
Edible Insects as a Novel Source of Bioactive Peptides: A Systematic Review

 ,
,  ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
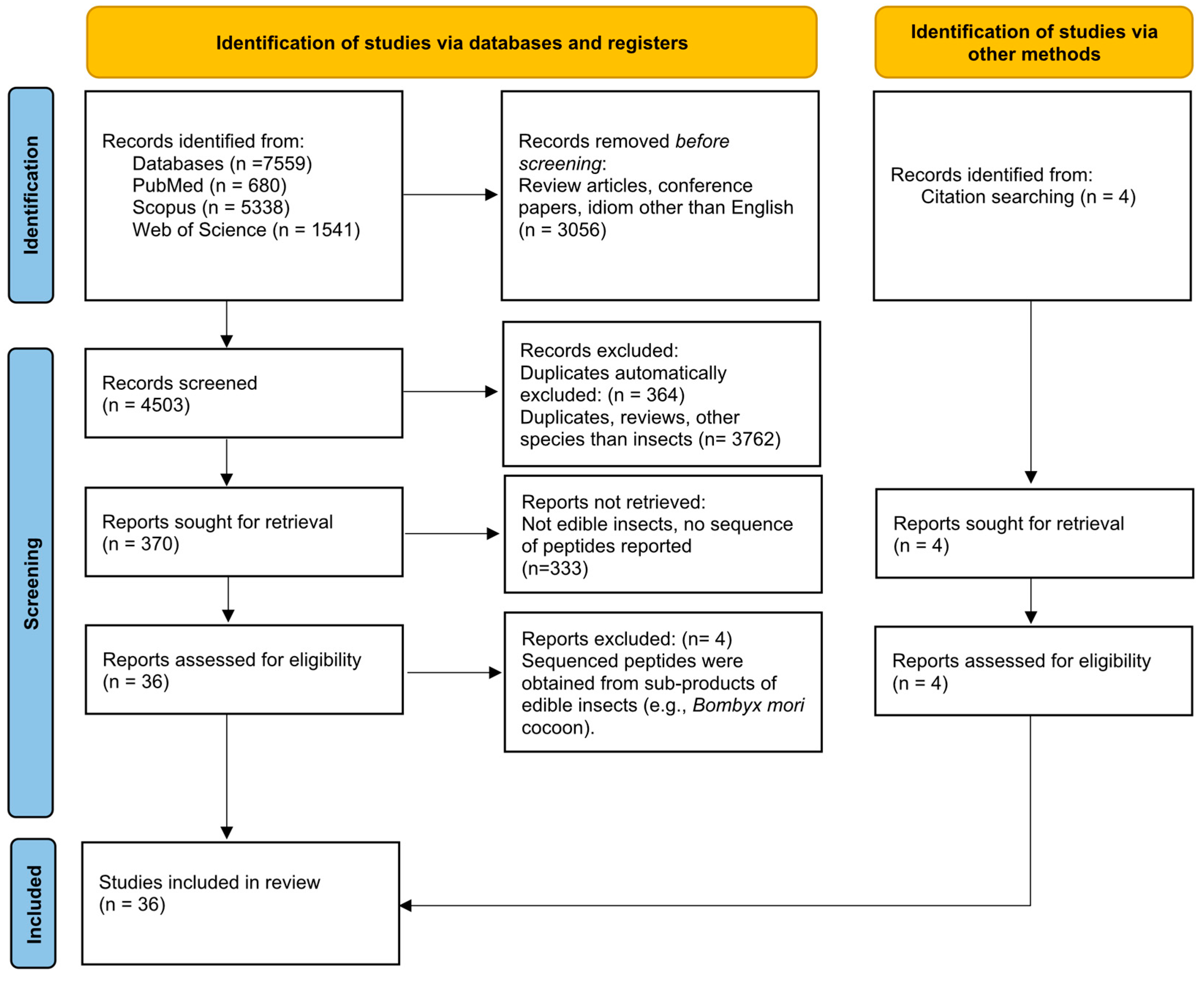
2.1. Search Strategy
2.2. Exclusion Criteria and Results
2.3. Data Extraction
3. Results
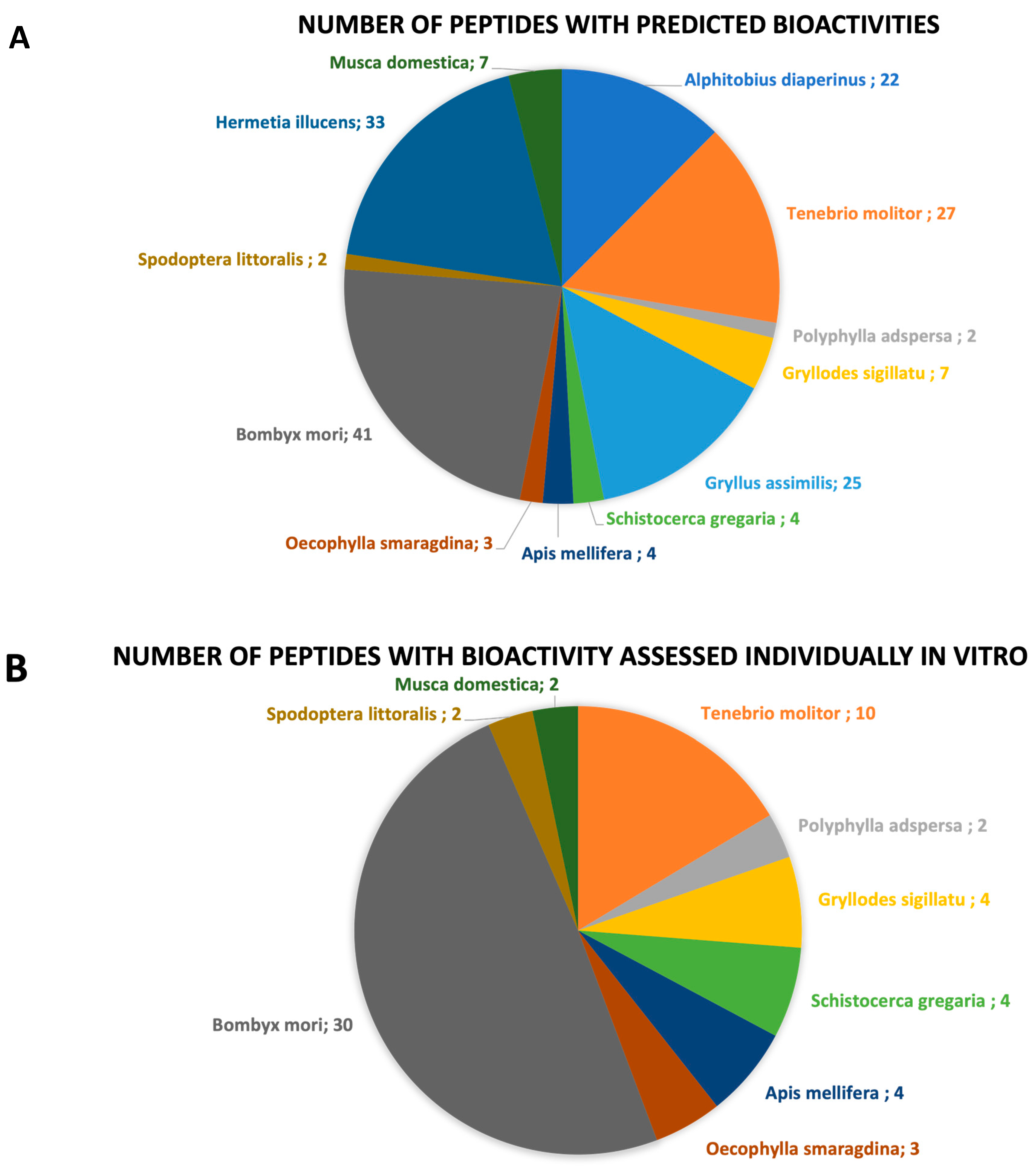
3.1. Species of Edible Insects with Sequenced Bioactive Peptides
3.1.1. Gryllodes sigillatus
3.1.2. Gryllus assimilis
3.1.3. Schistocerca gregaria
3.1.4. Alphitobius diaperinus
3.1.5. Tenebrio molitor
3.1.6. Polyphylla adspersa
3.1.7. Apis mellifera
3.1.8. Oecophylla smaragdina
3.1.9. Bombyx mori
3.1.10. Spodoptera littoralis
3.1.11. Hermetia illucens
3.1.12. Musca domestica
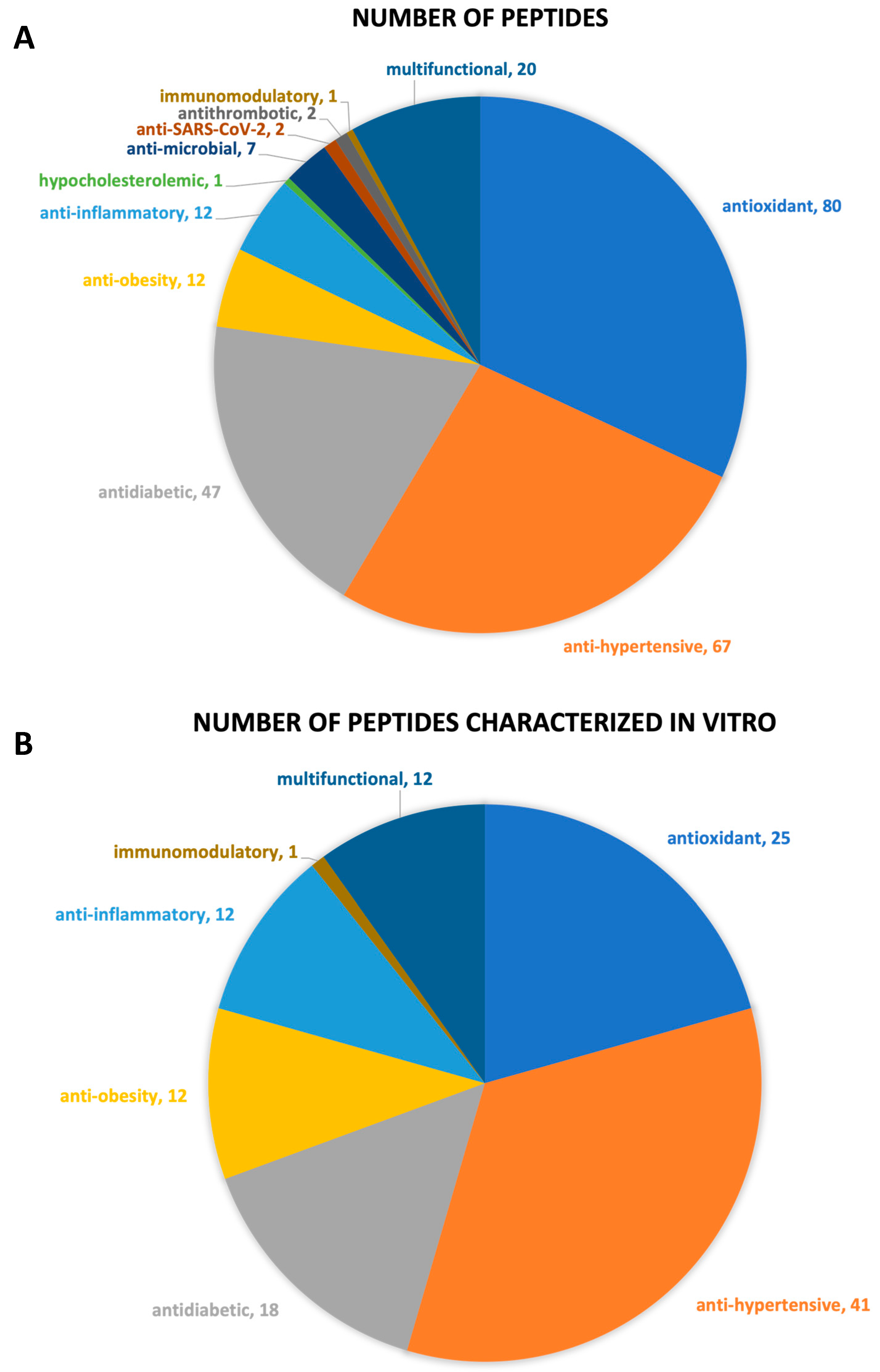
3.2. Bioactive Properties of the Identified Peptides
3.2.1. Antioxidant Peptides
3.2.2. Antihypertensive Peptides
3.2.3. Antidiabetic Peptides
3.2.4. Other Bioactive Peptides
4. Conclusions and Future Perspectives
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Meyer-Rochow, V.B. Can Insects Help to Ease Problem of World Food Shortage? Search 1975, 6, 261–262. [Google Scholar]
- Ramos-Elorduy, J. Anthropo-entomophagy: Cultures, evolution and sustainability. Entomol. Res. 2009, 39, 271–288. [Google Scholar] [CrossRef]
- Stork, N.E. How Many Species of Insects and Other Terrestrial Arthropods Are There on Earth? Annu. Rev. Entomol. 2018, 63, 31–45. [Google Scholar] [CrossRef] [PubMed]
- Kouřimská, L.; Adámková, A. Nutritional and sensory quality of edible insects. NFS J. 2016, 4, 22–26. [Google Scholar] [CrossRef]
- Xiaoming, C.; Ying, F.; Hong, Z.; Zhiyong, C. Review of the nutritive value of edible insects. In Forest Insects as Food: Humans Bite Back; Durst, P.B., Johnson, D.V., Leslie, R.N.S.K., Eds.; Food and Agriculture Organization of the United Nations: Rome, Italy, 2010; pp. 85–92. [Google Scholar]
- Ramos-Elorduy, J.; Moreno, J.M.P.; Prado, E.E.; Perez, M.A.; Otero, J.L.; de Guevara, O.L. Nutritional Value of Edible Insects from the State of Oaxaca, Mexico. J. Food Compos. Anal. 1997, 10, 142–157. [Google Scholar] [CrossRef]
- Quah, Y.; Tong, S.-R.; Bojarska, J.; Giller, K.; Tan, S.-A.; Ziora, Z.M.; Esatbeyoglu, T.; Chai, T.-T. Bioactive Peptide Discovery from Edible Insects for Potential Applications in Human Health and Agriculture. Molecules 2023, 28, 1233. [Google Scholar] [CrossRef]
- Vercruysse, L.; Van Camp, J.; Morel, N.; Rouge, P.; Herregods, G.; Smagghe, G. Ala-Val-Phe and Val-Phe: ACE inhibitory peptides derived from insect protein with antihypertensive activity in spontaneously hypertensive rats. Peptides 2010, 31, 482–488. [Google Scholar] [CrossRef]
- Rivero-Pino, F.; Guadix, A.; Guadix, E.M. Identification of novel dipeptidyl peptidase IV and α-glucosidase inhibitory peptides from Tenebrio molitor. Food Funct. 2021, 12, 873–880. [Google Scholar] [CrossRef]
- Zhu, D.; Huang, X.; Tu, F.; Wang, C.; Yang, F. Preparation, antioxidant activity evaluation, and identification of antioxidant peptide from black soldier fly (Hermetia illucens L.) larvae. J. Food Biochem. 2020, 44, e13186. [Google Scholar] [CrossRef]
- Zielińska, E.; Karaś, M.; Baraniak, B.; Jakubczyk, A. Evaluation of ACE, α-glucosidase, and lipase inhibitory activities of peptides obtained by in vitro digestion of selected species of edible insects. Eur. Food Res. Technol. 2020, 246, 1361–1369. [Google Scholar] [CrossRef]
- Li, Z.; Zhao, S.; Xin, X.; Zhang, B.; Thomas, A.; Charles, A.; Lee, K.S.; Jin, B.R.; Gui, Z. Purification and characterization of a novel immunomodulatory hexapeptide from alcalase hydrolysate of ultramicro-pretreated silkworm (Bombyx mori) pupa protein. J. Asia-Pac. Entomol. 2019, 22, 633–637. [Google Scholar] [CrossRef]
- Zielińska, E.; Baraniak, B.; Karaś, M. Identification of antioxidant and anti-inflammatory peptides obtained by simulated gastrointestinal digestion of three edible insects species (Gryllodes sigillatus, Tenebrio molitor, Schistocerca gragaria). Int. J. Food Sci. Technol. 2018, 53, 2542–2551. [Google Scholar] [CrossRef]
- Altomare, A.A.; Baron, G.; Aldini, G.; Carini, M.; D’Amato, A. Silkworm pupae as source of high-value edible proteins and of bioactive peptides. Food Sci. Nutr. 2020, 8, 2652–2661. [Google Scholar] [CrossRef]
- Ong, J.H.; Liang, C.E.; Wong, W.L.; Wong, F.C.; Chai, T.T. Multi-target anti-sars-cov-2 peptides from mealworm proteins: An in silico study. Malays. J. Biochem. Mol. Biol. 2021, 24, 83–91. [Google Scholar]
- Chen, F.; Jiang, H.; Lu, Y.; Chen, W.; Huang, G. Identification and in silico analysis of antithrombotic peptides from the enzymatic hydrolysates of Tenebrio molitor larvae. Eur. Food Res. Technol. 2019, 245, 2687–2695. [Google Scholar] [CrossRef]
- Meyer-Rochow, V.B.; Gahukar, R.T.; Ghosh, S.; Jung, C. Chemical Composition, Nutrient Quality and Acceptability of Edible Insects Are Affected by Species, Developmental Stage, Gender, Diet, and Processing Method. Foods 2021, 10, 1036. [Google Scholar] [CrossRef]
