochloris Incisa. J. Appl. Phycol. 2008, 20, 245–251. [Google Scholar] [CrossRef]
- Rise, M.; Cohen, E.; Vishkautsan, M.; Cojocaru, M.; Gottlieb, H.E.; Arad, S. Accumulation of Secondary Carotenoids in Chlorella zofingiensis. J. Plant Physiol. 1994, 144, 287–292. [Google Scholar] [CrossRef]
- Nzayisenga, J.C.; Farge, X.; Groll, S.L.; Sellstedt, A. Effects of Light Intensity on Growth and Lipid Production in Microalgae Grown in Wastewater. Biotechnol. Biofuels 2020, 13, 4. [Google Scholar] [CrossRef]
- Pereira, S.; Otero, A. Haematococcus pluvialis Bioprocess Optimization: Effect of Light Quality, Temperature and Irradiance on Growth, Pigment Content and Photosynthetic Response. Algal Res. 2020, 51, 102027. [Google Scholar] [CrossRef]
- Erdoğan, A.; Karataş, A.; Demirel, Z.; Dalay, M. Induction of Lutein Production in Scenedesmus obliquus under Different Culture Conditions Prior to Its Semipreparative Isolation. Turk. J. Chem. 2022, 46, 796–804. [Google Scholar] [CrossRef]
- Oliveira, C.Y.B.; Oliveira, C.D.L.; Prasad, R.; Ong, H.C.; Araujo, E.S.; Shabnam, N.; Gálvez, A.O. A Multidisciplinary Review of Tetradesmus obliquus: A Microalga Suitable for Large-Scale Biomass Production and Emerging Environmental Applications. Rev. Aquac. 2021, 13, 1594–1618. [Google Scholar] [CrossRef]
- Jallet, D.; Caballero, M.A.; Gallina, A.A.; Youngblood, M.; Peers, G. Photosynthetic Physiology and Biomass Partitioning in the Model Diatom Phaeodactylum tricornutum Grown in a Sinusoidal Light Regime. Algal Res. 2016, 18, 51–60. [Google Scholar] [CrossRef]
- Uwizeye, C.; Decelle, J.; Jouneau, P.-H.; Flori, S.; Gallet, B.; Keck, J.-B.; Bo, D.D.; Moriscot, C.; Seydoux, C.; Chevalier, F.; et al. Morphological Bases of Phytoplankton Energy Management and Physiological Responses Unveiled by 3D Subcellular Imaging. Nat. Commun. 2021, 12, 1049. [Google Scholar] [CrossRef]
- Leister, D. Enhancing the Light Reactions of Photosynthesis: Strategies, Controversies, and Perspectives. Mol. Plant 2023, 16, 4–22. [Google Scholar] [CrossRef]
- Metsoviti, M.N.; Papapolymerou, G.; Karapanagiotidis, I.T.; Katsoulas, N. Effect of Light Intensity and Quality on Growth Rate and Composition of Chlorella vulgaris. Plants 2019, 9, 31. [Google Scholar] [CrossRef]
- Niazkhani, A.; Mohammadi, A.; Mashhadi, H.; Mahmoudnia, F. An Investigation Amount of Cell Density, Biomass, Lipid and Biodiesel Production in Chlorella vulgaris Microalgae under Effect of Different Parameters. Period. Biol. 2022, 124, 1–10. [Google Scholar] [CrossRef]
- Pozzobon, V. Chlorella vulgaris Cultivation under Super High Light Intensity: An Application of the Flashing Light Effect. Algal Res. 2022, 68, 102874. [Google Scholar] [CrossRef]
- Abu-Ghosh, S.; Fixler, D.; Dubinsky, Z.; Iluz, D. Flashing Light in Microalgae Biotechnology. Bioresour. Technol. 2016, 203, 357–363. [Google Scholar] [CrossRef] [PubMed]
- Seyfabadi, J.; Ramezanpour, Z.; Khoeyi, Z.A. Protein, Fatty Acid, and Pigment Content of Chlorella vulgaris under Different Light Regimes. J. Appl. Phycol. 2010, 23, 721–726. [Google Scholar] [CrossRef]
- Simkin, A.J.; Kapoor, L.; Doss, C.G.P.; Hofmann, T.A.; Lawson, T.; Ramamoorthy, S. The Role of Photosynthesis Related Pigments in Light Harvesting, Photoprotection and Enhancement of Photosynthetic Yield in Planta. Photosynth. Res. 2022, 152, 23–42. [Google Scholar] [CrossRef]
- Zarei, Z.; Zamani, H. Biorefinery Approach to Stimulate Astaxanthin and Biofuel Generation in Microalga Haematococcus pluvialis under Different Light Irradiance. Clean. Technol. Environ. Policy 2024, 26, 3333–3347. [Google Scholar] [CrossRef]
- Raqiba, H.; Sibi, G. Light Emitting Diode (LED) Illumination for Enhanced Growth and Cellular Composition in Three Microalgae. Adv. Microbiol. Res. 2019, 3, 7. [Google Scholar] [CrossRef]
- Jungandreas, A.; Schellenberger Costa, B.; Jakob, T.; von Bergen, M.; Baumann, S.; Wilhelm, C. The Acclimation of Phaeodactylum tricornutum to Blue and Red Light Does Not Influence the Photosynthetic Light Reaction but Strongly Disturbs the Carbon Allocation Pattern. PLoS ONE 2014, 9, e99727. [Google Scholar] [CrossRef]
- Ritchie, R.J.; Sma-Air, S. Microalgae Grown under Different Light Sources. J. Appl. Phycol. 2023, 35, 551–566. [Google Scholar] [CrossRef]
- Anyanwu, R.C.; Rodriguez, C.; Durrant, A.; Olabi, A.G. Evaluation of Growth Rate and Biomass Productivity of Scenedesmus Quadricauda and Chlorella vulgaris under Different LED Wavelengths and Photoperiods. Sustainability 2022, 14, 6108. [Google Scholar] [CrossRef]
- Ma, R.; Thomas-Hall, S.R.; Chua, E.T.; Eltanahy, E.; Netzel, M.E.; Netzel, G.; Lu, Y.; Schenk, P.M. Blue Light Enhances Astaxanthin Biosynthesis Metabolism and Extraction Efficiency in Haematococcus pluvialis by Inducing Haematocyst Germination. Algal Res. 2018, 35, 215–222. [Google Scholar] [CrossRef]
- Cray, R.; Levine, I. Oxidative Stress Modulates Astaxanthin Synthesis in Haematococcus pluvialis. J. Appl. Phycol. 2022, 34, 2327–2338. [Google Scholar] [CrossRef]
- Hernández, H.; Nunes, M.C.; Prista, C.; Raymundo, A. Innovative and Healthier Dairy Products through the Addition of Microalgae: A Review. Foods 2022, 11, 755. [Google Scholar] [CrossRef]
- Araújo, R.; Peteiro, C. Algae as Food and Food Supplements in Europe; Publications Office of the EU: Luxembourg, 2021. [Google Scholar] [CrossRef]
- Hlaing, S.A.A.; Sadiq, M.B.; Anal, A.K. Enhanced Yield of Scenedesmus obliquus Biomacromolecules through Medium Optimization and Development of Microalgae Based Functional Chocolate. J. Food Sci. Technol. 2020, 57, 1090–1099. [Google Scholar] [CrossRef]
- Kazir, M.; Livney, Y.D. Plant-Based Seafood Analogs. Molecules 2021, 26, 1559. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, Y.; Chen, L.; Cheng, W.; Liu, T. Combined Production of Fucoxanthin and EPA from Two Diatom Strains Phaeodactylum tricornutum and Cylindrotheca Fusiformis Cultures. Bioprocess. Biosyst. Eng. 2018, 41, 1061–1071. [Google Scholar] [CrossRef] [PubMed]
- Dong, M.; Zhang, Y.; Yu, Q.; Liu, Q.; Zhou, X.; Jin, C. Regulation of Light Quality on Lipid Production, Biodiesel Quality, and Nutritional Quality of Phaeodactylum tricornutum. Aquacult. Int. 2023, 31, 1231–1251. [Google Scholar] [CrossRef]
- Gómez-Loredo, A.; Benavides, J.; Rito-Palomares, M. Growth Kinetics and Fucoxanthin Production of Phaeodactylum tricornutum and Isochrysis Galbana Cultures at Different Light and Agitation Conditions. J. Appl. Phycol. 2016, 28, 849–860. [Google Scholar] [CrossRef]
- Yi, Z.; Su, Y.; Cherek, P.; Nelson, D.R.; Lin, J.; Rolfsson, O.; Wu, H.; Salehi-Ashtiani, K.; Brynjolfsson, S.; Fu, W. Combined Artificial High-Silicate Medium and LED Illumination Promote Carotenoid Accumulation in the Marine Diatom Phaeodactylum tricornutum. Microb. Cell Fact. 2019, 18, 209. [Google Scholar] [CrossRef] [PubMed]
- Jin, C.; Zhou, Z.; Zhu, Y.; Liu, Q.; Zhou, X. Photosynthetic Activity and Astaxanthin Production of Haematococcus pluvialis Regulated by Manipulated Light Quality. Aquacult. Int. 2024, 32, 3617–3635. [Google Scholar] [CrossRef]
- Zhang, W.; Zhou, X.; Zhang, Y.; Cheng, P.; Ma, R.; Cheng, W.; Chu, H. Enhancing Astaxanthin Accumulation in Haematococcus pluvialis by Coupled Light Intensity and Nitrogen Starvation in Column Photobioreactors. J. Microbiol. Biotechnol. 2018, 28, 2019–2028. [Google Scholar] [CrossRef]