- Pali-Schöll, I.; Verhoeckx, K.; Mafra, I.; Bavaro, S.L.; Clare Mills, E.N.; Monaci, L. Allergenic and novel food proteins: State of the art and challenges in the allergenicity assessment. Trends Food Sci. Technol. 2019, 84, 45–48. [Google Scholar] [CrossRef]
- Tang, C.; Yang, D.; Liao, H.; Sun, H.; Liu, C.; Wei, L.; Li, F. Edible insects as a food source: A review. Food Prod. Process. Nutr. 2019, 1, 8. [Google Scholar] [CrossRef]
- European Union. Regulation (EU) 2015/2283. Regulation (EU) 2015/2283 of the European Parliament and of the Council of 25 November 2015 on novel foods, amending Regulation (EU) No 1169/2011 of the European Parliament and of the Council and repealing Regulation (EC) No 258/97 of the European Parliament and of the Council and Commission Regulation (EC) No 1852/2001. Off. J. Eur. Union 2015, L327, 1–22. [Google Scholar]
- European Union. Regulation (EU) 2021/882. Commission Implementing Regulation (EU) 2021/882 of 1 June 2021 authorising the placing on the market of dried Tenebrio molitor larva as a novel food under Regulation (EU) 2015/2283 of the European Parliament and of the Council, and amending Commission Implementing Regulation (EU) 2017/2470. Off. J. Eur. Union 2021, L194, 16–21. [Google Scholar]
- European Union. Regulation (EU) 2022/169. Commission Implementing Regulation (EU) 2022/169 of 8 February 2022 authorising the placing on the market of frozen, dried and powder forms of yellow mealworm (Tenebrio molitor larva) as a novel food under Regulation (EU) 2015/2283 of the European Parliament and of the Council, and amending Commission Implementing Regulation (EU) 2017/2470. Off. J. Eur. Union 2022, L28, 10–16. [Google Scholar]
- European Union. Regulation (EU) 2021/1975. Commission Implementing Regulation (EU) 2021/1975 of 12 November 2021 authorising the placing on the market of frozen, dried and powder forms of Locusta migratoria as a novel food under Regulation (EU) 2015/2283 of the European Parliament and of the Council and amending Commission Implementing Regulation (EU) 2017/2470. Off. J. Eur. Union 2021, L402, 10–16. [Google Scholar]
- European Union. Regulation (EU) 2022/188. Commission Implementing Regulation (EU) 2022/188 of 10 February 2022 authorising the placing on the market of frozen, dried and powder forms of Acheta domesticus as a novel food under Regulation (EU) 2015/2283 of the European Parliament and of the Council, and amending Commission Implementing Regulation (EU) 2017/2470. Off. J. Eur. Union 2022, L30, 108–114. [Google Scholar]
- European Union. Regulation (EU) 2023/58. Commission Implementing Regulation (EU) 2023/58 of 5 January 2023 authorising the placing on the market of the frozen, paste, dried and powder forms of Alphitobius diaperinus larvae (lesser mealworm) as a novel food and amending Implementing Regulation (EU) 2017/2470. Off. J. Eur. Union 2023, L5, 10–15. [Google Scholar]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. BMJ 2021, 372, n71. [Google Scholar] [CrossRef]
- Tejada, L.; Buendía-Moreno, L.; Hernández, I.; Abellán, A.; Cayuela, J.M.; Salazar, E.; Bueno-Gavilá, E. Bioactivities of Mealworm (Alphitobius diaperinus L.) Larvae Hydrolysates Obtained from Artichoke (Cynara scolymus L.) Proteases. Biology 2022, 11, 631. [Google Scholar] [CrossRef]
- Koh, J.A.; Ong, J.H.; Abd Manan, F.; Ee, K.Y.; Wong, F.C.; Chai, T.T. Discovery of Bifunctional Anti-DPP-IV and Anti-ACE Peptides from Housefly Larval Proteins After In silico Gastrointestinal Digestion. Biointerface Res. Appl. Chem. 2022, 12, 4929–4944. [Google Scholar] [CrossRef]
- de Matos, F.M.; de Lacerda, J.T.J.G.; Zanetti, G.; de Castro, R.J.S. Production of black cricket protein hydrolysates with α-amylase, α-glucosidase and angiotensin I-converting enzyme inhibitory activities using a mixture of proteases. Biocatal. Agric. Biotechnol. 2022, 39, 102276. [Google Scholar] [CrossRef]
- Cermeño, M.; Bascón, C.; Amigo-Benavent, M.; Felix, M.; FitzGerald, R.J. Identification of peptides from edible silkworm pupae (Bombyx mori) protein hydrolysates with antioxidant activity. J. Funct. Foods 2022, 92, 105052. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, J.; Zhu, Z.; Li, X.; Sun, S.; Wang, W.; Sadiq, F.A. Identification and characterization of two novel antioxidant peptides from silkworm pupae protein hydrolysates. Eur. Food Res. Technol. 2021, 247, 343–352. [Google Scholar] [CrossRef]
- Khammuang, S.; Sarnthima, R.; Sanachai, K. Purification and identification of novel antioxidant peptides from silkworm pupae (Bombyx mori) protein hydrolysate and molecular docking study. Biocatal. Agric. Biotechnol. 2022, 42, 102367. [Google Scholar] [CrossRef]