- Wu, R.A.; Ding, Q.; Yin, L.; Chi, X.; Sun, N.; He, R.; Luo, L.; Ma, H.; Li, Z. Comparison of the Nutritional Value of Mysore Thorn Borer (Anoplophora chinensis) and Mealworm Larva (Tenebrio molitor): Amino Acid, Fatty Acid, and Element Profiles. Food Chem. 2020, 323, 126818. [Google Scholar] [CrossRef]
- Manzoor, M.F.; Shabbir, U.; Gilani, S.M.; Sameen, A.; Ahmad, N.; Siddique, R.; Ahmed, Z.; Qayyum, A.; Rehman, A. Characterization of Bioactive Fatty Acids and Oxidative Stability of Microwave Vacuum Dried Fish Powder Supplemented Extruded Product. Food Sci. Technol. 2021, 42, e22720. [Google Scholar] [CrossRef]
- Munialo, C.D.; Vriesekoop, F. Plant-Based Foods as Meat and Fat Substitutes. Food Sci. Nutr. 2023, 11, 4898–4911. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Wen, S.; Ye, Q.; Lou, H.; Gao, Y.; Bajpai, V.K.; Carpena, M.; Prieto, M.-A.; Simal-Gandara, J.; Xiao, J.; et al. Advances on Delta 5-Unsaturated-Polymethylene-Interrupted Fatty Acids: Resources, Biosynthesis, and Benefits. Crit. Rev. Food Sci. Nutr. 2021, 63, 767–789. [Google Scholar] [CrossRef] [PubMed]
- Jesionowska, M.; Ovadia, J.; Hockemeyer, K.; Clews, A.C.; Xu, Y. EPA and DHA in Microalgae: Health Benefits, Biosynthesis, and Metabolic Engineering Advances. J. Am. Oil Chem. Soc. 2023, 100, 831–842. [Google Scholar] [CrossRef]
- Maréchal, E.; Lupette, J. Relationship between Acyl-Lipid and Sterol Metabolisms in Diatoms. Biochimie 2020, 169, 3–11. [Google Scholar] [CrossRef]
- Celi, C.; Fino, D.; Savorani, F. Phaeodactylum tricornutum as a Source of Value-Added Products: A Review on Recent Developments in Cultivation and Extraction Technologies. Bioresour. Technol. Rep. 2022, 19, 101122. [Google Scholar] [CrossRef]
- Butler, T.; Kapoore, R.V.; Vaidyanathan, S. Phaeodactylum tricornutum: A Diatom Cell Factory. Trends Biotechnol. 2020, 38, 606–622. [Google Scholar] [CrossRef]
- Cui, Y.; Thomas-Hall, S.R.; Chua, E.T.; Schenk, P.M. Development of High-Level Omega-3 Eicosapentaenoic Acid (EPA) Production from Phaeodactylum tricornutum. J. Phycol. 2021, 57, 258–268. [Google Scholar] [CrossRef]
- Conceição, D.; Lopes, R.G.; Derner, R.B.; Cella, H.; do Carmo, A.P.B.; Montes D’Oca, M.G.; Petersen, R.; Passos, M.F.; Vargas, J.V.C.; Galli-Terasawa, L.V.; et al. The Effect of Light Intensity on the Production and Accumulation of Pigments and Fatty Acids in Phaeodactylum tricornutum. J. Appl. Phycol. 2020, 32, 1017–1025. [Google Scholar] [CrossRef]
- Sirisuk, P.; Ra, C.-H.; Jeong, G.-T.; Kim, S.-K. Effects of Wavelength Mixing Ratio and Photoperiod on Microalgal Biomass and Lipid Production in a Two-Phase Culture System Using LED Illumination. Bioresour. Technol. 2018, 253, 175–181. [Google Scholar] [CrossRef]
- Peng, M.; Lin, S.; Shen, Y.; Peng, R.; Li, S.; Jiang, X.; Jiang, M. Effects of Light Quality on the Growth, Productivity, Fucoxanthin Accumulation, and Fatty Acid Composition of Thalassiosira Pseudonana. J. Appl. Phycol. 2024, 36, 1667–1678. [Google Scholar] [CrossRef]
- Liu, Y.; Qiao, Z.; Liu, W.; Hou, Z.; Zhang, D.; Huang, L.; Zhang, Y. Oleic Acid as a Protein Ligand Improving Intestinal Absorption and Ocular Benefit of Fucoxanthin in Water through Protein-Based Encapsulation. Food Funct. 2019, 10, 4381–4395. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.; Cassani, L.; Grosso, C.; Garcia-Oliveira, P.; Morais, S.L.; Echave, J.; Carpena, M.; Xiao, J.; Barroso, M.F.; Simal-Gandara, J.; et al. Recent Advances in Biological Properties of Brown Algae-Derived Compounds for Nutraceutical Applications. Crit. Rev. Food Sci. Nutr. 2022, 64, 1283–1311. [Google Scholar] [CrossRef]
- Bae, M.; Kim, M.-B.; Park, Y.-K.; Lee, J.-Y. Health Benefits of Fucoxanthin in the Prevention of Chronic Diseases. Biochim. Et. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2020, 1865, 158618. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Yang, S.; Zhao, W.; Kong, Q.; Zhu, C.; Fu, X.; Zhang, F.; Liu, Z.; Zhan, Y.; Mou, H.; et al. Fucoxanthin from Marine Microalgae: A Promising Bioactive Compound for Industrial Production and Food Application. Crit. Rev. Food Sci. Nutr. 2023, 63, 7996–8012. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, E.A.; El-Sayed, S.M.; Murad, S.A.; Abdallah, W.E.; El-Sayed, H.S. Evaluation of the Antioxidant and Antimicrobial Activities of Fucoxanthin from Dilophys Fasciola and as a Food Additive in Stirred Yoghurt. S. Afr. J. Sci. 2023, 119, 1–9. [Google Scholar] [CrossRef]
- Wang, S.; Wu, S.; Yang, G.; Pan, K.; Wang, L.; Hu, Z. A Review on the Progress, Challenges and Prospects in Commercializing Microalgal Fucoxanthin. Biotechnol. Adv. 2021, 53, 107865. [Google Scholar] [CrossRef]
- McClure, D.D.; Luiz, A.; Gerber, B.; Barton, G.W.; Kavanagh, J.M. An Investigation into the Effect of Culture Conditions on Fucoxanthin Production Using the Marine Microalgae Phaeodactylum tricornutum. Algal Res. 2018, 29, 41–48. [Google Scholar] [CrossRef]
- Truong, T.Q.; Park, Y.J.; Koo, S.Y.; Choi, J.-H.; Enkhbayar, A.; Song, D.-G.; Kim, S.M. Interdependence of Fucoxanthin Biosynthesis and Fucoxanthin-Chlorophyll a/c Binding Proteins in Phaeodactylum tricornutum under Different Light Intensities. J. Appl. Phycol. 2023, 35, 25–42. [Google Scholar] [CrossRef]
- Sharma, A.K.; Nymark, M.; Flo, S.; Sparstad, T.; Bones, A.M.; Winge, P. Simultaneous Knockout of Multiple LHCF Genes Using Single sgRNAs and Engineering of a High-Fidelity Cas9 for Precise Genome Editing in Marine Algae. Plant Biotechnol. J. 2021, 19, 1658–1669. [Google Scholar] [CrossRef]
- Wang, L.; Xie, X.; Gu, W.; Zheng, Z.; Chen, M.; Wang, G. LHCF15 Facilitates the Absorption of Longer Wavelength Light and Promotes Growth of Phaeodactylum tricornutum under Red Light. Algal Res. 2023, 75, 103249. [Google Scholar] [CrossRef]
- Song, Q.; Liu, C.; Xu, R.; Cai, L. Enhancement of Fucoxanthin Accumulation in Phaeodactylum tricornutum by Light Quality and Intensity Shift Strategy. Chem. Eng. J. 2025, 505, 159388. [Google Scholar] [CrossRef]
- Zhang, H.; Gong, P.; Cai, Q.; Zhang, C.; Gao, B. Maximizing Fucoxanthin Production in Odontella Aurita by Optimizing the Ratio of Red and Blue Light-Emitting Diodes in an Auto-Controlled Internally Illuminated Photobioreactor. Bioresour. Technol. 2022, 344, 126260. [Google Scholar] [CrossRef]
- Böcker, L.; Bertsch, P.; Wenner, D.; Teixeira, S.; Bergfreund, J.; Eder, S.; Fischer, P.; Mathys, A. Effect of Arthrospira platensis microalgae protein purification on emulsification mechanism and efficiency. J. Colloid Interface Sci. 2021, 584, 344–353. [Google Scholar] [CrossRef] [PubMed]
- Williams, P.J.l.B.; Laurens, L.M.L. Microalgae as Biodiesel & Biomass Feedstocks: Review & Analysis of the Biochemistry, Energetics & Economics. Energy Environ. Sci. 2010, 3, 554–590. [Google Scholar] [CrossRef]
- Caporgno, M.P.; Böcker, L.; Müssner, C.; Stirnemann, E.; Haberkorn, I.; Adelmann, H.; Handschin, S.; Windhab, E.J.; Mathys, A. Extruded Meat Analogues Based on Yellow, Heterotrophically Cultivated AuxenoChlorella protothecoides Microalgae. Innov. Food Sci. Emerg. Technol. 2020, 59, 102275. [Google Scholar] [CrossRef]
- Fu, Y.; Chen, T.; Chen, S.H.Y.; Liu, B.; Sun, P.; Sun, H.; Chen, F. The Potentials and Challenges of Using Microalgae as an Ingredient to Produce Meat Analogues. Trends Food Sci. Technol. 2021, 112, 188–200. [Google Scholar] [CrossRef]