- Lu, J.; Guo, Y.; Muhmood, A.; Zeng, B.; Qiu, Y.; Wang, P.; Ren, L. Probing the antioxidant activity of functional proteins and bioactive peptides in Hermetia illucens larvae fed with food wastes. Sci. Rep. 2022, 12, 2799. [Google Scholar] [CrossRef]
- Sun, T.; Zhang, S.; Yang, W.; Zhao, Z.; Yang, D. Housefly pupae-derived antioxidant peptides exerting neuroprotective effects on hydrogen peroxide-induced oxidative damage in PC12 cells. Molecules 2019, 24, 4486. [Google Scholar] [CrossRef]
- Pattarayingsakul, W.; Nilavongse, A.; Reamtong, O.; Chittavanich, P.; Mungsantisuk, I.; Mathong, Y.; Prasitwuttisak, W.; Panbangred, W. Angiotensin-converting enzyme inhibitory and antioxidant peptides from digestion of larvae and pupae of Asian weaver ant, Oecophylla smaragdina, Fabricius. J. Sci. Food Agric. 2017, 97, 3133–3140. [Google Scholar] [CrossRef]
- Khajepour-Zaveh, A.; Asoodeh, A.; Naderi-Manesh, H. Antioxidant enzyme regulating and intracellular ROS scavenging capacities of two novel bioactive peptides from white grub larvae (Polyphylla adstpersa) hydrolysate in A549 cells. Med. Chem. Res. 2020, 29, 2039–2049. [Google Scholar] [CrossRef]
- Xu, X.; Gao, Y.X. Purification and identification of angiotensin I-converting enzyme-inhibitory peptides from apalbumin 2 during simulated gastrointestinal digestion. J. Sci. Food Agric. 2015, 95, 906–914. [Google Scholar] [CrossRef]
- Yang, X.; Chen, K.; Liu, H.; Zhang, Y.; Luo, Y. Purification and identification of peptides with high angiotensin-I converting enzyme (ACE) inhibitory activity from honeybee pupae (Apis mellifera) hydrolysates with in silico gastrointestinal digestion. Eur. Food Res. Technol. 2019, 245, 535–544. [Google Scholar] [CrossRef]
- Wu, Q.; Jia, J.; Yan, H.; Du, J.; Gui, Z. A novel angiotensin-I converting enzyme (ACE) inhibitory peptide from gastrointestinal protease hydrolysate of silkworm pupa (Bombyx mori) protein: Biochemical characterization and molecular docking study. Peptides 2015, 68, 17–24. [Google Scholar] [CrossRef]
- Liu, L.; Wei, Y.; Chang, Q.; Sun, H.; Chai, K.; Huang, Z.; Zhao, Z.; Zhao, Z. Ultrafast Screening of a Novel, Moderately Hydrophilic Angiotensin-Converting-Enzyme-Inhibitory Peptide, RYL, from Silkworm Pupa Using an Fe-Doped-Silkworm-Excrement-Derived Biocarbon: Waste Conversion by Waste. J. Agric. Food Chem. 2017, 65, 11202–11211. [Google Scholar] [CrossRef]
- Tao, M.; Sun, H.; Liu, L.; Luo, X.; Lin, G.; Li, R.; Zhao, Z.; Zhao, Z. Graphitized Porous Carbon for Rapid Screening of Angiotensin-Converting Enzyme Inhibitory Peptide GAMVVH from Silkworm Pupa Protein and Molecular Insight into Inhibition Mechanism. J. Agric. Food Chem. 2017, 65, 8626–8633. [Google Scholar] [CrossRef]
- Jia, J.; Wu, Q.; Yan, H.; Gui, Z. Purification and molecular docking study of a novel angiotensin-I converting enzyme (ACE) inhibitory peptide from alcalase hydrolysate of ultrasonic-pretreated silkworm pupa (Bombyx mori) protein. Process Biochem. 2015, 50, 876–883. [Google Scholar] [CrossRef]
- Wang, W.; Wang, N.; Zhou, Y.; Zhang, Y.; Xu, L.; Xu, J.; Feng, F.; He, G. Isolation of a novel peptide from silkworm pupae protein components and interaction characteristics to angiotensin I-converting enzyme. Eur. Food Res. Technol. 2010, 232, 29–38. [Google Scholar] [CrossRef]
- Tao, M.; Wang, C.; Liao, D.; Liu, H.; Zhao, Z.; Zhao, Z. Purification, modification and inhibition mechanism of angiotensin I-converting enzyme inhibitory peptide from silkworm pupa (Bombyx mori) protein hydrolysate. Process Biochem. 2017, 54, 172–179. [Google Scholar] [CrossRef]
- Wang, W.; Zhang, Y.; Wang, N.; Zhu, Z. Molecular mechanisms of several novel dipeptides with angiotensin-iconverting enzyme inhibitory activity from in-silico screening of silkworm pupae protein. Curr. Pharm. Biotechnol. 2014, 15, 691–699. [Google Scholar] [CrossRef]
- Vercruysse, L.; Smagghe, G.; van der Bent, A.; van Amerongen, A.; Ongenaert, M.; Van Camp, J. Critical evaluation of the use of bioinformatics as a theoretical tool to find high-potential sources of ACE inhibitory peptides. Peptides 2009, 30, 575–582. [Google Scholar] [CrossRef]
- Hall, F.; Reddivari, L.; Liceaga, A.M. Identification and Characterization of Edible Cricket Peptides on Hypertensive and Glycemic In Vitro Inhibition and Their Anti-Inflammatory Activity on RAW 264.7 Macrophage Cells. Nutrients 2020, 12, 3588. [Google Scholar] [CrossRef]
- Brai, A.; Immacolata Trivisani, C.; Vagaggini, C.; Stella, R.; Angeletti, R.; Iovenitti, G.; Francardi, V.; Dreassi, E. Proteins from Tenebrio molitor: An interesting functional ingredient and a source of ACE inhibitory peptides. Food Chem. 2022, 393, 133409. [Google Scholar] [CrossRef]
- Vercruysse, L.; Smagghe, G.; Matsui, T.; Van Camp, J. Purification and identification of an angiotensin I converting enzyme (ACE) inhibitory peptide from the gastrointestinal hydrolysate of the cotton leafworm, Spodoptera littoralis. Process Biochem. 2008, 43, 900–904. [Google Scholar] [CrossRef]
- Dai, C.; Ma, H.; Luo, L.; Yin, X. Angiotensin I-converting enzyme (ACE) inhibitory peptide derived from Tenebrio molitor (L.) larva protein hydrolysate. Eur. Food Res. Technol. 2013, 236, 681–689. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, N.; Wang, W.; Wang, J.; Zhu, Z.; Li, X. Molecular mechanisms of novel peptides from silkworm pupae that inhibit α-glucosidase. Peptides 2016, 76, 45–50. [Google Scholar] [CrossRef]
- Luo, F.; Fu, Y.; Ma, L.; Dai, H.; Wang, H.; Chen, H.; Zhu, H.; Yu, Y.; Hou, Y.; Zhang, Y. Exploration of Dipeptidyl Peptidase-IV (DPP-IV) Inhibitory Peptides from Silkworm Pupae (Bombyx mori) Proteins Based on in Silico and in Vitro Assessments. J. Agric. Food Chem. 2022, 70, 3862–3871. [Google Scholar] [CrossRef]
- Cho, H.R.; Lee, S.O. Novel hepatoprotective peptides derived from protein hydrolysates of mealworm (Tenebrio molitor). Food Res. Int. 2020, 133, 109194. [Google Scholar] [CrossRef]
- Lourenço, F.; Calado, R.; Medina, I.; Ameixa, O.M.C.C. The Potential Impacts by the Invasion of Insects Reared to Feed Livestock and Pet Animals in Europe and Other Regions: A Critical Review. Sustainability 2022, 14, 6361. [Google Scholar] [CrossRef]
- Mazurkiewicz, A.; Tumialis, D.; Pezowicz, E.; Urbanski, J.; Galewski, P.; Goral, K. The effect of density on the breeding optimization of the tropical house cricket Gryllodes sigillatus (Walker) (Orthoptera: Gryllidae). Ann. Wars. Univ. Life Sci.-SGGW. Anim. Sci. 2013, 52, 135–139. [Google Scholar]
- Zielińska, E.; Baraniak, B.; Karaś, M.; Rybczyńska, K.; Jakubczyk, A. Selected species of edible insects as a source of nutrient composition. Food Res. Int. 2015, 77, 460–466. [Google Scholar] [CrossRef]
- Soares Araújo, R.R.; dos Santos Benfica, T.A.R.; Ferraz, V.P.; Moreira Santos, E. Nutritional composition of insects Gryllus assimilis and Zophobas morio: Potential foods harvested in Brazil. J. Food Compos. Anal. 2019, 76, 22–26. [Google Scholar] [CrossRef]
- Turck, D.; Bohn, T.; Castenmiller, J.; De Henauw, S.; Hirsch-Ernst, K.I.; Maciuk, A.; Mangelsdorf, I.; McArdle, H.J.; Naska, A.; Pelaez, C.; et al. Safety of frozen and freeze-dried formulations of the lesser mealworm (Alphitobius diaperinus larva) as a Novel food pursuant to Regulation (EU) 2015/2283. EFSA J. 2022, 20, e07325. [Google Scholar] [CrossRef]
- Roncolini, A.; Milanović, V.; Aquilanti, L.; Cardinali, F.; Garofalo, C.; Sabbatini, R.; Clementi, F.; Belleggia, L.; Pasquini, M.; Mozzon, M.; et al. Lesser mealworm (Alphitobius diaperinus) powder as a novel baking ingredient for manufacturing high-protein, mineral-dense snacks. Food Res. Int. 2020, 131, 109031. [Google Scholar] [CrossRef]
- Alvandi, J.; Karimi, J.; Ghadamyari, M.; Sharifi, M.; Asoodeh, A. Physiological defense of the white grub, Polyphylla adspersa Motschulsky (Col., Scarabaeidae) against entomopathogenic nematodes. J. Asia-Pac. Entomol. 2017, 20, 878–885. [Google Scholar] [CrossRef]
- Ghosh, S.; Meyer-Rochow, V.B.; Jung, C. Honey bees and their brood: A potentially valuable resource of food, worthy of greater appreciation and scientific attention. J. Ecol. Environ. 2021, 45, 31. [Google Scholar] [CrossRef]
- Ghosh, S.; Jung, C.; Meyer-Rochow, V.B. Nutritional value and chemical composition of larvae, pupae, and adults of worker honey bee, Apis mellifera ligustica as a sustainable food source. J. Asia-Pac. Entomol. 2016, 19, 487–495. [Google Scholar] [CrossRef]
- Chakravorty, J.; Ghosh, S.; Megu, K.; Jung, C.; Meyer-Rochow, V.B. Nutritional and anti-nutritional composition of Oecophylla smaragdina (Hymenoptera: Formicidae) and Odontotermes sp. (Isoptera: Termitidae): Two preferred edible insects of Arunachal Pradesh, India. J. Asia-Pac. Entomol. 2016, 19, 711–720. [Google Scholar] [CrossRef]
- Pereira, R.F.P.; Silva, M.M.; de Zea Bermudez, V. Bombyx moriSilk Fibers: An Outstanding Family of Materials. Macromol. Mater. Eng. 2015, 300, 1171–1198. [Google Scholar] [CrossRef]
- Zhou, J.; Han, D. Safety evaluation of protein of silkworm (Antheraea pernyi) pupae. Food Chem. Toxicol. 2006, 44, 1123–1130. [Google Scholar] [CrossRef]
- Tassoni, L.; Cappellozza, S.; Dalle Zotte, A.; Belluco, S.; Antonelli, P.; Marzoli, F.; Saviane, A. Nutritional Composition of Bombyx mori Pupae: A Systematic Review. Insects 2022, 13, 644. [Google Scholar] [CrossRef]
- Sayed, W.; Ibrahim, N.; Hatab, M.; Zhu, F.; Rumpold, B. Comparative Study of the Use of Insect Meal from Spodoptera littoralis and Bactrocera zonata for Feeding Japanese Quail Chicks. Animals 2019, 9, 136. [Google Scholar] [CrossRef]
- Lupi, D.; Savoldelli, S.; Leonardi, M.G.; Jucker, C. Feeding in the adult of Hermetia illucens (Diptera Stratiomyidae): Reality or fiction? J. Entomol. Acarol. Res. 2019, 51, 27–32. [Google Scholar] [CrossRef]