- Shah, M.A.R.; Zhu, F.; Cui, Y.; Hu, X.; Chen, H.; Kayani, S.-I.; Huo, S. Mechanistic Insights into the Nutritional and Therapeutic Potential of Spirulina (Arthrospira) spp.: Challenges and Opportunities. Trends Food Sci. Technol. 2024, 151, 104648. [Google Scholar] [CrossRef]
- Shao, W.; Ebaid, R.; El-Sheekh, M.; Abomohra, A.; Eladel, H. Pharmaceutical Applications and Consequent Environmental Impacts of Spirulina (Arthrospira): An Overview. Grasas Aceites 2019, 70, e292. [Google Scholar] [CrossRef]
- Wang, F.; Yu, X.; Cui, Y.; Xu, L.; Huo, S.; Ding, Z.; Hu, Q.; Xie, W.; Xiao, H.; Zhang, D. Efficient Extraction of Phycobiliproteins from Dry Biomass of Spirulina platensis Using Sodium Chloride as Extraction Enhancer. Food Chem. 2023, 406, 135005. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, X.; Li, Y.; Liu, S.; Chen, Y.; Jia, M.; Wang, X.; Zhang, L.; Gao, Q.; Zhang, L.; et al. Regulation of Different Light Conditions for Efficient Biomass Production and Protein Accumulation of Spirulina Platensis. J. Oceanol. Limnol. 2024, 42, 174–186. [Google Scholar] [CrossRef]
- Bhat, O.; Unpaprom, Y.; Ramaraj, R. Effect of Photoperiod and White LED on Biomass Growth and Protein Production by Spirulina. Mol. Biotechnol. 2023. [Google Scholar] [CrossRef]
- Bhat, O.; Unpaprom, Y.; Ramaraj, R. Spirulina Cultivation Under Different Light-Emitting Diodes for Boosting Biomass and Protein Production. Mol. Biotechnol. 2023. [Google Scholar] [CrossRef]
- Yan, B.; Chen, X.; Wang, Y.; Yuan, M.; Xian, J.; Lu, D.; Shao, Z.; Qiu, M.; Fu, T.; Zheng, X. Chlorella pyrenoidosa Ameliorates Ulcerative Colitis by Tuning Intestinal Microecology: Butyric Acid Is a Crucial Player. J. Funct. Foods 2024, 121, 106414. [Google Scholar] [CrossRef]
- Metsoviti, M.N.; Katsoulas, N.; Karapanagiotidis, I.T.; Papapolymerou, G. Effect of Nitrogen Concentration, Two-Stage and Prolonged Cultivation on Growth Rate, Lipid and Protein Content of Chlorella vulgaris. J. Chem. Technol. Biotechnol. 2019, 94, 1466–1473. [Google Scholar] [CrossRef]
- Atta, M.; Idris, A.; Bukhari, A.; Wahidin, S. Intensity of Blue LED Light: A Potential Stimulus for Biomass and Lipid Content in Fresh Water Microalgae Chlorella vulgaris. Bioresour. Technol. 2013, 148, 373–378. [Google Scholar] [CrossRef]
- Sui, Y.; Harvey, P.J. Effect of Light Intensity and Wavelength on Biomass Growth and Protein and Amino Acid Composition of Dunaliella salina. Foods 2021, 10, 1018. [Google Scholar] [CrossRef]
- Elbahnaswy, S.; Elshopakey, G.E. Recent Progress in Practical Applications of a Potential Carotenoid Astaxanthin in Aquaculture Industry: A Review. Fish Physiol. Biochem. 2024, 50, 97–126. [Google Scholar] [CrossRef]
- Abuzar; Sharif, H.R.; Sharif, M.K.; Arshad, R.; Rehman, A.; Ashraf, W.; Karim, A.; Awan, K.A.; Raza, H.; Khalid, W.; et al. Potential Industrial and Nutritional Applications of Shrimp By-Products: A Review. Int. J. Food Prop. 2023, 26, 3407–3432. [Google Scholar] [CrossRef]
- Xie, X.; Zhong, M.; Huang, X.; Yuan, X.; Mahna, N.; Mussagy, C.U.; Ren, M. Astaxanthin Biosynthesis for Functional Food Development and Space Missions. Crit. Rev. Biotechnol. 2024, 45, 923–937. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Wang, Y.; Zhu, L.; Zhao, X. Effects of Haematococcus pluvialis Addition on the Sensory Properties of Plant-Based Meat Analogues. Foods 2023, 12, 3435. [Google Scholar] [CrossRef]
- Samhat, K.; Kazbar, A.; Takache, H.; Ismail, A.; Pruvost, J. Influence of Light Absorption Rate on the Astaxanthin Production by the Microalga Haematococcus pluvialis during Nitrogen Starvation. Bioresour. Bioprocess. 2023, 10, 78. [Google Scholar] [CrossRef]