- Surendra, K.C.; Olivier, R.; Tomberlin, J.K.; Jha, R.; Khanal, S.K. Bioconversion of organic wastes into biodiesel and animal feed via insect farming. Renew. Energy 2016, 98, 197–202. [Google Scholar] [CrossRef]
- Čičková, H.; Newton, G.L.; Lacy, R.C.; Kozánek, M. The use of fly larvae for organic waste treatment. Waste Manag. 2015, 35, 68–80. [Google Scholar] [CrossRef]
- Pieterse, E.; Pretorius, Q. Nutritional evaluation of dried larvae and pupae meal of the housefly (Musca domestica) using chemical- and broiler-based biological assays. Anim. Prod. Sci. 2014, 54, 347–355. [Google Scholar] [CrossRef]
- Lorenzo, J.M.; Munekata, P.E.S.; Gómez, B.; Barba, F.J.; Mora, L.; Pérez-Santaescolástica, C.; Toldrá, F. Bioactive peptides as natural antioxidants in food products–A review. Trends Food Sci. Technol. 2018, 79, 136–147. [Google Scholar] [CrossRef]
- Whelton, P.K.; Carey, R.M.; Aronow, W.S.; Casey, D.E.; Collins, K.J.; Dennison Himmelfarb, C.; DePalma, S.M.; Gidding, S.; Jamerson, K.A.; Jones, D.W.; et al. 2017 ACC/AHA/AAPA/ABC/ACPM/AGS/APhA/ASH/ASPC/NMA/PCNA Guideline for the Prevention, Detection, Evaluation, and Management of High Blood Pressure in Adults. J. Am. Coll. Cardiol. 2018, 71, e127–e248. [Google Scholar] [CrossRef]
- Ponikowski, P.; Voors, A.A.; Anker, S.D.; Bueno, H.; Cleland, J.G.F.; Coats, A.J.S.; Falk, V.; González-Juanatey, J.R.; Harjola, V.-P.; Jankowska, E.A.; et al. 2016 ESC Guidelines for the diagnosis and treatment of acute and chronic heart failure. Eur. Heart J. 2016, 37, 2129–2200. [Google Scholar] [CrossRef]
- Messerli, F.H.; Bangalore, S.; Bavishi, C.; Rimoldi, S.F. Angiotensin-Converting Enzyme Inhibitors in Hypertension. J. Am. Coll. Cardiol. 2018, 71, 1474–1482. [Google Scholar] [CrossRef]
- Iwaniak, A.; Minkiewicz, P.; Darewicz, M. Food-Originating ACE Inhibitors, Including Antihypertensive Peptides, as Preventive Food Components in Blood Pressure Reduction. Compr. Rev. Food Sci. Food Saf. 2014, 13, 114–134. [Google Scholar] [CrossRef]
- Nakamura, K.; Oe, H.; Kihara, H.; Shimada, K.; Fukuda, S.; Watanabe, K.; Takagi, T.; Yunoki, K.; Miyoshi, T.; Hirata, K.; et al. DPP-4 inhibitor and alpha-glucosidase inhibitor equally improve endothelial function in patients with type 2 diabetes: EDGE study. Cardiovasc. Diabetol. 2014, 13, 110. [Google Scholar] [CrossRef]
- Blanco, J.; Nguyen, C.; Callebaut, C.; Jacotot, E.; Krust, B.; Mazaleyrat, J.-P.; Wakselman, M.; Hovanessian, A.G. Dipeptidyl-peptidase IV-beta. Further characterization and comparison to dipeptidyl-peptidase IV activity of CD26. Eur. J. Biochem. 1998, 256, 369–378. [Google Scholar] [CrossRef]
- Seo, C.; Sakamoto, M.; Nishimura, R.; Tsujino, D.; Ando, K.; Morimoto, A.; Utsunomiya, K. Comparison of Glycemic Variability in Patients with Type 2 Diabetes Given Sitagliptin or Voglibose: A Continuous Glucose Monitoring–Based Pilot Study. Diabetes Technol. Ther. 2013, 15, 378–385. [Google Scholar] [CrossRef]
- Alqahtani, A.S.; Hidayathulla, S.; Rehman, M.T.; ElGamal, A.A.; Al-Massarani, S.; Razmovski-Naumovski, V.; Alqahtani, M.S.; El Dib, R.A.; AlAjmi, M.F. Alpha-Amylase and Alpha-Glucosidase Enzyme Inhibition and Antioxidant Potential of 3-Oxolupenal and Katononic Acid Isolated from Nuxia oppositifolia. Biomolecules 2019, 10, 61. [Google Scholar] [CrossRef]
- Jaismy Jacob, P.; Manju, S.L.; Ethiraj, K.R.; Elias, G. Safer anti-inflammatory therapy through dual COX-2/5-LOX inhibitors: A structure-based approach. Eur. J. Pharm. Sci. 2018, 121, 356–381. [Google Scholar] [CrossRef]
- Liu, T.-T.; Liu, X.-T.; Chen, Q.-X.; Shi, Y. Lipase Inhibitors for Obesity: A Review. Biomed. Pharmacother. 2020, 128, 110314. [Google Scholar] [CrossRef] [PubMed]
- Mackman, N. Triggers, targets and treatments for thrombosis. Nature 2008, 451, 914–918. [Google Scholar] [CrossRef] [PubMed]
- Harter, K.; Levine, M.; Henderson, S. Anticoagulation Drug Therapy: A Review. West. J. Emerg. Med. 2015, 16, 11–17. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Order | Family | Species | Number of Studies | Number of Peptides with Predicted Bioactivities | Number of Peptides with Bioactivity Assessed In Vitro | Number of Peptides with Bioactivity Assessed In Vivo |
|---|---|---|---|---|---|---|
| Orthoptera | Gryllidae | Gryllodes sigillatu | 3 | 7 | 4 | _ |
| Gryllus assimilis | 1 | 25 a | _ | _ | ||
| Acrididae | Schistocerca gregaria | 2 | 4 | 4 | _ | |
| Coleoptera | Tenebrionidae | Alphitobius diaperinus | 1 | 22 | _ | _ |
| Tenebrio molitor | 8 | 27 a | 10 | 1 | ||
| Scarabaeidae | Polyphylla adspersa | 1 | 2 | 2 | _ | |
| Hymenoptera | Apidae | Apis mellifera | 2 | 4 | 4 | _ |
| Formicidae | Oecophylla smaragdina | 1 | 3 | 3 | _ | |
| Lepidoptera | Bombycidae | Bombyx mori | 15 | 41 | 30 | _ |
| Noctuidae | Spodoptera littoralis | 2 | 2 | 2 | 2 | |
| Diptera | Stratiomyidae | Hermetia illucens | 2 | 33 | _ | _ |
| Muscidae | Musca domestica | 2 | 7 b | 2 | _ | |
| Total | 40 c | 177 | 61 | 3 |
| Peptide Sequence | Bioactive Property | Species | Bioactivity Assessed Individually In Vitro | Simulated GI Digestion | Reference | |
|---|---|---|---|---|---|---|
| #1 | APVAVAHAAVPA | antioxidant/antihypertensive | A. diaperinus | no/no | yes | [27] |
| #2 | ASVVEKLGDY | antioxidant/antihypertensive | A. diaperinus | no/no | yes | [27] |
| #3 | GLIGAPIAAPIAA | antioxidant/antihypertensive | A. diaperinus | no/no | yes | [27] |
| #4 | AF | antidiabetic/antihypertensive | M. domestica | no/no | yes | [28] |
| #5 | GW | antidiabetic/antihypertensive | M. domestica | no/no | yes | [28] |
| #6 | GY | antidiabetic/antihypertensive | M. domestica | no/no | yes | [28] |
| #7 | PH | antidiabetic/antihypertensive | M. domestica | no/no | yes | [28] |
| #8 | IIAPPER | antioxidant/anti-inflammatory | G. sigillatus | yes/yes | yes | [13] |
| antihypertensive/antidiabetic/antiobesity | G. sigillatus | yes/yes/yes | yes | [11] | ||
| #9 | LAPSTIK | antioxidant/anti-inflammatory | G. sigillatus | yes/yes | yes | [13] |
| antihypertensive/antidiabetic/antiobesity | G. sigillatus | yes/yes/yes | yes | [11] | ||
| #10 | VAPEEHPV | antioxidant/anti-inflammatory | G. sigillatus | yes/yes | yes | [13] |
| antihypertensive/antidiabetic/antiobesity | G. sigillatus | yes/yes/yes | yes | [11] | ||
| #11 | KVEGDLK | antioxidant/anti-inflammatory | G. sigillatus | yes/yes | yes | [13] |
| antihypertensive/antidiabetic/antiobesity | G. sigillatus | yes/yes/yes | yes | [11] | ||
| #12 | GKDAVIV | antioxidant/anti-inflammatory | S. gregaria | yes/yes | yes | [13] |
| antihypertensive/antidiabetic/antiobesity | S. gregaria | yes/yes/yes | yes | [11] | ||
| #13 | AIGVGAIER | antioxidant/anti-inflammatory | S. gregaria | yes/yes | yes | [13] |
| antihypertensive/antidiabetic/antiobesity | S. gregaria | yes/yes/yes | yes | [11] | ||
| #14 | FDPFPK | antioxidant/anti-inflammatory | S. gregaria | yes/yes | yes | [13] |
| antihypertensive/antidiabetic/antiobesity | S. gregaria | yes/yes/yes | yes | [11] | ||
| #15 | YETGNGIK | antioxidant/anti-inflammatory | S. gregaria | yes/yes | yes | [13] |
| antihypertensive/antidiabetic/antiobesity | S. gregaria | yes/yes/yes | yes | [11] | ||
| #16 | NYVADGLG | antioxidant/anti-inflammatory | T. molitor | yes/yes | yes | [13] |
| antihypertensive/antidiabetic/antiobesity | T. molitor | yes/yes/yes | yes | [11] | ||
| #17 | AAAPVAVAK | antioxidant/anti-inflammatory | T. molitor | yes/yes | yes | [13] |
| antihypertensive/antidiabetic/antiobesity | T. molitor | yes/yes/yes | yes | [11] | ||
| #18 | YDDGSYKPH | antioxidant/anti-inflammatory | T. molitor | yes/yes | yes | [13] |
| antihypertensive/antidiabetic/antiobesity | T. molitor | yes/yes/yes | yes | [11] | ||
| #19 | AGDDAPR | antioxidant/anti-inflammatory | T. molitor | yes/yes | yes | [13] |
| antihypertensive/antidiabetic/antiobesity | T. molitor | yes/yes/yes | yes | [11] | ||
| antihypertensive/antidiabetic | G. assimilis | no/no | no | [29] | ||
| #20 | GLIGAPIAAPI | antioxidant | A. diaperinus | no | yes | [27] |
| #21 | AYVGPDGVTY | antioxidant | A. diaperinus | no | yes | [27] |
| #22 | AESEVAALNR | antioxidant | A. diaperinus | no | yes | [27] |
| #23 | GLIGAPIAAPIA | antioxidant | A. diaperinus | no | yes | [27] |
| #24 | VDAAVLEKLE | antioxidant | A. diaperinus | no | yes | [27] |
| #25 | ASVVEKLGDYL | antioxidant | A. diaperinus | no | yes | [27] |
| #26 | VDAAVLEKLEA | antioxidant | A. diaperinus | no | yes | [27] |
| #27 | AGFAGDDAPRAVF | antioxidant | A. diaperinus | no | yes | [27] |
| #28 | GLIGAPIAAPIAAPL | antioxidant | A. diaperinus | no | yes | [27] |
| #29 | ASLEAEAKGKAEAL | antioxidant | A. diaperinus | no | yes | [27] |
| #30 | AIANAAEKKQKAF | antioxidant | A. diaperinus | no | yes | [27] |
| #31 | FSLPHAILRLDL | antioxidant | A. diaperinus | no | yes | [27] |
| #32 | YALPHAILRIDL | antioxidant | A. diaperinus | no | yes | [27] |
| #33 | VDAAVLEKLEAGF | antioxidant | A. diaperinus | no | yes | [27] |
| #34 | GLIGAPIAAPIAAPLA | antioxidant | A. diaperinus | no | yes | [27] |
| #35 | PADTPEVAAAKVAHA | antioxidant | A. diaperinus | no | yes | [27] |
| #36 | LKVDDLAAELDASQ | antioxidant | A. diaperinus | no | yes | [27] |
| #37 | VAYSPAAVVSH | antioxidant | A. diaperinus | no | yes | [27] |
| #38 | NDVLFF | antioxidant | B. mori | yes | no | [30] |
| #39 | SWFVTPF | antioxidant | B. mori | yes | no | [30] |
| #40 | FKGPACA | antioxidant | B. mori | yes | yes | [31] |
| #41 | SVLGTGC | antioxidant | B. mori | yes | yes | [31] |
| #42 | AAEYPA | antioxidant | B. mori | yes | no | [32] |
| #43 | AKPGVY | antioxidant | B. mori | yes | no | [32] |
| #44 | VEEPPKEE | antioxidant | H. illucens | no | no | [10] |
| #45 | EEKNPKATE | antioxidant | H. illucens | no | no | [10] |
| #46 | PTTAPSATIN | antioxidant | H. illucens | no | no | [10] |