- Morgado, D.; Fanesi, A.; Martin, T.; Tebbani, S.; Bernard, O.; Lopes, F. Exploring the Dynamics of Astaxanthin Production in Haematococcus pluvialis Biofilms Using a Rotating Biofilm-Based System. Biotechnol. Bioeng. 2023, 121, 991–1004. [Google Scholar] [CrossRef]
- Rizzo, A.; Ross, M.E.; Norici, A.; Jesus, B. A Two-Step Process for Improved Biomass Production and Non-Destructive Astaxanthin and Carotenoids Accumulation in Haematococcus pluvialis. Appl. Sci. 2022, 12, 1261. [Google Scholar] [CrossRef]
- Ahirwar, A.; Meignen, G.; Jahir Khan, M.; Sirotiya, V.; Harish; Scarsini, M.; Roux, S.; Marchand, J.; Schoefs, B.; Vinayak, V. Light Modulates Transcriptomic Dynamics Upregulating Astaxanthin Accumulation in Haematococcus: A Review. Bioresour. Technol. 2021, 340, 125707. [Google Scholar] [CrossRef] [PubMed]
- Cifuentes, A.S.; González, M.A.; Vargas, S.; Hoeneisen, M.; González, N. Optimization of Biomass, Total Carotenoids and Astaxanthin Production in Haematococcus pluvialis Flotow Strain Steptoe (Nevada, USA) under Laboratory Conditions. Biol. Res. 2003, 36, 343–357. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Cui, D.; Sun, X.; Shi, J.; Xu, N. Primary Metabolism Is Associated with the Astaxanthin Biosynthesis in the Green Algae Haematococcus pluvialis under Light Stress. Algal Res. 2020, 46, 101768. [Google Scholar] [CrossRef]
- Boussiba, S. Carotenogenesis in the Green Alga Haematococcus pluvialis: Cellular Physiology and Stress Response. Physiol. Plant. 2000, 108, 111–117. [Google Scholar] [CrossRef]
- Huang, L.; Gao, B.; Wu, M.; Wang, F.; Zhang, C. Comparative Transcriptome Analysis of a Long-Time Span Two-Step Culture Process Reveals a Potential Mechanism for Astaxanthin and Biomass Hyper-Accumulation in Haematococcus pluvialis JNU35. Biotechnol. Biofuels 2019, 12, 18. [Google Scholar] [CrossRef] [PubMed]
- He, B.; Hou, L.; Dong, M.; Shi, J.; Huang, X.; Ding, Y.; Cong, X.; Zhang, F.; Zhang, X.; Zang, X. Transcriptome Analysis in Haematococcus pluvialis: Astaxanthin Induction by High Light with Acetate and Fe2+. Int. J. Mol. Sci. 2018, 19, 175. [Google Scholar] [CrossRef]
- Gwak, Y.; Hwang, Y.; Wang, B.; Kim, M.; Jeong, J.; Lee, C.-G.; Hu, Q.; Han, D.; Jin, E. Comparative Analyses of Lipidomes and Transcriptomes Reveal a Concerted Action of Multiple Defensive Systems against Photooxidative Stress in Haematococcus pluvialis. J. Exp. Bot. 2014, 65, 4317–4334. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Yong, H.I.; Kim, M.; Choi, Y.-S.; Jo, C. Status of Meat Alternatives and Their Potential Role in the Future Meat Market—A Review. Asian-Australas. J. Anim. Sci. 2020, 33, 1533–1543. [Google Scholar] [CrossRef]
- Kim, T.-K.; Yong, H.I.; Cha, J.Y.; Park, S.-Y.; Jung, S.; Choi, Y.-S. Drying-Induced Restructured Jerky Analog Developed Using a Combination of Edible Insect Protein and Textured Vegetable Protein. Food Chem. 2022, 373, 131519. [Google Scholar] [CrossRef] [PubMed]
- Han, J.-H.; Keum, D.-H.; Hong, S.-J.; Kim, Y.-J.; Han, S.-G. Comparative Evaluation of Polysaccharide Binders on the Quality Characteristics of Plant-Based Patties. Foods 2023, 12, 3731. [Google Scholar] [CrossRef]
- Ouyang, Y.; Qiu, Y.; Liu, Y.; Zhu, R.; Chen, Y.; El-Seedi, H.R.; Chen, X.; Zhao, C. Cancer-Fighting Potentials of Algal Polysaccharides as Nutraceuticals. Food Res. Int. 2021, 147, 110522. [Google Scholar] [CrossRef]
- Wang, M.; Yin, Z.; Zeng, M. Microalgae as a Promising Structure Ingredient in Food: Obtained by Simple Thermal and High-Speed Shearing Homogenization. Food Hydrocoll. 2022, 131, 107743. [Google Scholar] [CrossRef]
- Guo, W.; Zhu, S.; Li, S.; Feng, Y.; Wu, H.; Zeng, M. Microalgae Polysaccharides Ameliorates Obesity in Association with Modulation of Lipid Metabolism and Gut Microbiota in High-Fat-Diet Fed C57BL/6 Mice. Int. J. Biol. Macromol. 2021, 182, 1371–1383. [Google Scholar] [CrossRef]