| #47 | VEEPPKEEKNPK | antioxidant | H. illucens | no | no | [10] |
| #48 | MAAGTNLLDTK | antioxidant | H. illucens | no | no | [10] |
| #49 | ETKNDEASVEQIK | antioxidant | H. illucens | no | no | [10] |
| #50 | RPEELGPNK | antioxidant | H. illucens | no | no | [10] |
| #51 | FPGGETEALRR | antioxidant | H. illucens | no | no | [10] |
| #52 | AGGGGGGGGGGGKNL | antioxidant | H. illucens | no | no | [10] |
| #53 | IHKAGGGGGGGGGGGK | antioxidant | H. illucens | no | no | [10] |
| #54 | HPERPIPEH | antioxidant | H. illucens | no | no | [10] |
| #55 | DQAKAFLEKDNK | antioxidant | H. illucens | no | no | [10] |
| #56 | NWDLKEVGGGALP | antioxidant | H. illucens | no | no | [10] |
| #57 | SATTAIYMNALL | antioxidant | H. illucens | no | no | [10] |
| #58 | KDNEEAEAKPT | antioxidant | H. illucens | no | no | [10] |
| #59 | SLGGEMKQTAK | antioxidant | H. illucens | no | no | [10] |
| #60 | LTSGSANATGSR | antioxidant | H. illucens | no | no | [10] |
| #61 | GYGFGGGAGCLSMDTGAHLNR | antioxidant | H. illucens | no | no | [33] |
| #62 | AGLQFPVGR | antioxidant | H. illucens | no | no | [33] |
| #63 | HFQAPSHIR | antioxidant | H. illucens | no | no | [33] |
| #64 | VGIKAPGIIPR | antioxidant | H. illucens | no | no | [33] |
| #65 | GFIGPGVDVPAPDMGTGER | antioxidant | H. illucens | no | no | [33] |
| #66 | SQINFPIGGPTER | antioxidant | H. illucens | no | no | [33] |
| #67 | AVDSLVPIGR | antioxidant | H. illucens | no | no | [33] |
| #68 | VVPSANRAMVGIVAGGGRIDKPILK | antioxidant | H. illucens | no | no | [33] |
| #69 | GFKDQIQDVFK | antioxidant | H. illucens | no | no | [33] |
| #70 | TQLEPPISTPHCAR | antioxidant | H. illucens | no | no | [33] |
| #71 | TIRYPDPLIK | antioxidant | H. illucens | no | no | [33] |
| #72 | SKIPFNVTPGSEQIR | antioxidant | H. illucens | no | no | [33] |
| #73 | RIPFSHDDR | antioxidant | H. illucens | no | no | [33] |
| #74 | VLVDGPLTGVPR | antioxidant | H. illucens | no | no | [33] |
| #75 | GVEEDWLSAR | antioxidant | H. illucens | no | no | [33] |
| #76 | IGGIGTVPVGR | antioxidant | H. illucens | no | no | [33] |
| #77 | DFTPVCTTELGR | antioxidant | M. domestica | yes | yes | [34] |
| #78 | ARFEELCSDLFR | antioxidant | M. domestica | yes | yes | [34] |
| #79 | CTKKHKPNC | antioxidant | O. smaragdina | yes | yes | [35] |
| #80 | YPQSLRWRAK | antioxidant | P. adspersa | yes | no | [36] |
| #81 | LPLFFYDVRP | antioxidant | P. adspersa | yes | no | [36] |
| #82 | WDDMEK | antioxidant | G. assimilis | no | no | [29] |
| #83 | LEKDNALDRAAM | antihypertensive | A. diaperinus | no | yes | [27] |
| #84 | LLKPY | antihypertensive | A. mellifera | yes | yes | [37] |
| #85 | AVFPSIVGR | antihypertensive | A. mellifera | yes | yes | [38] |
| #86 | PGKVHIT | antihypertensive | A. mellifera | yes | yes | [35] |
| #87 | PPVLVFV | antihypertensive | A. mellifera | yes | yes | [35] |
| #88 | ASL | antihypertensive | B. mori | yes | yes | [39] |
| #89 | RYL | antihypertensive | B. mori | yes | yes | [40] |
| #90 | GAMVVH | antihypertensive | B. mori | yes | yes | [41] |
| #91 | KHV | antihypertensive | B. mori | yes | yes | [42] * |
| #92 | APPPKK | antihypertensive | B. mori | yes | no | [43] * |
| #93 | GNPWM | antihypertensive | B. mori | yes | yes | [44] * |
| #94 | IF | antihypertensive | B. mori | yes | yes | [45] |
| #95 | GD | antihypertensive | B. mori | yes | yes | [45] |
| #96 | DA | antihypertensive | B. mori | yes | yes | [45] |
| #97 | TE | antihypertensive | B. mori | yes | yes | [45] |
| #98 | TA | antihypertensive | B. mori | yes | yes | [45] |
| #99 | ES | antihypertensive | B. mori | yes | yes | [45] |
| #100 | SS | antihypertensive | B. mori | yes | yes | [45] |
| #101 | ST | antihypertensive | B. mori | yes | yes | [45] |
| #102 | SD | antihypertensive | B. mori | yes | yes | [45] |
| #103 | QD | antihypertensive | B. mori | yes | yes | [45] |
| #104 | QE | antihypertensive | B. mori | yes | yes | [45] |
| #105 | EG | antihypertensive | B. mori | no | yes | [46] |
| #106 | DL | antihypertensive | B. mori | no | yes | [46] |
| #107 | GM | antihypertensive | B. mori | no | yes | [46] |
| #108 | QK | antihypertensive | B. mori | no | yes | [46] |
| #109 | YKPRP | antihypertensive | G. sigillatus | no | yes | [47] |
| #110 | PHGAP | antihypertensive | G. sigillatus | no | yes | [47] |
| #111 | VGPPQ | antihypertensive | G. sigillatus | no | yes | [47] |
| #112 | AFLL | antihypertensive | G. assimilis | no | no | [29] |
| #113 | LPLP | antihypertensive | G. assimilis | no | no | [29] |
| #114 | DM(+15.99)EKIWH | antihypertensive | G. assimilis | no | no | [29] |
| #115 | VFPSIVGRPR | antihypertensive | G. assimilis | no | no | [29] |
| #116 | ASTSLEKSY | antihypertensive | G. assimilis | no | no | [29] |
| #117 | NILFSGTNVAAGKAR | antihypertensive | G. assimilis | no | no | [29] |
| #118 | NPEGLLTGRPR | antihypertensive | G. assimilis | no | no | [29] |