- Frick, K.; Ebbing, T.; Yeh, Y.-C.; Schmid-Staiger, U.; Tovar, G.E.M. Influence of Light Conditions on the Production of Chrysolaminarin Using Phaeodactylum tricornutum in Artificially Illuminated Photobioreactors. MicrobiologyOpen 2023, 12, e1378. [Google Scholar] [CrossRef]
- González-Fernández, C.; Ballesteros, M. Linking Microalgae and Cyanobacteria Culture Conditions and Key-Enzymes for Carbohydrate Accumulation. Biotechnol. Adv. 2012, 30, 1655–1661. [Google Scholar] [CrossRef] [PubMed]
- Xia, S.; Gao, B.; Li, A.; Xiong, J.; Ao, Z.; Zhang, C. Preliminary Characterization, Antioxidant Properties and Production of Chrysolaminarin from Marine Diatom Odontella Aurita. Mar. Drugs 2014, 12, 4883–4897. [Google Scholar] [CrossRef]
- Ran, X.; Shen, Y.; Jiang, D.; Wang, C.; Li, X.; Zhang, H.; Pan, Y.; Xie, C.; Xie, T.; Zhang, Y.; et al. Nutrient Deprivation Coupled with High Light Exposure for Bioactive Chrysolaminarin Production in the Marine Microalga Isochrysis Zhangjiangensis. Mar. Drugs 2022, 20, 351. [Google Scholar] [CrossRef]
- Schulze, C.; Wetzel, M.; Reinhardt, J.; Schmidt, M.; Felten, L.; Mundt, S. Screening of Microalgae for Primary Metabolites Including β-Glucans and the Influence of Nitrate Starvation and Irradiance on β-Glucan Production. J. Appl. Phycol. 2016, 28, 2719–2725. [Google Scholar] [CrossRef]
- Arumugam, M.; Agarwal, A.; Arya, M.C.; Ahmed, Z. Influence of Nitrogen Sources on Biomass Productivity of Microalgae Scenedesmus bijugatus. Bioresour. Technol. 2013, 131, 246–249. [Google Scholar] [CrossRef] [PubMed]
- Navarro, F.E.; Damiani, M.C.; Leonardi, P.I.; Popovich, C.A. Temperature and Salinity Effect on Tolerance and Lipid Accumulation in Halamphora Coffeaeformis: An Approach for Outdoor Bioenergy Cultures. Bioenerg. Res. 2022, 15, 1545–1554. [Google Scholar] [CrossRef]
- Zhou, J.; Li, P.; Wang, J.; Fu, W. Growth, Photosynthesis, and Nutrient Uptake at Different Light Intensities and Temperatures in Lettuce. HortScience 2019, 54, 1925–1933. [Google Scholar] [CrossRef]
- Terada, R.; Shindo, A.; Moriyama, H.; Shimboku, N.; Nishihara, G.N. The Response of Photosynthesis to Temperature and Irradiance in a Green Alga, Ryuguphycus kuaweuweu (Ulvales) Reveals Adaptation to a Subtidal Environment in the Northern Ryukyu Islands. Algal Res. 2023, 74, 103189. [Google Scholar] [CrossRef]
- Ova Ozcan, D.; Ovez, B. Evaluation of the Interaction of Temperature and Light Intensity on the Growth of Phaeodactylum tricornutum: Kinetic Modeling and Optimization. Biochem. Eng. J. 2020, 154, 107456. [Google Scholar] [CrossRef]
- Cella, H.; Nader, C.; Bastolla, C.L.V.; Bonomi-Barufi, J.; Oliveira, C.Y.B.; Lopes, R.G.; Mattos, J.J.; Bauer, C.M.; Maraschin, M.; Rörig, L.R.; et al. PAR Regulation of Photoprotection in Phaeodactylum tricornutum (Bacillariophyceae): Roles of Doses and Irradiances. J. Appl. Phycol. 2023, 35, 2177–2191. [Google Scholar] [CrossRef]
- Şirin, P.A.; Serdar, S. Effects of Nitrogen Starvation on Growth and Biochemical Composition of Some Microalgae Species. Folia Microbiol. 2024, 69, 889–902. [Google Scholar] [CrossRef] [PubMed]
- Scibilia, L.; Girolomoni, L.; Berteotti, S.; Alboresi, A.; Ballottari, M. Photosynthetic Response to Nitrogen Starvation and High Light in Haematococcus pluvialis. Algal Res. 2015, 12, 170–181. [Google Scholar] [CrossRef]
- Li, F.; Cai, M.; Wu, Y.; Lian, Q.; Qian, Z.; Luo, J.; Zhang, Y.; Zhang, N.; Li, C.; Huang, X. Effects of Nitrogen and Light Intensity on the Astaxanthin Accumulation in Motile Cells of Haematococcus pluvialis. Front. Mar. Sci. 2022, 9. [Google Scholar] [CrossRef]
- Li, Q.; Li, L.; Zhang, Y.; Gao, H.; Zhao, Y.; Yu, X. Chemical Inducers Regulate ROS Signalling to Stimulate Astaxanthin Production in Haematococcus pluvialis under Environmental Stresses: A Review. Trends Food Sci. Technol. 2023, 136, 181–193. [Google Scholar] [CrossRef]
- Wong, F.-C.; Xiao, J.; Wang, S.; Ee, K.-Y.; Chai, T.-T. Advances on the Antioxidant Peptides from Edible Plant Sources. Trends Food Sci. Technol. 2020, 99, 44–57. [Google Scholar] [CrossRef]
- Zhao, Y.; Xing, H.; Li, X.; Geng, S.; Ning, D.; Ma, T.; Yu, X. Physiological and Metabolomics Analyses Reveal the Roles of Fulvic Acid in Enhancing the Production of Astaxanthin and Lipids in Haematococcus pluvialis under Abiotic Stress Conditions. J. Agric. Food Chem. 2019, 67, 12599–12609. [Google Scholar] [CrossRef] [PubMed]
- Acheampong, A.; Wang, R.; Elsherbiny, S.M.; Bondzie-Quaye, P.; Huang, Q. Exogenous Arginine Promotes the Coproduction of Biomass and Astaxanthin under High-Light Conditions in Haematococcus pluvialis. Bioresour. Technol. 2024, 393, 130001. [Google Scholar] [CrossRef]
- Bo, Y.; Wang, S.; Ma, F.; Yurevich Manyakhin, A.; Zhang, G.; Li, X.; Zhou, C.; Ge, B.; Yan, X.; Ruan, R.; et al. The Influence of Spermidine on the Build-up of Fucoxanthin in Isochrysis sp. Acclimated to Varying Light Intensities. Bioresour. Technol. 2023, 387, 129688. [Google Scholar] [CrossRef]
- Ferreira de Oliveira, A.P.; Bragotto, A.P.A. Microalgae-Based Products: Food and Public Health. Future Foods 2022, 6, 100157. [Google Scholar] [CrossRef]
- European Parliament and Council of the European Union. Regulation (EU) 2015/2283 of the European Parliament and of the Council of 25 November 2015 on Novel Foods, Amending Regulation (EU) No 1169/2011 of the European Parliament and of the Council and Repealing Regulation (EC) No 258/97 of the European Parliament and of the Council and Commission Regulation (EC) No 1852/2001; Official Journal of the European Union L 327/1; European Parliament and Council of the European Union: Strasbourg, France, 2015. [Google Scholar]
- Martín-Girela, I.; Albero, B.; Tiwari, B.K.; Miguel, E.; Aznar, R. Screening of Contaminants of Emerging Concern in Microalgae Food Supplements. Separations 2020, 7, 28. [Google Scholar] [CrossRef]
- Nethravathy, M.U.; Mehar, J.G.; Mudliar, S.N.; Shekh, A.Y. Recent Advances in Microalgal Bioactives for Food, Feed, and Healthcare Products: Commercial Potential, Market Space, and Sustainability. Compr. Rev. Food Sci. Food Saf. 2019, 18, 1882–1897. [Google Scholar] [CrossRef] [PubMed]
- Kozai, T. Benefits, Problems and Challenges of Plant Factories with Artificial Lighting (PFALs): A Short Review. Acta Hortic. 2018, 25–30. [Google Scholar] [CrossRef]
- Teng, S.Y.; Yew, G.Y.; Sukačová, K.; Show, P.L.; Máša, V.; Chang, J.-S. Microalgae with Artificial Intelligence: A Digitalized Perspective on Genetics, Systems and Products. Biotechnol. Adv. 2020, 44, 107631. [Google Scholar] [CrossRef] [PubMed]
- Imamoglu, E. Artificial Intelligence and/or Machine Learning Algorithms in Microalgae Bioprocesses. Bioengineering 2024, 11, 1143. [Google Scholar] [CrossRef]
- Kushwaha, O.S.; Uthayakumar, H.; Kumaresan, K. Modeling of Carbon Dioxide Fixation by Microalgae Using Hybrid Artificial Intelligence (AI) and Fuzzy Logic (FL) Methods and Optimization by Genetic Algorithm (GA). Environ. Sci. Pollut. Res. 2023, 30, 24927–24948. [Google Scholar] [CrossRef]
- Lim, H.R.; Khoo, K.S.; Chia, W.Y.; Chew, K.W.; Ho, S.-H.; Show, P.L. Smart Microalgae Farming with Internet-of-Things for Sustainable Agriculture. Biotechnol. Adv. 2022, 57, 107931. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).