| #119 | RYDPNRVF | antihypertensive | G. assimilis | no | no | [29] |
| #120 | KPYDLGGRMF | antihypertensive | G. assimilis | no | no | [29] |
| #121 | YPLDL | antihypertensive | G. assimilis | no | no | [29] |
| #122 | WGPTKPP | antihypertensive | G. assimilis | no | no | [29] |
| #123 | FFGT | antihypertensive | O. smaragdina | yes | yes | [35] |
| #124 | LSRVP | antihypertensive | O. smaragdina | yes | yes | [35] |
| #125 | QGLGY | antihypertensive | T. molitor | yes | yes | [48] |
| #126 | NIKY | antihypertensive | T. molitor | yes | yes | [48] |
| #127 | HILG | antihypertensive | T. molitor | yes | yes | [48] |
| #128 | AVF | antihypertensive | S. littoralis | yes | yes | [49] |
| antihypertensive | S. littoralis | yes | yes | [8] | ||
| #129 | YAN | antihypertensive | T. molitor | yes | no | [50] |
| antihypertensive | T. molitor | yes | yes | [48] | ||
| #130 | VF | antidiabetic/antihypertensive | M. domestica | no | yes | [28] |
| antihypertensive | S. littoralis | yes | yes | [8] | ||
| #131 | QPGR | antidiabetic | B. mori | yes | yes | [51] * |
| #132 | SQSPA | antidiabetic | B. mori | yes | yes | [51] * |
| #133 | QPPT | antidiabetic | B. mori | yes | yes | [51] * |
| #134 | NSPR | antidiabetic | B. mori | yes | yes | [51] * |
| #135 | LPPEHDWR | antidiabetic | B. mori | yes | yes | [52] |
| #136 | LPAVTIR | antidiabetic | B. mori | yes | yes | [52] |
| #137 | APSTIKIKIIAPPER | antidiabetic | G. assimilis | no | no | [29] |
| #138 | EITALAPSTIKIK | antidiabetic | G. assimilis | no | no | [29] |
| #139 | Q(-17.03)RPEELPLLR | antidiabetic | G. assimilis | no | no | [29] |
| #140 | LAMVEA | antidiabetic | G. assimilis | no | no | [29] |
| #141 | LPPPP | antidiabetic | G. assimilis | no | no | [29] |
| #142 | ALLVVW | antidiabetic | G. assimilis | no | no | [29] |
| #143 | DSYPL | antidiabetic | G. assimilis | no | no | [29] |
| #144 | EKEEEFENTR | antidiabetic | G. assimilis | no | no | [29] |
| #145 | DGMEVPRTP | antidiabetic | G. assimilis | no | no | [29] |
| #146 | YPGDV | antidiabetic | G. assimilis | no | no | [29] |
| #147 | LPLPL | antidiabetic | G. assimilis | no | no | [29] |
| #148 | APVAH | antidiabetic | T. molitor | no | no | [9] |
| #149 | AVTTK | antidiabetic | T. molitor | no | no | [9] |
| #150 | AAGAPP | antidiabetic | T. molitor | no | no | [9] |
| #151 | SLAPK | antidiabetic | T. molitor | no | no | [9] |
| #152 | VHCSE | antidiabetic | T. molitor | no | no | [9] |
| #153 | PALLL | antidiabetic | T. molitor | no | no | [9] |
| #154 | PAALST | antidiabetic | T. molitor | no | no | [9] |
| #155 | AR | antidiabetic | T. molitor | no | no | [9] |
| #156 | CSR | antidiabetic | T. molitor | no | no | [9] |
| #157 | ATAL | antidiabetic | T. molitor | no | no | [9] |
| #158 | RVGS | antidiabetic | T. molitor | no | no | [9] |
| #159 | AGGP | antidiabetic | T. molitor | no | no | [9] |
| #160 | APYF | antidiabetic | T. molitor | no | no | [9] |
| #161 | DNKDCFL | antimicrobial | B. mori | no | yes | [14] |
| #162 | NNKMNCM | antimicrobial | B. mori | no | yes | [14] |
| #163 | TREQWF | antimicrobial | B. mori | no | yes | [14] |
| #164 | DNGSGMCK | antimicrobial | B. mori | no | yes | [14] |
| #165 | ESCMNCR | antimicrobial | B. mori | no | yes | [14] |
| #166 | NDNRINF | antimicrobial | B. mori | no | yes | [14] |
| #167 | KDCYTNM | antimicrobial | B. mori | no | yes | [14] |
| #168 | SLVDAIGMGP | antithrombotic | T. molitor | no | yes | [16] |
| #169 | AGFAGDDAPR | antithrombotic | T. molitor | no | yes | [16] |
| #170 | AKKHKE | antioxidant (hepatoprotective) | T. molitor | yes | no | [53] |
| #171 | LE | antioxidant (hepatoprotective) | T. molitor | yes | no | [53] |
| #172 | PKWF | anti-SARS-CoV-2 | T. molitor | no | no | [15] |
| #173 | VHRKCF | anti-SARS-CoV-2 | T. molitor | no | no | [15] |
| #174 | VGVL | hypocholesterolemic | G. assimilis | no | no | [29] |
| #175 | PNPNTN | immunomodulatory | B. mori | yes | yes | [12] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Teixeira, C.S.S.; Villa, C.; Costa, J.; Ferreira, I.M.P.L.V.O.; Mafra, I. Edible Insects as a Novel Source of Bioactive Peptides: A Systematic Review. Foods 2023, 12, 2026. https://doi.org/10.3390/foods12102026
Teixeira CSS, Villa C, Costa J, Ferreira IMPLVO, Mafra I. Edible Insects as a Novel Source of Bioactive Peptides: A Systematic Review. Foods. 2023; 12(10):2026. https://doi.org/10.3390/foods12102026
Chicago/Turabian StyleTeixeira, Carla S. S., Caterina Villa, Joana Costa, Isabel M. P. L. V. O. Ferreira, and Isabel Mafra. 2023. "Edible Insects as a Novel Source of Bioactive Peptides: A Systematic Review" Foods 12, no. 10: 2026. https://doi.org/10.3390/foods12102026
APA StyleTeixeira, C. S. S., Villa, C., Costa, J., Ferreira, I. M. P. L. V. O., & Mafra, I. (2023). Edible Insects as a Novel Source of Bioactive Peptides: A Systematic Review. Foods, 12(10), 2026. https://doi.org/10.3390/foods12102026